
Pleiotrophin and the Expression of Its Receptors during Development of the Human Cerebellar Cortex
 ,
,  , ,
, ,  and
and 
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Human Paraffin-Embedded Tissues
2.2. Tissue Preparation and Histological Staining
2.3. Immunohistochemistry (IHC)
2.4. Histological Analysis
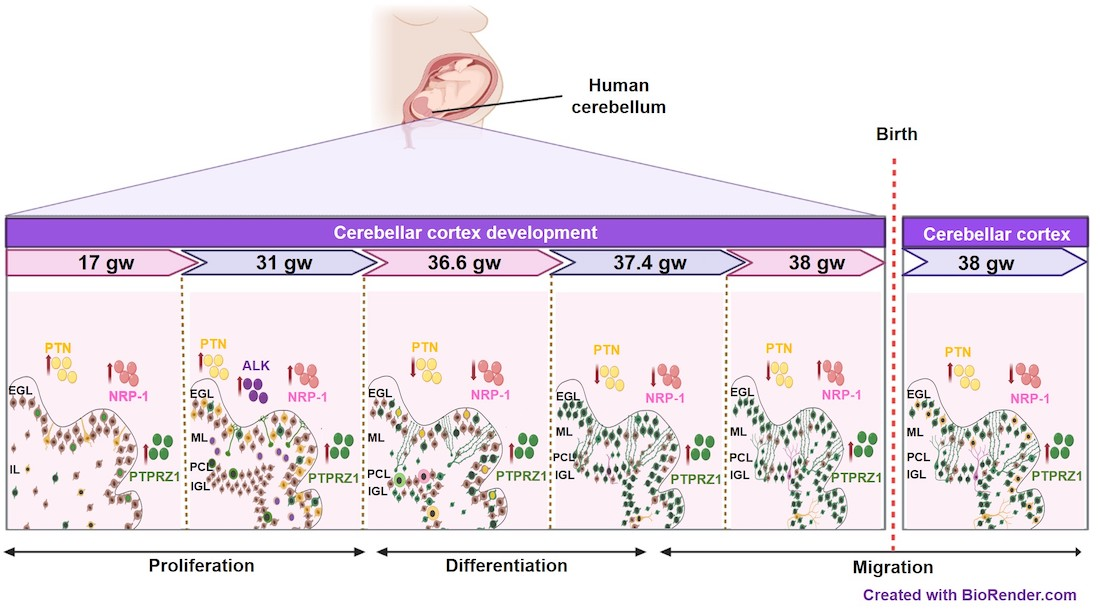
3. Results
3.1. The Topological Architecture of the Developing Human Cerebellar Cortex
3.2. Pleiotrophin Cell-Type Expression Changes during Cerebellar Cortex Development in Humans
3.3. PTN Receptor Expression Spatiotemporally Changes in Cerebellar Cortex Development
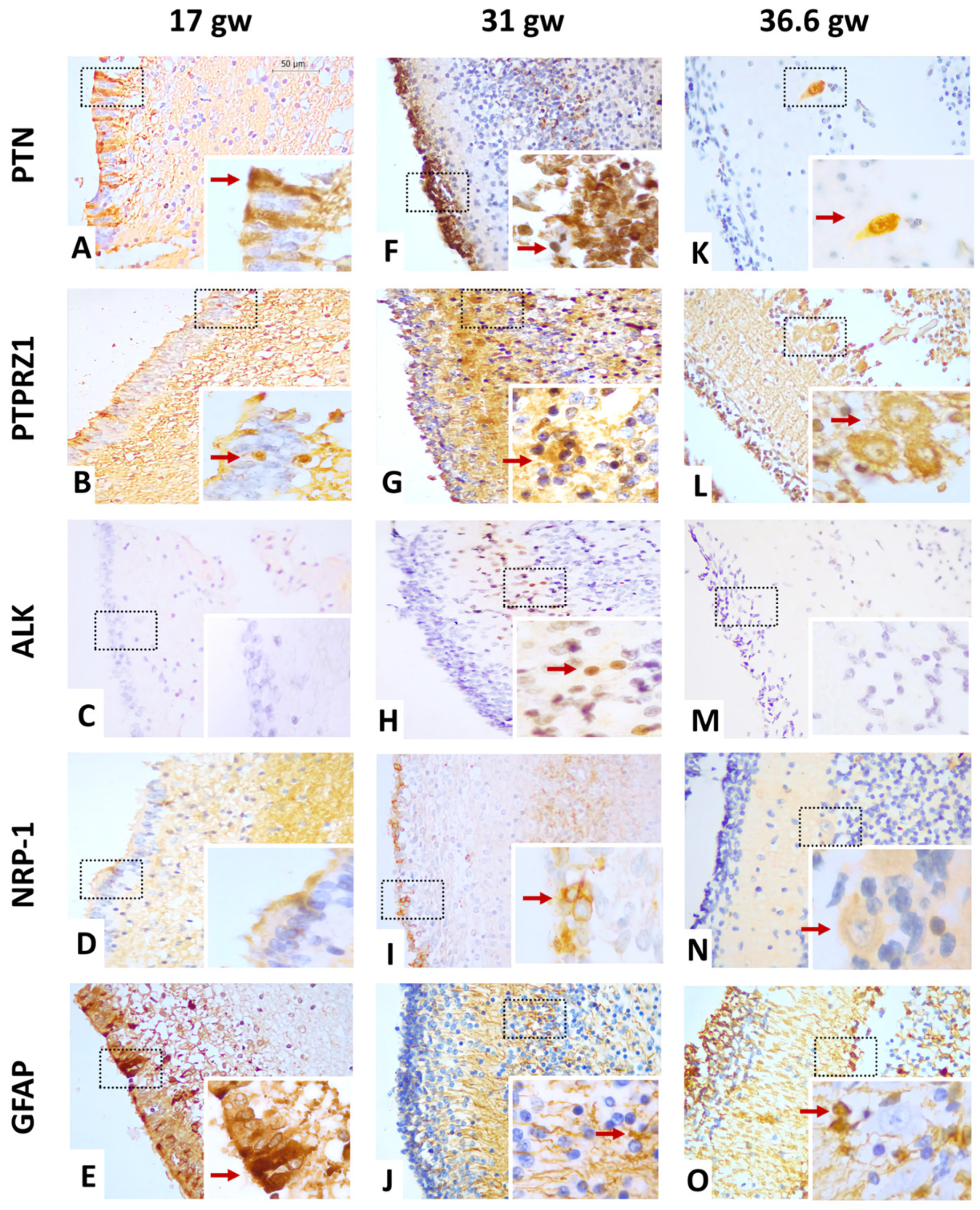
3.3.1. Gestational Week 17
3.3.2. Gestational Week 31
3.3.3. Gestational Week 36.6
3.3.4. Gestational Week 37.4
3.3.5. Gestational Week 38
3.3.6. Neonatal Sample
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Haldipur, P.; Bharti, U.; Alberti, C.; Sarkar, C.; Gulati, G.; Iyengar, S.; Gressens, P.; Mani, S. Preterm Delivery Disrupts the Developmental Program of the Cerebellum. PLoS ONE 2011, 6, e23449. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Larsell, O. The Development of the Cerebellum in Man in Relation to Its Comparative Anatomy. J. Comp. Neurol. 1947, 87, 85–129. [Google Scholar] [CrossRef] [PubMed]
- Rakic, P.; Sidman, R.L. Histogenesis of Cortical Layers in Human Cerebellum, Particularly the Lamina Dissecans. J. Comp. Neurol. 1970, 139, 473–500. [Google Scholar] [CrossRef] [PubMed]
- Amore, G.; Spoto, G.; Ieni, A.; Vetri, L.; Quatrosi, G.; Di Rosa, G.; Nicotera, A.G. A Focus on the Cerebellum: From Embryogenesis to an Age-Related Clinical Perspective. Front. Syst. Neurosci. 2021, 15, 646052. [Google Scholar] [CrossRef]
- Millen, K.J.; Gleeson, J.G. Cerebellar Development and Disease. Curr. Opin. Neurobiol. 2008, 18, 12–19. [Google Scholar] [CrossRef] [Green Version]
- Van Essen, M.J.; Nayler, S.; Becker, E.B.E.; Jacob, J. Deconstructing Cerebellar Development Cell by Cell. PLoS Genet. 2020, 16, e1008630. [Google Scholar] [CrossRef] [Green Version]
- Yachnis, A.T.; Rorke, L.B.; Lee, V.M.-Y.; Trojanowski, J.Q. Expression of Neuronal and Glial Polypeptides during Histogenesis of the Human Cerebellar Cortex Including Observations on the Dentate Nucleus. J. Comp. Neurol. 1993, 334, 356–369. [Google Scholar] [CrossRef]
- Levitt, P.; Rakic, P. Immunoperoxidase Localization of Glial Fibrillary Acidic Protein in Radial Glial Cells and Astrocytes of the Developing Rhesus Monkey Brain. J. Comp. Neurol. 1980, 193, 815–840. [Google Scholar] [CrossRef]
- DeAzevedo, L.C.; Fallet, C.; Moura-Neto, V.; Daumas-Duport, C.; Hedin-Pereira, C.; Lent, R. Cortical Radial Glial Cells in Human Fetuses: Depth-Correlated Transformation into Astrocytes. J. Neurobiol. 2003, 55, 288–298. [Google Scholar] [CrossRef]
- Sievers, J.; Pehlemann, F.W.; Gude, S.; Hartmann, D.; Berry, M. The Development of the Radial Glial Scaffold of the Cerebellar Cortex from GFAP-Positive Cells in the External Granular Layer. J. Neurocytol. 1994, 23, 97–115. [Google Scholar] [CrossRef]
- Williamson, D.; Schwalbe, E.C.; Bailey, S.; Clifford, S.C. Timing Is Everything: A Connection between Medulloblastoma Prognosis and Fetal Cerebellar Development. Neuropathol. Appl. Neurobiol. 2023, 49, e12903. [Google Scholar] [CrossRef]
- Matsumoto, K.; Wanaka, A.; Takatsuji, K.; Muramatsu, H.; Muramatsu, T.; Tohyama, M. A Novel Family of Heparin-Binding Growth Factors, Pleiotrophin and Midkine, Is Expressed in the Developing Rat Cerebral Cortex. Dev. Brain Res. 1994, 79, 229–241. [Google Scholar] [CrossRef]
- Hamza, M.M.; Rey, S.A.; Hilber, P.; Arabo, A.; Collin, T.; Vaudry, D.; Burel, D. Early Disruption of Extracellular Pleiotrophin Distribution Alters Cerebellar Neuronal Circuit Development and Function. Mol. Neurobiol. 2016, 53, 5203–5216. [Google Scholar] [CrossRef] [Green Version]
- Basille-Dugay, M.; Hamza, M.M.; Tassery, C.; Parent, B.; Raoult, E.; Bénard, M.; Raisman-Vozari, R.; Vaudry, D.; Burel, D.C. Spatio-Temporal Characterization of the Pleiotrophinergic System in Mouse Cerebellum: Evidence for Its Key Role during Ontogenesis. Exp. Neurol. 2013, 247, 537–551. [Google Scholar] [CrossRef]
- González-Castillo, C.; Ortuño-Sahagún, D.; Guzmán-Brambila, C.; Márquez-Aguirre, A.L.; Raisman-Vozari, R.; Pallás, M.; Rojas-Mayorquín, A.E. The Absence of Pleiotrophin Modulates Gene Expression in the Hippocampus in Vivo and in Cerebellar Granule Cells in Vitro. Mol. Cell. Neurosci. 2016, 75, 113–121. [Google Scholar] [CrossRef]
- Wewetzer, K.; Rauvala, H.; Unsicker, K. Immunocytochemical Localization of the Heparin-Binding Growth-Associated Molecule (HB-GAM) in the Developing and Adult Rat Cerebellar Cortex. Brain Res. 1995, 693, 31–38. [Google Scholar] [CrossRef]
- González-Castillo, C.; Ortuño-Sahagún, D.; Guzmán-Brambila, C.; Pallás, M.; Rojas-Mayorquín, A.E. Pleiotrophin as a Central Nervous System Neuromodulator, Evidences from the Hippocampus. Front. Cell. Neurosci. 2015, 8, 443. [Google Scholar] [CrossRef] [Green Version]
- Knudsen, A.M.; Halle, B.; Cédile, O.; Burton, M.; Baun, C.; Thisgaard, H.; Anand, A.; Hubert, C.; Thomassen, M.; Michaelsen, S.R.; et al. Surgical Resection of Glioblastomas Induces Pleiotrophin-Mediated Self-Renewal of Glioblastoma Stem Cells in Recurrent Tumors. Neuro-Oncol. 2022, 24, 1074–1087. [Google Scholar] [CrossRef]
- Hu, B.; Qin, C.; Li, L.; Wei, L.; Mo, X.; Fan, H.; Lei, Y.; Wei, F.; Zou, D. Midkine Promotes Glioblastoma Progression via PI3K-Akt Signaling. Cancer Cell Int. 2021, 21, 509. [Google Scholar] [CrossRef]
- Hauser, S.; Kaminski, A.; Syring, I.; Holdenrieder, S.; Dieckmann, K.-P.; Muller, S.C.; Ellinger, J. Evaluation of Serum Biomarkers (FGF-2, HGF, MIF and PTN) in Patients with Testicular Germ Cell Cancer. In Vivo 2019, 33, 1935–1940. [Google Scholar] [CrossRef]
- Zheng, L.; Li, H.; Huang, J.; Shin, J.; Luo, S.; Guo, C.; Zhao, Y.; Li, F. Serum Midkine Levels for the Diagnosis and Assessment of Response to Interventional Therapy in Patients with Hepatocellular Carcinoma. J. Interv. Med. 2021, 4, 39–45. [Google Scholar] [CrossRef] [PubMed]
- Kilpeläinen, I.; Kaksonen, M.; Kinnunen, T.; Avikainen, H.; Fath, M.; Linhardt, R.J.; Raulo, E.; Rauvala, H. Heparin-Binding Growth-Associated Molecule Contains Two Heparin-Binding β-Sheet Domains That Are Homologous to the Thrombospondin Type I Repeat. J. Biol. Chem. 2000, 275, 13564–13570. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rojas-Mayorquín, A.E.; Ortuño-Sahagún, D. Pleiotrophin. In Encyclopedia of Signaling Molecules; Choi, S., Ed.; Springer: New York, NY, USA, 2016; pp. 1–15. ISBN 978-1-4614-6438-9. [Google Scholar]
- Rauvala, H.; Vanhala, A.; Castre’n, E.; Nolo, R.; Raulo, E.; Merenmies, J.; Panula, P. Expression of HB-GAM (Heparin-Binding Growth-Associated Molecules) in the Pathways of Developing Axonal Processes in Vivo and Neurite Outgrowth in Vitro Induced by HB-GAM. Dev. Brain Res. 1994, 79, 157–176. [Google Scholar] [CrossRef] [PubMed]
- Maeda, N.; Noda, M. Involvement of Receptor-like Protein Tyrosine Phosphatase ζ/RPTPβ and Its Ligand Pleiotrophin/Heparin-Binding Growth-Associated Molecule (HB-GAM) in Neuronal Migration. J. Cell Biol. 1998, 142, 203–216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yanagisawa, H.; Komuta, Y.; Kawano, H.; Toyoda, M.; Sango, K. Pleiotrophin Induces Neurite Outgrowth and Up-Regulates Growth-Associated Protein (GAP)-43 MRNA through the ALK/GSK3β/β-Catenin Signaling in Developing Mouse Neurons. Neurosci. Res. 2010, 66, 111–116. [Google Scholar] [CrossRef] [PubMed]
- Elahouel, R.; Blanc, C.; Carpentier, G.; Frechault, S.; Cascone, I.; Destouches, D.; Delbé, J.; Courty, J.; Hamma-Kourbali, Y. Pleiotrophin Exerts Its Migration and Invasion Effect through the Neuropilin-1 Pathway. Neoplasia 2015, 17, 613–624. [Google Scholar] [CrossRef] [Green Version]
- Papadimitriou, E.; Mourkogianni, E.; Ntenekou, D.; Christopoulou, M.; Koutsioumpa, M.; Lamprou, M. On the Role of Pleiotrophin and Its Receptors in Development and Angiogenesis. Int. J. Dev. Biol. 2022, 66, 115–124. [Google Scholar] [CrossRef]
- Canoll, P.D.; Barnea, G.; Levy, J.B.; Sap, J.; Ehrlich, M.; Silvennoinen, O.; Schlessinger, J.; Musacchio, J.M. The Expression of a Novel Receptor-Type Tyrosine Phosphatase Suggests a Role in Morphogenesis and Plasticity of the Nervous System. Dev. Brain Res. 1993, 75, 293–298. [Google Scholar] [CrossRef]
- Meng, K.; Rodríguez-Peña, A.; Dimitrov, T.; Chen, W.; Yamin, M.; Noda, M.; Deuel, T.F. Pleiotrophin Signals Increased Tyrosine Phosphorylation of β-Catenin through Inactivation of the Intrinsic Catalytic Activity of the Receptor-Type Protein Tyrosine Phosphatase β/ζ. Proc. Natl. Acad. Sci. USA 2000, 97, 2603–2608. [Google Scholar] [CrossRef] [Green Version]
- Nishiwaki, T.; Maeda, N.; Noda, M. Characterization and Developmental Regulation of Proteoglycan-Type Protein Tyrosine Phosphatase /RPTP Isoforms. J. Biochem. 1998, 123, 458–467. [Google Scholar] [CrossRef]
- Morris, S.W.; Naeve, C.; Mathew, P.; James, P.L.; Kirstein, M.N.; Cui, X.; Witte, D.P. ALK, the Chromosome 2 Gene Locus Altered by the t(2;5) in Non-Hodgkin’s Lymphoma, Encodes a Novel Neural Receptor Tyrosine Kinase That Is Highly Related to Leukocyte Tyrosine Kinase (LTK). Oncogene 1997, 14, 2175–2188. [Google Scholar] [CrossRef] [Green Version]
- Vernersson, E.; Khoo, N.K.S.; Henriksson, M.L.; Roos, G.; Palmer, R.H.; Hallberg, B. Characterization of the Expression of the ALK Receptor Tyrosine Kinase in Mice. Gene Expr. Patterns 2006, 6, 448–461. [Google Scholar] [CrossRef]
- Papadimitriou, E.; Pantazaka, E.; Castana, P.; Tsalios, T.; Polyzos, A.; Beis, D. Pleiotrophin and Its Receptor Protein Tyrosine Phosphatase Beta/Zeta as Regulators of Angiogenesis and Cancer. Biochim. Biophys. Acta BBA Rev. Cancer 2016, 1866, 252–265. [Google Scholar] [CrossRef]
- Ellis, L.M. The Role of Neuropilins in Cancer. Mol. Cancer Ther. 2006, 5, 1099–1107. [Google Scholar] [CrossRef] [Green Version]
- Kitsukawa, T.; Shimizu, M.; Sanbo, M.; Hirata, T.; Taniguchi, M.; Bekku, Y.; Yagi, T.; Fujisawa, H. Neuropilin–Semaphorin III/D-Mediated Chemorepulsive Signals Play a Crucial Role in Peripheral Nerve Projection in Mice. Neuron 1997, 19, 995–1005. [Google Scholar] [CrossRef] [Green Version]
- Cheng, L.; Jia, H.; Löhr, M.; Bagherzadeh, A.; Holmes, D.I.R.; Selwood, D.; Zachary, I. Anti-Chemorepulsive Effects of Vascular Endothelial Growth Factor and Placental Growth Factor-2 in Dorsal Root Ganglion Neurons Are Mediated via Neuropilin-1 and Cyclooxygenase-Derived Prostanoid Production. J. Biol. Chem. 2004, 279, 30654–30661. [Google Scholar] [CrossRef] [Green Version]
- Bankhead, P.; Loughrey, M.B.; Fernández, J.A.; Dombrowski, Y.; McArt, D.G.; Dunne, P.D.; McQuaid, S.; Gray, R.T.; Murray, L.J.; Coleman, H.G.; et al. QuPath: Open Source Software for Digital Pathology Image Analysis. Sci. Rep. 2017, 7, 16878. [Google Scholar] [CrossRef] [Green Version]
- Snyder, S.E.; Li, J.; Elyse Schauwecker, P.; McNeill, T.H.; Salton, S.R.J. Comparison of RPTPζ/β, Phosphacan, and TrkB MRNA Expression in the Developing and Adult Rat Nervous System and Induction of RPTPζ/β and Phosphacan MRNA Following Brain Injury. Mol. Brain Res. 1996, 40, 79–96. [Google Scholar] [CrossRef]
- Perez-Pinera, P.; Chang, Y.; Deuel, T.F. Pleiotrophin, A Multifunctional Tumor Promoter Through Induction of Tumor Angiogenesis, Remodeling of the Tumor Microenvironment, and Activation of Stromal Fibroblasts. Cell Cycle 2007, 6, 2877–2883. [Google Scholar] [CrossRef] [Green Version]
- Di Paolo, D.; Pastorino, F.; Brignole, C.; Corrias, M.V.; Emionite, L.; Cilli, M.; Tamma, R.; Priddy, L.; Amaro, A.; Ferrari, D.; et al. Combined Replenishment of MiR-34a and Let-7b by Targeted Nanoparticles Inhibits Tumor Growth in Neuroblastoma Preclinical Models. Small 2020, 16, 1906426. [Google Scholar] [CrossRef]
- Mangieri, R.A.; Maier, E.Y.; Buske, T.R.; Lasek, A.W.; Morrisett, R.A. Anaplastic Lymphoma Kinase Is a Regulator of Alcohol Consumption and Excitatory Synaptic Plasticity in the Nucleus Accumbens Shell. Front. Pharmacol. 2017, 8, 533. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.; Zhang, Y.; Petrus, M.N.; Xiao, W.; Nicolae, A.; Raffeld, M.; Pittaluga, S.; Bamford, R.N.; Nakagawa, M.; Ouyang, S.T.; et al. Cytokine Receptor Signaling Is Required for the Survival of ALK− Anaplastic Large Cell Lymphoma, Even in the Presence of JAK1/STAT3 Mutations. Proc. Natl. Acad. Sci. USA 2017, 114, 3975–3980. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lopez-Delisle, L.; Pierre-Eugène, C.; Louis-Brennetot, C.; Surdez, D.; Raynal, V.; Baulande, S.; Boeva, V.; Grossetête-Lalami, S.; Combaret, V.; Peuchmaur, M.; et al. Activated ALK Signals through the ERK–ETV5–RET Pathway to Drive Neuroblastoma Oncogenesis. Oncogene 2018, 37, 1417–1429. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Slupianek, A.; Nieborowska-Skorska, M.; Hoser, G.; Morrione, A.; Majewski, M.; Xue, L.; Morris, S.W.; Wasik, M.A.; Skorski, T. Role of Phosphatidylinositol 3-Kinase-Akt Pathway in Nucleophosmin/Anaplastic Lymphoma Kinase-Mediated Lymphomagenesis. Cancer Res. 2001, 61, 2194–2199. [Google Scholar] [PubMed]
- Volpe, J.J. Cerebellum of the Premature Infant: Rapidly Developing, Vulnerable, Clinically Important. J. Child Neurol. 2009, 24, 1085–1104. [Google Scholar] [CrossRef] [Green Version]
- Iwahara, T.; Fujimoto, J.; Wen, D.; Cupples, R.; Bucay, N.; Arakawa, T.; Mori, S.; Ratzkin, B.; Yamamoto, T. Molecular Characterization of ALK, a Receptor Tyrosine Kinase Expressed Specifically in the Nervous System. Oncogene 1997, 14, 439–449. [Google Scholar] [CrossRef] [Green Version]
- Carén, H.; Abel, F.; Kogner, P.; Martinsson, T. High Incidence of DNA Mutations and Gene Amplifications of the ALK Gene in Advanced Sporadic Neuroblastoma Tumours. Biochem. J. 2008, 416, 153–159. [Google Scholar] [CrossRef] [Green Version]
- Janoueix-Lerosey, I.; Lequin, D.; Brugières, L.; Ribeiro, A.; de Pontual, L.; Combaret, V.; Raynal, V.; Puisieux, A.; Schleiermacher, G.; Pierron, G.; et al. Somatic and Germline Activating Mutations of the ALK Kinase Receptor in Neuroblastoma. Nature 2008, 455, 967–970. [Google Scholar] [CrossRef]
- Maris, J.M. Recent Advances in Neuroblastoma. N. Engl. J. Med. 2010, 362, 2202–2211. [Google Scholar] [CrossRef] [Green Version]
- Marshall, G.M.; Carter, D.R.; Cheung, B.B.; Liu, T.; Mateos, M.K.; Meyerowitz, J.G.; Weiss, W.A. The Prenatal Origins of Cancer. Nat. Rev. Cancer 2014, 14, 277–289. [Google Scholar] [CrossRef] [Green Version]
- Tanaka, M.; Maeda, N.; Noda, M.; Marunouchi, T. A Chondroitin Sulfate Proteoglycan PTPζ/RPTPβ Regulates the Morphogenesis of Purkinje Cell Dendrites in the Developing Cerebellum. J. Neurosci. 2003, 23, 2804–2814. [Google Scholar] [CrossRef] [Green Version]
- Haldipur, P.; Dang, D.; Millen, K.J. Embryology. In Handbook of Clinical Neurology; Elsevier: Amsterdam, The Netherlands, 2018; Volume 154, pp. 29–44. ISBN 978-0-444-63956-1. [Google Scholar]
- Chizhikov, V.; Millen, K.J. Development and Malformations of the Cerebellum in Mice. Mol. Genet. Metab. 2003, 80, 54–65. [Google Scholar] [CrossRef]
- Kolodkin, A.L.; Levengood, D.V.; Rowe, E.G.; Tai, Y.-T.; Giger, R.J.; Ginty, D.D. Neuropilin Is a Semaphorin III Receptor. Cell 1997, 90, 753–762. [Google Scholar] [CrossRef] [Green Version]
- He, Z.; Tessier-Lavigne, M. Neuropilin Is a Receptor for the Axonal Chemorepellent Semaphorin III. Cell 1997, 90, 739–751. [Google Scholar] [CrossRef] [Green Version]
- Neufeld, G.; Kessler, O.; Herzog, Y. The Interaction of Neuropilin-1 and Neuropilin-2 with Tyrosine-Kinase Receptors for VEGF. In Neuropilin; Bagnard, D., Ed.; Advances in Experimental Medicine and Biology; Springer: Boston, MA, USA, 2002; Volume 515, pp. 81–90. ISBN 978-0-306-47416-3. [Google Scholar]
- Aldinger, K.A.; Thomson, Z.; Phelps, I.G.; Haldipur, P.; Deng, M.; Timms, A.E.; Hirano, M.; Santpere, G.; Roco, C.; Rosenberg, A.B.; et al. Spatial and Cell Type Transcriptional Landscape of Human Cerebellar Development. Nat. Neurosci. 2021, 24, 1163–1175. [Google Scholar] [CrossRef]
- Zecevic, N.; Rakic, P. Differentiation of Purkinje Cells and Their Relationship to Other Components of Developing Cerebellar Cortex in Man. J. Comp. Neurol. 1976, 167, 27–47. [Google Scholar] [CrossRef]
- Lafont, D.; Adage, T.; Gréco, B.; Zaratin, P. A Novel Role for Receptor like Protein Tyrosine Phosphatase Zeta in Modulation of Sensorimotor Responses to Noxious Stimuli: Evidences from Knockout Mice Studies. Behav. Brain Res. 2009, 201, 29–40. [Google Scholar] [CrossRef]
- Cohen, S.; Shoshana, O.; Zelman-Toister, E.; Maharshak, N.; Binsky-Ehrenreich, I.; Gordin, M.; Hazan-Halevy, I.; Herishanu, Y.; Shvidel, L.; Haran, M.; et al. The Cytokine Midkine and Its Receptor RPTPζ Regulate B Cell Survival in a Pathway Induced by CD74. J. Immunol. 2012, 188, 259–269. [Google Scholar] [CrossRef] [Green Version]
- Shintani, T.; Watanabe, E.; Maeda, N.; Noda, M. Neurons as Well as Astrocytes Express Proteoglycan-Type Protein Tyrosine Phosphatase ζ/RPTPβ: Analysis of Mice in Which the PTPζ/RPTPβ Gene Was Replaced with the LacZ Gene. Neurosci. Lett. 1998, 247, 135–138. [Google Scholar] [CrossRef]
- Del Campo, M.; Fernández-Calle, R.; Vicente-Rodríguez, M.; Martín Martínez, S.; Gramage, E.; Zapico, J.M.; Haro, M.; Herradon, G. Role of Receptor Protein Tyrosine Phosphatase β/ζ in Neuron–Microglia Communication in a Cellular Model of Parkinson’s Disease. Int. J. Mol. Sci. 2021, 22, 6646. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Age | Sex | Cause of Death |
|---|---|---|
| Fetal samples | ||
| 17 gw | Male | Interruption of placental blood flow |
| 31 gw | Female | Congenital heart disease |
| 36.6 gw | Male | Umbilical venous thrombosis |
| 37.4 gw | Female | Amniotic fluid aspiration |
| 38 gw | Female | Placental hypoxia–ischemia |
| Neonatal sample | ||
| 38 gw | Female | Respiratory failure |
| Gestational Weeks | EGL Thickness (µm) (Mean ± SD) | EGL Cell Density (Number/5000 µm2) (Mean ± SD) | PC Area (µm2) (Mean ± SD) | ML Thickness (µm) (Mean ± SD) |
|---|---|---|---|---|
| 17 | 29.4 ± 6.3 | 47.0 ± 8.4 | N/A | N/A |
| 31 | 42.1 ± 10.9 | 77.3 ± 22.3 | 81.7 ± 25.9 | 48.6 ± 14.7 |
| 36.6 | 76.9 ± 29.3 | 143.0 ± 43.8 | 247.6 ± 44.9 | 111.5 ± 22.0 |
| 37.4 | 32.2 ± 6.3 | 66.3 ± 11.9 | 261.9 ± 11.5 | 54.4 ± 7.9 |
| 38 | 51.4 ± 9.1 | 81.0 ± 9.9 | 278.7 ± 23.5 | 56.6 ± 11.8 |
| Postnatal | 26.9 ± 4.1 | 51.5 ± 5.6 | 307.2 ± 30.5 | 49.3 ± 6.4 |
| EGL | PCL | ML | IGL | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Staining Intensity | 1+ | 2+ | 3+ | 1+ | 2+ | 3+ | 1+ | 2+ | 3+ | 1+ | 2+ | 3+ |
| 17 gw | 5.5% | 6.1% | 1.7% | N/A | N/A | N/A | N/A | N/A | N/A | 0% | 0% | 0% |
| 31 gw | 7.5% | 9.8% | 15.9% | 0% | 0% | 0% | 0% | 0% | 0% | 21.3% | 12.0% | 6.9% |
| 36.6 gw | 24.0% | 7.4% | 1.8% | 0% | 0% | 66.% | 0% | 0% | 0% | 0% | 0% | 0% |
| 37.4 gw | 0% | 0% | 0% | 33.3% | 0% | 33.3% | 0% | 0% | 0% | 0% | 0% | 0% |
| 38 gw | 0% | 0% | 0% | 33.3% | 0% | 0% | 0% | 0% | 0% | 0% | 0% | 0% |
| Postnatal | 20.6% | 8.9% | 3.7% | 0% | 0% | 0% | 0% | 0% | 0% | 0% | 0% | 0% |
| EGL | PCL | ML | IGL | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Staining Intensity | 1+ | 2+ | 3+ | 1+ | 2+ | 3+ | 1+ | 2+ | 3+ | 1+ | 2+ | 3+ |
| 17 gw | 4.3% | 6.3% | 2.5% | N/A | N/A | N/A | N/A | N/A | N/A | 2.2% | 9.3% | 34.4% |
| 31 gw | 6.6% | 12.7% | 13.8% | 0% | 11.1% | 22.2% | 0.1% | 8.1% | 24.9% | 4.2% | 17.6% | 15.2% |
| 36.6 gw | 1.8% | 7.3% | 24.0% | 7.6% | 20.4% | 5.1% | 5.5% | 10.2% | 8.8% | 8.1% | 13.4% | 11.6% |
| 37.4 gw | 11.9% | 10.3% | 10.2% | 0% | 14.2% | 19.0% | 2.3% | 11.9% | 16.2% | 7.3% | 15.9% | 9.6% |
| 38 gw | 24.4% | 7.5% | 1.3% | 0% | 0% | 33.3% | 4% | 16% | 13.3% | 20.2% | 9.4% | 3.6% |
| Postnatal | 21.8% | 11.0% | 0.4% | 0% | 9.0% | 24.2% | 30.7% | 20% | 2.6% | 16.1% | 15.1% | 1.9% |
| EGL | PCL | ML | IGL | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Staining Intensity | 1+ | 2+ | 3+ | 1+ | 2+ | 3+ | 1+ | 2+ | 3+ | 1+ | 2+ | 3+ |
| 17 gw | 0% | 0% | 0% | N/A | N/A | N/A | N/A | N/A | N/A | 0% | 0% | 0% |
| 31 gw | 0% | 0% | 0% | 0% | 0% | 0% | 12.9% | 14% | 17.9% | 0% | 0% | 0% |
| 36.6 gw | 0% | 0% | 0% | 0% | 0% | 0% | 0% | 0% | 0% | 0% | 0% | 0% |
| 37.4 gw | 0% | 0% | 0% | 0% | 0% | 0% | 0% | 0% | 0% | 0% | 0% | 0% |
| 38 gw | 0% | 0% | 0% | 0% | 0% | 0% | 0% | 0% | 0% | 0% | 0% | 0% |
| Postnatal | 0% | 0% | 0% | 0% | 0% | 0% | 0% | 0% | 0% | 0% | 0% | 0% |
| EGL | PCL | ML | Neuropil | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Staining Intensity | 1+ | 2+ | 3+ | 1+ | 2+ | 3+ | 1+ | 2+ | 3+ | 1+ | 2+ | 3+ |
| 17 gw | 0% | 0% | 0% | N/A | N/A | N/A | N/A | N/A | N/A | 8.2% | 20.1% | 4.9% |
| 31 gw | 4.4% | 9.7% | 18.4% | 0% | 0% | 0% | 29.1% | 4.1% | 0% | 24.7% | 6.8% | 1.6% |
| 36.6 gw | 0% | 0% | 0% | 22.5% | 4.7% | 0% | 0% | 0% | 0% | 0% | 0% | 0% |
| 37.4 gw | 0% | 0% | 0% | 0% | 25% | 8.3% | 0% | 0% | 0% | 0% | 0% | 0% |
| 38 gw | 0% | 0% | 0% | 0% | 0% | 33.3% | 0% | 0% | 0% | 0% | 0% | 0% |
| Postnatal | 0% | 0% | 0% | 30% | 3.3% | 0% | 0% | 0% | 0% | 0% | 0% | 0% |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Santana-Bejarano, M.B.; Grosso-Martínez, P.R.; Puebla-Mora, A.G.; Martínez-Silva, M.G.; Nava-Villalba, M.; Márquez-Aguirre, A.L.; Ortuño-Sahagún, D.; Godínez-Rubí, M. Pleiotrophin and the Expression of Its Receptors during Development of the Human Cerebellar Cortex. Cells 2023, 12, 1733. https://doi.org/10.3390/cells12131733
Santana-Bejarano MB, Grosso-Martínez PR, Puebla-Mora AG, Martínez-Silva MG, Nava-Villalba M, Márquez-Aguirre AL, Ortuño-Sahagún D, Godínez-Rubí M. Pleiotrophin and the Expression of Its Receptors during Development of the Human Cerebellar Cortex. Cells. 2023; 12(13):1733. https://doi.org/10.3390/cells12131733
Chicago/Turabian StyleSantana-Bejarano, Margarita Belem, Paula Romina Grosso-Martínez, Ana Graciela Puebla-Mora, María Guadalupe Martínez-Silva, Mario Nava-Villalba, Ana Laura Márquez-Aguirre, Daniel Ortuño-Sahagún, and Marisol Godínez-Rubí. 2023. "Pleiotrophin and the Expression of Its Receptors during Development of the Human Cerebellar Cortex" Cells 12, no. 13: 1733. https://doi.org/10.3390/cells12131733