Simultaneous Detection of Antigen-Specific IgG- and IgM-Secreting Cells with a B Cell Fluorospot Assay

Abstract
:1. Introduction
2. Results and Discussion
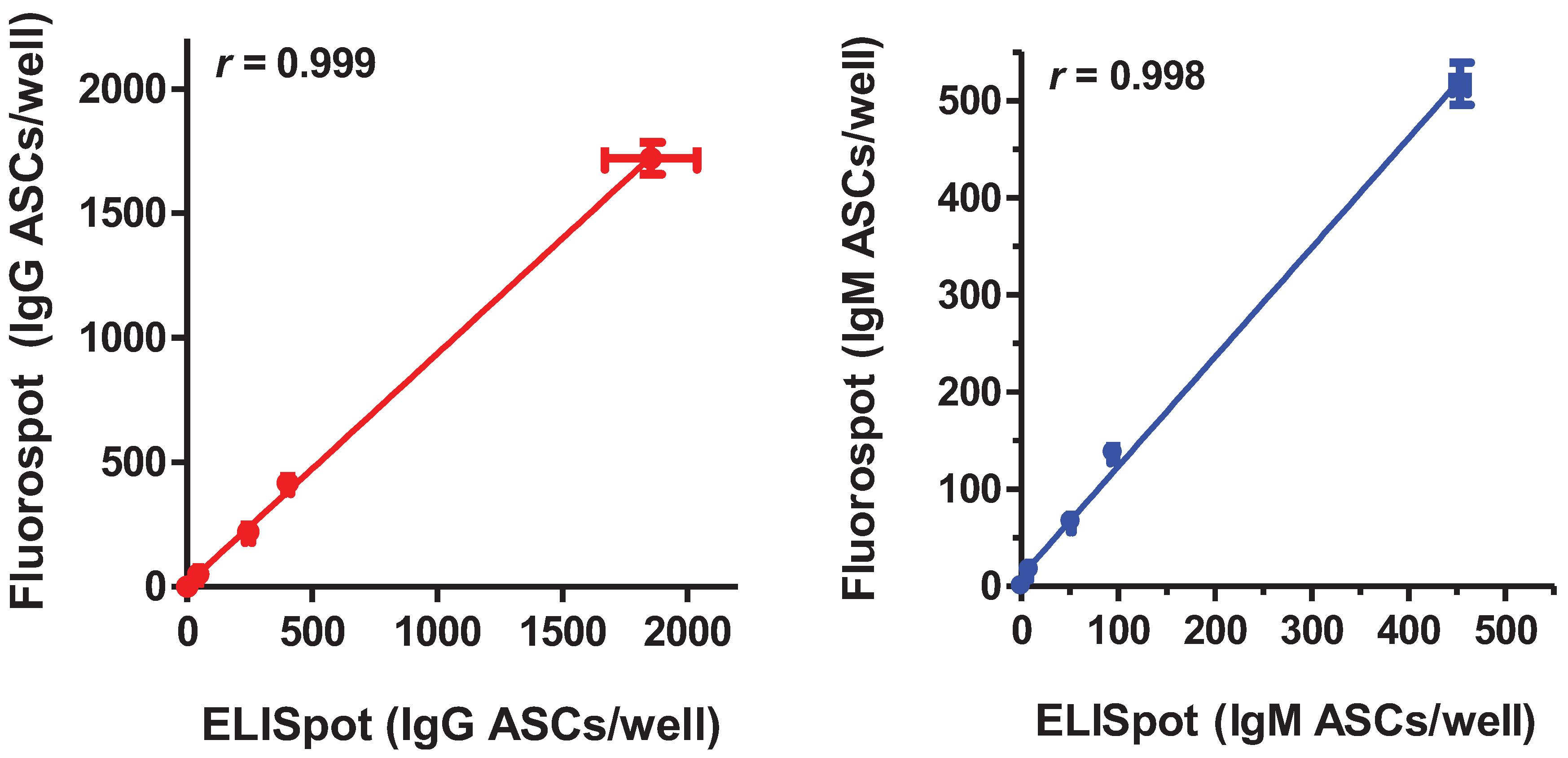
2.1. Comparison of the Sensitivity and Linear Range of Detection of the B cell Fluorospot Assay and the Traditional ELISpot Assay in Enumerating IgG and IgM ASCs

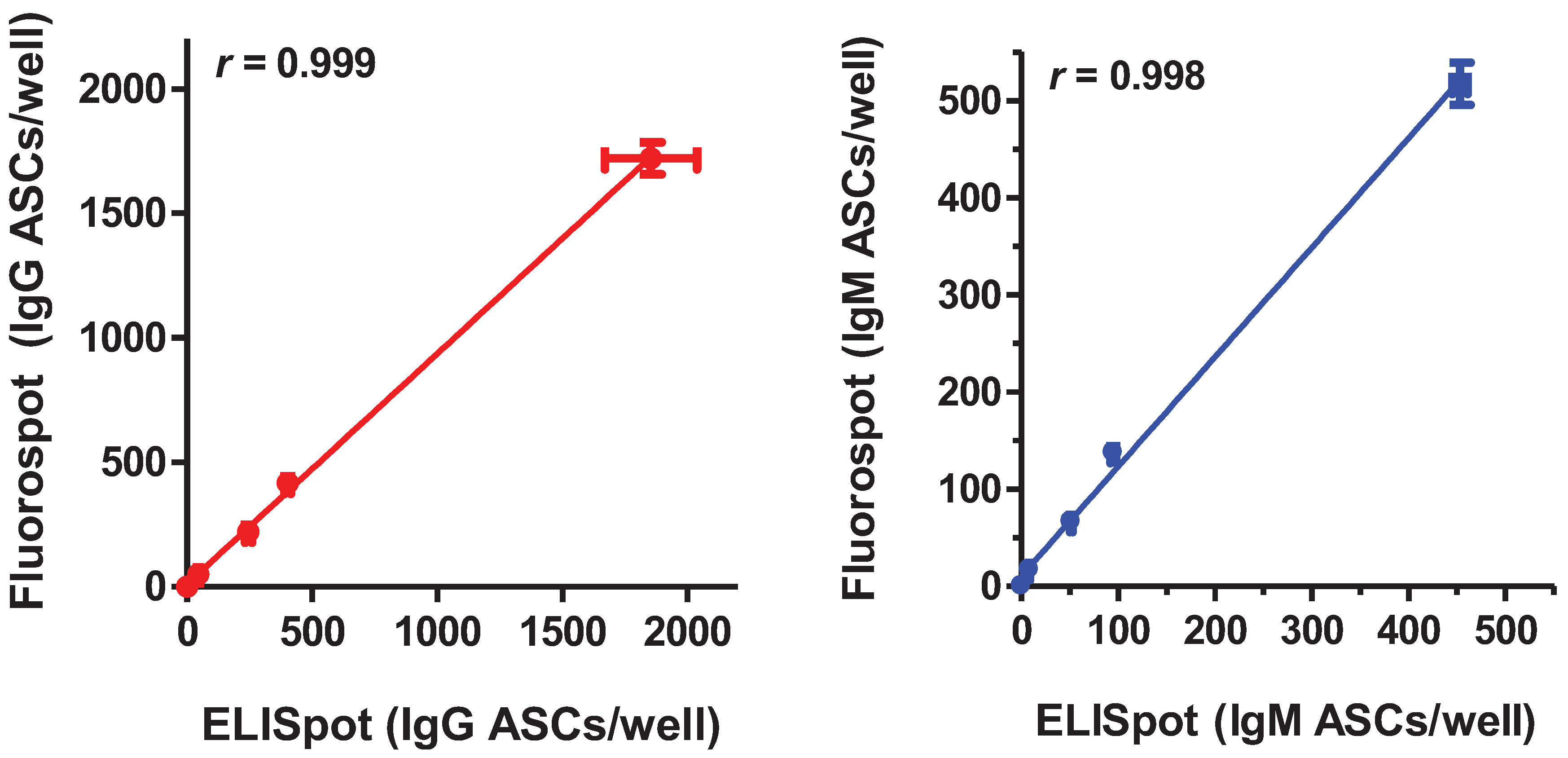{kind=link}
{kind=link}
{kind=link}
| cells/well | 7B2-IgG | |||
|---|---|---|---|---|
| ELISpot | Fluorospot | |||
| ASC/wella | spot:cell ratio | ASC/well a | spot:cell ratio | |
| 0 | 0 | n/a | 0 | n/a |
| 100 | 52 ± 9 | 0.52 | 50 ± 11 | 0.50 |
| 500 | 232 ± 19 | 0.46 | 222 ± 18 | 0.44 |
| 1,000 | 416 ± 18 | 0.42 | 417 ± 12 | 0.42 |
| 5,000 | 1,669 ± 260 | 0.33 | 1,721 ± 64 | 0.34 |
| Linearity of the range of detection | ||||
| r | 0.99<0.0001 | 0.998<0.0001 | ||
| p | ||||
| cells/well | 2B9-IgM | |||
|---|---|---|---|---|
| ELISpot | Fluorospot | |||
| ASC/well a | spot:cell ratio | ASC/well a | spot:cell ratio | |
| 0 | 0 | n/a | 0 | n/a |
| 100 | 9 ± 1 | 0.09 | 17 ± 0 | 0.17 |
| 500 | 52 ± 2 | 0.10 | 67 ± 1 | 0.13 |
| 1,000 | 95 ± 4 | 0.10 | 138 ± 8 | 0.14 |
| 5,000 | 453 ± 11 | 0.09 | 518 ± 22 | 0.10 |
| Linearity of the range of detection | ||||
| r | 0.999<0.0001 | 0.997<0.0001 | ||
| p | ||||
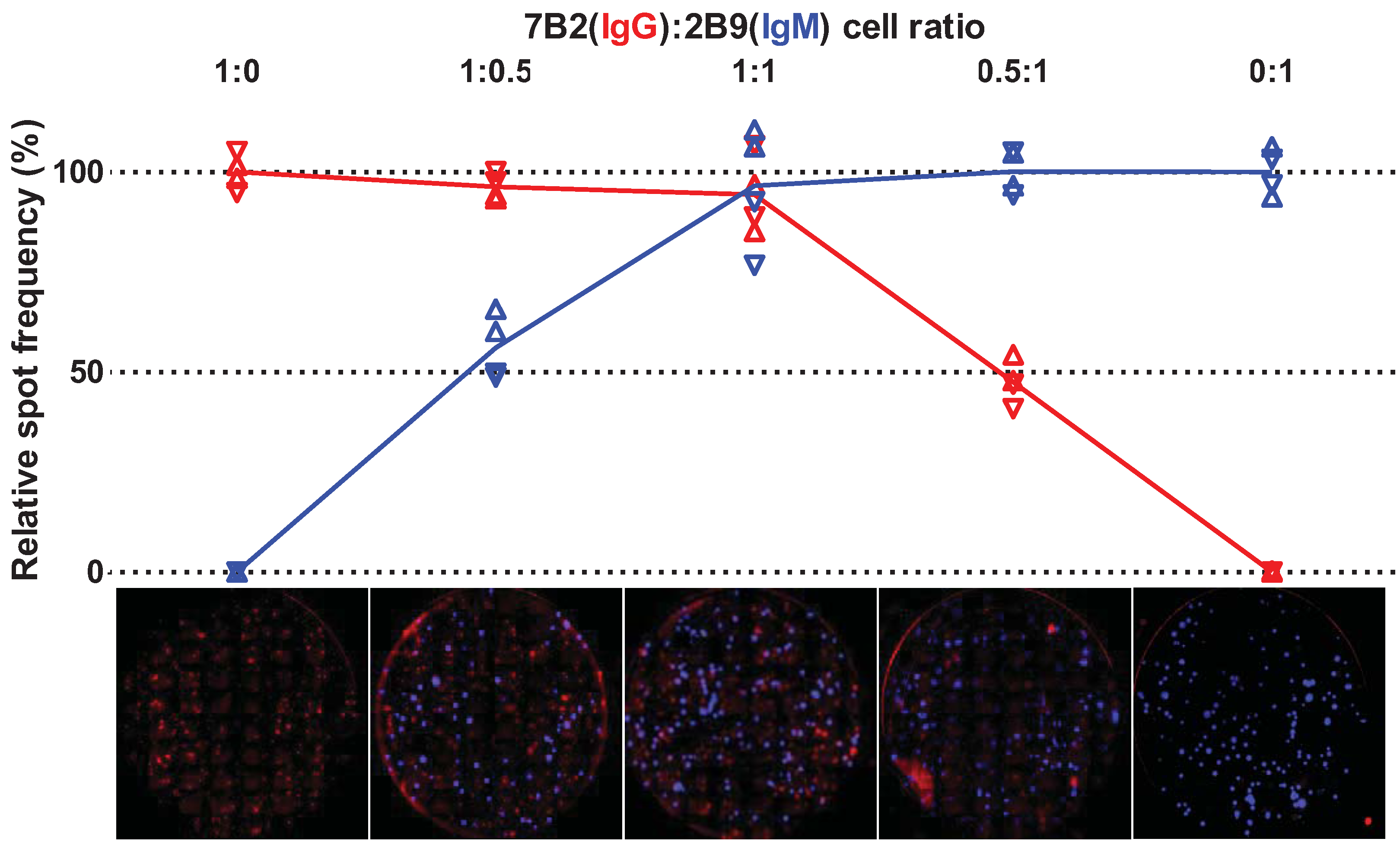
2.2. Multiplex Detection of IgG and IgM Antibody-Secreting Cells
| Condition | cells/well | IgG ASCs | IgM ASCs | |||||||
|---|---|---|---|---|---|---|---|---|---|---|
| IgG-7B2 | IgM-2B9 | Well 1 | Well 2 | Average | Well 1 | Well 2 | Average | |||
| Experiment 1 | ||||||||||
| 1:0 | 1,000 | 0 | 955 | 987 | 971 | 0 | 0 | 0 | ||
| 1:0.5 | 1,000 | 500 | 915 | 906 | 911 | 43 | 47 | 45 | ||
| 1:1 | 1,000 | 1,000 | 827 | 940 | 884 | 79 | 77 | 78 | ||
| 0.5:1 | 500 | 1,000 | 465 | 527 | 496 | 69 | 75 | 72 | ||
| 0:1 | 0 | 1,000 | 0 | 0 | 0 | 76 | 67 | 72 | ||
| Experiment 2 | ||||||||||
| 1:0 | 500 | 0 | 360 | 325 | 343 | 0 | 0 | 0 | ||
| 1:0.5 | 500 | 1,000 | 343 | 334 | 339 | 94 | 92 | 93 | ||
| 1:1 | 500 | 2,000 | 304 | 365 | 335 | 175 | 145 | 160 | ||
| 0.5:1 | 250 | 2,000 | 161 | 140 | 151 | 178 | 198 | 188 | ||
| 0:1 | 0 | 2,000 | 0 | 0 | 0 | 195 | 183 | 189 | ||
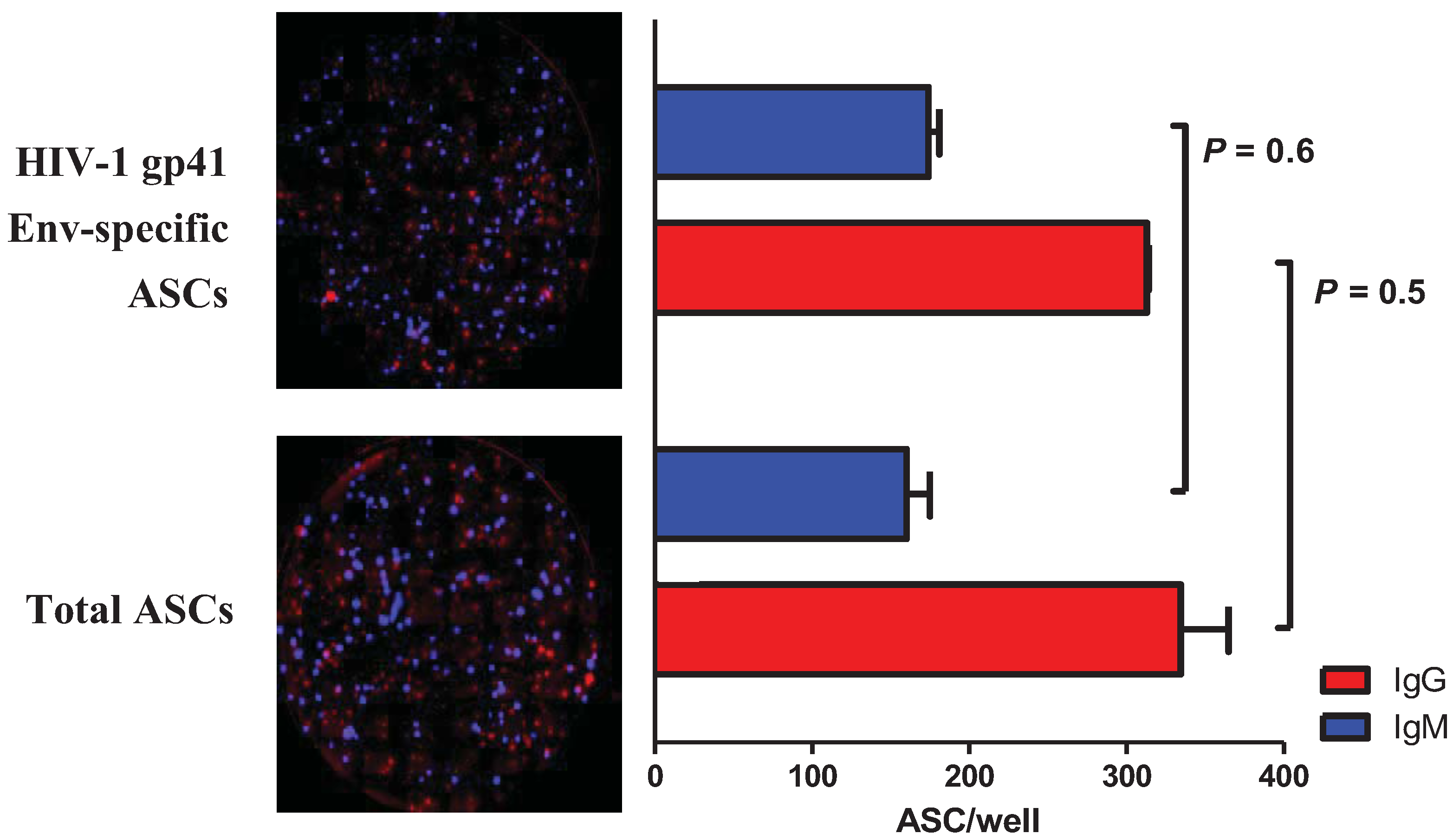
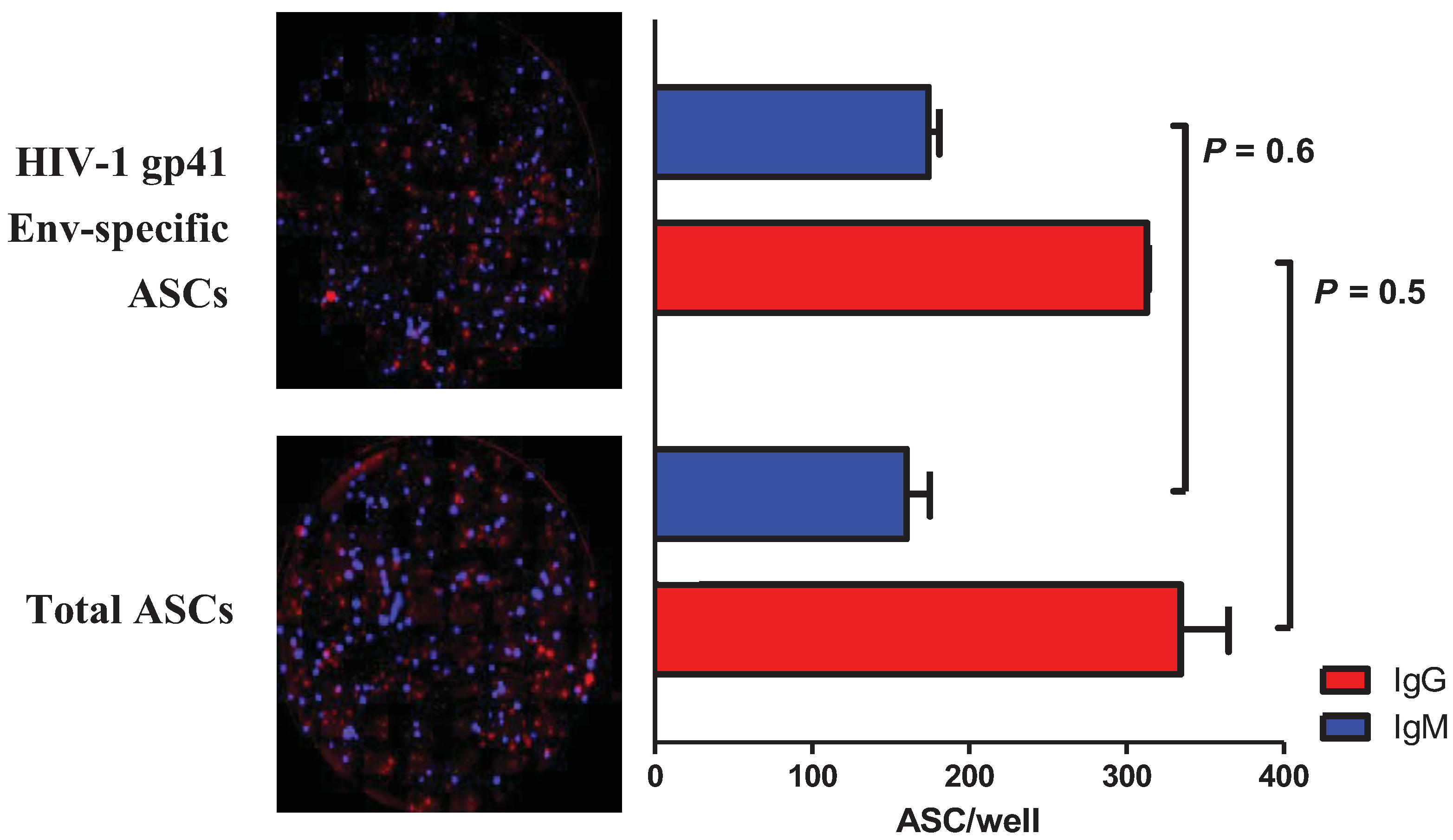
2.3. Multiplex Detection of HIV-1 gp41 Env-Specific ASCs
2.4. Discussion
3. Experimental Section
3.1. Cell Lines
3.2. Conjugation of Anti-Human IgG and IgM to Quantum-Dot Nanocrystals
3.3. B Cell Fluorospot Assay
3.4. Acquisition of Fluorescent Images
3.5. Traditional ELISpot Assay
4. Conclusions
Acknowledgments
Conflict of Interest
References
- Sedgwick, J.D.; Holt, P.G. A solid-phase immunoenzymatic technique for the enumeration of specific antibody-secreting cells. J. Immunol. Methods 1983, 57, 301–309. [Google Scholar] [CrossRef]
- Czerkinsky, C.C.; Nilsson, L.A.; Nygren, H.; Ouchterlony, O.; Tarkowski, A. A solid-phase enzyme-linked immunospot (ELISPOT) assay for enumeration of specific antibody-secreting cells. J. Immunol. Methods 1983, 65, 109–121. [Google Scholar] [CrossRef]
- Crotty, S.; Aubert, R.D.; Glidewell, J.; Ahmed, R. Tracking human antigen-specific memory B cells: A sensitive and generalized ELISPOT system. J. Immunol. Methods 2004, 286, 111–122. [Google Scholar] [CrossRef]
- Tuaillon, E.; Tabaa, Y.A.; Petitjean, G.; Huguet, M.F.; Pajeaux, G.; Fondere, J.M.; Ponseille, B.; Ducos, J.; Blanc, P.; Vendrell, J.P. Detection of memory B lymphocytes specific to hepatitis B virus (HBV) surface antigen (HBsAg) from HBsAg-vaccinated or HBV-immunized subjects by ELISPOT assay. J. Immunol. Methods 2006, 315, 144–152. [Google Scholar] [CrossRef]
- Bonsignori, M.; Moody, M.A.; Parks, R.J.; Holl, T.M.; Kelsoe, G.; Hicks, C.B.; Vandergrift, N.; Tomaras, G.D.; Haynes, B.F. HIV-1 envelope induces memory B cell responses that correlate with plasma antibody levels after envelope gp120 protein vaccination or HIV-1 infection. J. Immunol. 2009, 183, 2708–2717. [Google Scholar] [CrossRef]
- Malyguine, A.; Strobl, S.; Zaritskaya, L.; Baseler, M.; Shafer-Weaver, K. New approaches for monitoring CTL activity in clinical trials. Adv. Exp. Med. Biol. 2007, 601, 273–284. [Google Scholar] [CrossRef]
- Sachdeva, N.; Asthana, D. Cytokine quantitation: Technologies and applications. Front. Biosci. 2007, 12, 4682–4695. [Google Scholar] [CrossRef]
- Vasan, S.; Schlesinger, S.J.; Arrode, G. T cell immune responses to HIV-1. Front. Biosci. 2007, 12, 2330–2343. [Google Scholar] [CrossRef]
- Bromage, E.; Stephens, R.; Hassoun, L. The third dimension of ELISPOTs: Quantifying antibody secretion from individual plasma cells. J. Immunol. Methods 2009, 346, 75–79. [Google Scholar] [CrossRef]
- Gazagne, A.; Malkusch, W.; Vingert, B.; Fridman, W.H.; Tartour, E. Fluorospot assay: Methodological analysis. Methods Mol. Biol. 2005, 302, 289–296. [Google Scholar]
- Ahlborg, N.; Axelsson, B. Dual- and triple-color fluorospot. Methods Mol. Biol. 2012, 792, 77–85. [Google Scholar] [CrossRef]
- Rebhahn, J.A.; Bishop, C.; Divekar, A.A.; Jiminez-Garcia, K.; Kobie, J.J.; Lee, F.E.; Maupin, G.M.; Snyder-Cappione, J.E.; Zaiss, D.M.; Mosmann, T.R. Automated analysis of two- and three-color fluorescent Elispot (Fluorospot) assays for cytokine secretion. Comput. Methods Programs Biomed. 2008, 92, 54–65. [Google Scholar] [CrossRef]
- Czerkinsky, C.; Moldoveanu, Z.; Mestecky, J.; Nilsson, L.A.; Ouchterlony, O. A novel two colour ELISPOT assay. I. Simultaneous detection of distinct types of antibody-secreting cells. J. Immunol. Methods 1988, 115, 31–37. [Google Scholar] [CrossRef]
- Cordoba, F.; Lavabre-Bertrand, T.; Salhi, S.L.; Huguet, M.F.; Gerfaux, J.; Rossi, J.F.; Vendrell, J.P. Spontaneous monoclonal immunoglobulin-secreting peripheral blood mononuclear cells as a marker of disease severity in multiple myeloma. Br. J. Haematol. 2000, 108, 549–558. [Google Scholar] [CrossRef]
- Fondere, J.M.; Huguet, M.F.; Macura-Biegun, A.; Baillat, V.; Ohayon, V.; Reynes, J.; Vendrell, J.P. Detection and enumeration of circulating HIV-1-specific memory B cells in HIV-1-infected patients. J. Acquir. Immune Defic. Syndr. 2004, 35, 114–119. [Google Scholar] [CrossRef]
- Okamoto, Y.; Abe, T.; Niwa, T.; Mizuhashi, S.; Nishida, M. Development of a dual color enzyme-linked immunospot assay for simultaneous detection of murine T helper type 1- and T helper type 2-cells. Immunopharmacology 1998, 39, 107–116. [Google Scholar] [CrossRef]
- Okamoto, Y.; Gotoh, Y.; Shiraishi, H.; Nishida, M. A human dual-color enzyme-linked immunospot assay for simultaneous detection of interleukin 2- and interleukin 4-secreting cells. Int. Immunopharmacol. 2004, 4, 149–156. [Google Scholar] [CrossRef]
- Okamoto, Y.; Nishida, M. Dual-color ELISPOT assay for analyzing cytokine balance. Methods Mol. Biol. 2005, 302, 263–272. [Google Scholar]
- Boulet, S.; Ndongala, M.L.; Peretz, Y.; Boisvert, M.P.; Boulassel, M.R.; Tremblay, C.; Routy, J.P.; Sekaly, R.P.; Bernard, N.F. A dual color ELISPOT method for the simultaneous detection of IL-2 and IFN-gamma HIV-specific immune responses. J. Immunol. Methods 2007, 320, 18–29. [Google Scholar] [CrossRef]
- Quast, S.; Zhang, W.; Shive, C.; Kovalovski, D.; Ott, P.A.; Herzog, B.A.; Boehm, B.O.; Tary-Lehmann, M.; Karulin, A.Y.; Lehmann, P.V. IL-2 absorption affects IFN-gamma and IL-5, but not IL-4 producing memory T cells in double color cytokine ELISPOT assays. Cell. Immunol. 2005, 237, 28–36. [Google Scholar] [CrossRef]
- Snyder, J.E.; Bowers, W.J.; Livingstone, A.M.; Lee, F.E.; Federoff, H.J.; Mosmann, T.R. Measuring the frequency of mouse and human cytotoxic T cells by the Lysispot assay: Independent regulation of cytokine secretion and short-term killing. Nat. Med. 2003, 9, 231–235. [Google Scholar] [CrossRef]
- Karulin, A.Y.; Hesse, M.D.; Tary-Lehmann, M.; Lehmann, P.V. Single-cytokine-producing CD4 memory cells predominate in type 1 and type 2 immunity. J. Immunol. 2000, 164, 1862–1872. [Google Scholar]
- Gazagne, A.; Claret, E.; Wijdenes, J.; Yssel, H.; Bousquet, F.; Levy, E.; Vielh, P.; Scotte, F.; Goupil, T.L.; Fridman, W.H.; et al. A Fluorospot assay to detect single T lymphocytes simultaneously producing multiple cytokines. J. Immunol. Methods 2003, 283, 91–98. [Google Scholar] [CrossRef]
- Cromey, D.W. Avoiding twisted pixels: Ethical guidelines for the appropriate use and manipulation of scientific digital images. Sci. Eng. Ethics 2010, 16, 639–667. [Google Scholar] [CrossRef]
- Homepage of ImageJ. Available online: http://imagej.nih.gov/ij (accessed on 15 March 2012).
- Abramoff, M.D.; Magalhaes, P.J.; Ram, S.J. Image processing with image. J. Biophotonics Int. 2004, 11, 36–42. [Google Scholar]
© 2012 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Bonsignori, M.; Moody, M.A. Simultaneous Detection of Antigen-Specific IgG- and IgM-Secreting Cells with a B Cell Fluorospot Assay. Cells 2012, 1, 15-26. https://doi.org/10.3390/cells1010015
Bonsignori M, Moody MA. Simultaneous Detection of Antigen-Specific IgG- and IgM-Secreting Cells with a B Cell Fluorospot Assay. Cells. 2012; 1(1):15-26. https://doi.org/10.3390/cells1010015
Chicago/Turabian StyleBonsignori, Mattia, and M. Anthony Moody. 2012. "Simultaneous Detection of Antigen-Specific IgG- and IgM-Secreting Cells with a B Cell Fluorospot Assay" Cells 1, no. 1: 15-26. https://doi.org/10.3390/cells1010015