Forage Yield and Quality Indices of Silage-Corn Following Organic and Inorganic Phosphorus Amendments in Podzol Soil under Boreal Climate


 , ,
, , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Location and Treatments
2.2. Crop Management and Forage Sampling
2.3. Forage Quality Analysis
2.4. Statistical Analyses
3. Results
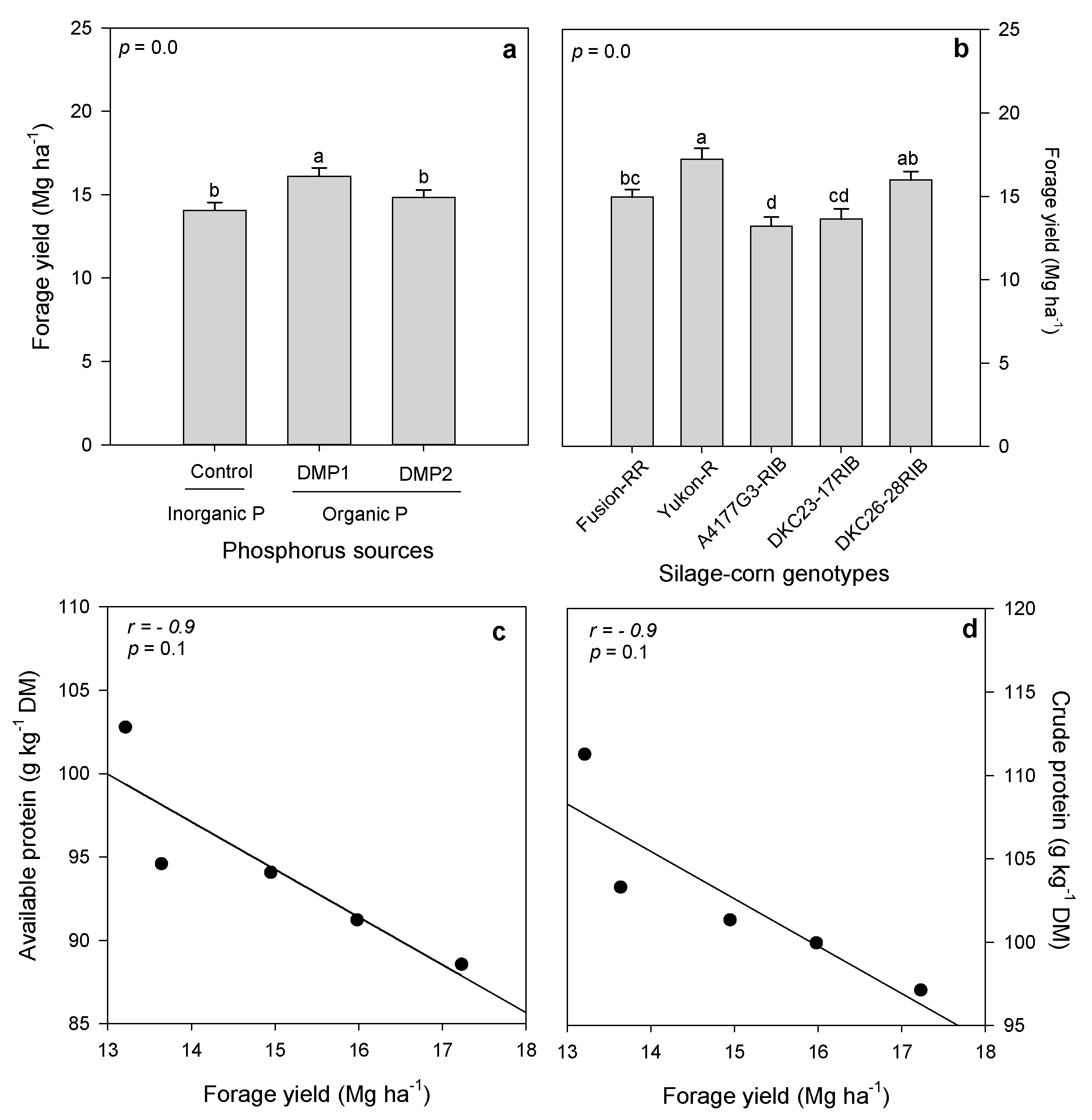
3.1. Forage Yield
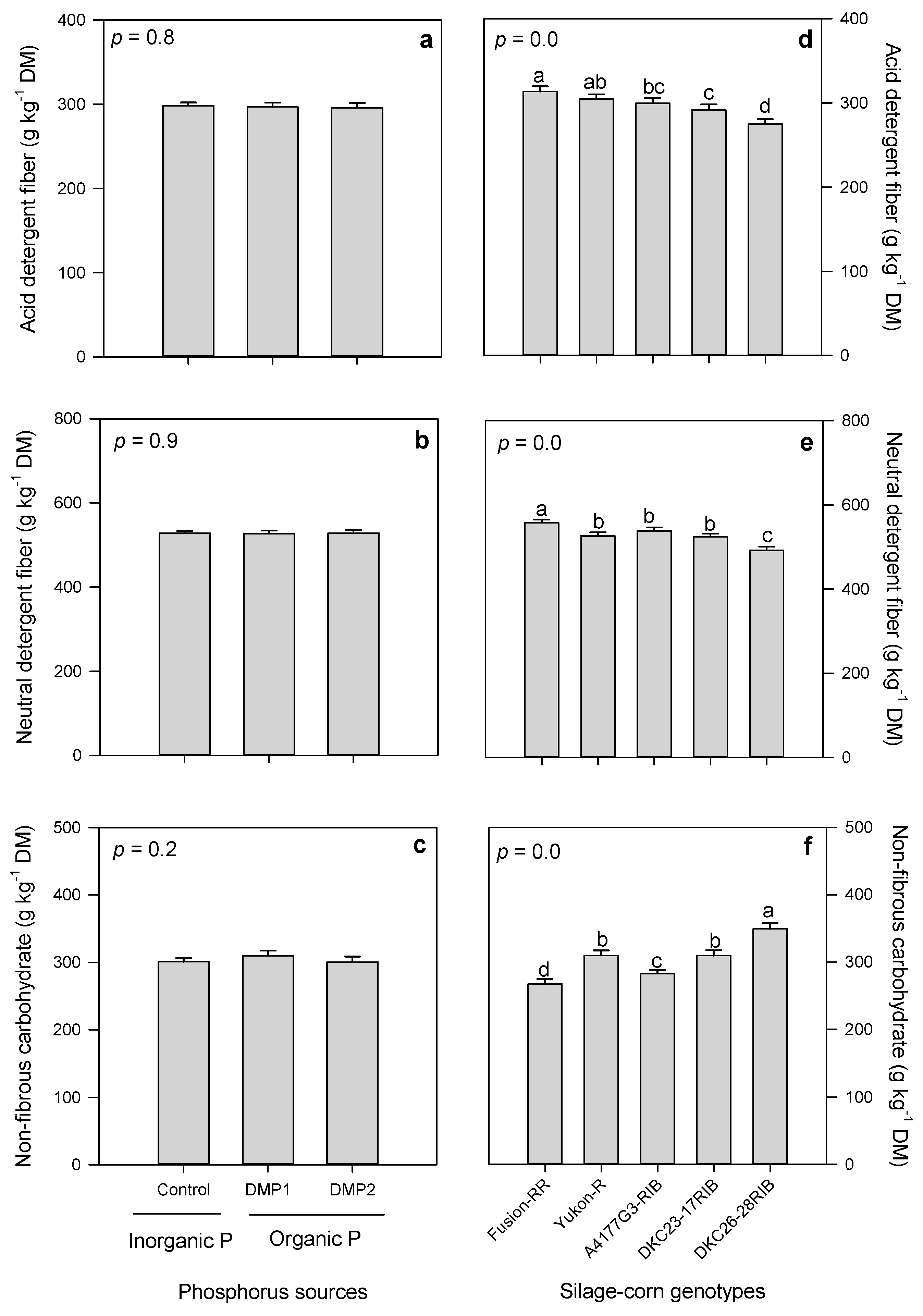
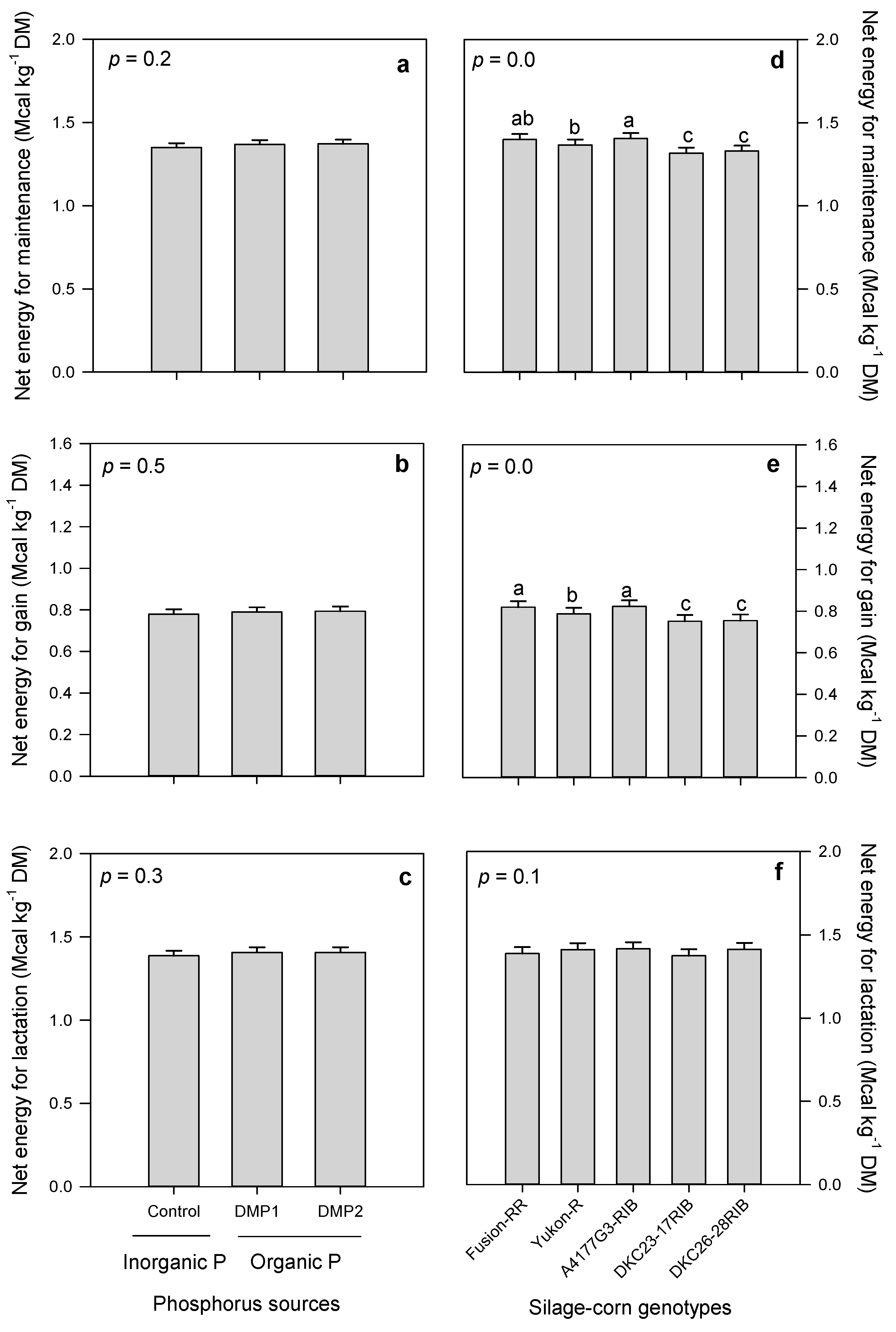
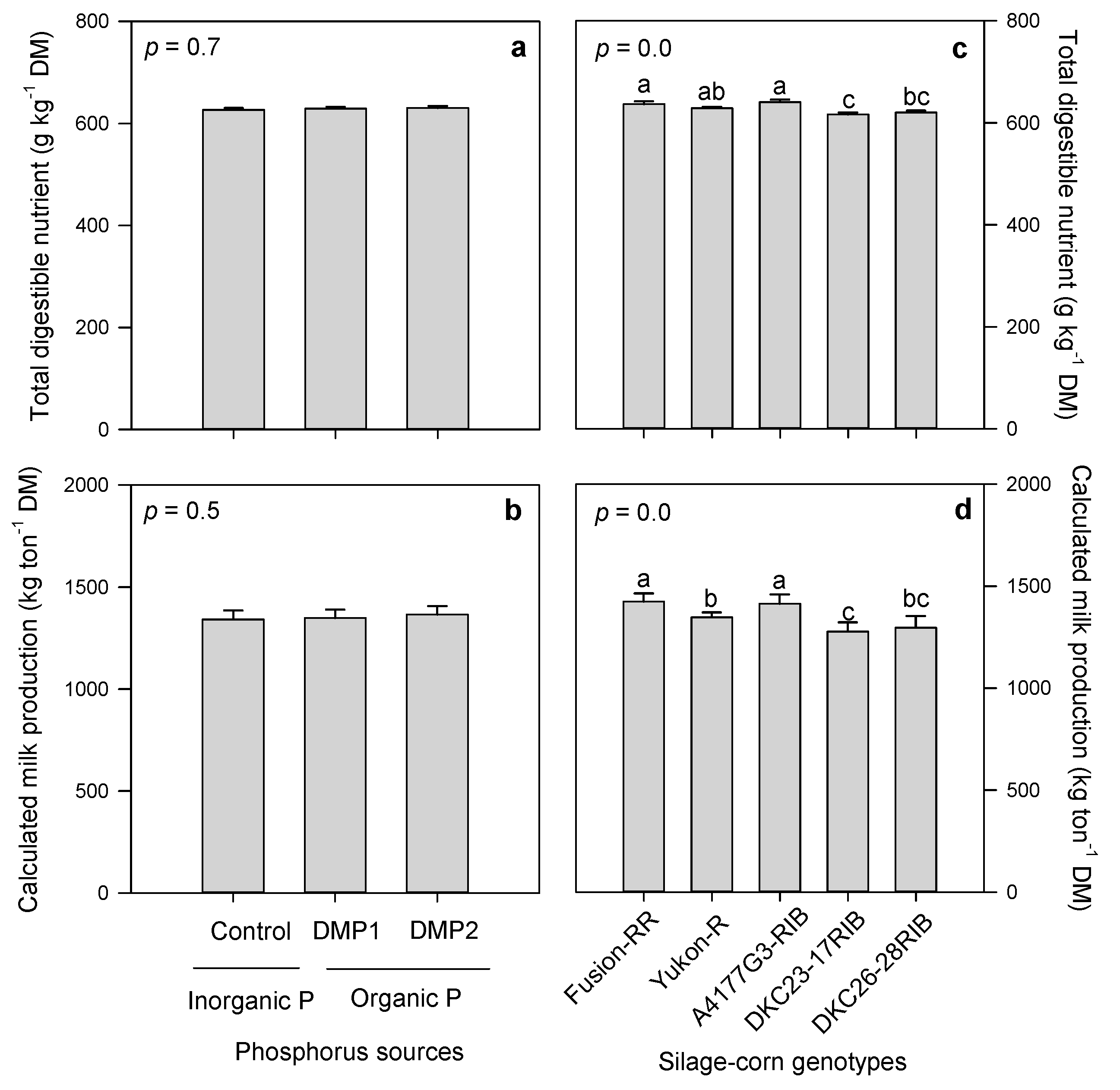
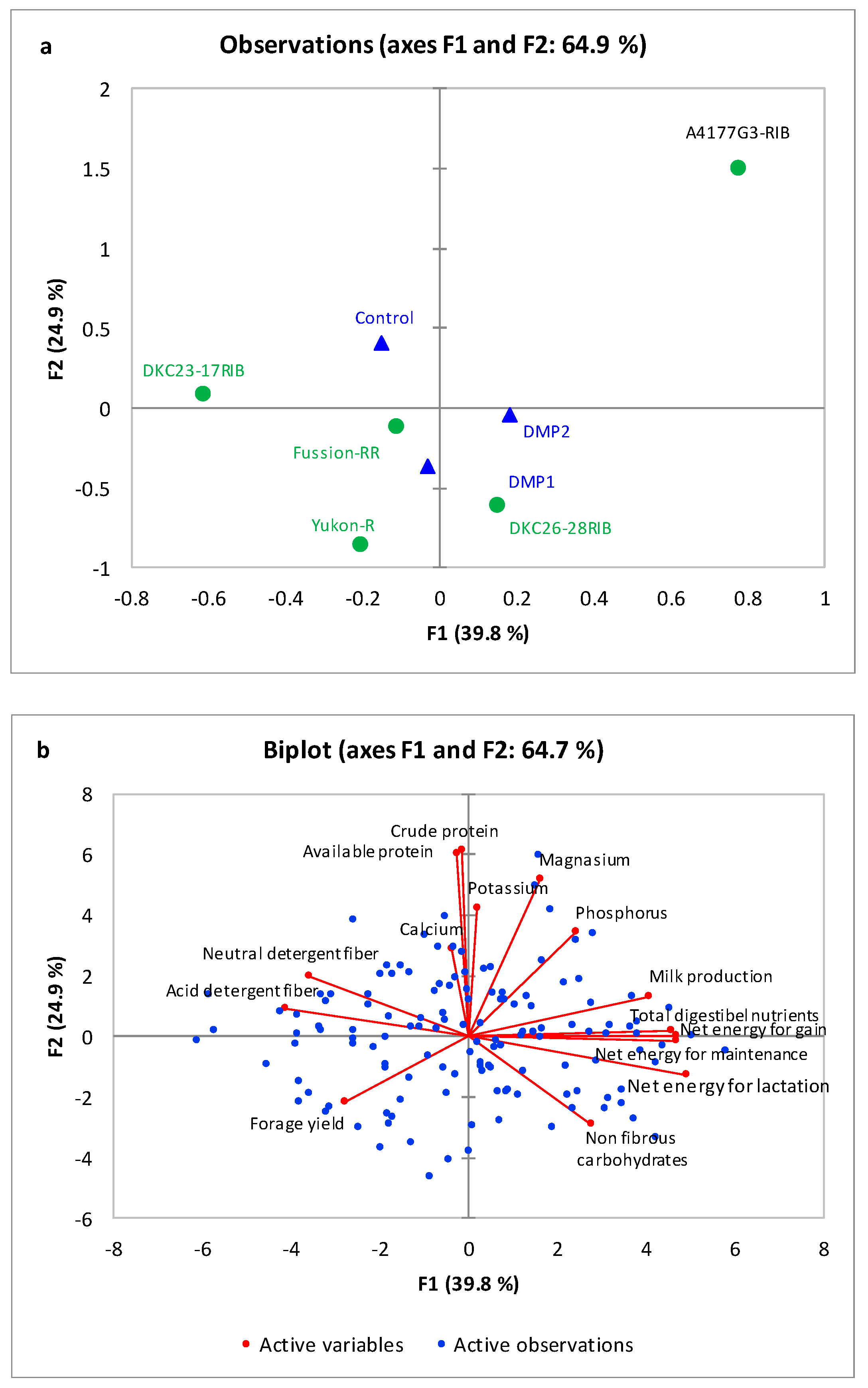
3.2. Forage Quality
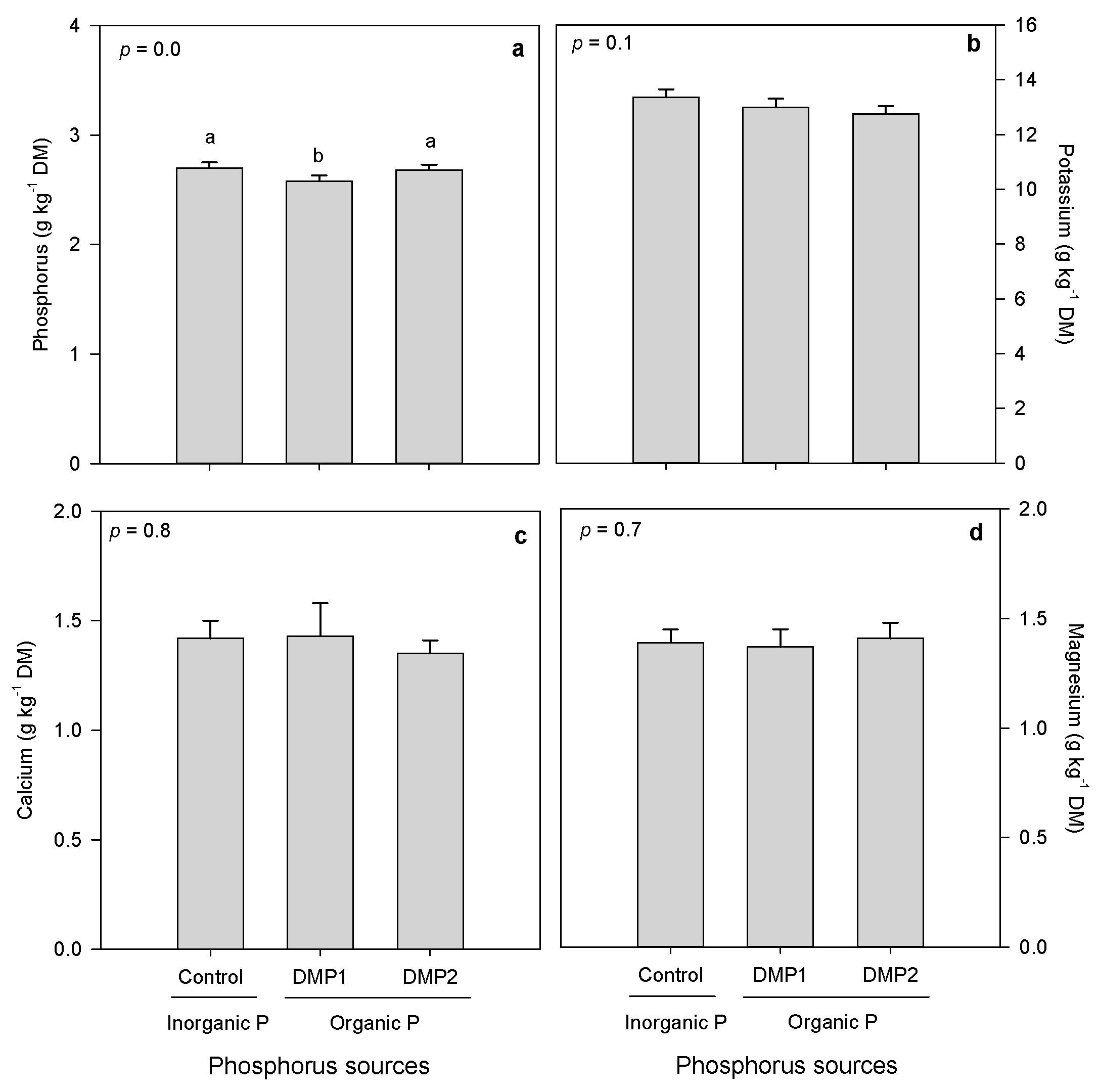
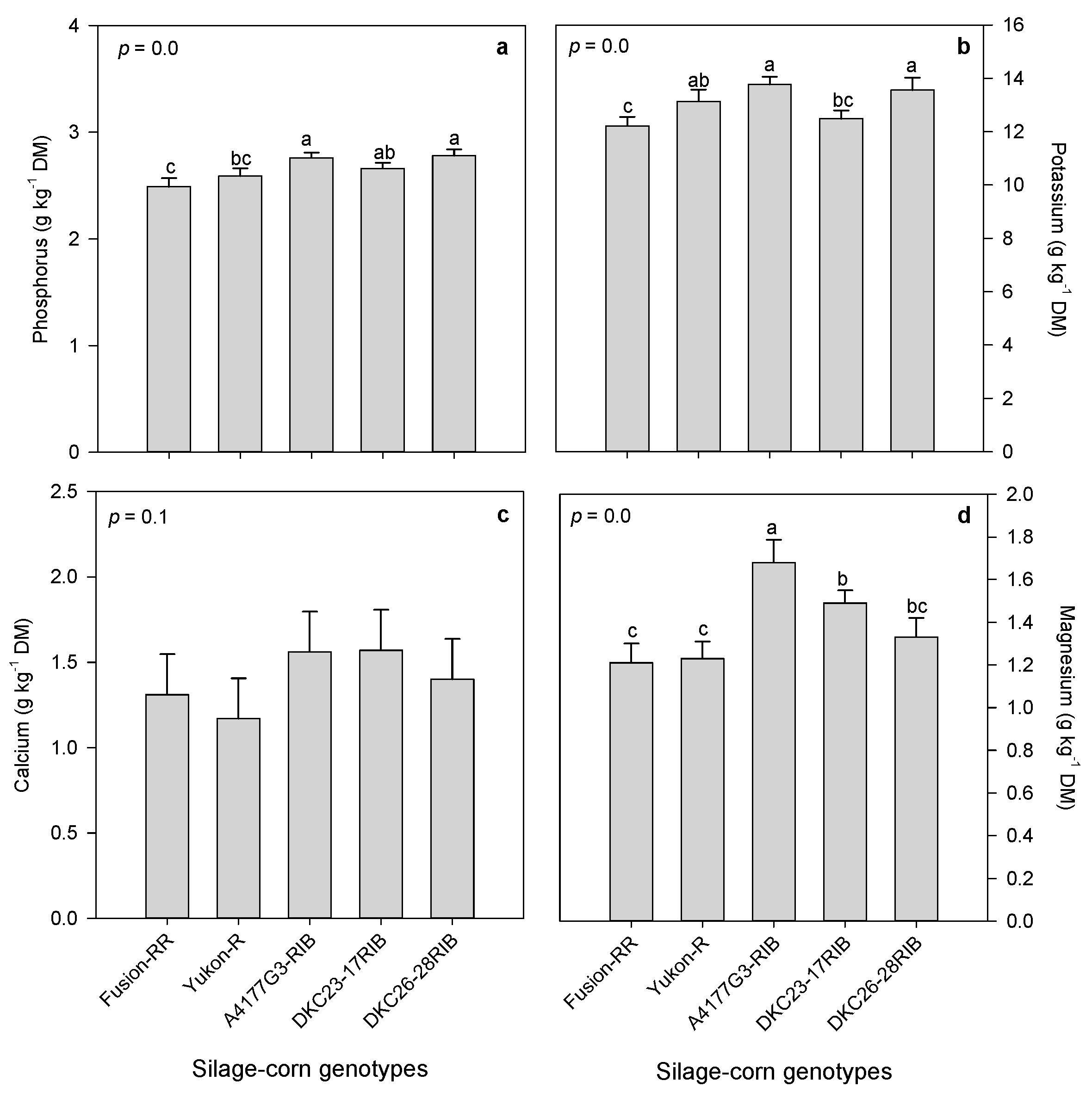
3.2.1. Mineral Composition
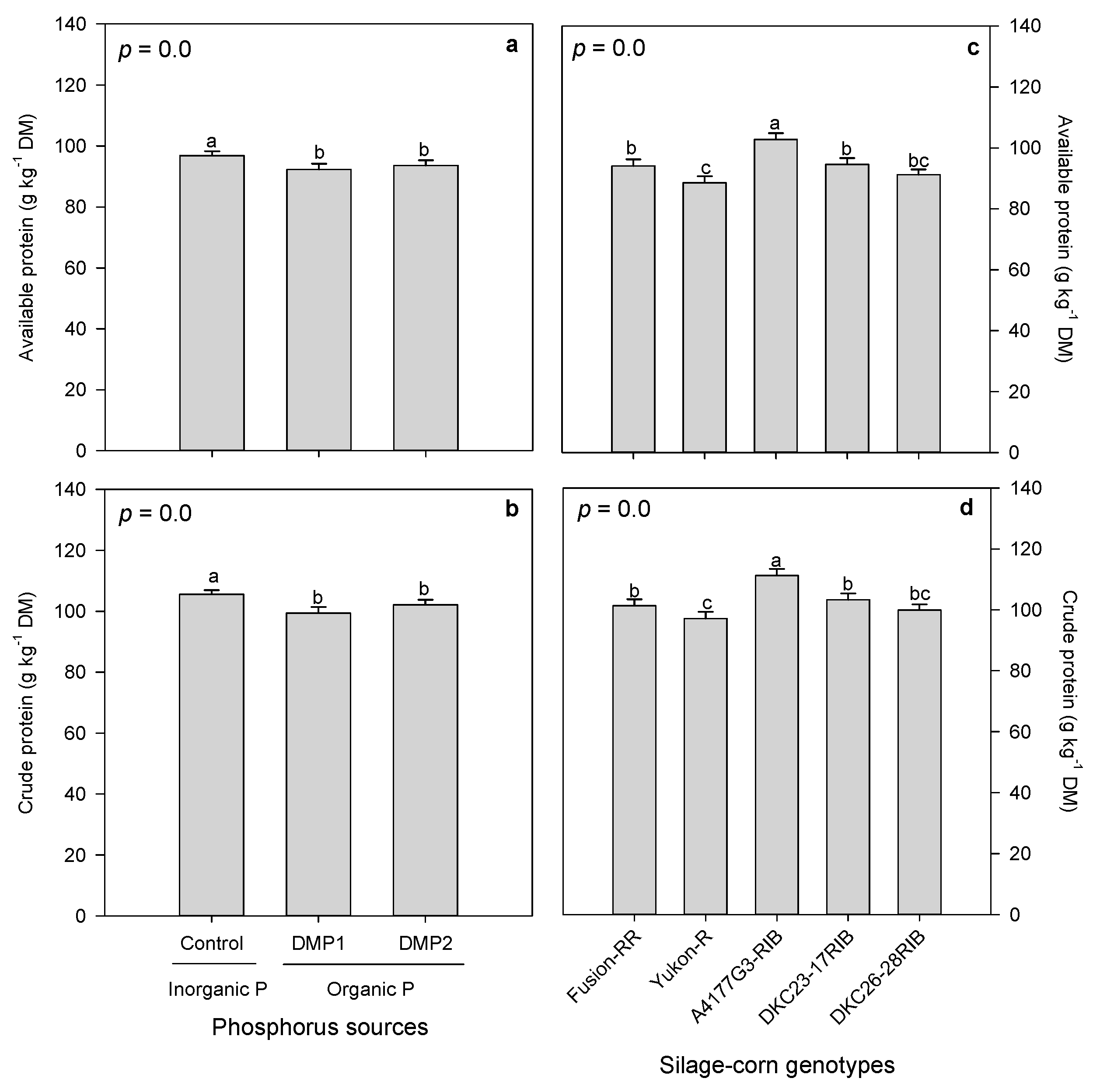
3.2.2. Forage Proteins
3.2.3. Fiber Contents
3.2.4. Forage Energies
3.2.5. Total Digestible Nutrients and Calculated Milk Production
4. Discussion
4.1. Forage Yield
4.2. Forage Quality
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Tilman, D.; Balzer, C.; Hill, J.; Befort, B.L. Global food demand and the sustainable intensification of agriculture. Proc. Natl. Acad. Sci. USA 2011, 108, 20260–20264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fedoroff, N.V. Food in a future of 10 billion. Agric. Food Secur. 2015, 4, 11. [Google Scholar] [CrossRef] [Green Version]
- FAO. How to Feed the World in 2050. Available online: http://www.fao.org/fileadmin/templates/wsfs/docs/expert_paper/How_to_Feed_the_World_in_2050.pdf (accessed on 15 March 2019).
- Ittersum, V. Food Security in 2050 on a Global Scale Achievable but Greatly Challenging. Available online: https://phys.org/news/2011-05-food-global-scale-greatly.html (accessed on 9 May 2019).
- Ranum, P.; Pena-Rosas, J.P.; Garcia-Casal, M.N. Global maize production, utilization, and consumption. Ann. N. Y. Acad. Sci. 2014, 1312, 105–112. [Google Scholar] [CrossRef] [PubMed]
- Khan, N.A.; Yu, P.; Ali, M.; Cone, J.W.; Hendriks, W.H. Nutritive value of maize silage in relation to dairy cow performance and milk quality. J. Sci. Food Agric. 2015, 95, 238–252. [Google Scholar] [CrossRef] [PubMed]
- Rankin, M. Choosing between Alfalfa and Corn Silage; University of Wisconsin-Extension: Madison, WI, USA, 2014. [Google Scholar]
- Kirkland, R.M.; Steen, R.W.J.; Gordon, F.J.; Keady, T.W.J. The influence of grass and maize silage quality on apparent diet digestibility, metabolizable energy concentration and intake of finishing beef cattle. Grass Forage Sci. 2005, 60, 244–253. [Google Scholar] [CrossRef]
- Karsten, H.D.; Roth, G.W.; Muller, L.D. Evaluation of corn hybrids at two stages of development for grazing heifers. Agrono. J. 2003, 95, 870–877. [Google Scholar] [CrossRef]
- Fitzgerald, J.J.; Murphy, J.J. A comparison of low starch maize silage and grass silage and the effect of concentrate supplementation of the forages or inclusion of maize grain with the maize silage on milk production by dairy cows. Livest. Prod. Sci. 1999, 57, 95–111. [Google Scholar] [CrossRef]
- Martin, N.P.; Russelle, M.P.; Powell, J.M.; Sniffen, C.J.; Smith, S.I.; Tricarico, J.M.; Grant, R.J. Invited review: Sustainable forage and grain crop production for the US dairy industry. J. Dairy Sci. 2017, 100, 9479–9494. [Google Scholar] [CrossRef] [Green Version]
- Hart, J.M.; Marx, E.S.; Christensen, N.W.; Moore, J.A. Nutrient management strategies. J. Dairy Sci. 1997, 80, 2659–2666. [Google Scholar] [CrossRef]
- Walsh, J.J.; Jones, D.L.; Edwards-Jones, G.; Williams, A.P. Replacing inorganic fertilizer with anaerobic digestate may maintain agricultural productivity at less environmental cost. J. Plant Nutr. Soil Sci. 2012, 175, 840–845. [Google Scholar] [CrossRef]
- Agriculture, N.L. A Factsheet Series on Environmental Guidelines for Livestock Producers; Agriculture, Newfoundland and Labrador; Publication SLM 045; Department of Forest Resources and Agrifoods: Carbonear, NL, Canada, 2002. [Google Scholar]
- Seleiman, M.F.; Selim, S.; Jaakkola, S.; Mäkelä, P.S.A. Chemical composition and in vitro digestibility of whole-crop maize fertilized with synthetic fertilizer or digestate and harvested at two maturity stages in Boreal growing conditions. Agric. Food Sci. 2017, 26, 47–55. [Google Scholar] [CrossRef]
- Seleiman, M.F.; Santanen, A.; Stoddard, F.L.; Makela, P. Feedstock quality and growth of bioenergy crops fertilized with sewage sludge. Chemosphere 2012, 89, 1211–1217. [Google Scholar] [CrossRef] [PubMed]
- Glass, A.D.M.; Beaton, J.D.; Bomke, A. Role of P in plant nutrition. In Proceedings of the Western Canada Phosphate Symposium, Calgary, AB, Canada, 11–12 March 1980; pp. 357–368. [Google Scholar]
- Plénet, D.; Etchebest, S.; Mollier, A.; Pellerin, S. Growth analysis of maize field crops under phosphorus deficiency-I. Leaf growth. Plant Soil 2000, 223, 117–130. [Google Scholar] [CrossRef]
- Assuero, S.G.; Mollier, A.; Pellerin, S. The decrease in growth of phosphorus-deficient maize leaves is related to a lower cell production. Plant Cell Environ. 2004, 27, 887–895. [Google Scholar] [CrossRef]
- Raghothama, K.G. Phosphate acquisition. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1999, 50, 665–693. [Google Scholar] [CrossRef] [PubMed]
- Kulhánek, M.; Balík, J.; Černý, J.; Nedvěd, V.; Kotková, B. The influence of different intensities of phosphorus fertilizing on available phosphorus contents in soils and uptake by plants. Plant Soil Environ. 2007, 53, 382–387. [Google Scholar] [CrossRef]
- Hinsinger, P. Bioavailability of soil inorganic P in the rhizosphere as affected by root-induced chemical changes: A review. Plant Soil 2001, 237, 173–195. [Google Scholar] [CrossRef]
- Vance, C.P.; Uhde-Stone, C.; Allan, D.L. Phosphorus acquisition and use: Critical adaptations by plants for securing a nonrenewable resource. New Phytol. 2003, 157, 423–447. [Google Scholar] [CrossRef]
- Cordell, D.; Drangert, J.O.; White, S. The story of phosphorus: Global food security and food for thought. Glob. Environ. Chang. 2009, 19, 292–305. [Google Scholar] [CrossRef]
- Newton, G.L.; Bernard, J.K.; Hubbard, R.K.; Allison, J.R.; Lowrance, R.R.; Gascho, G.J.; Gates, R.N.; Vellidis, G. Managing manure nutrients through multi-crop forage production. J. Dairy Sci. 2003, 86, 2243–2252. [Google Scholar] [CrossRef]
- Cherney, J.H.; Cherney, D.J.R. Impact of fertilization on forage production and animal performance. In Proceedings of the Colloque sur les plantes fourrageres, Drummondville, QC, Canada, 9 December 2015; pp. 1–7. [Google Scholar]
- Moreno-Reséndez, A.; Cantú Brito, J.E.; Reyes-Carrillo, J.L.; Contreras-Villarreal, V. Forage maize nutritional quality according to organic and inorganic fertilization. Sci. Agropecu. 2017, 8, 127–135. [Google Scholar] [CrossRef]
- Corrales, I.; Amenós, M.; Poschenrieder, C.; Barceló, J. Phosphorus efficiency and root exudates in two contrasting tropical maize varieties. J. Plant Nutr. 2007, 30, 887–900. [Google Scholar] [CrossRef]
- Ozturk, L.; Eker, S.; Torun, B.; Cakmak, I. Variation in phosphorus efficiency among 73 bread and durum wheat genotypes grown in a phosphorus-deficient calcareous soil. Plant Soil 2005, 269, 69–80. [Google Scholar] [CrossRef]
- Amodu, J.T.; Akpensuen, T.T.; Dung, D.D.; Tanko, R.J.; Musa, A.; Abubakar, S.A.; Hassan, M.R.; Jegede, J.O.; Sani, I. Evaluation of maize accessions for nutrients composition, forage and silage yields. J. Agric. Sci. 2014, 6, 178–187. [Google Scholar] [CrossRef]
- Faisal, S.; Shah, S.N.M.; Majid, A.; Khan, A. Effect of organic and inorganic fertilizers on protein, yield and related traits of maize varieties. Int. J. Agric. Crop Sci. 2013, 6, 1299–1303. [Google Scholar]
- Peña-Ramos, A.; Núñez-Hernández, G.; González-Castañeda, F. Forage potential of some maize populations and relationships between their agronomic characteristics and nutritional quality. Téc. Pecu. Méx. 2002, 40, 215–228. [Google Scholar]
- Eghball, B.; Power, J.F. Phosphorus- and nitrogen-based manure and compost applications: Corn production and soil phosphorus. Soil Sci. Soc. Am. J. 1999, 63, 895–901. [Google Scholar] [CrossRef]
- Hirzel, J.; Matus, I.; Novoa, F.; Walter, I.; Walter, I. Effect of poultry litter on silage maize (Zea mays L.) production and nutrient uptake. Span. J. Agric. Res. 2007, 5, 8. [Google Scholar] [CrossRef]
- Toth, J.D.; Dou, Z.; Ferguson, J.D.; Galligan, D.T.; Ramberg, C.F. Nitrogen- vs. phosphorus-based dairy manure applications to field crops. J. Environ. Qual. 2006, 35, 2302–2312. [Google Scholar] [CrossRef]
- Group, S.C.W. The Canadian System of Soi1 Classification, 3rd ed.; Agriculture and Agrifoods Canada, National Research Council of Canada: Ottawa, ON, Canada, 1998. [Google Scholar]
- Kirby, G.E. Soils of the Pasadena-Deer Lake Area, Newfoundland; Soil and Land Irlanagement Division: St. John’s, NL, Canada, 1988; Available online: http://sis.agr.gc.ca/cansis/publications/surveys/nf/nf17/nf17report.pdf (accessed on 15 March 2019).
- Bal, M.A.; Coors, J.G.; Shaver, R.D. Impact of the maturity of corn for use as silage in the diets of dairy cows on intake, digestion, and milk production. J. Dairy Sci. 1997, 80, 2497–2503. [Google Scholar] [CrossRef]
- Kwabiah, A.B. Growth, maturity, and yield responses of silage maize (Zea mays L.) to hybrid, planting date and plastic mulch. J. New Seeds 2005, 7, 37–59. [Google Scholar] [CrossRef]
- Guyader, J.; Baron, V.S.; Beauchemin, K.A. Corn forage yield and quality for silage in short growing season areas of the Canadian Prairies. Agronomy 2018, 8, 164. [Google Scholar] [CrossRef]
- Min, D.H.; Vough, L.R.; Reeves, J.B. Dairy slurry effects on forage quality of orchardgrass, reed canarygrass and alfalfa-grass mixtures. Anim. Feed Sci. Technol. 2002, 95, 143–157. [Google Scholar] [CrossRef]
- Ferreira, G.; Alfonso, M.; Depino, S.; Alessandri, E. Effect of planting density on nutritional quality of green-chopped corn for silage. J. Dairy Sci. 2014, 97, 5918–5921. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gallo, A.; Moschini, M.; Cerioli, C.; Masoero, F. Use of principal component analysis to classify forages and predict their calculated energy content. Anim. Int. J. Anim. Biosci. 2013, 7, 930–939. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Horwitz, W. Official methods of analysis. In The Association of Officeal Analytical Chemists; Benjamin Frankline Statio: Washington, DC, USA, 1980; p. 1018. [Google Scholar]
- Van Soest, P.J.; Fox, D.G. Discounts for net energy and protein-5th Revision. In Proceedings of the Cornell Nutrition Conference for Feed Manufacturers, Rochester, NY, USA, 13–15 October 1992; p. 40. [Google Scholar]
- Weiss, W.P.; Conrad, H.R.; St. Pierre, N.R. A theoretically-based model for predicting total digestible nutrient values of forages and concentrates. Anim. Feed Sci. Technol. 1992, 39, 95–110. [Google Scholar] [CrossRef]
- National, R.C. Nutrient Requirements of Dairy Cattle, 7th ed.; National Academy of Science: Washington, DC, USA, 2001. [Google Scholar]
- Norell, R.J. The Wisconsin system fo revaluating corn silage. In Proceedings of the 2005 Idaho Alfalfa and Forage Conference, Twin Falls, ID, USA, 7–8 February 2005; pp. 44–51. [Google Scholar]
- King, M.; Altdorff, D.; Li, P.; Galagedara, L.; Holden, J.; Unc, A. Northward shift of the agricultural climate zone under 21(st)-century global climate change. Sci. Rep. 2018, 8, 7904. [Google Scholar] [CrossRef]
- Yan, F.; Schubert, S.; Mengel, K. Soil pH changes during legume growth and application of plant material. Biol. Fertil. Soils 1996, 23, 236–242. [Google Scholar] [CrossRef]
- Carstensen, A.; Herdean, A.; Schmidt, S.B.; Sharma, A.; Spetea, C.; Pribil, M.; Husted, S. The Impacts of phosphorus deficiency on the photosynthetic electron transport chain. Plant Physiol. 2018, 177, 271–284. [Google Scholar] [CrossRef]
- Efthimiadou, A.; Bilalis, D.; Karkanis, A.; Froud-Williams, B. Combined organic/inorganic fertilization enhance soil quality and increased yield, photosynthesis and sustainability of sweet maize crop. Aust. J. Crop Sci. 2009, 4, 722–729. [Google Scholar]
- Elhindi, K. Evaluation of composted green waste fertigation through surface and subsurface drip irrigation systems on pot marigold plants (Calendula officinalis L.) grown on sandy soil. Aust. J. Crop Sci. 2012, 6, 1249–1259. [Google Scholar]
- Nadeem, M.; Pham, T.H.; Thomas, R.; Galagedara, L.; Kavanagh, V.; Zhu, X.; Ali, W.; Cheema, M. Potential role of root membrane phosphatidic acid in superior agronomic performance of silage-corn cultivated in cool climate cropping systems. Physiol. Plant. 2019. [Google Scholar] [CrossRef] [PubMed]
- Kronqvist, C. Minerals to Dairy Cows with Focus on Calcium and Magnesium Balance. Ph.D. Thesis, Swedish University of Agricultural Sciences, Uppsala, Sweden, 2011. [Google Scholar]
- Plaizier, J.C.; Garner, T.; Droppo, T.; Whiting, T. Nutritional practices on Manitoba dairy farms. Can. J. Anim. Sci. 2004, 84, 501–509. [Google Scholar] [CrossRef]
- Holtenius, K.; Kronqvist, C.; Briland, E.; Sporndly, R. Magnesium absorption by lactating dairy cows on a grass silage-based diet supplied with different potassium and magnesium levels. J. Dairy Sci. 2008, 91, 743–748. [Google Scholar] [CrossRef] [PubMed]
- Leytem, A.B.; Dungun, R.S.; Moore, A. Nutrient availability to corn from dairy manures and fertilizer in a calcareous soil. Soil Sci. 2011, 176, 1–9. [Google Scholar] [CrossRef]
- Suttle, N. Mineral Nutrition of Livestock, 4th ed.; CABI: Wallingford, UK, 2010. [Google Scholar]
- Roth, G.W.; Heinrichs, A.J. Maize Silage Production and Management; Agronomy fact-18; Information and Communication Technology, The College of Agricultural Sciences, Pennsylvania State University: State College, PA, USA, 2001. [Google Scholar]
- Ballard, C.S.; Thomas, E.D.; Tsang, D.S.; Mandebvu, P.; Sniffen, C.J.; Endres, M.I.; Carter, M.P. Effect of corn silage hybrid on dry matter yield, nutrient composition, in vitro digestion, intake by dairy heifers, and milk production by dairy cows. J. Dairy Sci. 2001, 84, 442–452. [Google Scholar] [CrossRef]
- Garibay, S.V.; Stamp, P.; Ammon, H.U.; Feil, B. Yield and quality components of silage maize in killed and live cover crops sods. Eur. J. Agron. 1997, 6, 179–190. [Google Scholar] [CrossRef]
- Seleiman, M.F.; Santanen, A.; Kleemola, J.; Stoddard, F.L.; Mäkelä, P.S.A. Improved sustainability of feedstock production with sludge and interacting mycorrhiza. Chemosphere 2013, 91, 1236–1242. [Google Scholar] [CrossRef] [PubMed]
- Fletcher, A.L.; Chakwizira, E. Developing a critical nitrogen dilution curve for forage brassicas. Grass Forage Sci. 2012, 67, 13–23. [Google Scholar] [CrossRef]
- Bélanger, G.; Gastal, F. Nitrogen utilization by forage grasses. Can. J. Plant Sci. 2000, 80, 11–20. [Google Scholar] [CrossRef]
- Castillo-Jimenez, M.; Roja-Bourrillon, A.; WingChing-Jones, R. Nutritional value of silage made with a mixture of corn and mung bean (Vigna radiata). Agron. Costarric. 2009, 33, 133–146. [Google Scholar]
- Gallegos-Ponce, A.; Martínez-Ríos, A.; Fernando-Sánchez, M.; Figueroa-Viramontes, R.; Berumen-Padilla, S.; Venegas-Soto, J.; Quevedo-Guillen, J.D.D.; Escobedo-López, D.; Silos-Calzada, M.C. Nutritional quality of forage maize (Zea mays L.) under limited water logging conditions. Agrofaz 2012, 12, 59–66. [Google Scholar]
- Johnson, C.R.; Reiling, B.A.; Mislevy, P.; Hall, M.B. Effects of nitrogen fertilization and harvest date on yield, digestibility, fiber, and protein fractions of tropical grasses. J. Anim. Sci. 2001, 79, 2439–2448. [Google Scholar] [CrossRef] [PubMed]
- Nazli, R.I.; Inal, I.; Kusvuran, A.; Demirbas, A.; Tansi, V. Effects of different organic materials on forage yield and nutrient uptake of silage maize (Zea mays L.). J. Plant Nutr. 2016, 39, 912–921. [Google Scholar] [CrossRef]
- Schwab, E.C.; Shaver, R.D.; Lauer, J.G.; Coors, J.G. Estimating silage energy value and milk yield to rank corn hybrids. Anim. Feed Sci. Technol. 2003, 109, 1–18. [Google Scholar] [CrossRef]
- DePeters, E.J.; Fadel, J.G.; Arana, M.J.; Ohanesian, N.; Etchebarne, M.A.; Hamilton, C.A.; Hinders, R.G.; Maloney, M.D.; Old, C.A.; Riordan, T.J.; et al. Variability in the chemical composition of seventeen selected by-product feedstuffs used by the California dairy industry. Prof. Anim. Sci. 2000, 16, 69–99. [Google Scholar] [CrossRef]
- Harris, B., Jr. CIR1122 Nonstructrual and Structural Carbohydrates in Dairy Cattle Rations; University of Florida IFAS Extension: Gainesville, FL, USA, 1993; pp. 1–5. [Google Scholar]
- Tine, M.A.; McLeod, K.R.; Erdman, R.A.; Baldwin, R.L.T. Effects of brown midrib corn silage on the energy balance of dairy cattle. J. Dairy Sci. 2001, 84, 885–895. [Google Scholar] [CrossRef]
- Di Marco, O.N.; Aello, M.S.; Nomdedeu, M.; Van Houtte, S. Effect of maize crop maturity on silage chemical composition and digestibility (in vivo, in situ and in vitro). Anim. Feed Sci. Technol. 2002, 99, 37–43. [Google Scholar] [CrossRef]
- de la Cruz-Lázaro, E.; Rodríguez-Herrera, S.A.; Palomo-Gil, A.; López Benítez, A.; Robledo-Torres, V.; Gómez-Vázquez, A.; Osorio-Osorio, R. Combining ability of protein high quality maíz inbred lines for forage characteristics. Ecosistemas Recur. Agropecu. 2007, 23, 57–67. [Google Scholar]
- Núñez-Hernández, G.; Contreras, G.F.; Faz-Contreras, R. Yield, chemical composition and in vitro digestibility of tropical and temperate corn hybrids in the arid region of Mexico. Av. Investig. Agropecu. 2004, 8, 1–19. [Google Scholar]
- Posada, O.S.; Rosero, N.R.; Rodríguez, N.; Costa, C.A. Comparison of methods to determine the energy value of feeds for ruminants. Rev. MVZ Córdoba 2012, 17, 3184–3192. [Google Scholar]
- Kim, J.D.; Kwon, C.H.; Kim, D.A. Yield and quality of silage corn as affected by hybrid maturity, Pplanting date and harvest stage. Asian Australas. J. Anim. Sci. 2001, 14, 1705–1711. [Google Scholar] [CrossRef]
- Olague-Ramirez, J.; Montemayor-Trejo, J.A.; Bravo-Sánchez, S.R.; Fortis-Hernández, M.; Aldaco-Nuncio, R.A.; Ruiz-Cerda, E. Agronomic characteristic and forage quality corn with subsurface drip irrigation. Tec. Pecu. Mex. 2006, 44, 351–357. [Google Scholar]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Soil Properties | Average of Three Growing Seasons (2015–2017) |
|---|---|
| Soil pH | 6.7 ± 0.1 |
| Phosphorus (mg L−1) | 78.3 ± 2.2 |
| Potassium (mg L−1) | 44.0 ± 3.2 |
| Calcium (mg L−1) | 1339.7 ± 156.7 |
| Magnesium (mg L−1) | 241.3 ± 13.6 |
| Organic matter (%) | 3.0 ± 0.0 |
| Sulphur (mg L−1) | 14.5 ± 0.4 |
| Zinc (mg L−1) | 1.3 ± 0.4 |
| Copper (mg L−1) | 1.4 ± 0.3 |
| Sodium (mg L−1) | 7.3 ± 0.9 |
| Iron (mg L−1) | 146.7 ± 8.8 |
| Boron (mg L−1) | 0.3 ± 0.1 |
| Manganese (mg L−1) | 17.0 ± 0.6 |
| Aluminum (mg L−1) | 1421.3 ± 46.3 |
| Growth Period | Crop Growing Seasons’ Average (2015–2017) | Thirty Years Average (1985–2014) | ||||||
|---|---|---|---|---|---|---|---|---|
| Mean Average Temperature (°C) | Mean Maximum Temperature (°C) | Mean Minimum Temperature (°C) | Rainfall (mm) | Mean Average Temperature (°C) | Mean Maximum Temperature (°C) | Mean Minimum Temperature (°C) | Rainfall (mm) | |
| May 16–31 | 9.0 ± 1.3 | 15.6 ± 1.4 | 2.4 ± 1.2 | 32.0 ± 15.0 | 8.5 | 14.2 | 2.8 | 44.1 |
| June 1–15 | 10.1 ± 0.6 | 17.5 ± 2.4 | 2.7 ± 1.2 | 65.0 ± 21.6 | 11.1 | 16.7 | 5.3 | 38.8 |
| June 16–30 | 14.5 ± 0.7 | 22.0 ± 0.6 | 7.1 ± 1.0 | 37.3 ± 5.8 | 14.2 | 20.2 | 8.1 | 33.3 |
| July 1–15 | 15.8 ± 0.7 | 22.7 ± 0.8 | 8.8 ± 0.6 | 30.7 ± 6.1 | 16.4 | 22.1 | 10.6 | 44.1 |
| July 16–31 | 16.5 ± 0.8 | 23.4 ± 1.1 | 9.6 ± 0.8 | 43.7 ± 19.4 | 17.7 | 23.0 | 12.2 | 43.5 |
| August 1–15 | 16.7 ± 0.1 | 24.1 ± 0.7 | 9.4 ± 0.4 | 32.3 ± 10.7 | 17.8 | 23.1 | 12.5 | 44.7 |
| August 16–31 | 16.9 ± 1.7 | 23.2 ± 1.5 | 10.3 ± 2.1 | 51.0 ± 31.2 | 16.1 | 20.9 | 11.2 | 57.0 |
| September 1–15 | 14.0 ± 0.9 | 19.8 ± 1.4 | 8.0 ± 0.5 | 76.7 ± 13.4 | 13.8 | 18.5 | 9.0 | 54.7 |
| September 16–30 | 10.3 ± 1.4 | 17.9 ± 3.7 | 2.2 ± 0.5 | 38.0 ± 21.4 | 11.6 | 16.3 | 7.0 | 48.4 |
| October 1–15 | 8.9 ± 1.5 | 16.2 ± 2.2 | 2.0 ± 0.8 | 80.7 ± 20.3 | 8.5 | 12.6 | 4.4 | 48.5 |
| 487.3 ± 103.6 | 457.1 | |||||||
| Genotype | Corn Heat Units (CHU) for Grain | Maturity Zones and CHU for Silage | Plant Height | Genetic Traits | Source |
|---|---|---|---|---|---|
| Fusion-RR | 2200 | 2000–2300 | Tall | Roundup Ready® Corn 2 (RR2) | Brett-Young Seeds Limited, Canada |
| Yukon-R | 2150 | 1900–2200 | Very tall | Roundup Ready® Corn 2 (RR2) | Brett-Young Seeds Limited, Canada |
| A4177G3-RIB | 2175 | 2000–2200 | Medium | Roundup Ready® Corn (VT3P-RR) | Pride Seeds, Canada |
| DKC23-17RIB | 2075 | 1800–2000 | Medium | Roundup Ready® Corn 2 (VT2P) | DEKALB®, Canada |
| DKC26-28RIB | 2150 | – | Medium to tall | Roundup Ready® Corn 2 (VT2PRIB) | DEKALB®, Canada |
| Manure Properties | Average of Three Growing Seasons | |
|---|---|---|
| DMP1 | DMP2 | |
| Dry matter (%) | 9.9 ± 0.5 | 2.9 ± 0.6 |
| pH | 6.9 ± 0.1 | 7.1 ± 0.1 |
| Nitrogen (g kg−1) | 3.8 ± 0.3 | 1.4 ± 0.1 |
| Phosphorus (g kg−1) | 0.7 ± 0.1 | 0.3 ± 0.0 |
| Potassium (g kg−1) | 3.6 ± 0.2 | 1.2 ± 0.0 |
| Total Calcium (g kg−1) | 1.7 ± 0.1 | 0.6 ± 0.1 |
| Total Magnesium (g kg−1) | 0.7 ± 0.0 | 0.3 ± 0.0 |
| Total Iron (mg kg−1) | 58.3 ± 5.5 | 18.0 ± 6.1 |
| Total Manganese (mg kg−1) | 22.7 ± 0.9 | 8.7 ± 2.0 |
| Total Copper (mg kg−1) | 4.5 ± 0.1 | 25.3 ± 3.9 |
| Total Zinc (mg kg−1) | 18.0 ± 1.5 | 8.0 ± 1.7 |
| Total Boron (mg kg−1) | 3.1 ± 0.1 | 0.8 ± 0.2 |
| Total Sodium (mg kg−1) | 826.7 ± 80.9 | 332.0 ± 74.7 |
| Forage Quality and Yield Indices | Genotype (G) | Phosphorus Source (P) | G × P | |
|---|---|---|---|---|
| Forage | Forage yield (Mg ha−1) | * | * | NS |
| Minerals | Phosphorus (g ka−1 DM) | * | * | NS |
| Potassium (g ka−1 DM) | * | NS | NS | |
| Calcium (g ka−1 DM) | NS | NS | NS | |
| Magnesium (g ka−1 DM) | * | NS | NS | |
| Proteins | Crude protein (g kg−1 DM) | * | * | NS |
| Available protein (g kg−1 DM) | * | * | NS | |
| Fibers | Acid detergent fiber (g kg−1 DM) | * | NS | NS |
| Neutral detergent fiber (g kg−1 DM) | * | NS | NS | |
| NFC | Non-fibrous carbohydrates (g kg−1 DM) | * | NS | NS |
| TDN | Total digestible nutrients (g kg−1 DM) | * | NS | NS |
| Energy | Net energy for lactation (Mcal kg−1 DM) | NS | NS | NS |
| Net energy for maintenance (Mcal kg−1 DM) | * | NS | NS | |
| Net energy for gain (Mcal kg−1 DM) | * | NS | NS | |
| Milk production | Calculated milk production (Ib ton−1 DM) | * | NS | NS |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ali, W.; Nadeem, M.; Ashiq, W.; Zaeem, M.; Thomas, R.; Kavanagh, V.; Cheema, M. Forage Yield and Quality Indices of Silage-Corn Following Organic and Inorganic Phosphorus Amendments in Podzol Soil under Boreal Climate. Agronomy 2019, 9, 489. https://doi.org/10.3390/agronomy9090489
Ali W, Nadeem M, Ashiq W, Zaeem M, Thomas R, Kavanagh V, Cheema M. Forage Yield and Quality Indices of Silage-Corn Following Organic and Inorganic Phosphorus Amendments in Podzol Soil under Boreal Climate. Agronomy. 2019; 9(9):489. https://doi.org/10.3390/agronomy9090489
Chicago/Turabian StyleAli, Waqas, Muhammad Nadeem, Waqar Ashiq, Muhammad Zaeem, Raymond Thomas, Vanessa Kavanagh, and Mumtaz Cheema. 2019. "Forage Yield and Quality Indices of Silage-Corn Following Organic and Inorganic Phosphorus Amendments in Podzol Soil under Boreal Climate" Agronomy 9, no. 9: 489. https://doi.org/10.3390/agronomy9090489