Exportin-1-Dependent Nuclear Export of DEAD-box Helicase DDX3X is Central to its Role in Antiviral Immunity

 , ,
, ,  and
and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Cell Lines
2.2. Molecular Cloning
2.3. hPIV-3 Infection and Plaque Assays
2.4. DNA Transfections
2.5. Cell Sorting
2.6. Co-Immunoprecipitation and Immunoblotting
2.7. NanoString RNA Profiling
2.8. Enzyme-Linked Immunosorbent Assay
2.9. Immunocytochemistry
2.10. Live and Indirect Immunofluorescence Microscopy
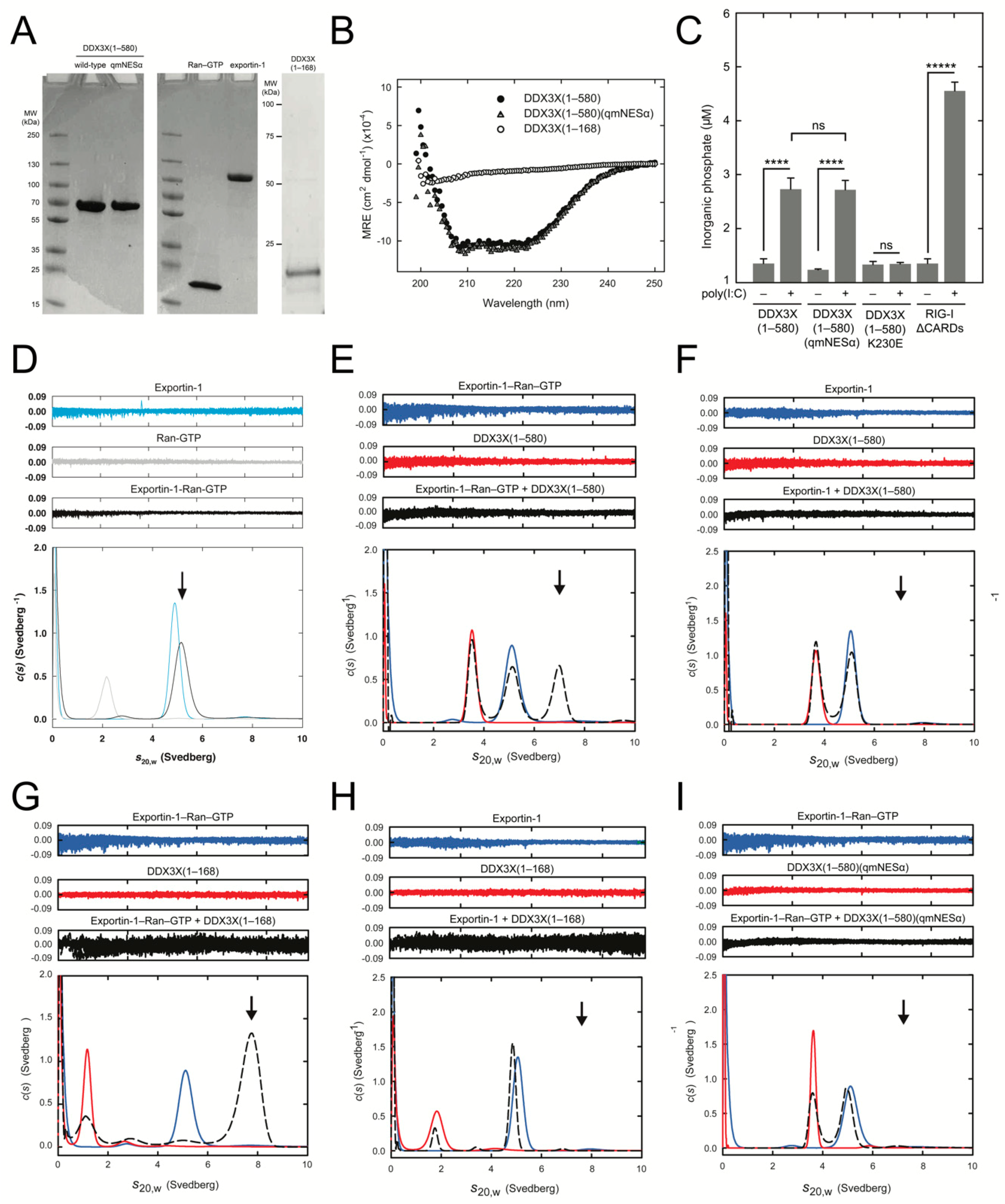
2.11. Expression and Purification of DDX3X
2.12. Expression and Assembly of Exportin-1-Ran-GTP
2.13. Circular Dichroism
2.14. RNA-Dependent ATP Hydrolysis Assays
2.15. Analytical Ultracentrifugation
2.16. Quantification and Statistical Analysis
3. Results
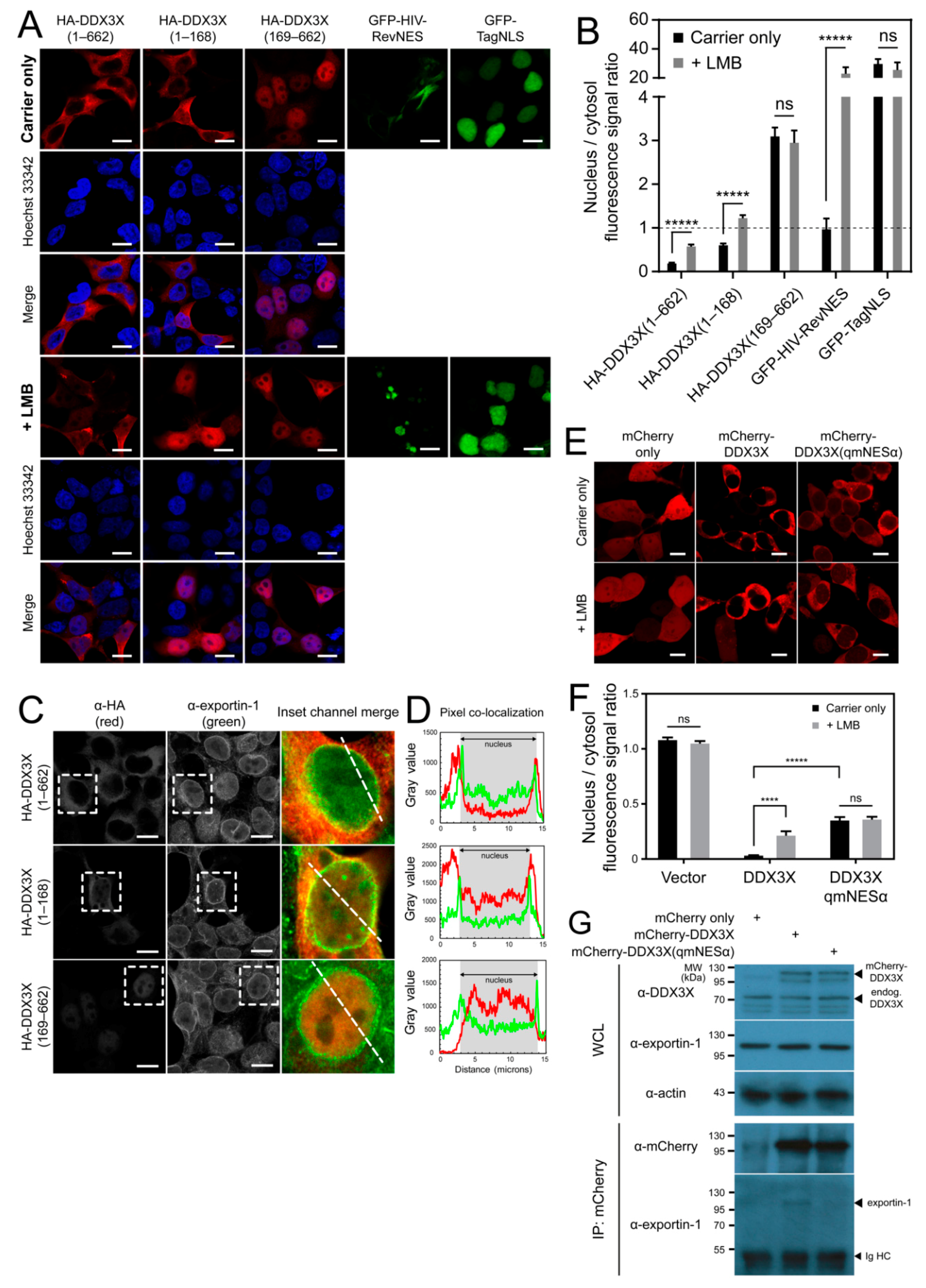
3.1. The DDX3X N-Terminus Mediates Its Nuclear Export
3.2. Exportin-1 Is a DDX3X Nuclear Export Receptor
3.3. DDX3X Harbors an Exportin-1 Recognized NES in the N-Terminus
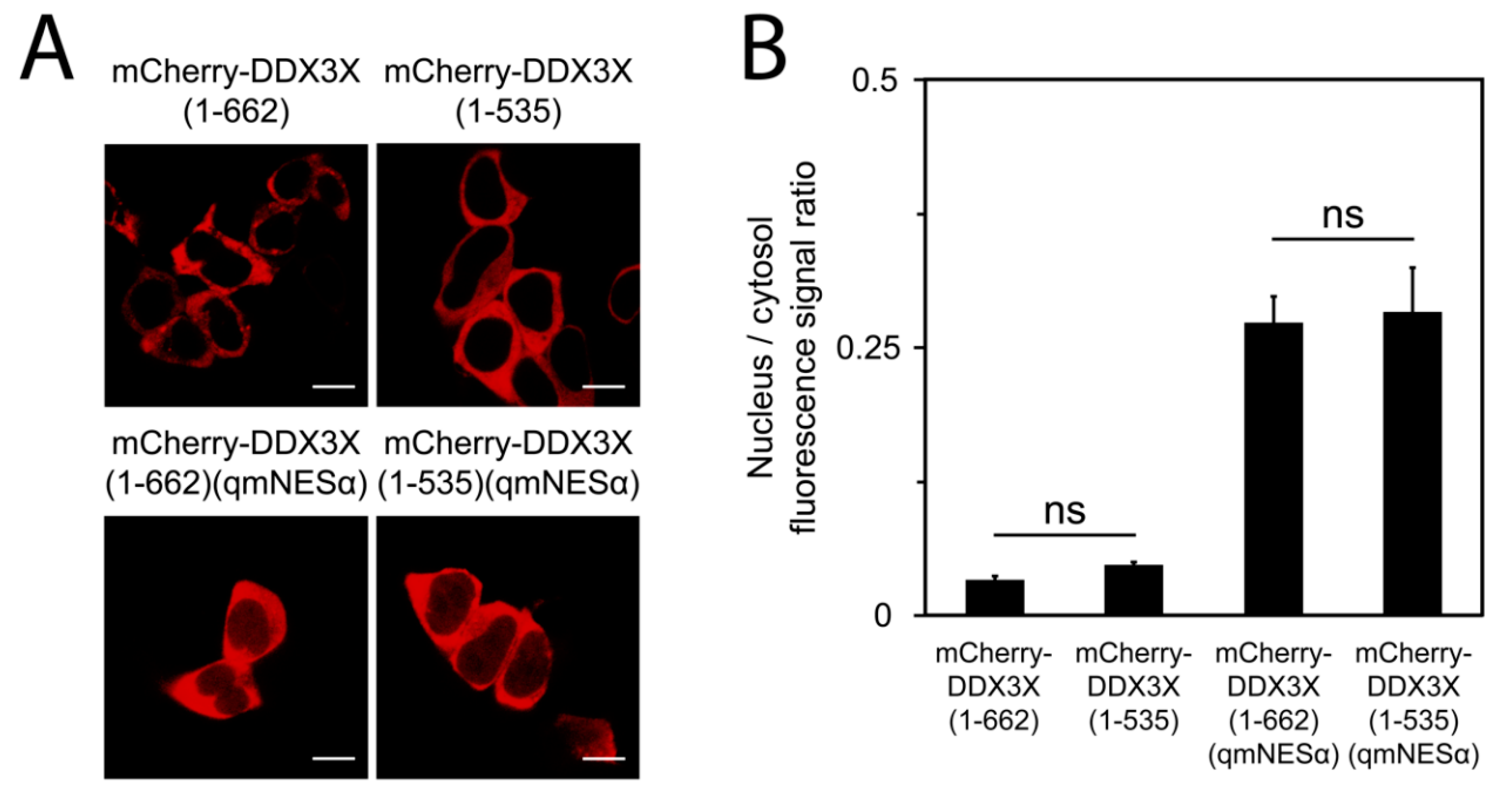
3.4. DDX3X’s C-Terminal Tail Is Dispensable for Nuclear Export
3.5. DDX3X-exportin-1 Binding Requires Ran-GTP
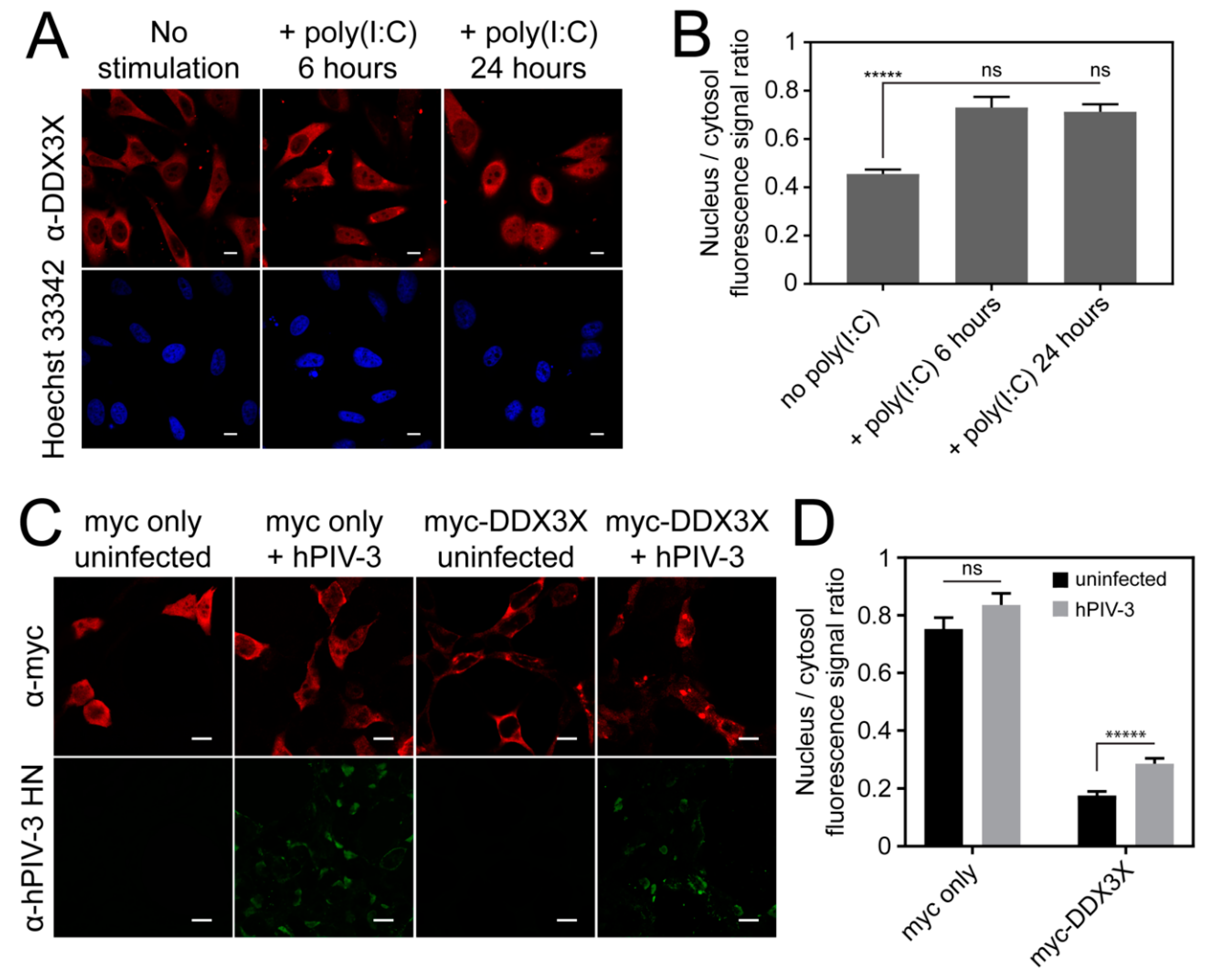
3.6. Invasive RNA Triggers DDX3X Nuclear Accumulation
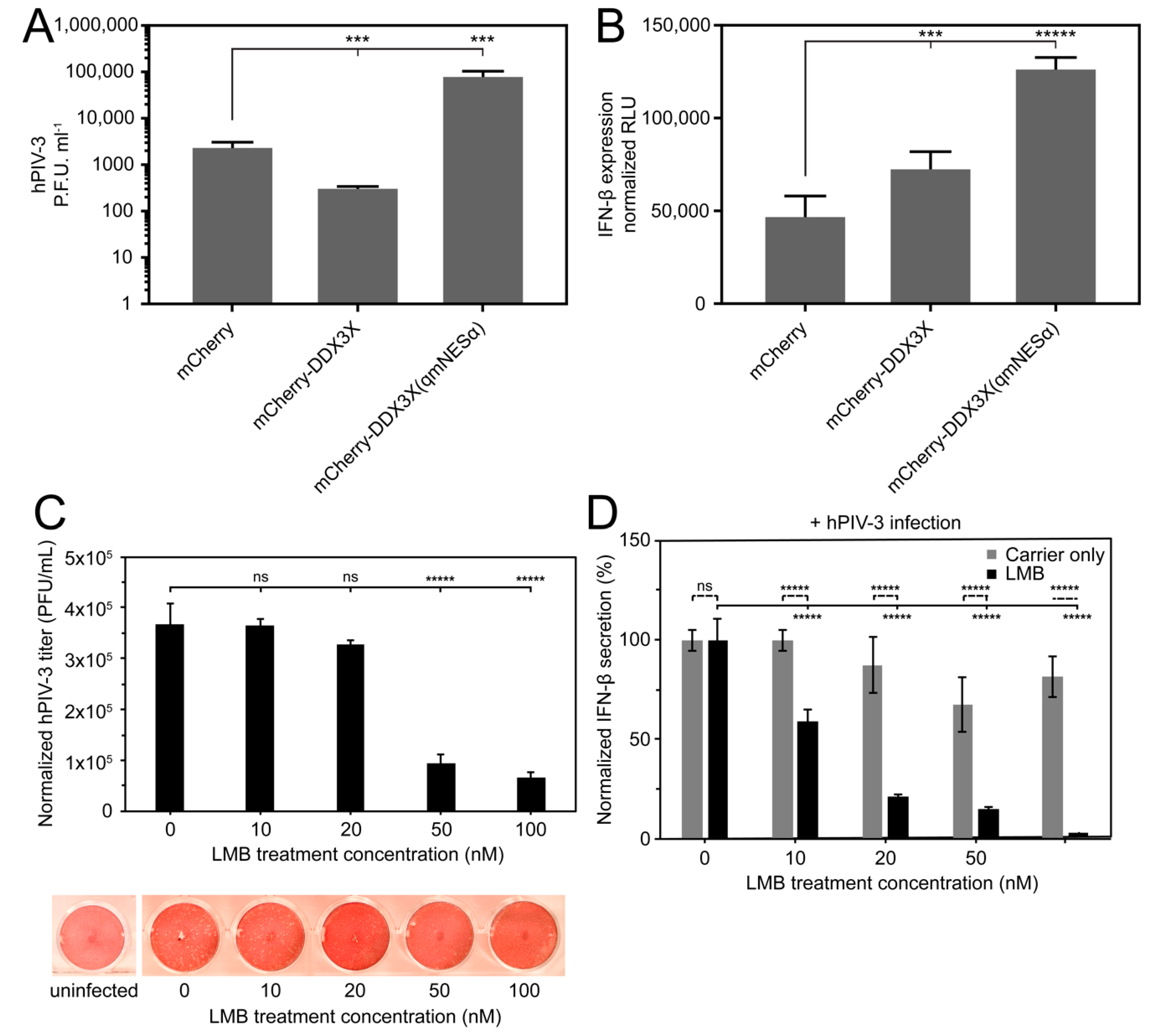
3.7. Overexpression of Wild-Type But Not Nuclear Export Defective DDX3X Can Protect Against hPIV-3 Infection
3.8. Exportin-1 Is Important to hPIV-3 Replication
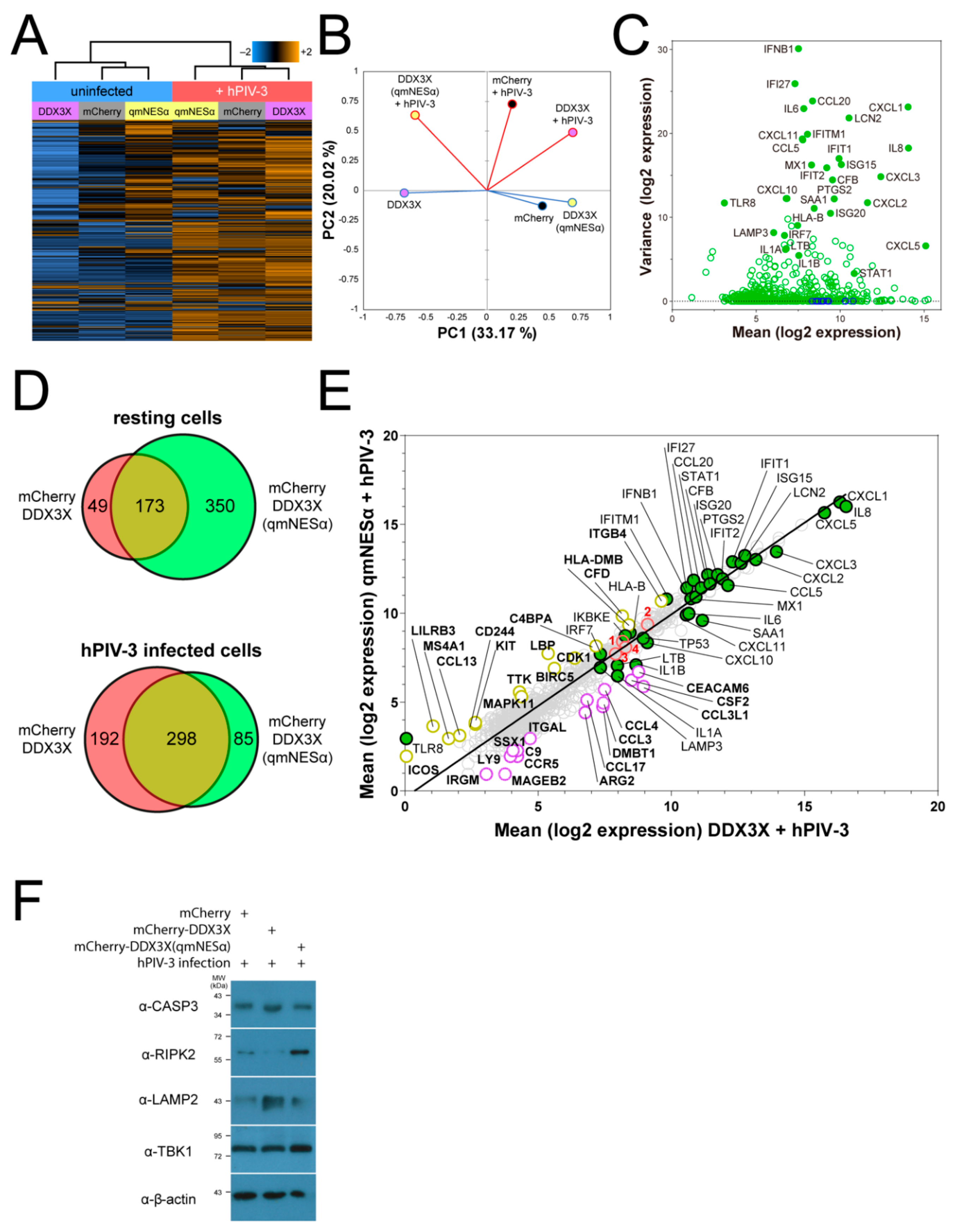
3.9. DDX3X’s Nuclear Trafficking Potentiates Immune Gene Induction
3.10. Nuclear DDX3X Contributes to IFNB1 Transcription and Influences ISG Subset Induction
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Valiente-Echeverria, F.; Hermoso, M.A.; Soto-Rifo, R.; Valiente-Echeverría, F.; Soto-Rifo, R. RNA helicase DDX3: At the crossroad of viral replication and antiviral immunity. Rev. Med. Virol. 2015, 25, 286–299. [Google Scholar] [CrossRef] [PubMed]
- Solis, M.; Nakhaei, P.; Jalalirad, M.; Lacoste, J.; Douville, R.; Arguello, M.; Zhao, T.; Laughrea, M.; Wainberg, M.A.; Hiscott, J. RIG-I-mediated antiviral signaling is inhibited in HIV-1 infection by a protease-mediated sequestration of RIG-I. J. Virol. 2011, 85, 1224–1236. [Google Scholar] [CrossRef] [PubMed]
- Sabbah, A.; Bose, S. Retinoic acid inducible gene I Activates innate antiviral response against human parainfluenza virus type 3. Virol. J. 2009, 6, 200. [Google Scholar] [CrossRef] [PubMed]
- Cortez, K.J.; Erdman, D.D.; Peret, T.C.T.; Gill, V.J.; Childs, R.; Barrett, A.J.; Bennett, J.E. Outbreak of human parainfluenza virus 3 infections in a hematopoietic stem cell transplant population. J. Infect. Dis. 2001, 184, 1093–1097. [Google Scholar] [CrossRef] [PubMed]
- Sydnor, E.R.; Greer, A.; Budd, A.P.; Pehar, M.; Munshaw, S.; Neofytos, D.; Perl, T.M.; Valsamakis, A. An outbreak of human parainfluenza virus 3 infection in an outpatient hematopoietic stem cell transplantation clinic. Am. J. Infect. Control. 2012, 40, 601–605. [Google Scholar] [CrossRef] [PubMed]
- Gu, L.; Fullam, A.; McCormack, N.; Hohn, Y.; Schroder, M. DDX3 directly regulates TRAF3 ubiquitination and acts as a scaffold to co-ordinate assembly of signalling complexes downstream from MAVS. Biochem. J 2017, 474, 571–587. [Google Scholar] [CrossRef] [PubMed]
- Oshiumi, H.; Sakai, K.; Matsumoto, M.; Seya, T. DEAD/H BOX 3 (DDX3) helicase binds the RIG-I adaptor IPS-1 to up-regulate IFN-beta-inducing potential. Eur. J. Immunol. 2010, 40, 940–948. [Google Scholar] [CrossRef]
- Schröder, M.; Baran, M.; Bowie, A.G. Viral targeting of DEAD box protein 3 reveals its role in TBK1/IKKepsilon-mediated IRF activation. EMBO J. 2008, 27, 2147–2157. [Google Scholar] [CrossRef] [PubMed]
- Heaton, S.M.; Borg, N.A.; Dixit, V.M. Ubiquitin in the activation and attenuation of innate antiviral immunity. J. Exp. Med. 2016, 213, 1–13. [Google Scholar] [CrossRef]
- Soulat, D.; Bürckstümmer, T.; Westermayer, S.; Goncalves, A.; Bauch, A.; Stefanovic, A.; Hantschel, O.; Bennett, K.L.; Decker, T.; Superti-Furga, G. The DEAD-box helicase DDX3X is a critical component of the TANK-binding kinase 1-dependent innate immune response. EMBO J. 2008, 27, 2135–2146. [Google Scholar] [CrossRef]
- Mohr, D.; Frey, S.; Fischer, T.; Güttler, T.; Görlich, D. Characterisation of the passive permeability barrier of nuclear pore complexes. EMBO J. 2009, 28, 2541–2553. [Google Scholar] [CrossRef] [PubMed]
- Fornerod, M.; Ohno, M.; Yoshida, M.; Mattaj, I.W. CRM1 is an export receptor for leucine-rich nuclear export signals. Cell 1997, 90, 1051–1060. [Google Scholar] [CrossRef]
- Fukuda, M.; Asano, S.; Nakamura, T.; Adachi, M.; Yoshida, M.; Yanagida, M.; Nishida, E. CRM1 is responsible for intracellular transport mediated by the nuclear export signal. Nature 1997, 390, 308–311. [Google Scholar] [CrossRef] [PubMed]
- Engelsma, D.; Bernad, R.; Calafat, J.; Fornerod, M. Supraphysiological nuclear export signals bind CRM1 independently of RanGTP and arrest at Nup358. EMBO J. 2004, 23, 3643–3652. [Google Scholar] [CrossRef] [PubMed]
- Sun, Q.; Carrasco, Y.P.; Hu, Y.; Guo, X.; Mirzaei, H.; Macmillan, J.; Chook, Y.M. Nuclear export inhibition through covalent conjugation and hydrolysis of Leptomycin B by CRM1. Proc. Natl. Acad. Sci. USA 2013, 110, 1303–1308. [Google Scholar] [CrossRef] [PubMed]
- Kudo, N.; Matsumori, N.; Taoka, H.; Fujiwara, D.; Schreiner, E.P.; Wolff, B.; Yoshida, M.; Horinouchi, S. Leptomycin B inactivates CRM1/exportin 1 by covalent modification at a cysteine residue in the central conserved region. Proc. Natl. Acad. Sci. USA 1999, 96, 9112–9117. [Google Scholar] [CrossRef] [PubMed]
- Yedavalli, V.S.; Neuveut, C.; Chi, Y.H.; Kleiman, L.; Jeang, K.T. Requirement of DDX3 DEAD Box RNA helicase for HIV-1 Rev-RRE export function. Cell 2004, 119, 381–392. [Google Scholar] [CrossRef] [PubMed]
- Lai, M.C.; Lee, Y.H.W.; Tarn, W.Y. The DEAD-box RNA helicase DDX3 associates with export messenger ribonucleoproteins as well as tip-associated protein and participates in translational control. Mol. Biol. Cell 2008, 19, 3847–3858. [Google Scholar] [CrossRef] [PubMed]
- Sekiguchi, T.; Iida, H.; Fukumura, J.; Nishimoto, T. Human DDX3Y, the Y-encoded isoform of RNA helicase DDX3, rescues a hamster temperature-sensitive ET24 mutant cell line with a DDX3X mutation. Exp. Cell Res. 2004, 300, 213–222. [Google Scholar] [CrossRef]
- Brennan, R.; Haap-Hoff, A.; Gu, L.; Gautier, V.; Long, A.; Schröder, M. Investigating nucleo-cytoplasmic shuttling of the human DEAD-box helicase DDX3. Eur. J. Cell Biol. 2018, 97, 501–511. [Google Scholar] [CrossRef]
- Sharma, D.; Bhattacharya, J. Evolutionary constraints acting on DDX3X protein potentially interferes with Rev-Mediated nuclear export of HIV-1 RNA. PLoS ONE 2010, 5, e9613. [Google Scholar] [CrossRef]
- Mahboobi, S.H.; Javanpour, A.A.; Mofrad, M.R.K. The interaction of RNA helicase DDX3 with HIV-1 Rev-CRM1-RanGTP complex during the HIV replication cycle. PLoS ONE 2015, 10, e0112969. [Google Scholar] [CrossRef]
- Atkinson, S.C.; Audsley, M.D.; Lieu, K.G.; Marsh, G.A.; Thomas, D.R.; Heaton, S.M.; Paxman, J.J.; Wagstaff, K.M.; Buckle, A.M.; Moseley, G.W.; et al. Recognition by host nuclear transport proteins drives disorder-to-order transition in Hendra virus V. Sci. Rep. 2018, 8, 358. [Google Scholar] [CrossRef] [PubMed]
- Rusinova, I.; Forster, S.; Yu, S.; Kannan, A.; Masse, M.; Cumming, H.; Chapman, R.; Hertzog, P.J. Interferome v2.0: An updated database of annotated interferon-regulated genes. Nucl. Acids Res. 2013, 41, D1040–D1046. [Google Scholar] [CrossRef] [PubMed]
- Fraser, J.E.; Rawlinson, S.M.; Heaton, S.M.; Jans, D.A. Dynamic nucleolar targeting of Dengue virus polymerase NS5 in response to extracellular pH. J. Virol. 2016, 90, 5797–5807. [Google Scholar] [CrossRef]
- Laue, T.M.; Shah, B.D.; Ridgeway, T.M.; Pelletier, S.L. Computer-aided interpretation of analytical sedimentation data for proteins. In Analytical Ultracentrifugation in Biochemistry and Polymer Science; Harding, S.E., Ed.; The Royal Society of Chemistry: Cambridge, UK, 1992; pp. 90–125. [Google Scholar]
- Schuck, P. Size-distribution analysis of macromolecules by sedimentation velocity ultracentrifugation and Lamm equation modeling. Biophys. J. 2000, 78, 1606–1619. [Google Scholar] [CrossRef]
- Moseley, G.W.; Roth, D.M.; DeJesus, M.A.; Leyton, D.L.; Filmer, R.P.; Pouton, C.W.; Jans, D.A. Dynein light chain association sequences can facilitate nuclear protein import. Mol. Biol. Cell 2007, 18, 3204–3213. [Google Scholar] [CrossRef]
- Li, J.-Z.; Chen, X.; Gong, X.-L.; Hu, H.-Y.; Shi, D.; Lu, Y.-M.; Qiu, L.; Lu, F.; Hu, Z.-L.; Zhang, J.-P. Identification of a functional nuclear localization signal mediating nuclear import of the zinc finger transcription factor ZNF24. PLoS ONE 2013, 8, e79910. [Google Scholar] [CrossRef]
- Van De Garde, M.D.B.; Pas, S.D.; Van Oord, G.W.; Gama, L.; Choi, Y.; De Man, R.A.; Boonstra, A.; Vanwolleghem, T. Interferon-alpha treatment rapidly clears Hepatitis E virus infection in humanized mice. Sci. Rep. 2017, 7, 8267. [Google Scholar] [CrossRef]
- Askjaer, P. The specificity of the CRM1-Rev nuclear export signal interaction is mediated by RanGTP. J. Biol. Chem. 1998, 273, 33414–33422. [Google Scholar] [CrossRef]
- Hübner, S.; Xiao, C.-Y.; Jans, D.A. The protein kinase CK2 site (Ser111/112) enhances recognition of the simian virus 40 large T-antigen nuclear localization sequence by importin. J. Biol. Chem. 1997, 272, 17191–17195. [Google Scholar] [CrossRef] [PubMed]
- Huang, T.T.; Kudo, N.; Yoshida, M.; Miyamoto, S. A nuclear export signal in the N-terminal regulatory domain of Ikappa Balpha controls cytoplasmic localization of inactive NF-kappa B/Ikappa Balpha complexes. Proc. Natl. Acad. Sci. USA 2000, 97, 1014–1019. [Google Scholar] [CrossRef] [PubMed]
- Senchi, K.; Matsunaga, S.; Hasegawa, H.; Kimura, H.; Ryo, A. Development of oligomannose-coated liposome-based nasal vaccine against human parainfluenza virus type 3. Front. Microbiol. 2013, 4, 346. [Google Scholar] [CrossRef] [PubMed]
- Ghildyal, R.; Ho, A.; Dias, M.; Soegiyono, L.; Bardin, P.G.; Tran, K.C.; Teng, M.N.; Jans, D.A. The respiratory syncytial virus matrix protein possesses a Crm1-mediated nuclear export mechanism. J. Virol. 2009, 83, 5353–5362. [Google Scholar] [CrossRef] [PubMed]
- Lundberg, L.; Pinkham, C.; Baer, A.; Amaya, M.; Narayanan, A.; Wagstaff, K.M.; Jans, D.A.; Kehn-Hall, K. Nuclear import and export inhibitors alter capsid protein distribution in mammalian cells and reduce Venezuelan Equine Encephalitis Virus replication. Antiviral. Res. 2013, 100, 662–672. [Google Scholar] [CrossRef] [PubMed]
- Yu, S.; Chen, J.; Wu, M.; Chen, H.; Kato, N.; Yuan, Z. Hepatitis B virus polymerase inhibits RIG-I- and Toll-like receptor 3-mediated beta interferon induction in human hepatocytes through interference with interferon regulatory factor 3 activation and dampening of the interaction between TBK1/IKKepsilon and DDX3. J. Gen. Virol. 2010, 91, 2080–2090. [Google Scholar] [CrossRef]
- Ariumi, Y.; Kuroki, M.; Abe, K.; Dansako, H.; Ikeda, M.; Wakita, T.; Kato, N. DDX3 DEAD-Box RNA helicase is required for hepatitis C virus RNA replication. J. Virol. 2007, 81, 13922–13926. [Google Scholar] [CrossRef] [PubMed]
- Raman, S.N.T.; Liu, G.; Pyo, H.M.; Cui, Y.C.; Xu, F.; Ayalew, L.E.; Tikoo, S.K.; Zhou, Y. DDX3 interacts with influenza A virus NS1 and NP proteins and exerts antiviral function through regulation of stress granule formation. J. Virol. 2016, 90, 3661–3675. [Google Scholar] [CrossRef]
- Li, C.; Ge, L.L.; Li, P.P.; Wang, Y.; Dai, J.J.; Sun, M.X.; Huang, L.; Shen, Z.Q.; Hu, X.C.; Ishag, H.; et al. Cellular DDX3 regulates Japanese encephalitis virus replication by interacting with viral un-translated regions. Virology 2014, 449, 70–81. [Google Scholar] [CrossRef]
- Chahar, H.S.; Chen, S.; Manjunath, N. P-body components LSM1, GW182, DDX3, DDX6 and XRN1 are recruited to WNV replication sites and positively regulate viral replication. Virology 2013, 436, 1–7. [Google Scholar] [CrossRef]
- Khadka, S.; Vangeloff, A.D.; Zhang, C.; Siddavatam, P.; Heaton, N.S.; Wang, L.; Sengupta, R.; Sahasrabudhe, S.; Randall, G.; Gribskov, M.; et al. A physical interaction network of dengue virus and human proteins. Mol. Cell. Proteom. 2011, 10. [Google Scholar] [CrossRef] [PubMed]
- Askjaer, P.; Bachi, A.; Wilm, M.; Bischoff, F.R.; Weeks, D.L.; Ogniewski, V.; Ohno, M.; Niehrs, C.; Kjems, J.; Mattaj, I.W.; et al. RanGTP-regulated interactions of CRM1 with nucleoporins and a shuttling DEAD-Box helicase. Mol. Cell. Biol. 1999, 19, 6276–6285. [Google Scholar] [CrossRef] [PubMed]
- Hauk, G.; Bowman, G.D. Formation of a trimeric Xpo1-Ran[GTP]-Ded1 exportin complex modulates ATPase and helicase activities of Ded1. PLoS ONE 2015, 10, e0131690. [Google Scholar] [CrossRef] [PubMed]
- Eberle, K.C.; McGill, J.L.; Reinhardt, T.A.; Sacco, R.E. Parainfluenza virus 3 blocks antiviral mediators downstream of the interferon lambda receptor by modulating Stat1 phosphorylation. J. Virol. 2015, 90, 2948–2958. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.; De, B.P.; Das, T.; Banerjee, A.K. Inhibition of human parainfluenza virus-3 replication by interferon and human MxA. Virology 1996, 220, 330–338. [Google Scholar] [CrossRef] [PubMed][Green Version]
- E Dorman, S.; Uzel, G.; Roesler, J.; Bradley, J.S.; Bastian, J.; Billman, G.; King, S.; Filie, A.; Schermerhorn, J.; Holland, S.M. Viral infections in interferon-gamma receptor deficiency. J. Pediatr. 1999, 135, 640–643. [Google Scholar] [CrossRef]
- Perwitasari, O.; Johnson, S.; Yan, X.; Register, E.; Crabtree, J.; Gabbard, J.; Howerth, E.; Shacham, S.; Carlson, R.; Tamir, S.; et al. Antiviral efficacy of verdinexor in vivo in two animal models of influenza A virus infection. PLoS ONE 2016, 11, e0167221. [Google Scholar] [CrossRef] [PubMed]
- Easton, A.; Gould, P.; Marsh, A. Use of DDX3X inhibitors for the treatment of pneumovirus infections. US Patent WO2015136292A1, 2012. [Google Scholar]
- Pickens, J.A.; Tripp, R.A. Verdinexor Targeting of CRM1 is a promising therapeutic approach against RSV and influenza viruses. Viruses 2018, 10, 48. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Individual Proteins | Mr1 | M2 | s20,w3 | f/f04 |
| Exportin-1 | 123386 | 117954 | 4.9 | 1.7 |
| Ran-GTP | 24423 | 25682 | 2.2 | 1.3 |
| DDX3X(1-168) | 19403 | 15856 | 1.7 | 1.7 |
| DDX3X(1-580) | 65945 | 71566 | 3.6 | 1.7 |
| DDX3X(1-580)(qmNESα) | 65743 | 70076 | 3.7 | 1.6 |
| Protein Complexes | Mr1 | M2 | s20,w3 | f/f04 |
| Exportin-1-Ran-GTP | 147809 | 133003 | 5.1 | 1.7 |
| DDX3X(1-168) + Exportin-1-Ran-GTP | 167212 | 128675 | 7.8 | 1.9 |
| DDX3X(1-580) + Exportin-1-Ran-GTP | 213754 | 179465 | 7.0 | 1.6 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Heaton, S.M.; Atkinson, S.C.; Sweeney, M.N.; Yang, S.N.Y.; Jans, D.A.; Borg, N.A. Exportin-1-Dependent Nuclear Export of DEAD-box Helicase DDX3X is Central to its Role in Antiviral Immunity. Cells 2019, 8, 1181. https://doi.org/10.3390/cells8101181
Heaton SM, Atkinson SC, Sweeney MN, Yang SNY, Jans DA, Borg NA. Exportin-1-Dependent Nuclear Export of DEAD-box Helicase DDX3X is Central to its Role in Antiviral Immunity. Cells. 2019; 8(10):1181. https://doi.org/10.3390/cells8101181
Chicago/Turabian StyleHeaton, Steven M., Sarah C. Atkinson, Melissa N. Sweeney, Sundy N. Y. Yang, David A. Jans, and Natalie A. Borg. 2019. "Exportin-1-Dependent Nuclear Export of DEAD-box Helicase DDX3X is Central to its Role in Antiviral Immunity" Cells 8, no. 10: 1181. https://doi.org/10.3390/cells8101181
APA StyleHeaton, S. M., Atkinson, S. C., Sweeney, M. N., Yang, S. N. Y., Jans, D. A., & Borg, N. A. (2019). Exportin-1-Dependent Nuclear Export of DEAD-box Helicase DDX3X is Central to its Role in Antiviral Immunity. Cells, 8(10), 1181. https://doi.org/10.3390/cells8101181