Siglecs in Brain Function and Neurological Disorders


Abstract
1. Introduction
The Siglec Family
2. Siglecs and Their Role in Neurological Disorders
2.1. General Biology of Siglecs That are Expressed in the Brain
2.2. Role of Siglecs in Brain Functions
2.3. Role of Siglecs in Neurological Disorders
3. Conclusions

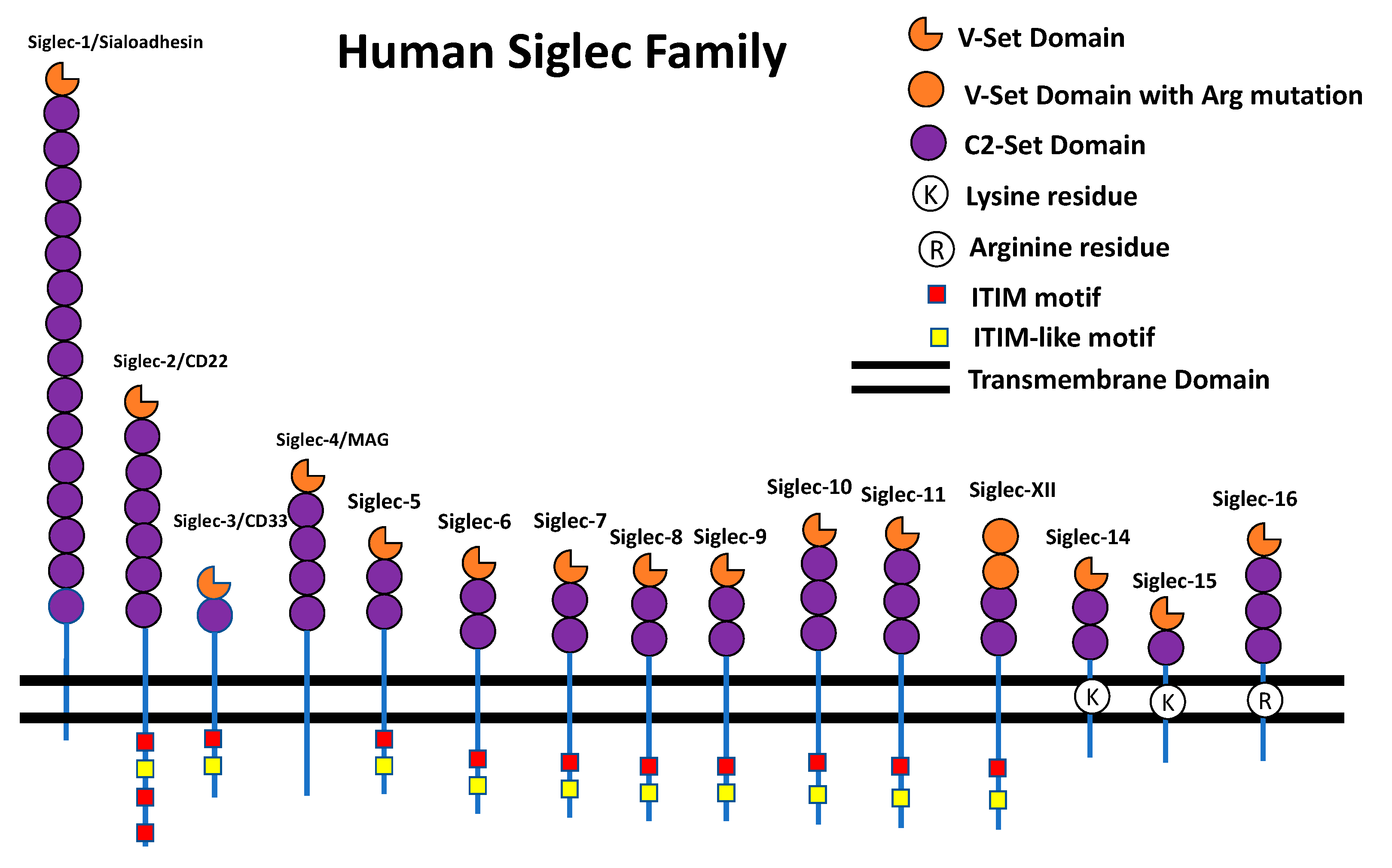{kind=link}
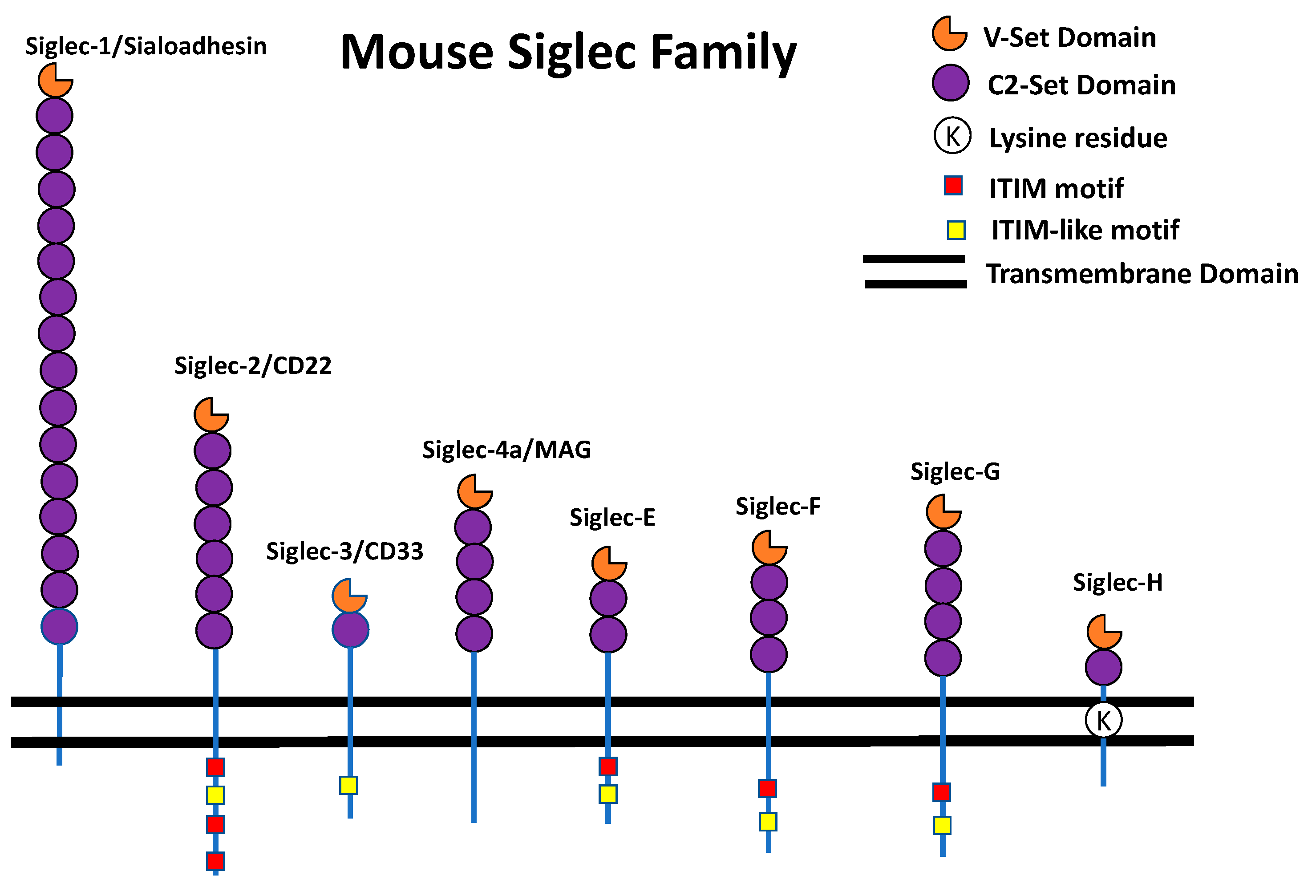{kind=link}
| Siglec Name | Alternate Name | Major Role | Major Role in Brain Disease/Disorder | Animal Model Exist | References |
|---|---|---|---|---|---|
| Siglec-1 | Sialoadhesin, CD169 | Clearance of Sialylated pathogens | Upregulation in ceroid lipofuscinosis (CLN) | Yes. Knockout mouse model exist | [34,82,104] |
| Siglec-2 | CD22, B-Lymphocyte Cell Adhesion Molecule (BL-CAM) | Regulation of B cell response upon inflammation | Microglial phagocytosis in aging brains | Yes. Knockout mouse model exist. | [35,68,105,106] |
| Siglec-3 | CD33, gp67, p67 | Inhibition of pathogenesis and clearance of amyloid plaques | Implicated in LOAD (Late Onset of Alzheimer’s Disease) | Yes. Knockout mouse model exist but mouse and human CD33 are very different (explained in the text) | [14,27,90,94] |
| Siglec-4 | Myelin associated glycoprotein (MAG) | Inhibition of neurite outgrowth and stimulation of axon growth cone collapse | Multiple sclerosis and benign monoclonal gammopathies | Yes. Knockout mouse model exist | [98,107,108,109] |
| Siglec-11 | NA | Reduction of inflammation, Impairment of phagocytosis and decrease in microglial neurotoxicity | Autism, Schizophrenia and bipolar disorder (indirect evidence) | Yes. Humanized knock in mouse model exist. | [4,78,79,101,102,110] |
| Siglec-E | NA | Neuroprotection, Inhibition of phagocytosis of neural debris and regulation of reactive oxygen species (ROS) production | NA | Yes. Knock out mouse model and knowk in E16 mouse model exist. | [22,23,53,60] |
| Siglec-F | NA | Protection of neuronal integrity | Neurodegenerative diseases | Yes. Knockout mouse model exist | [63,111,112] |
| Siglec-H | NA | Enhancement of phagocytosis | Amyotrophic lateral sclerosis and brain tumor | Yes. Knockout mouse model exist. | [28,81,113] |
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Freeze, H.H.; Eklund, E.A.; Ng, B.G.; Patterson, M.C. Neurological Aspects of Human Glycosylation Disorders. Annu. Rev. Neurosci. 2015, 38, 105–125. [Google Scholar] [CrossRef]
- Scott, H.; Panin, V.M. N-Glycosylation in Regulation of the Nervous System. In Glycobiology of the Nervous System; Yu, R.K., Schengrund, C.-L., Eds.; Springer: New York, NY, USA, 2009; Volume 9, pp. 367–394. [Google Scholar]
- Varki, A.; Schnaar, R.L.; Schauer, R. Sialic Acids and Other Nonulosonic Acids. In Essentials of Glycobiology, 3rd ed.; Varki, A., Cummings, R.D., Esko, J.D., Eds.; Cold Spring Harbor Laboratory Press: New York, NY, USA, 2017. [Google Scholar]
- Schnaar, R.L.; Gerardy-Schahn, R.; Hildebrandt, H. Sialic Acids in the Brain: Gangliosides and Polysialic Acid in Nervous System Development, Stability, Disease, and Regeneration. Physiol. Rev. 2014, 94, 461–518. [Google Scholar] [CrossRef] [PubMed]
- Pillai, S.; Netravali, I.A.; Cariappa, A.; Mattoo, H. Siglecs and immune regulation. Annu. Rev. Immunol. 2012, 30, 357–392. [Google Scholar] [CrossRef] [PubMed]
- Stamenkovic, I.; Seed, B. The B-cell antigen CD22 mediates monocyte and erythrocyte adhesion. Nature 1990, 345, 74–77. [Google Scholar] [CrossRef] [PubMed]
- Stamenkovic, I.; Sgroi, D.; Aruffo, A.; Sy, M.S.; Anderson, T. The B lymphocyte adhesion molecule CD22 interacts with leukocyte common antigen CD45RO on T cells and α2-6 sialyltransferase, CD75, on B cells. Cell 1991, 66, 1133–1144. [Google Scholar] [CrossRef]
- Crocker, P.; Kelm, S.; Dubois, C.; Martin, B.; McWilliam, A.; Shotton, D.; Paulson, J.; Gordon, S. Purification and properties of sialoadhesin, a sialic acid-binding receptor of murine tissue macrophages. EMBO J. 1991, 10, 1661–1669. [Google Scholar] [CrossRef]
- Crocker, P.; Mucklow, S.; Bouckson, V.; McWilliam, A.; Willis, A.; Gordon, S.; Milon, G.; Kelm, S.; Bradfield, P. Sialoadhesin, a macrophage sialic acid binding receptor for haemopoietic cells with 17 immunoglobulin-like domains. EMBO J. 1994, 13, 4490–4503. [Google Scholar] [CrossRef]
- Kelm, S.; Pelz, A.; Schauer, R.; Filbin, M.T.; Tang, S.; De Bellard, M.-E.; Schnaar, R.L.; Mahoney, J.A.; Hartnell, A.; Bradfield, P.; et al. Sialoadhesin, myelin-associated glycoprotein and CD22 define a new family of sialic acid-dependent adhesion molecules of the immunoglobulin superfamily. Curr. Boil. 1994, 4, 965–972. [Google Scholar] [CrossRef]
- Crocker, P.R.; Clark, E.A.; Filbin, M.; Gordon, S.; Jones, Y.; Kehrl, J.H.; Kelm, H.; Douarin, N.L.; Powell, L.; Roder, J.; et al. Siglecs: A family of sialic-acid binding lectins. Glycobiology 2012. [Google Scholar] [CrossRef]
- Varki, A.; Angata, T. Siglecs-The major subfamily of I-type lectins. Glycobiology 2006, 16, 1R–27R. [Google Scholar] [CrossRef]
- Crocker, P.R.; Varki, A. Siglecs, sialic acids and innate immunity. Trends Immunol. 2001, 22, 337–342. [Google Scholar] [CrossRef]
- Der Linden, E.C.M.B.-V.; Angata, T.; Reynolds, S.A.; Powell, L.D.; Hedrick, S.M.; Varki, A. CD33/Siglec-3 Binding Specificity, Expression Pattern, and Consequences of Gene Deletion in Mice. Mol. Cell. Boil. 2003, 23, 4199–4206. [Google Scholar] [CrossRef]
- Paulson, J.C.; Macauley, M.S.; Kawasaki, N. Siglecs as sensors of self in innate and adaptive immune responses. Ann. New York Acad. Sci. 2012, 1253, 37–48. [Google Scholar] [CrossRef]
- Angata, T.; Hayakawa, T.; Yamanaka, M.; Varki, A.; Nakamura, M. Discovery of Siglec-14, a novel sialic acid receptor undergoing concerted evolution with Siglec-5 in primates. FASEB J. 2006, 20, 1964–1973. [Google Scholar] [CrossRef]
- Cao, H.; Lakner, U.; De Bono, B.; Traherne, J.A.; Trowsdale, J.; Barrow, A.D. SIGLEC16encodes a DAP12-associated receptor expressed in macrophages that evolved from its inhibitory counterpartSIGLEC11and has functional and non-functional alleles in humans. Eur. J. Immunol. 2008, 38, 2303–2315. [Google Scholar] [CrossRef]
- Kameda, Y.; Takahata, M.; Komatsu, M.; Mikuni, S.; Hatakeyama, S.; Shimizu, T.; Angata, T.; Kinjo, M.; Minami, A.; Iwasaki, N. Siglec-15 Regulates Osteoclast Differentiation by Modulating RANKL-Induced Phosphatidylinositol 3-Kinase/Akt and Erk Pathways in Association With Signaling Adaptor DAP12. J. Bone Miner. Res. 2013, 28, 2463–2475. [Google Scholar] [CrossRef]
- Takamiya, R.; Ohtsubo, K.; Takamatsu, S.; Taniguchi, N.; Angata, T. The interaction between Siglec-15 and tumor-associated sialyl-Tn antigen enhances TGF-β secretion from monocytes/macrophages through the DAP12-Syk pathway. Glycobiology 2013, 23, 178–187. [Google Scholar] [CrossRef]
- Ishida-Kitagawa, N.; Tanaka, K.; Bao, X.; Kimura, T.; Miura, T.; Kitaoka, Y.; Hayashi, K.; Sato, M.; Maruoka, M.; Ogawa, T.; et al. Siglec-15 Protein Regulates Formation of Functional Osteoclasts in Concert with DNAX-activating Protein of 12 kDa (DAP12). J. Boil. Chem. 2012, 287, 17493–17502. [Google Scholar] [CrossRef]
- Ali, S.R.; Fong, J.J.; Carlin, A.F.; Busch, T.D.; Linden, R.; Angata, T.; Areschoug, T.; Parast, M.; Varki, N.; Murray, J.; et al. Siglec-5 and Siglec-14 are polymorphic paired receptors that modulate neutrophil and amnion signaling responses to group B Streptococcus. J. Exp. Med. 2014, 211, 1231–1242. [Google Scholar] [CrossRef]
- Schwarz, F.; Landig, C.S.; Siddiqui, S.; Secundino, I.; Olson, J.; Varki, N.; Nizet, V.; Varki, A. Paired Siglec receptors generate opposite inflammatory responses to a human-specific pathogen. EMBO J. 2017, 36, 751–760. [Google Scholar] [CrossRef]
- Stanczak, M.A.; Siddiqui, S.S.; Trefny, M.P.; Thommen, D.S.; Boligan, K.F.; Von Gunten, S.; Tzankov, A.; Tietze, L.; Lardinois, D.; Heinzelmann-Schwarz, V.; et al. Self-associated molecular patterns mediate cancer immune evasion by engaging Siglecs on T cells. J. Clin. Investig. 2018, 128, 4912–4923. [Google Scholar] [CrossRef]
- Brinkman-Van Der linden, E.C.M.; Hurtado-Ziola, N.; Hayakawa, T.; Wiggleton, L.; Benirschke, K.; Varki, A.; Varki, N. Human-specific expression of Siglec-6 in the placenta. Glycobiology 2007, 17, 922–931. [Google Scholar] [CrossRef]
- Mitra, N.; Banda, K.; Altheide, T.K.; Schaffer, L.; Johnson-Pais, T.L.; Beuten, J.; Leach, R.J.; Angata, T.; Varki, N.; Varki, A. SIGLEC12, a Human-specific Segregating (Pseudo)gene, Encodes a Signaling Molecule Expressed in Prostate Carcinomas. J. Boil. Chem. 2011, 286, 23003–23011. [Google Scholar] [CrossRef]
- Dharmadhikari, G.; Stolz, K.; Hauke, M.; Morgan, N.G.; Varki, A.; De Koning, E.; Kelm, S.; Maedler, K. Siglec-7 restores β-cell function and survival and reduces inflammation in pancreatic islets from patients with diabetes. Sci. Rep. 2017, 7. [Google Scholar] [CrossRef]
- Estus, S.; Shaw, B.C.; Devanney, N.; Katsumata, Y.; Press, E.E.; Fardo, D.W. Evaluation of CD33 as a genetic risk factor for Alzheimer’s disease. Acta Neuropathol. 2019, 138, 187–199. [Google Scholar] [CrossRef]
- Siew, J.J.; Chern, Y. Microglial Lectins in Health and Neurological Diseases. Front. Mol. Neurosci. 2018, 11. [Google Scholar] [CrossRef]
- Linnartz-Gerlach, B.; Mathews, M.; Neumann, H. Sensing the neuronal glycocalyx by glial sialic acid binding immunoglobulin-like lectins. Neuroscience 2014, 275, 113–124. [Google Scholar] [CrossRef]
- Linnartz-Gerlach, B.; Kopatz, J.; Neumann, H. Siglec functions of microglia. Glycobiology 2014, 24, 794–799. [Google Scholar] [CrossRef]
- Landig, C.S.; Hazel, A.; Kellman, B.P.; Fong, J.J.; Schwarz, F.; Agarwal, S.; Varki, N.; Massari, P.; Lewis, N.E.; Ram, S.; et al. Evolution of the exclusively human pathogen Neisseria gonorrhoeae: Human-specific engagement of immunoregulatory Siglecs. Evol. Appl. 2019, 12, 337–349. [Google Scholar] [CrossRef]
- Tecle, E.; Reynoso, H.S.; Wang, R.; Gagneux, P. The female reproductive tract contains multiple innate sialic acid-binding immunoglobulin-like lectins (Siglecs) that facilitate sperm survival. J. Boil. Chem. 2019, 294, 11910–11919. [Google Scholar] [CrossRef]
- Hartnell, A.; Steel, J.; Turley, H.; Jones, M.; Jackson, D.G.; Crocker, P.R. Characterization of human sialoadhesin, a sialic acid binding receptor expressed by resident and inflammatory macrophage populations. Blood 2001, 97, 288–296. [Google Scholar] [CrossRef]
- Crocker, P.R.; Hartnell, A.; Munday, J.; Nath, D. The potential role of sialoadhesin as a macrophage recognition molecule in health and disease. Glycoconj. J. 1997, 14, 601–609. [Google Scholar] [CrossRef]
- Jellusova, J.; Nitschke, L. Regulation of B Cell Functions by the Sialic Acid-Binding Receptors Siglec-G and CD22. Front. Immunol. 2012, 2. [Google Scholar] [CrossRef]
- Andrews, R.G.; Torok-Storb, B.; Bernstein, I.D. Myeloid-associated differentiation antigens on stem cells and their progeny identified by monoclonal antibodies. Blood 1983, 62, 124–132. [Google Scholar]
- Freeman, S.D.; Kelm, S.; Barber, E.K.; Crocker, P.R. Characterization of CD33 as a new member of the sialoadhesin family of cellular interaction molecules. Blood 1995, 85, 2005–2012. [Google Scholar]
- Martínez-Esparza, M.; Miguel, R.C.-S.; Hernández-Caselles, T.; Pérez-Oliva, A.B.; Vicente-Fernández, J.J.; García-Peñarrubia, P. Epitope mapping, expression and post-translational modifications of two isoforms of CD33 (CD33M and CD33m) on lymphoid and myeloid human cells. Glycobiol. 2011, 21, 757–770. [Google Scholar]
- Hernández-Caselles, T.; Martínez-Esparza, M.; Pérez-Oliva, A.B.; Quintanilla-Cecconi, A.M.; García-Alonso, A.; Alvarez-López, D.M.R.; García-Peñarrubia, P. A study of CD33 (SIGLEC-3) antigen expression and function on activated human T and NK cells: Two isoforms of CD33 are generated by alternative splicing. J. Leukoc. Boil. 2006, 79, 46–58. [Google Scholar] [CrossRef]
- Quarles, R.H. Myelin sheaths: Glycoproteins involved in their formation, maintenance and degeneration. Cell. Mol. Life Sci. 2002, 59, 1851–1871. [Google Scholar] [CrossRef]
- Quarles, R.H.; Everly, J.L.; Brady, R.O. Evidence for the close association of a glycoprotein with myelin in rat brain. J. Neurochem. 1973, 21, 1177–1191. [Google Scholar] [CrossRef]
- Arquint, M.; Roder, J.; Chia, L.S.; Down, J.; Wilkinson, D.; Bayley, H.; Braun, P.; Dunn, R. Molecular cloning and primary structure of myelin-associated glycoprotein. Proc. Natl. Acad. Sci. USA 1987, 84, 600–604. [Google Scholar] [CrossRef]
- Sutcliffe, J.; Milner, R.J.; Shinnick, T.M.; Bloom, F.E. Identifying the protein products of brain-specific genes with antibodies to chemically synthesized peptides. Cell 1983, 33, 671–682. [Google Scholar] [CrossRef]
- Vinson, M.; Strijbos, P.J.L.M.; Rowles, A.; Facci, L.; Moore, S.E.; Simmons, D.L.; Walsh, F.S. Myelin-associated glycoprotein interacts with ganglioside GT1b. A mechanism for neurite outgrowth inhibition. J. Boil. Chem. 2001, 276, 20280–20285. [Google Scholar] [CrossRef]
- Collins, B.E.; Yang, L.J.-S.; Mukhopadhyay, G.; Filbin, M.T.; Kiso, M.; Hasegawa, A.; Schnaar, R.L. Sialic Acid Specificity of Myelin-associated Glycoprotein Binding. J. Boil. Chem. 1997, 272, 1248–1255. [Google Scholar] [CrossRef]
- Trapp, B.D. Myelin-Associated Glycoprotein Location and Potential Functions. Ann. N. Y. Acad. Sci. 1990, 605, 29–43. [Google Scholar] [CrossRef]
- Angata, T.; Kerr, S.C.; Greaves, D.R.; Varki, N.M.; Crocker, P.R.; Varki, A. Cloning and characterization of human Siglec-11: A recently evolved signaling molecule that can interact with SHP-1 and SHP-2 and is expressed by tissue macrophages, including brain microglia. J. Biol. Chem. 2002, 27, 24466–24474. [Google Scholar] [CrossRef]
- Wang, X.; Mitra, N.; Cruz, P.; Deng, L.; Varki, N.; Angata, T.; Green, E.D.; Mullikin, J.; Hayakawa, T.; Varki, A. Evolution of Siglec-11 and Siglec-16 Genes in Hominins. Mol. Boil. Evol. 2012, 29, 2073–2086. [Google Scholar] [CrossRef]
- Hayakawa, T.; Angata, T.; Lewis, A.L.; Mikkelsen, T.S.; Varki, N.M.; Varki, A. Evolution: A human-specific gene in microglia. Science 2005, 309. [Google Scholar] [CrossRef]
- YU, Z.; MAOUI, M.; WU, L.; BANVILLE, D.; SHEN, S.-H. mSiglec-E, a novel mouse CD33-related siglec (sialic acid-binding immunoglobulin-like lectin) that recruits Src homology 2 (SH2)-domain-containing protein tyrosine phosphatases SHP-1 and SHP-2. Biochem. J. 2015, 353, 483–492. [Google Scholar] [CrossRef]
- Siddiqui, S.; Schwarz, F.; Springer, S.; Khedri, Z.; Yu, H.; Deng, L.; Verhagen, A.; Naito-Matsui, Y.; Jiang, W.; Kim, D.; et al. Studies on the detection, expression, glycosylation, dimerization, and ligand binding properties of mouse Siglec-E. J. Biol. Chem. 2017, 292, 1029–1037. [Google Scholar] [CrossRef]
- Zhang, J.Q.; Biedermann, B.; Nitschke, L.; Crocker, P.R. The murine inhibitory receptor mSiglec-E is expressed broadly on cells of the innate immune system whereas mSiglec-F is restricted to eosinophils. Eur. J. Immunol. 2004, 34, 1175–1184. [Google Scholar] [CrossRef]
- Uchiyama, S.; Sun, J.; Fukahori, K.; Ando, N.; Wu, M.; Schwarz, F.; Siddiqui, S.S.; Varki, A.; Marth, J.D.; Nizet, V. Dual actions of group B Streptococcus capsular sialic acid provide resistance to platelet-mediated antimicrobial killing. Proc. Natl. Acad. Sci. USA 2019, 116, 7465–7470. [Google Scholar] [CrossRef]
- Spence, S.; Greene, M.K.; Fay, F.; Hams, E.; Saunders, S.P.; Hamid, U.; Fitzgerald, M.; Beck, J.; Bains, B.K.; Smyth, P.; et al. Targeting Siglecs with a sialic acid-decorated nanoparticle abrogates inflammation. Sci. Transl. Med. 2015, 7, ra140. [Google Scholar] [CrossRef]
- Läubli, H.; Pearce, O.M.T.; Schwarz, F.; Siddiqui, S.S.; Deng, L.; Stanczak, M.A.; Deng, L.; Verhagen, A.; Secrest, P.; Lusk, C.; et al. Engagement of myelomonocytic Siglecs by tumor-associated ligands modulates the innate immune response to cancer. Proc. Natl. Acad. Sci. USA 2014, 111, 14211–14216. [Google Scholar] [CrossRef]
- Boyd, C.R.; Orr, S.J.; Spence, S.; Burrows, J.F.; Elliott, J.; Carroll, H.P.; Brennan, K.; Ni Gabhann, J.; Coulter, W.A.; Johnston, J.A.; et al. Siglec-E is up-regulated and phosphorylated following lipopolysaccharide stimulation in order to limit TLR-driven cytokine production. J. Immunol. 2010, 183, 7703–7709. [Google Scholar] [CrossRef]
- Chen, G.-Y.; Brown, N.K.; Wu, W.; Khedri, Z.; Yu, H.; Chen, X.; van de Vlekkert, D.; D’Azzo, A.; Zheng, P.; Liu, Y. Broad and direct interaction between TLR and Siglec families of pattern recognition receptors and its regulation by Neu1. Elife 2014, 3. [Google Scholar] [CrossRef]
- McMillan, S.J.; Sharma, R.S.; McKenzie, E.J.; Richards, H.E.; Zhang, J.; Prescott, A.; Crocker, P.R. Siglec-E is a negative regulator of acute pulmonary neutrophil inflammation and suppresses CD11b b2-integrin-dependent signaling. Blood 2013, 121, 2084–2094. [Google Scholar] [CrossRef]
- Schwarz, F.; Pearce, O.M.; Wang, X.; Samraj, A.N.; Läubli, H.; Garcia, J.O.; Lin, H.; Fu, X.; Garcia-Bingman, A.; Secrest, P.; et al. Siglec receptors impact mammalian lifespan by modulating oxidative stress. Elife 2015, 4. [Google Scholar] [CrossRef]
- Claude, J.; Linnartz-Gerlach, B.; Kudin, A.P.; Kunz, W.S.; Neumann, H. Microglial CD33-Related Siglec-E Inhibits Neurotoxicity by Preventing the Phagocytosis-Associated Oxidative Burst. J. Neurosci. 2013, 33, 18270–18276. [Google Scholar] [CrossRef]
- Angata, T. Cloning and Characterization of a Novel Mouse Siglec, mSiglec-F. DIFFERENTIAL EVOLUTION OF THE MOUSE AND HUMAN (CD33) Siglec-3-RELATED GENE CLUSTERS. J. Boil. Chem. 2001, 276, 45128–45136. [Google Scholar] [CrossRef]
- Tateno, H.; Crocker, P.R.; Paulson, J.C. Mouse Siglec-F and human Siglec-8 are functionally convergent paralogs that are selectively expressed on eosinophils and recognize 6′-sulfo-sialyl Lewis X as a preferred glycan ligand. Glycobiology 2005, 15, 1125–1135. [Google Scholar] [CrossRef]
- Wielgat, P.; Braszko, J.J. The participation of sialic acids in microglia–neuron interactions. Cell. Immunol. 2012, 273, 17–22. [Google Scholar] [CrossRef]
- Blasius, A.L.; Colonna, M. Sampling and signaling in plasmacytoid dendritic cells: The potential roles of Siglec-H. Trends Immunol. 2006, 27, 255–260. [Google Scholar] [CrossRef]
- Konishi, H.; Kobayashi, M.; Kunisawa, T.; Imai, K.; Sayo, A.; Malissen, B.; Crocker, P.R.; Sato, K.; Kiyama, H. Siglec-H is a microglia-specific marker that discriminates microglia from CNS-associated macrophages and CNS-infiltrating monocytes. Glia 2017, 65, 1927–1943. [Google Scholar] [CrossRef]
- Perry, V.H.; Crocker, P.R.; Gordon, S. The blood–brain barrier regulates the expression of a macrophage sialic acid-binding receptor on microglia. J. Cell Sci. 1992, 101, 201–207. [Google Scholar]
- Mott, R.T.; Ait-Ghezala, G.; Town, T.; Mori, T.; Vendrame, M.; Zeng, J.; Ehrhart, J.; Mullan, M.; Tan, J.; Mullan, M.; et al. Neuronal expression of CD22: Novel mechanism for inhibiting microglial proinflammatory cytokine production. Glia 2004, 46, 369–379. [Google Scholar] [CrossRef]
- Pluvinage, J.V.; Haney, M.S.; Smith, B.A.H.; Sun, J.; Iram, T.; Bonanno, L.; Li, L.; Lee, D.P.; Morgens, D.W.; Yang, A.C.; et al. CD22 blockade restores homeostatic microglial phagocytosis in ageing brains. Nature 2019, 568, 187–192. [Google Scholar] [CrossRef]
- Johnson, P.W.; Abramow-Newerly, W.; Seilheimer, B.; Sadoul, R.; Tropak, M.B.; Arquint, M.; Dunn, R.J.; Schachner, M.; Roder, J.C. Recombinant myelin-associated glycoprotein confers neural adhesion and neurite outgrowth function. Neuron 1989, 3, 377–385. [Google Scholar] [CrossRef]
- Turnley, A.M.; Bartlett, P.F. MAG and MOG enhance neurite outgrowth of embryonic mouse spinal cord neurons. NeuroReport 1998, 9, 1987–1990. [Google Scholar] [CrossRef]
- Mukhopadhyay, G.; Doherty, P.; Walsh, F.S.; Crocker, P.R.; Filbin, M.T. A novel role for myelin-associated glycoprotein as an inhibitor of axonal regeneration. Neuron 1994, 13, 757–767. [Google Scholar] [CrossRef]
- Quarles, R.H. Myelin-associated glycoprotein (MAG): Past, present and beyond. J. Neurochem. 2007, 100, 1431–1448. [Google Scholar] [CrossRef]
- Cai, D.; Qiu, J.; Cao, Z.; McAtee, M.; Bregman, B.S.; Filbin, M.T. Neuronal Cyclic AMP Controls the Developmental Loss in Ability of Axons to Regenerate. J. Neurosci. 2001, 21, 4731–4739. [Google Scholar] [CrossRef] [PubMed]
- McKerracher, L.; Rosen, K.M. MAG, myelin and overcoming growth inhibition in the CNS. Front. Mol. Neurosci. 2015, 8. [Google Scholar] [CrossRef] [PubMed]
- Bartsch, U.; Bandtlow, C.E.; Schnell, L.; Bartsch, S.; Spillmann, A.A.; Rubin, B.P.; Hillenbrand, R.; Schwab, D.M.E.; Schachner, M. Lack of evidence that myelin-associated glycoprotein is a major inhibitor of axonal regeneration in the CNS. Neuron 1995, 15, 1375–1381. [Google Scholar] [CrossRef]
- Tang, S.; Woodhall, R.W.; Shen, Y.J.; Debellard, M.E.; Saffell, J.L.; Doherty, P.; Walsh, F.S.; Filbin, M.T. Soluble myelin-associated glycoprotein (MAG) found in vivo inhibits axonal regeneration. Mol. Cell. Neurosci. 1997, 9, 333–346. [Google Scholar] [CrossRef] [PubMed]
- Tang, S.; Qiu, J.; Nikulina, E.; Filbin, M.T. Soluble Myelin-Associated Glycoprotein Released from Damaged White Matter Inhibits Axonal Regeneration. Mol. Cell. Neurosci. 2001, 18, 259–269. [Google Scholar] [CrossRef] [PubMed]
- Linnartz, B.; Wang, Y.; Neumann, H. Microglial Immunoreceptor Tyrosine-Based Activation and Inhibition Motif Signaling in Neuroinflammation. Int. J. Alzheimer’s Dis. 2010, 2010, 1–7. [Google Scholar] [CrossRef]
- Wang, Y.; Neumann, H. Alleviation of Neurotoxicity by Microglial Human Siglec-11. J. Neurosci. 2010, 30, 3482–3488. [Google Scholar] [CrossRef] [PubMed]
- Pshezhetsky, A.V.; Ashmarina, M. Keeping it trim: Roles of neuraminidases in CNS function. Glycoconj. J. 2018, 35, 375–386. [Google Scholar] [CrossRef]
- Kopatz, J.; Beutner, C.; Welle, K.; Bodea, L.-G.; Reinhardt, J.; Claude, J.; Linnartz-Gerlach, B.; Neumann, H.; Linnartz-Gerlach, B. Siglec-h on activated microglia for recognition and engulfment of glioma cells. Glia 2013, 61, 1122–1133. [Google Scholar] [CrossRef]
- Groh, J.; Ribechini, E.; Stadler, D.; Schilling, T.; Lutz, M.B.; Martini, R. Sialoadhesin promotes neuroinflammation-related disease progression in two mouse models of CLN disease. Glia 2016, 64, 792–809. [Google Scholar] [CrossRef]
- Mabbott, N. How do PrPSc Prions Spread between Host Species, and within Hosts? Pathogens 2017, 6, 60. [Google Scholar] [CrossRef]
- Bradford, B.M.; Crocker, P.R.; Mabbott, N.A. Peripheral prion disease pathogenesis is unaltered in the absence of sialoadhesin (Siglec-1/CD169). Immunology 2014, 143, 120–129. [Google Scholar] [CrossRef]
- Ip, C.W.; Kroner, A.; Crocker, P.R.; Nave, K.-A.; Martini, R. Sialoadhesin deficiency ameliorates myelin degeneration and axonopathic changes in the CNS of PLP overexpressing mice. Neurobiol. Dis. 2007, 25, 105–111. [Google Scholar] [CrossRef]
- Bertram, L.; Lange, C.; Mullin, K.; Parkinson, M.; Hsiao, M.; Hogan, M.F.; Schjeide, B.M.M.; Hooli, B.; DiVito, J.; Ionita, I.; et al. Genome-wide Association Analysis Reveals Putative Alzheimer’s Disease Susceptibility Loci in Addition to APOE. Am. J. Hum. Genet. 2008, 83, 623. [Google Scholar] [CrossRef]
- Hollingworth, P.; Harold, D.; Sims, R.; Gerrish, A.; Lambert, J.C.; Carrasquillo, M.M.; Abraham, R.; Hamshere, M.L.; Pahwa, J.S.; Moskvina, V.; et al. Common variants at ABCA7, MS4A6A/MS4A4E, EPHA1, CD33 and CD2AP are associated with Alzheimer’s disease. Nat. Genet. 2011, 43, 429–435. [Google Scholar] [CrossRef]
- Walker, D.G.; Whetzel, A.M.; Serrano, G.; Sue, L.I.; Beach, T.G.; Lue, L.F. Association of CD33 polymorphism rs3865444 with Alzheimer’s disease pathology and CD33 expression in human cerebral cortex. Neurobiol. Aging 2015, 36, 571–582. [Google Scholar] [CrossRef]
- Naj, A.C.; Jun, G.; Beecham, G.W.; Wang, L.S.; Vardarajan, B.N.; Buros, J.; Gallins, P.J.; Buxbaum, J.D.; Jarvik, G.P.; Crane, P.K.; et al. Common variants at MS4A4/MS4A6E, CD2AP, CD33 and EPHA1 are associated with late-onset Alzheimer’s disease. Nat. Genet. 2011, 43, 436–441. [Google Scholar] [CrossRef]
- Griciuc, A.; Serrano-Pozo, A.; Parrado, A.R.; Lesinski, A.N.; Asselin, C.N.; Mullin, K.; Hooli, B.; Choi, S.H.; Hyman, B.T.; Tanzi, R.E. Alzheimer’s Disease Risk Gene CD33 Inhibits Microglial Uptake of Amyloid Beta. Neuron 2013, 78, 631–643. [Google Scholar] [CrossRef]
- Malik, M.; Simpson, J.F.; Parikh, I.; Wilfred, B.R.; Fardo, D.W.; Nelson, P.T.; Estus, S. CD33 Alzheimer’s Risk-Altering Polymorphism, CD33 Expression, and Exon 2 Splicing. J. Neurosci. 2013, 33, 13320–13325. [Google Scholar] [CrossRef]
- Raj, T.; Ryan, K.J.; Replogle, J.M.; Chibnik, L.B.; Rosenkrantz, L.; Tang, A.; Rothamel, K.; Stranger, B.E.; Bennett, D.A.; Evans, D.A.; et al. CD33: Increased inclusion of exon 2 implicates the Ig V-set domain in Alzheimer’s disease susceptibility. Hum. Mol. Genet. 2014, 23, 2729–2736. [Google Scholar] [CrossRef]
- Bradshaw, E.M.; Chibnik, L.B.; Keenan, B.T.; Ottoboni, L.; Raj, T.; Tang, A.; Rosenkrantz, L.L.; Imboywa, S.; Lee, M.; Von Korff, A.; et al. CD33 Alzheimer’s disease locus: Altered monocyte function and amyloid biology. Nat. Neurosci. 2013, 16, 848–850. [Google Scholar] [CrossRef]
- Siddiqui, S.S.; Springer, S.A.; Verhagen, A.; Sundaramurthy, V.; Alisson-Silva, F.; Jiang, W.; Ghosh, P.; Varki, A. The Alzheimer’s Disease–protective CD33 splice variant mediates adaptive loss of function via diversion to an intracellular pool. J. Biol. Chem. 2017, 292, 15312–15320. [Google Scholar] [CrossRef]
- Fruttiger, M.; Montag, D.; Schachner, M.; Martini, R. Crucial Role for the Myelin-associated Glycoprotein in the Maintenance of Axon-Myelin Integrity. Eur. J. Neurosci. 1995, 7, 511–515. [Google Scholar] [CrossRef]
- Bjartmar, C.; Trapp, B.D. Axonal and neuronal degeneration in multiple sclerosis: Mechanisms and functional consequences. Curr. Opin. Neurol. 2001, 14, 271–278. [Google Scholar] [CrossRef]
- Barbay, S.; Plautz, E.J.; Zoubina, E.; Frost, S.B.; Cramer, S.C.; Nudo, R.J. Effects of Postinfarct Myelin-Associated Glycoprotein Antibody Treatment on Motor Recovery and Motor Map Plasticity in Squirrel Monkeys. Stroke 2015, 46, 1620–1625. [Google Scholar] [CrossRef][Green Version]
- Dalakas, M.C. Advances in the diagnosis, immunopathogenesis and therapies of IgM-anti-MAG antibody-mediated neuropathies. Ther. Adv. Neurol. Disord. 2018, 11. [Google Scholar] [CrossRef]
- Ilyas, A.A.; Quarles, R.H.; MacIntosh, T.D.; Dobersen, M.J.; Trapp, B.D.; Dalakas, M.C.; Brady, R.O. IgM in a human neuropathy related to paraproteinemia binds to a carbohydrate determinant in the myelin-associated glycoprotein and to a ganglioside. Proc. Natl. Acad. Sci. USA 1984, 81, 1225–1229. [Google Scholar] [CrossRef]
- Dalakas, M.C. Pathogenesis and Treatment of Anti-MAG Neuropathy. Curr. Treat. Options Neurol. 2010, 12, 71–83. [Google Scholar] [CrossRef]
- Brennaman, L.H.; Maness, P.F. NCAM in Neuropsychiatric and Neurodegenerative Disorders. In Structure and Function of the Neural Cell Adhesion Molecule NCAM; Berezin, V., Ed.; Springer: New York, NY, USA, 2009. [Google Scholar]
- Vawter, M.P. Dysregulation of the neural cell adhesion molecule and neuropsychiatric disorders. Eur. J. Pharmacol. 2000, 405, 385–395. [Google Scholar] [CrossRef]
- Pham, M.T.; Pollock, K.M.; Rose, M.D.; Cary, W.A.; Stewart, H.R.; Zhou, P.; Nolta, J.A.; Waldau, B. Generation of human vascularized brain organoids. NeuroReport 2018, 29, 588–593. [Google Scholar] [CrossRef]
- Oetke, C.; Vinson, M.C.; Jones, C.; Crocker, P.R. Sialoadhesin-Deficient Mice Exhibit Subtle Changes in B- and T-Cell Populations and Reduced Immunoglobulin M Levels. Mol. Cell. Boil. 2006, 26, 1549–1557. [Google Scholar] [CrossRef]
- Nitschke, L.; Carsetti, R.; Ocker, B.; Köhler, G.; Lamers, M.C. CD22 is a negative regulator of B-cell receptor signalling. Curr. Boil. 1997, 7, 133–143. [Google Scholar] [CrossRef]
- Samardzic, T.; Marinkovic, D.; Danzer, C.P.; Gerlach, J.; Nitschke, L.; Wirth, T. Reduction of marginal zone B cells in CD22-deficient mice. Eur. J. Immunol. 2002, 32, 561–567. [Google Scholar] [CrossRef]
- Li, M.; Shibata, A.; Li, C.; Braun, P.E.; McKerracher, L.; Roder, J.; Kater, S.B.; David, S. Myelin-associated glycoprotein inhibits neurite/axon growth and causes growth cone collapse. J. Neurosci. Res. 1996, 46, 404–414. [Google Scholar] [CrossRef]
- DeBellard, M.-E.; Tang, S.; Mukhopadhyay, G.; Shen, Y.-J.; Filbin, M.T. Myelin-Associated Glycoprotein Inhibits Axonal Regeneration from a Variety of Neurons via Interaction with a Sialoglycoprotein. Mol. Cell. Neurosci. 1996, 7, 89–101. [Google Scholar] [CrossRef]
- Montag, D.; Giese, K.P.; Bartsch, U.; Martini, R.; Lang, Y.; Blüthmann, H.; Karthigasan, J.; Kirschner, D.A.; Wintergerst, E.S.; Nave, K.-A.; et al. Mice deficient for the glycoprotein show subtle abnormalities in myelin. Neuron 1994, 13, 229–246. [Google Scholar] [CrossRef]
- Karlstetter, M.; Kopatz, J.; Aslanidis, A.; Shahraz, A.; Caramoy, A.; Linnartz-Gerlach, B.; Lin, Y.; Lückoff, A.; Fauser, S.; Düker, K.; et al. Polysialic acid blocks mononuclear phagocyte reactivity, inhibits complement activation, and protects from vascular damage in the retina. EMBO Mol. Med. 2017, 9, 154–166. [Google Scholar] [CrossRef]
- Zhang, M.; Angata, T.; Cho, J.Y.; Miller, M.; Broide, D.H.; Varki, A. Defining the in vivo function of Siglec-F, a CD33-related Siglec expressed on mouse eosinophils. Blood 2007, 109, 4280–4287. [Google Scholar] [CrossRef]
- McMillan, S.J.; Richards, H.E.; Crocker, P.R. Siglec-F-dependent negative regulation of allergen-induced eosinophilia depends critically on the experimental model. Immunol. Lett. 2014, 160, 11–16. [Google Scholar] [CrossRef]
- Schmitt, H.; Sell, S.; Koch, J.; Seefried, M.; Sonnewald, S.; Daniel, C.; Winkler, T.H.; Nitschke, L. Siglec-H protects from virus-triggered severe systemic autoimmunity. J. Exp. Med. 2016, 213, 1627–1644. [Google Scholar] [CrossRef]


© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Siddiqui, S.S.; Matar, R.; Merheb, M.; Hodeify, R.; Vazhappilly, C.G.; Marton, J.; Shamsuddin, S.A.; Al Zouabi, H. Siglecs in Brain Function and Neurological Disorders. Cells 2019, 8, 1125. https://doi.org/10.3390/cells8101125
Siddiqui SS, Matar R, Merheb M, Hodeify R, Vazhappilly CG, Marton J, Shamsuddin SA, Al Zouabi H. Siglecs in Brain Function and Neurological Disorders. Cells. 2019; 8(10):1125. https://doi.org/10.3390/cells8101125
Chicago/Turabian StyleSiddiqui, Shoib Sarwar, Rachel Matar, Maxime Merheb, Rawad Hodeify, Cijo George Vazhappilly, John Marton, Syed Azharuddin Shamsuddin, and Hussain Al Zouabi. 2019. "Siglecs in Brain Function and Neurological Disorders" Cells 8, no. 10: 1125. https://doi.org/10.3390/cells8101125
APA StyleSiddiqui, S. S., Matar, R., Merheb, M., Hodeify, R., Vazhappilly, C. G., Marton, J., Shamsuddin, S. A., & Al Zouabi, H. (2019). Siglecs in Brain Function and Neurological Disorders. Cells, 8(10), 1125. https://doi.org/10.3390/cells8101125