Cytoskeletal Regulation of Dermal Regeneration

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Regeneration vs. Repair
3. Fingertip Regeneration
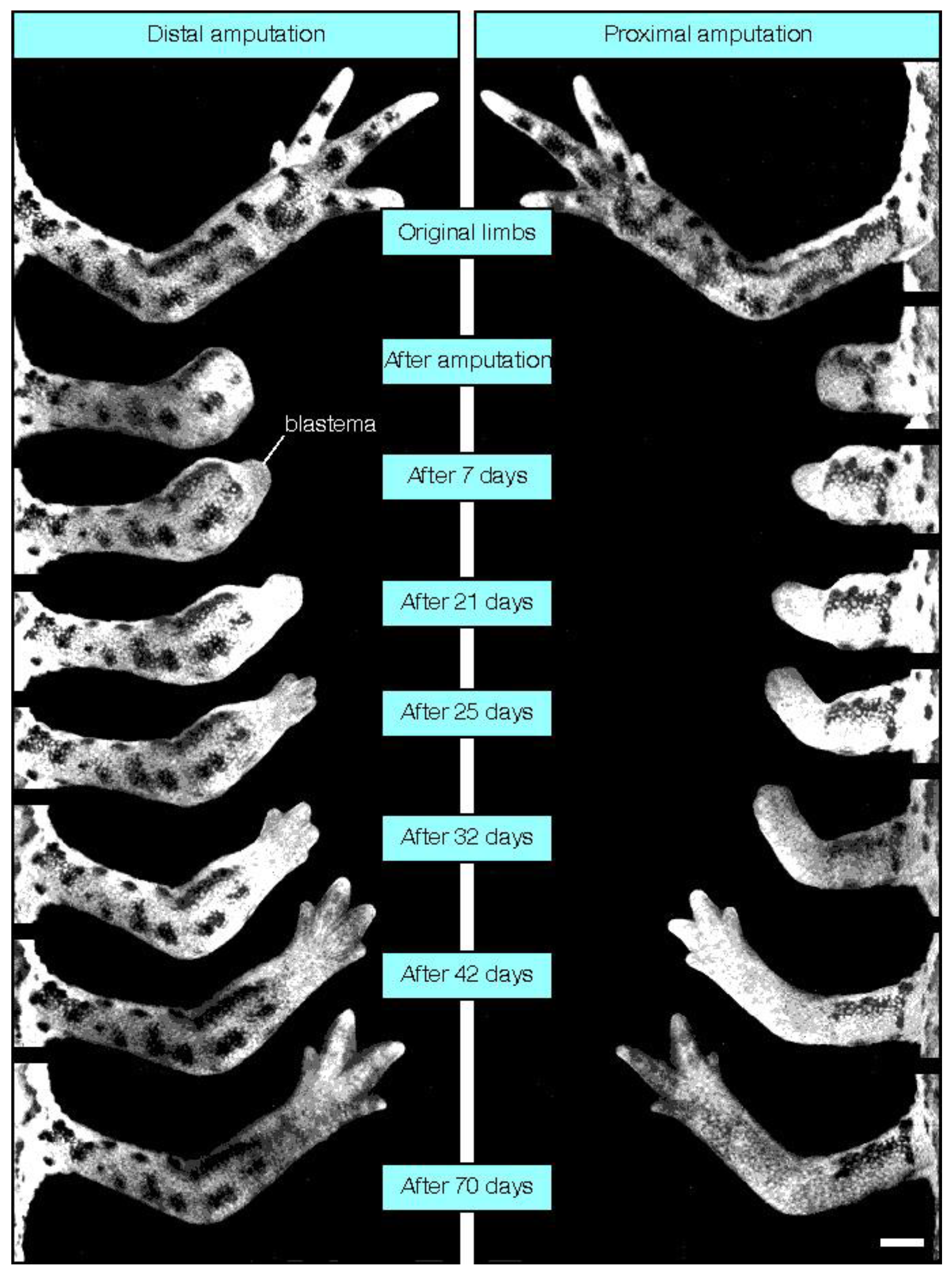

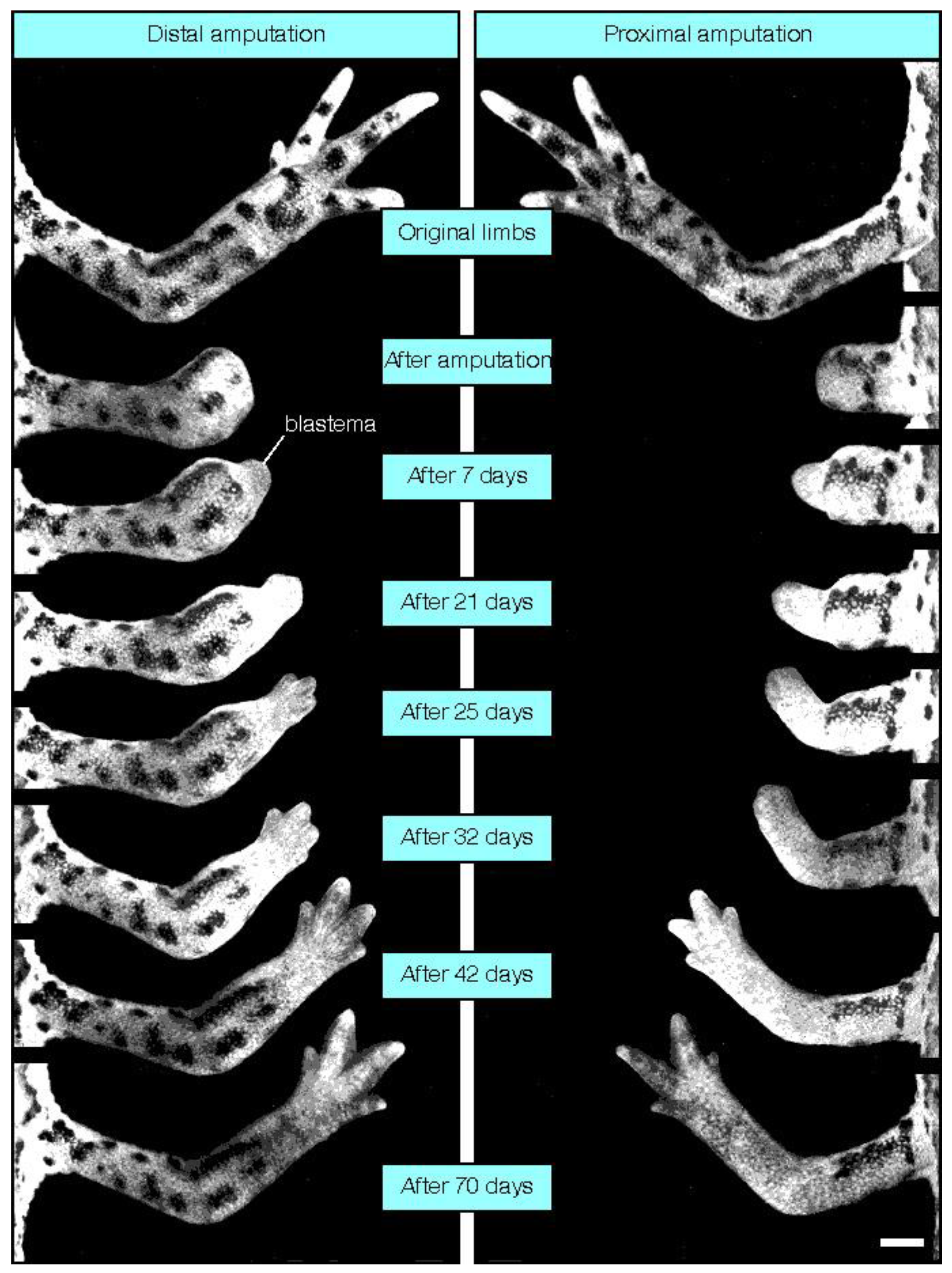
4. Mammalian Regenerative Capacity
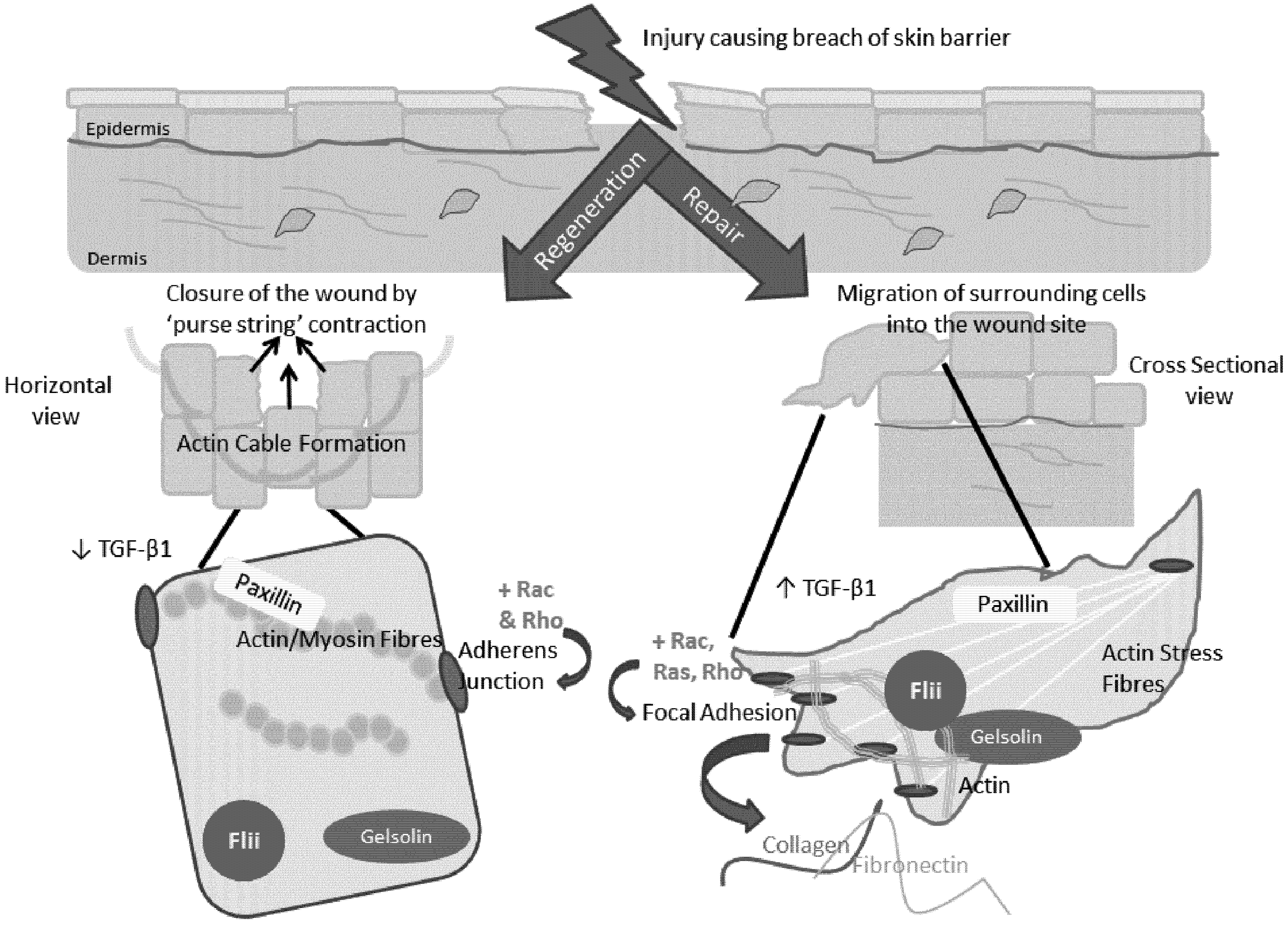
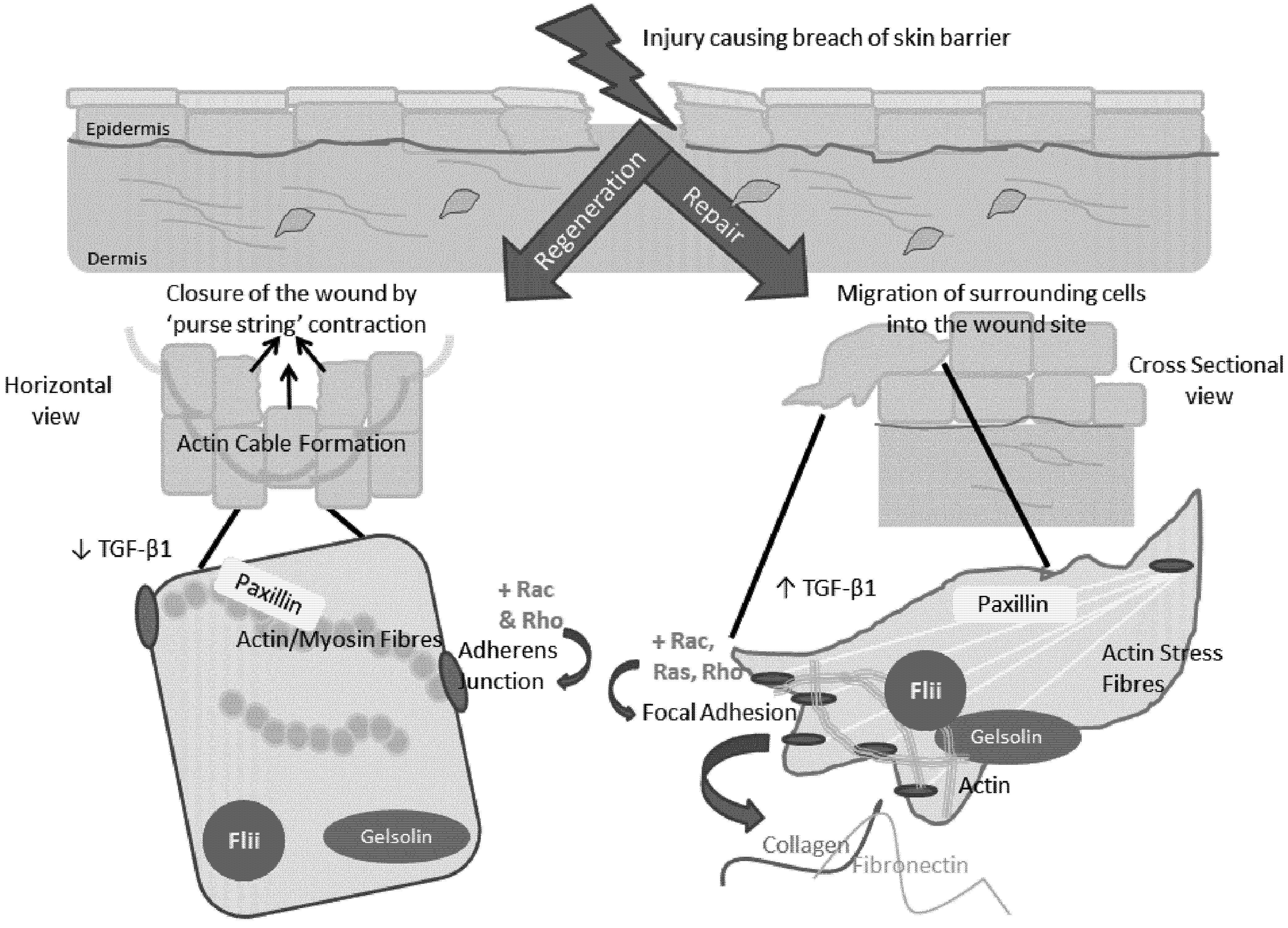
5. The Role of the Actin Cytoskeleton in Regeneration and Repair
6. Regenerative Fetal vs Reparative Adult Wound Healing

7. Involvement of the Actin Cytoskeleton during Regeneration and Repair
8. Flightless I in Wound Regeneration and Repair
9. Conclusion
Acknowledgements
References
- Morasso, M.I. The influence of flightless i: Regeneration versus wound healing. J. Invest. Dermatol. 2011, 131, 816–817. [Google Scholar] [CrossRef]
- Sanchez Alvarado, A. Regeneration in the metazoans: Why does it happen? Bioessays 2000, 22, 578–590. [Google Scholar] [CrossRef]
- Maiorana, V.C. Tail autonomy, functional conflicts and their resolution by a salamander. Nature 1997, 2265, 533–535. [Google Scholar]
- McKim, L.H. Regeneration of the distal phalanx. Can. Med. Assoc. J. 1932, 26, 549–550. [Google Scholar]
- Soderberg, T.; Nystrom, A.; Hallmans, G.; Hulten, J. Treatment of fingertip amputations with bone exposure. A comparative study between surgical and conservative treatment methods. Scand. J. Plast. Reconstr. Surg. Suppl. 1983, 17, 147–152. [Google Scholar] [CrossRef]
- Neufeld, D.A.; Zhao, W. Bone regrowth after digit tip amputation in mice is equivalent in adults and neonates. Wound Repair Regen. 1995, 3, 461–466. [Google Scholar]
- Han, M.; Yang, X.; Lee, J.; Allan, C.H.; Muneoka, K. Development and regeneration of the neonatal digit tip in mice. Dev. Biol. 2008, 315, 125–135. [Google Scholar] [CrossRef]
- Chadwick, R.B.; Bu, L.; Yu, H.; Hu, Y.; Wergedal, J.E.; Mohan, S.; Baylink, D.J. Digit tip regrowth and differential gene expression in mrl/mpj, dba/2, and c57bl/6 mice. Wound Repair Regen. 2007, 15, 275–284. [Google Scholar] [CrossRef]
- Singer, A.J.; Clark, R.A. Cutaneous wound healing. N. Engl. J. Med. 1999, 341, 738–746. [Google Scholar] [CrossRef]
- Yu, L.; Han, M.; Yan, M.; Lee, E.C.; Lee, J.; Muneoka, K. Bmp signaling induces digit regeneration in neonatal mice. Development 2010, 137, 551–559. [Google Scholar] [CrossRef]
- Fernando, W.A.; Leininger, E.; Simkin, J.; Li, N.; Malcom, C.A.; Sathyamoorthi, S.; Han, M.; Muneoka, K. Wound healing and blastema formation in regenerating digit tips of adult mice. Dev. Biol. 2011, 350, 301–310. [Google Scholar] [CrossRef]
- Cowin, A.J.; Adams, D.H.; Strudwick, X.L.; Chan, H.; Hooper, J.A.; Sander, G.R.; Rayner, T.E.; Matthaei, K.I.; Powell, B.C.; Campbell, H.D. Flightless i deficiency enhances wound repair by increasing cell migration and proliferation. J. Pathol. Bacteriol. 2007, 211, 572–581. [Google Scholar] [CrossRef]
- Lehoczky, J.A.; Robert, B.; Tabin, C.J. Mouse digit tip regeneration is mediated by fate-restricted progenitor cells. Proc. Natl. Acad. Sci. USA 2011, 108, 20609–20614. [Google Scholar]
- Rinkevich, Y.; Lindau, P.; Ueno, H.; Longaker, M.T.; Weissman, I.L. Germ-layer and lineage-restricted stem/progenitors regenerate the mouse digit tip. Nature 2011, 476, 409–413. [Google Scholar]
- Goss, R.J. Principles of regeneration; Academic Press: New York, NY, USA, 1969. [Google Scholar]
- Vidal, P.; Dickson, M.G. Regeneration of the distal phalanx. A case report. J. Hand. Surg. 1993, 18, 230–233. [Google Scholar]
- Muneoka, K.; Allan, C.H.; Yang, X.; Lee, J.; Han, M. Mammalian regeneration and regenerative medicine. Birth Defects Res. C Embryo Today 2008, 84, 265–280. [Google Scholar] [CrossRef]
- Jahoda, C.A.; Oliver, R.F.; Reynolds, A.J.; Forrester, J.C.; Horne, K.A. Human hair follicle regeneration following amputation and grafting into the nude mouse. J. Invest. Dermatol. 1996, 107, 804–807. [Google Scholar]
- Gat, U.; DasGupta, R.; Degenstein, L.; Fuchs, E. De novo hair follicle morphogenesis and hair tumors in mice expressing a truncated beta-catenin in skin. Cell 1998, 95, 605–614. [Google Scholar] [CrossRef]
- Fathke, C.; Wilson, L.; Shah, K.; Kim, B.; Hocking, A.; Moon, R.; Isik, F. Wnt signaling induces epithelial differentiation during cutaneous wound healing. BMC Cell Biol. 2006, 7, 4. [Google Scholar] [CrossRef]
- Michalopoulos, G.K.; DeFrances, M.C. Liver regeneration. Science 1997, 276, 60–66. [Google Scholar] [CrossRef]
- Thomas, D.W.; Harding, K.G. Wound healing. Br. J. Surg. 2002, 89, 1203–1205. [Google Scholar]
- Stephens, P.; Davies, K.J.; Occleston, N.; Pleass, R.D.; Kon, C.; Daniels, J.; Khaw, P.T.; Thomas, D.W. Skin and oral fibroblasts exhibit phenotypic differences in extracellular matrix reorganization and matrix metalloproteinase activity. Br. J. Dermatol. 2001, 144, 229–237. [Google Scholar] [CrossRef]
- Abreu-Blanco, M.T.; Watts, J.J.; Verboon, J.M.; Parkhurst, S.M. Cytoskeleton responses in wound repair. Cell. Mol. Life Sci. 2012, 2469–2483. [Google Scholar]
- Cowin, A. Role of the actin cytoskeleton in wound healing and scar formation. Primary Intention 2006, 14, 39–42. [Google Scholar]
- Desai, A.; Mitchison, T.J. Microtubule polymerization dynamics. Annu. Rev. Cell Dev. Biol. 1997, 13, 83–117. [Google Scholar] [CrossRef]
- Hall, A. Rho gtpases and the actin cytoskeleton. Science 1998, 279, 509–514. [Google Scholar] [CrossRef]
- Turner, C.E. Paxillin interactions. J. Cell. Sci. 2000, 113, 4139–4140. [Google Scholar]
- Martin, P. Wound healing--aiming for perfect skin regeneration. Science 1997, 276, 75–81. [Google Scholar] [CrossRef]
- Stelnicki, E.J.; Chin, G.S.; Gittes, G.K.; Longaker, M.T. Fetal wound repair: Where do we go from here? Semin. Pediatr. Surg. 1999, 8, 124–130. [Google Scholar]
- Cowin, A.J.; Hatzirodos, N.; Teusner, J.T.; Belford, D.A. Differential effect of wounding on actin and its associated proteins, paxillin and gelsolin, in fetal skin explants. J. Invest. Dermatol. 2003, 120, 1118–1129. [Google Scholar] [CrossRef]
- Hopkinson-Woolley, J.; Hughes, D.; Gordon, S.; Martin, P. Macrophage recruitment during limb development and wound healing in the embryonic and foetal mouse. J. Cell. Sci. 1994, 107, 1159–1167. [Google Scholar]
- Cowin, A.J.; Brosnan, M.P.; Holmes, T.M.; Ferguson, M.W. Endogenous inflammatory response to dermal wound healing in the fetal and adult mouse. Dev. Dyn. 1998, 212, 385–393. [Google Scholar] [CrossRef]
- Longaker, M.T.; Peled, Z.M.; Chang, J.; Krummel, T.M. Fetal wound healing: Progress report and future directions. Surgery 2001, 130, 785–787. [Google Scholar] [CrossRef]
- Cowin, A.J.; Hatzirodos, N.; Holding, C.A.; Dunaiski, V.; Harries, R.H.; Rayner, T.E.; Fitridge, R.; Cooter, R.D.; Schultz, G.S.; Belford, D.A. Effect of healing on the expression of transforming growth factor beta(s) and their receptors in chronic venous leg ulcers. J. Invest. Dermatol. 2001, 117, 1282–1289. [Google Scholar] [CrossRef]
- Ferguson, M.W. Skin wound healing: Transforming growth factor beta antagonists decrease scarring and improve quality. J. Interferon Res. 1994, 14, 303–304. [Google Scholar] [CrossRef]
- Jacinto, A.; Martinez-Arias, A.; Martin, P. Mechanisms of epithelial fusion and repair. Nat. Cell Biol. 2001, 3, E117–123. [Google Scholar] [CrossRef]
- Danjo, Y.; Gipson, I.K. Actin 'purse string' filaments are anchored by e-cadherin-mediated adherens junctions at the leading edge of the epithelial wound, providing coordinated cell movement. J. Cell. Sci. 1998, 111, 3323–3332. [Google Scholar]
- Belford, D.A. The mechanism of excisional fetal wound repair in vitro is responsive to growth factors. Endocrinology 1997, 138, 3987–3996. [Google Scholar] [CrossRef]
- Ihara, S.; Motobayashi, Y.; Nagao, E.; Kistler, A. Ontogenetic transition of wound healing pattern in rat skin occurring at the fetal stage. Development 1990, 110, 671–680. [Google Scholar]
- Amadeu, T.P.; Coulomb, B.; Desmouliere, A.; Costa, A.M. Cutaneous wound healing: Myofibroblastic differentiation and in vitro models. Int. J. Low. Extrem. Wounds 2003, 2, 60–68. [Google Scholar] [CrossRef]
- Dallon, J.; Sherratt, J.; Maini, P.; Ferguson, M. Biological implications of a discrete mathematical model for collagen deposition and alignment in dermal wound repair. IMA J. Math. Appl. Med. Biol. 2000, 17, 379–393. [Google Scholar] [Green Version]
- O'Kane, S. Wound remodelling and scarring. J Wound Care 2002, 11, 296–299. [Google Scholar] [Green Version]
- Silacci, P.; Mazzolai, L.; Gauci, C.; Stergiopulos, N.; Yin, H.L.; Hayoz, D. Gelsolin superfamily proteins: Key regulators of cellular functions. Cell. Mol. Life Sci. 2004, 61, 2614–2623. [Google Scholar] [CrossRef]
- Sun, H.Q.; Yamamoto, M.; Mejillano, M.; Yin, H.L. Gelsolin, a multifunctional actin regulatory protein. J. Biol. Chem. 1999, 274, 33179–33182. [Google Scholar] [CrossRef]
- Lee, W.M.; Galbraith, R.M. The extracellular actin-scavenger system and actin toxicity. N. Engl. J. Med. 1992, 326, 1335–1341. [Google Scholar] [CrossRef]
- Mounzer, K.C.; Moncure, M.; Smith, Y.R.; Dinubile, M.J. Relationship of admission plasma gelsolin levels to clinical outcomes in patients after major trauma. Am. J. Respir. Crit. Care Med. 1999, 160, 1673–1681. [Google Scholar]
- Ridley, A.J.; Hall, A. The small gtp-binding protein rho regulates the assembly of focal adhesions and actin stress fibers in response to growth factors. Cell 1992, 70, 389–399. [Google Scholar] [CrossRef]
- Gardel, M.L.; Schneider, I.C.; Aratyn-Schaus, Y.; Waterman, C.M. Mechanical integration of actin and adhesion dynamics in cell migration. Annu. Rev. Cell Dev. Biol. 2010, 26, 315–333. [Google Scholar] [CrossRef]
- Ridley, A.J.; Paterson, H.F.; Johnston, C.L.; Diekmann, D.; Hall, A. The small gtp-binding protein rac regulates growth factor-induced membrane ruffling. Cell 1992, 70, 401–410. [Google Scholar] [CrossRef]
- Nobes, C.D.; Hall, A. Rho, rac, and cdc42 gtpases regulate the assembly of multimolecular focal complexes associated with actin stress fibers, lamellipodia, and filopodia. Cell 1995, 81, 53–62. [Google Scholar] [CrossRef]
- Wozniak, M.A.; Modzelewska, K.; Kwong, L.; Keely, P.J. Focal adhesion regulation of cell behavior. Biochim. Biophys. Acta 2004, 1692, 103–119. [Google Scholar] [CrossRef]
- Millan, J.; Cain, R.J.; Reglero-Real, N.; Bigarella, C.; Marcos-Ramiro, B.; Fernandez-Martin, L.; Correas, I.; Ridley, A.J. Adherens junctions connect stress fibres between adjacent endothelial cells. BMC Biol. 2010, 8, 11. [Google Scholar]
- Liu, Y.T.; Yin, H.L. Identification of the binding partners for flightless i, a novel protein bridging the leucine-rich repeat and the gelsolin superfamilies. J. Biol. Chem. 1998, 273, 7920–7927. [Google Scholar] [CrossRef]
- Claudianos, C.; Campbell, H.D. The novel flightless-i gene brings together two gene families, actin-binding proteins related to gelsolin and leucine-rich-repeat proteins involved in ras signal transduction. Mol. Biol. Evol. 1995, 12, 405–414. [Google Scholar] [Green Version]
- Campbell, H.D.; Fountain, S.; McLennan, I.S.; Berven, L.A.; Crouch, M.F.; Davy, D.A.; Hooper, J.A.; Waterford, K.; Chen, K.S.; Lupski, J.R.; et al. Fliih, a gelsolin-related cytoskeletal regulator essential for early mammalian embryonic development. Mol. Cell. Biol. 2002, 22, 3518–3526. [Google Scholar] [CrossRef]
- Davy, D.A.; Ball, E.E.; Matthaei, K.I.; Campbell, H.D.; Crouch, M.F. The flightless i protein localizes to actin-based structures during embryonic development. Immunol. Cell Biol. 2000, 78, 423–429. [Google Scholar] [CrossRef]
- Lin, C.H.; Waters, J.M.; Powell, B.C.; Arkell, R.M.; Cowin, A.J. Decreased expression of flightless i, a gelsolin family member and developmental regulator, in early-gestation fetal wounds improves healing. Mamm. Genome 2011, 22, 341–352. [Google Scholar] [CrossRef]
- Jackson, J.E.; Kopecki, Z.; Adams, D.H.; Cowin, A.J. Flii neutralizing antibodies improve wound healing in porcine preclinical studies. Wound Repair Regen. 2012, 20, 523–536. [Google Scholar]
- Davy, D.A.; Campbell, H.D.; Fountain, S.; de Jong, D.; Crouch, M.F. The flightless i protein colocalizes with actin- and microtubule-based structures in motile swiss 3t3 fibroblasts: Evidence for the involvement of pi 3-kinase and ras-related small gtpases. J. Cell Sci. 2001, 114, 549–562. [Google Scholar]
- Higashi, T.; Ikeda, T.; Murakami, T.; Shirakawa, R.; Kawato, M.; Okawa, K.; Furuse, M.; Kimura, T.; Kita, T.; Horiuchi, H. Flightless-i (fli-i) regulates the actin assembly activity of diaphanous-related formins (drfs) daam1 and mdia1 in cooperation with active rho gtpase. J. Biol. Chem. 2010, 285, 16231–16238. [Google Scholar]
- Begum, R.; Nur, E.K.M.S.; Zaman, M.A. The role of rho gtpases in the regulation of the rearrangement of actin cytoskeleton and cell movement. Exp. Mol. Med. 2004, 36, 358–366. [Google Scholar]
- Bhowmick, N.A.; Ghiassi, M.; Bakin, A.; Aakre, M.; Lundquist, C.A.; Engel, M.E.; Arteaga, C.L.; Moses, H.L. Transforming growth factor-beta1 mediates epithelial to mesenchymal transdifferentiation through a rhoa-dependent mechanism. Mol. Biol. Cell 2001, 12, 27–36. [Google Scholar]
- Hall, A.; Nobes, C.D. Rho gtpases: Molecular switches that control the organization and dynamics of the actin cytoskeleton. Philos. Trans. R Soc. Lond B Biol. Sci. 2000, 355, 965–970. [Google Scholar] [CrossRef]
- Wang, W.; Koka, V.; Lan, H.Y. Transforming growth factor-beta and smad signalling in kidney diseases. Nephrology 2005, 10, 48–56. [Google Scholar] [CrossRef]
- Kopecki, Z.; Arkell, R.; Powell, B.C.; Cowin, A.J. Flightless i regulates hemidesmosome formation and integrin-mediated cellular adhesion and migration during wound repair. J. Invest. Dermatol. 2009, 129, 2031–2045. [Google Scholar] [CrossRef]
- Ben-Ze'ev, A. Cytoskeletal and adhesion proteins as tumor suppressors. Curr. Opin. Cell. Biol. 1997, 9, 99–108. [Google Scholar] [CrossRef]
- Kaibuchi, K.; Kuroda, S.; Amano, M. Regulation of the cytoskeleton and cell adhesion by the rho family gtpases in mammalian cells. Annu. Rev. Biochem. 1999, 68, 459–486. [Google Scholar] [CrossRef]
- Wilkes, M.C.; Mitchell, H.; Penheiter, S.G.; Dore, J.J.; Suzuki, K.; Edens, M.; Sharma, D.K.; Pagano, R.E.; Leof, E.B. Transforming growth factor-beta activation of phosphatidylinositol 3-kinase is independent of smad2 and smad3 and regulates fibroblast responses via p21-activated kinase-2. Cancer Res. 2005, 65, 10431–10440. [Google Scholar]
- Shah, M.; Foreman, D.M.; Ferguson, M.W. Neutralising antibody to tgf-beta 1,2 reduces cutaneous scarring in adult rodents. J. Cell. Sci. 1994, 107, 1137–1157. [Google Scholar]
- Waters, J.M.; Lindo, J.E.; Arkell, R.M.; Cowin, A.J. Regeneration of hair follicles is modulated by flightless i (flii) in a rodent vibrissa model. J. Invest. Dermatol. 2011. [Google Scholar]
- Lee, Y.H.; Stallcup, M.R. Interplay of fli-i and flap1 for regulation of beta-catenin dependent transcription. Nucleic. Acids Res. 2006, 34, 5052–5059. [Google Scholar] [CrossRef]
- Archer, S.K.; Behm, C.A.; Claudianos, C.; Campbell, H.D. The flightless i protein and the gelsolin family in nuclear hormone receptor-mediated signalling. Biochem. Soc. Trans. 2004, 32, 940–942. [Google Scholar] [CrossRef]
- Lee, Y.H.; Campbell, H.D.; Stallcup, M.R. Developmentally essential protein flightless i is a nuclear receptor coactivator with actin binding activity. Mol. Cell. Biol. 2004, 24, 2103–2117. [Google Scholar] [CrossRef]
- Fong, K.S.; de Couet, H.G. Novel proteins interacting with the leucine-rich repeat domain of human flightless-i identified by the yeast two-hybrid system. Genomics 1999, 58, 146–157. [Google Scholar] [CrossRef]
- Seward, M.E.; Easley, C.A.T.; McLeod, J.J.; Myers, A.L.; Tombes, R.M. Flightless-i, a gelsolin family member and transcriptional regulator, preferentially binds directly to activated cytosolic camk-ii. FEBS Lett. 2008, 582, 2489–2495. [Google Scholar] [CrossRef]
- Lei, N.; Franken, L.; Ruzehaji, N.; Offenhauser, C.; Cowin, A.J.; Murray, R.Z. Flightless, secreted through a late endosome/lysosome pathway, binds lps and dampens cytokine secretion. J. Cell. Sci. 2012. [Google Scholar]
- Stallcup, M.R.; Kim, J.H.; Teyssier, C.; Lee, Y.H.; Ma, H.; Chen, D. The roles of protein-protein interactions and protein methylation in transcriptional activation by nuclear receptors and their coactivators. J. Steroid Biochem. Mol. Biol. 2003, 85, 139–145. [Google Scholar] [CrossRef]
- Yang, P.; An, H.; Liu, X.; Wen, M.; Zheng, Y.; Rui, Y.; Cao, X. The cytosolic nucleic acid sensor lrrfip1 mediates the production of type i interferon via a beta-catenin-dependent pathway. Nat. Immunol. 2010, 11, 487–494. [Google Scholar]
- Liu, J.; Bang, A.G.; Kintner, C.; Orth, A.P.; Chanda, S.K.; Ding, S.; Schultz, P.G. Identification of the wnt signaling activator leucine-rich repeat in flightless interaction protein 2 by a genome-wide functional analysis. Proc. Natl. Acad. Sci. USA 2005, 102, 1927–1932. [Google Scholar]
- Carre, A.L.; James, A.W.; MacLeod, L.; Kong, W.; Kawai, K.; Longaker, M.T.; Lorenz, H.P. Interaction of wingless protein (wnt), transforming growth factor-beta1, and hyaluronan production in fetal and postnatal fibroblasts. Plast. Reconstr. Surg. 2010, 125, 74–88. [Google Scholar]
© 2012 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Strudwick, X.L.; Cowin, A.J. Cytoskeletal Regulation of Dermal Regeneration. Cells 2012, 1, 1313-1327. https://doi.org/10.3390/cells1041313
Strudwick XL, Cowin AJ. Cytoskeletal Regulation of Dermal Regeneration. Cells. 2012; 1(4):1313-1327. https://doi.org/10.3390/cells1041313
Chicago/Turabian StyleStrudwick, Xanthe L., and Allison J. Cowin. 2012. "Cytoskeletal Regulation of Dermal Regeneration" Cells 1, no. 4: 1313-1327. https://doi.org/10.3390/cells1041313
APA StyleStrudwick, X. L., & Cowin, A. J. (2012). Cytoskeletal Regulation of Dermal Regeneration. Cells, 1(4), 1313-1327. https://doi.org/10.3390/cells1041313