Establishment and Characterization of Callus and Cell Suspension Cultures of Selected Sorghum bicolor (L.) Moench Varieties: A Resource for Gene Discovery in Plant Stress Biology

 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Seed Surface Sterilisation and Germination
2.3. Initiation and Maintenance of Callus and Cell Suspension Cultures
2.4. Characterisation of Cell Suspension Cultures
2.5. Osmotic Stress Treatment
2.6. Viability Assays of Sorghum Cell Cultures
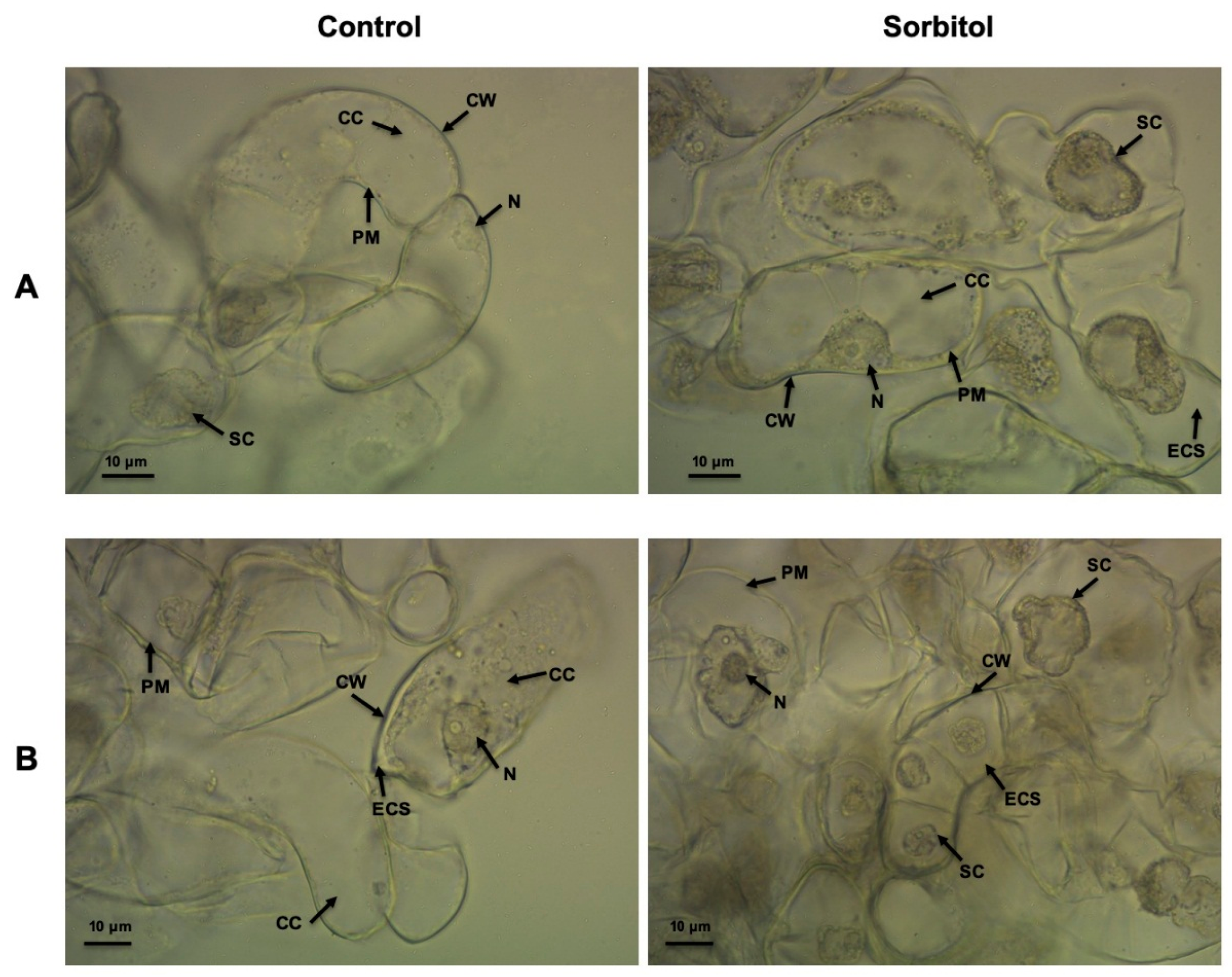
2.7. Light Microscopic Analysis of Osmotic Stressed Cell Cultures
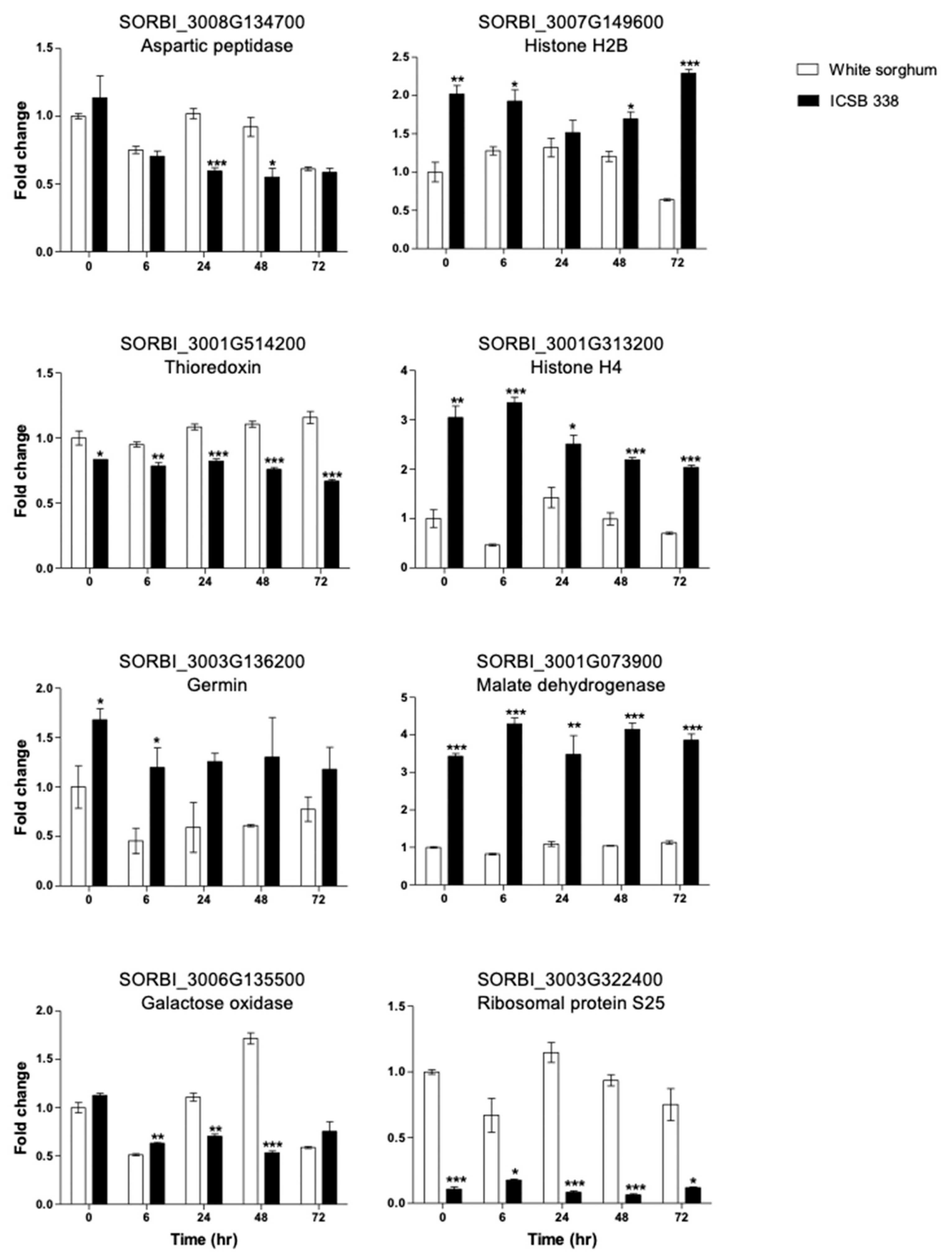
2.8. RNA Extraction and Gene Expression Analysis
2.9. Statistical Analysis
3. Results
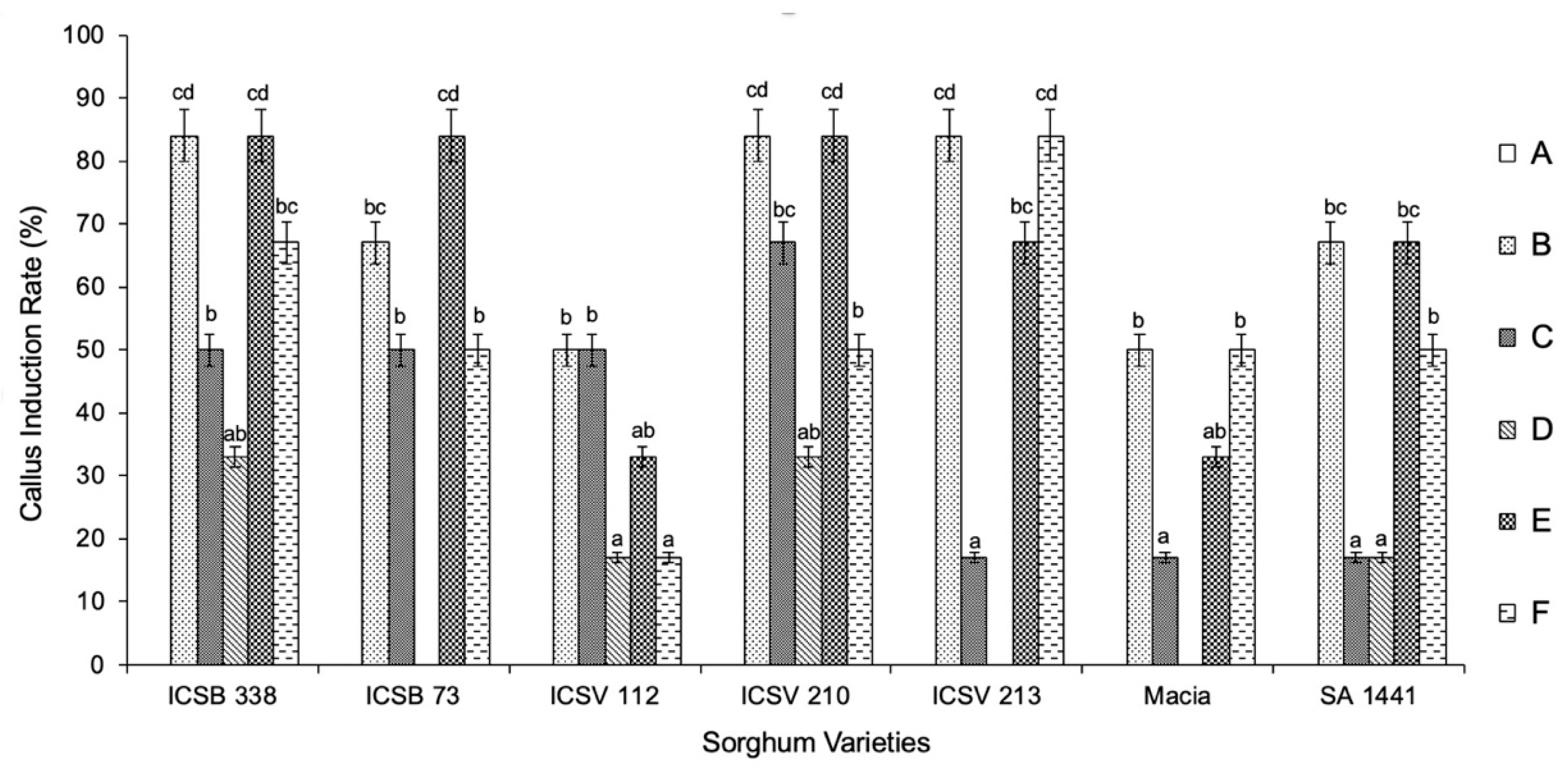
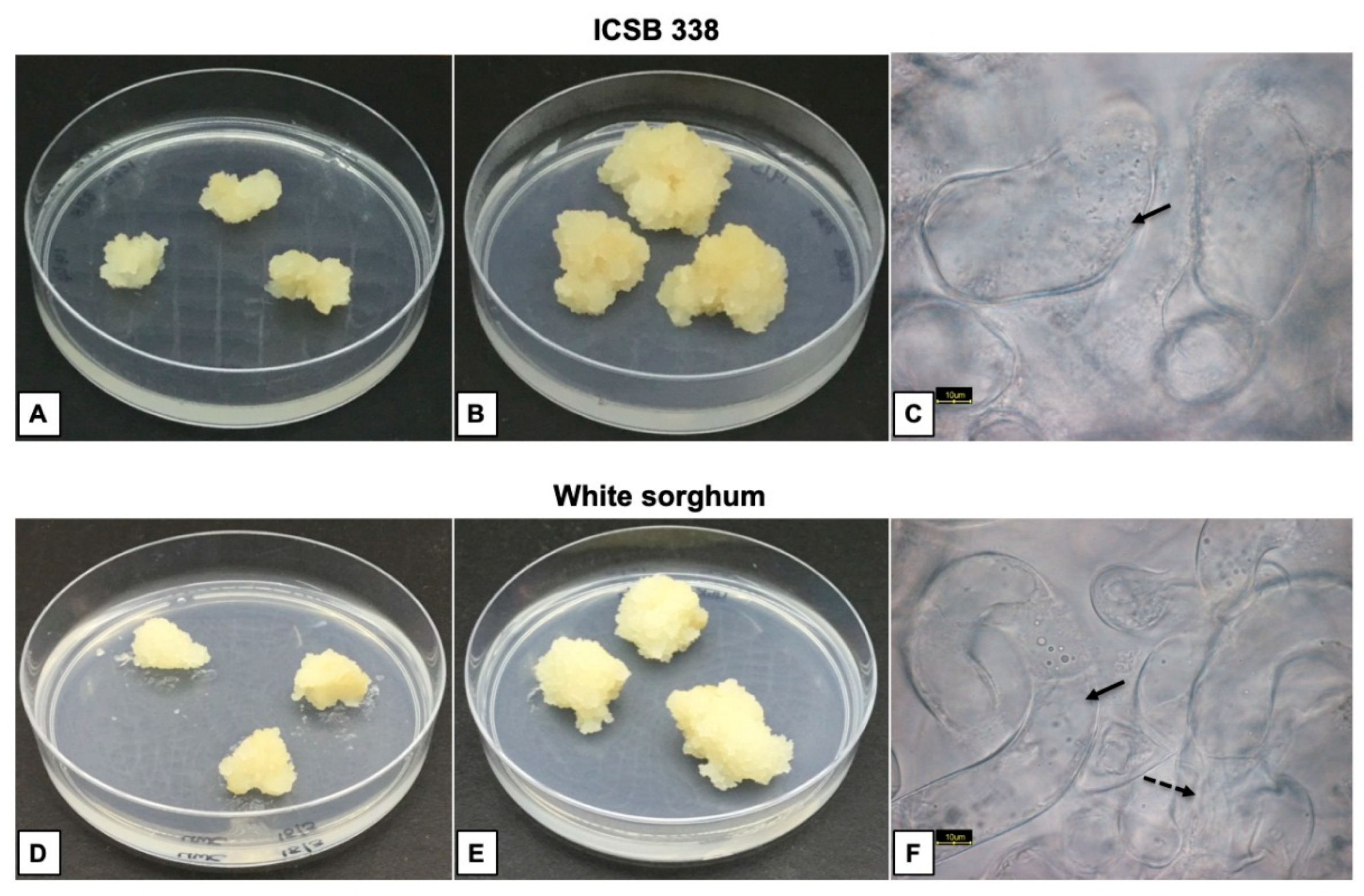
3.1. Callus Induction
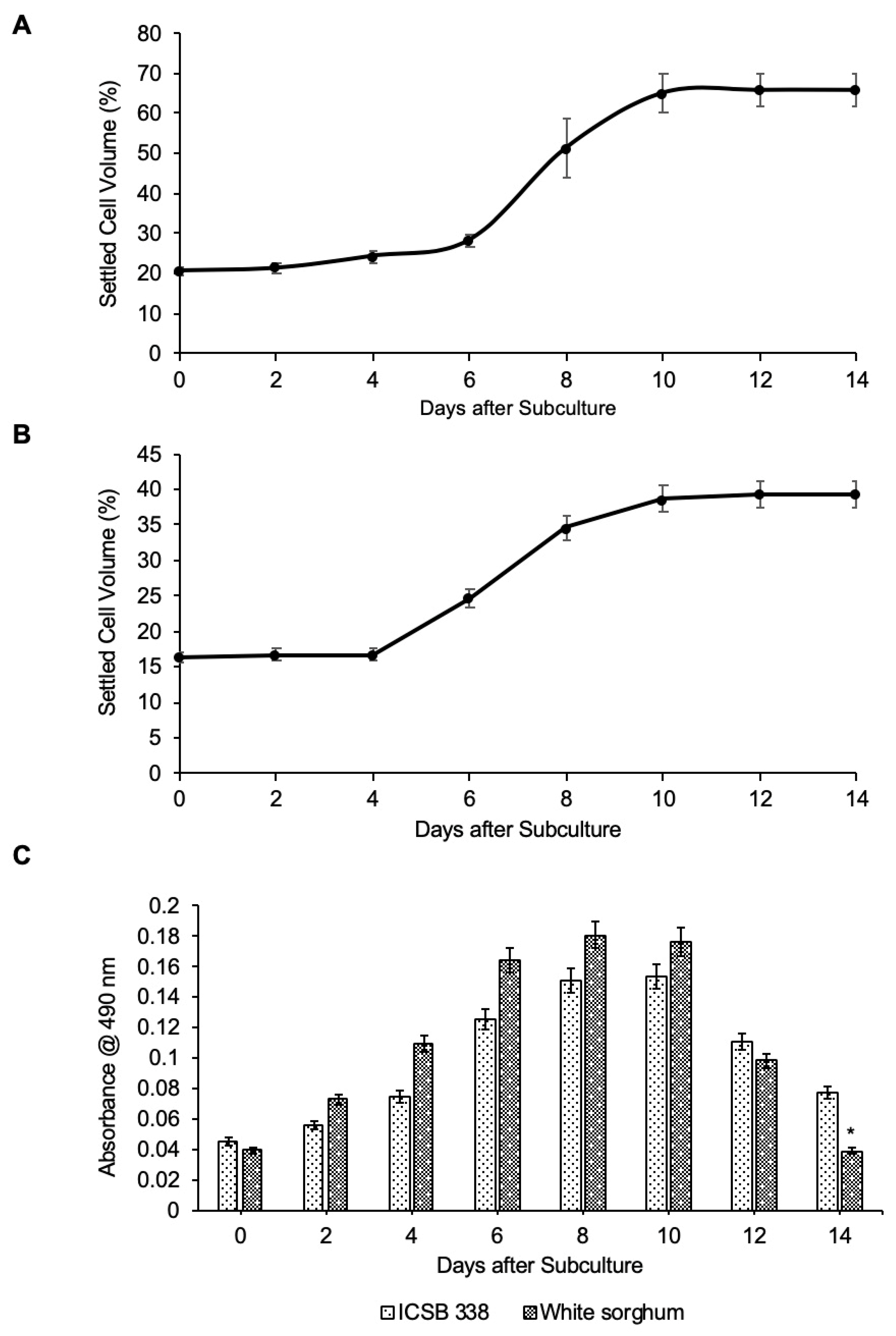
3.2. Establishment and Characterisation of Cell Suspension Cultures
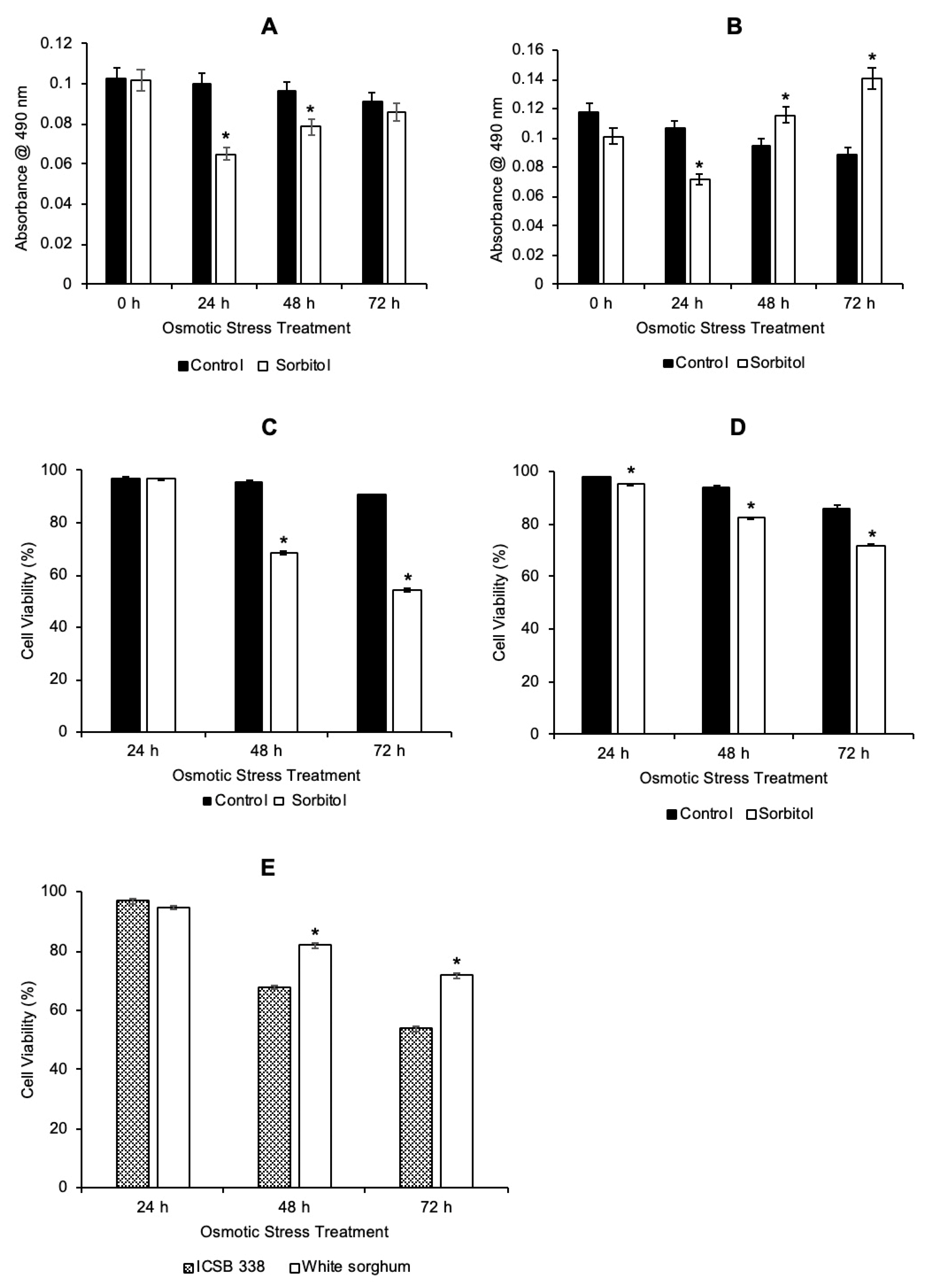
3.3. Comparative Analysis of the Two Sorghum Cell Suspension Cultures Following Osmotic Stress Treatment
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- FAOSTAT. 2017. Available online: http://www.fao.org (accessed on 22 March 2019).
- Kimber, C.T.; Dahlberg, J.A.; Kresovich, S. The gene pool of Sorghum bicolor and its improvement. In Genomics of the Saccharinae; Paterson, A.H., Ed.; Springer: New York, NY, USA, 2013; Volume 11, pp. 23–41. [Google Scholar]
- Motlhaodi, T.; Geleta, M.; Chite, S.; Fatih, M.; Ortiz, R.; Bryngelsson, T. Genetic diversity in sorghum [Sorghum bicolor (L.) Moench] germplasm from Southern Africa as revealed by microsatellite markers and agro-morphological traits. Genet. Resour. Crop Evol. 2017, 64, 599–610. [Google Scholar] [CrossRef]
- Rosenow, D.T.; Quisenberry, J.E.; Wendt, C.W.; Clark, L.E. Drought tolerant sorghum and cotton germplasm. Agric. Water Manag. 1983, 7, 207–222. [Google Scholar] [CrossRef]
- Doggett, H. Sorghum, 2nd ed.; Longman Scientific & Technical: New York, NY, USA, 1988. [Google Scholar]
- IPCC. Summary for Policymakers; IPCC: Cambridge, UK, 2007; pp. 7–22. [Google Scholar]
- Nelson, G.C.; Rosegrant, M.; Koo, J.; Robertson, R.; Sulser, T.; Zhu, T.; Msangi, S.; Ringler, C.; Palazzo, A.; Batka, M.; et al. Climate Change. Impact on Agriculture and Costs of Adapation; International Food Policy Research Institure: Washington, DC, USA, 2009; p. 32. [Google Scholar]
- Pola, S. Isolation and culture of suspension cultures of Sorghum bicolor. Int. J. Bioassays 2014, 3. [Google Scholar] [CrossRef]
- Liu, G.; Gilding, E.K.; Godwin, I.D. A robust tissue culture system for sorghum [Sorghum bicolor (L.) Moench. S. Afr. J. Bot. 2015, 98, 157–160. [Google Scholar] [CrossRef]
- Belide, S.; Vanhercke, T.; Petrie, J.R.; Singh, S.P. Robust genetic transformation of sorghum (Sorghum bicolor L.) using differentiating embryogenic callus induced from immature embryos. Plant Methods 2017, 13, 109. [Google Scholar] [CrossRef] [PubMed]
- George, E.F.; Hall, M.A.; De Klerk, G.J. Plant tissue culture procedure-background. In Plant Propagation by Tissue Culture; George, E.F., Hall, M.A., De Klerk, G.J., Eds.; Springer: Dordrecht, The Netherlands, 2008; pp. 1–28. [Google Scholar]
- Evans, D.E.; Coleman, J.O.D.; Kearns, A. Plant Cell Culture; BIOS Scientific Publishers: London, UK, 2003. [Google Scholar]
- Ikeuchi, M.; Sugimoto, K.; Iwase, A. Plant callus: Mechanisms of induction and repression. Plant Cell 2013, 25, 3159–3173. [Google Scholar] [CrossRef] [PubMed]
- Collin, H.A.; Edwards, S. Plant Cell Culture; BIOS Scientific Publishers: Oxford, UK, 1998. [Google Scholar]
- Frank, M.; Rupp, H.M.; Prinsen, E.; Motyka, V.; Van Onckelen, H.; Schmulling, T. Hormone autotrophic growth and differentiation identifies mutant lines of Arabidopsis with altered cytokinin and auxin content or signaling. Plant Physiol. 2000, 122, 721–729. [Google Scholar] [CrossRef] [PubMed]
- Chourey, P.S.; Sharpe, D.Z. Callus formation from protoplasys of Sorghum cell suspension cultures. Plant Sci. 1985, 39, 171–175. [Google Scholar] [CrossRef]
- Cai, T.; Daly, B.; Butler, L. Callus induction and plant regeneration from shoot portions of mature embryos of high tannin sorghums. Plant Cell Tissue Organ Cult. 1987, 9, 245–252. [Google Scholar] [CrossRef]
- Anjum, S.A.; Xie, X.-Y.; Wang, L.C.; Saleem, M.F.; Man, C.; Lei, W. Morphological, physiological and biochemical responses of plants to drought stress. Afr. J. Agric. Res. 2011, 6, 2026–2032. [Google Scholar]
- Ngara, R.; Ramulifho, E.; Movahedi, M.; Shargie, N.G.; Brown, A.P.; Chivasa, S. Identifying differentially expressed proteins in sorghum cell cultures exposed to osmotic stress. Sci. Rep. 2018, 8, 8671. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.-L.; Lee, P.-D.; Liu, L.-F.; Su, J.-C. Effect of sorbitol induced osmotic stress on the changes of carbohydrate and free amino acid pools in sweet potato cell suspension cultures. Bot. Bull. Acad. Sin. 1999, 40, 219–225. [Google Scholar]
- Monetti, E.; Kadono, T.; Tran, D.; Azzarello, E.; Arbelet-Bonnin, D.; Biligui, B.; Briand, J.; Kawano, T.; Mancuso, S.; Bouteau, F. Deciphering early events involved in hyperosmotic stress-induced programmed cell death in tobacco BY-2 cells. J. Exp. Bot. 2014, 65, 1361–1375. [Google Scholar] [CrossRef] [Green Version]
- Gupta, P.; Sharma, S.; Saxena, S. Biomass yield and steviol glycoside production in callus and suspension culture of Stevia rebaudiana treated with proline and polyethylene glycol. Appl. Biochem. Biotechnol. 2015, 176, 863–874. [Google Scholar] [CrossRef] [PubMed]
- Patade, V.Y.; Bhargava, S.; Suprasanna, P. Effects of NaCl and iso-osmotic PEG stress on growth, osmolytes accumulation and antioxidant defense in cultured sugarcane cells. Plant Cell Tissue Organ Cult. 2012, 108, 279–286. [Google Scholar] [CrossRef]
- Muilu-Mäkelä, R.; Vuosku, J.; Hamberg, L.; Latva-Mäenpää, H.; Häggman, H.; Sarjala, T. Osmotic stress affects polyamine homeostasis and phenolic content in proembryogenic liquid cell cultures of scots pine. Plant Cell Tissue Organ Cult. 2015, 122, 709–726. [Google Scholar] [CrossRef]
- Wang, L.; He, J.; Ding, H.; Liu, H.; Lu, B.; Liang, J.; Wang, L.; He, J.; Ding, H.D.; Liu, H.; et al. Overexpression of AT14A confers tolerance to drought stress-induced oxidative damage in suspension cultured cells of Arabidopsis thaliana. Protoplasma 2015, 252, 1111–1120. [Google Scholar] [CrossRef]
- Alqurashi, M.; Chiapello, M.; Bianchet, C.; Paolocci, F.; Lilley, K.S.; Gehring, C. Early responses to severe drought stress in the Arabidopsis thaliana cell suspension culture proteome. Proteomes 2018, 6, 38. [Google Scholar] [CrossRef]
- Ngara, R.; Rees, J.; Ndimba, B.K. Establishment of sorghum cell suspension culture system for proteomics studies. Afr. J. Biotechnol. 2008, 7, 744–749. [Google Scholar]
- Ngara, R. A Proteomic Analysis of Drought and Salt Stress Responsive Proteins of Different Sorghum Varieties. Ph.D. Thesis, University of the Western Cape, Cape Town, South Africa, 2009. [Google Scholar]
- Murashige, T.; Skoog, F. A revised medium for rapid growth and bio assays with tobacco tissue cultures. Physiol. Plant. 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Ngara, R.; Ndimba, B.K. Mapping and characterisation of the sorghum cell suspension culture secretome. Afr. J. Biotechnol. 2011, 10, 253–266. [Google Scholar]
- Goche, T. Characterisation of the Physiological, Biochemical and Molecular Responses of Sorghum to Drought Stress. Ph.D. Thesis, University of the Free State, Bloemfontein, South Africa, 2018. [Google Scholar]
- Johnson, S.M.; Lim, F.L.; Finkler, A.; Fromm, H.; Slabas, A.R.; Knight, M.R. Transcriptomic analysis of Sorghum bicolor responding to combined heat and drought stress. BMC Genom. 2014, 15, 456. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Fan, F.; Wang, L.; Zhan, Q.; Wu, P.; Du, J.; Yang, X.; Liu, Y. Cloning and expression analysis of cinnamoyl-CoA reductase (CCR) genes in sorghum. PeerJ 2016, 4, e2005. [Google Scholar] [CrossRef] [PubMed]
- Ye, J.; Coulouris, G.; Zaretskaya, I.; Cutcutache, I.; Rozen, S.; Madden, T.L. Primer-blast: A tool to design target-specific primers for polymerase chain reaction. BMC Bioinform. 2012, 13, 134. [Google Scholar] [CrossRef]
- Cai, T.; Butler, L. Plant regeneration from embryogenic callus initiated from immature inflorescences of several high-tannin sorghums. Plant Cell Tissue Organ Cult. 1990, 20, 101–110. [Google Scholar] [CrossRef]
- Sai Kishore, N.; Visarada, K.B.; Aravinda Lakshmi, Y.; Pashupatinath, E.; Rao, S.V.; Seetharama, N. In vitro culture methods in sorghum with shoot tip as the explant material. Plant Cell Rep. 2006, 25, 174–182. [Google Scholar]
- Pola, S.; Mani, N.S.; Ramana, T. Plant tissue culture studies in Sorghum bicolor: Immature embryo explants as the source material. Int. J. Plant Prod. 2008, 2, 1–14. [Google Scholar]
- Rao, R.V.R.; Kumar, G.P.; Rao, M.V.S.; Manga, V. Differential in vitro response of genotypes of sorghum. Phytomorphology 2000, 50, 253–260. [Google Scholar]
- Gupta, S.; Khanna, V.K.; Singh, R.; Garg, G.K. Strategies for overcoming genotypic limitations of in vitro regeneration and determination of genetic components of variability of plant regeneration traits in sorghum. Plant Cell Tissue Organ Cult. 2006, 86, 379–388. [Google Scholar] [CrossRef]
- Shafi, A.; Gill, T.; Sreenivasulu, Y.; Kumar, S.; Ahuja, P.S.; Singh, A.K. Improved callus induction, shoot regeneration, and salt stress tolerance in Arabidopsis overexpressing superoxide dismutase from Potentilla atrosanguinea. Protoplasma 2015, 252, 41–51. [Google Scholar] [CrossRef]
- Chawla, H.S. Introduction to Plant Biotechnology; Science Publishers: Enfield, NH, USA, 2009. [Google Scholar]
- George, E.F.; Hall, M.A.; De Klerk, G.J. Stock plant physiological factors affecting growth and morphogenesis. In Plant Propagation by Tissue Culture; George, E.F., Hall, M.A., De Klerk, G., Eds.; Springer: Dordrecht, The Netherlands, 2008; pp. 403–422. [Google Scholar]
- Gupta, S.; Wardhan, V.; Verma, S.; Gayali, S.; Rajamani, U.; Datta, A.; Chakraborty, S.; Chakraborty, N. Characterization of the secretome of chickpea suspension culture reveals pathway abundance and the expected and unexpected secreted proteins. J. Proteome Res. 2011, 10, 5006–5015. [Google Scholar] [CrossRef] [PubMed]
- Rahman, M.S.; Miah, M.A.B.; Hossain, M.S.; Kabir, A.H.; Rahman, M.M. Establishment of cell suspension culture and plant regeneration in Abrus precatorius L., a rare medicinal plant. Not. Sci. Biol. 2012, 4, 86–93. [Google Scholar] [CrossRef]
- Angius, F.; Floris, A. Liposomes and MTT cell viability assay: An incompatible affair. Toxicol. In Vitro 2015, 29, 314–319. [Google Scholar] [CrossRef] [PubMed]
- Baker, C.J.; Mock, N.M. An improved method for monitoring cell death in cell suspension and leaf disc assays using evans blue. Plant Cell Tissue Organ Cult. 1994, 39, 7–12. [Google Scholar] [CrossRef]
- Ramos, L.Y.S.; Carballo, L.M.; Melara, M.V. Establishment of cell suspension cultures of two Costa Rican Jatropha species (Euphorbiaceae). Rev. Biol. Trop. 2013, 61, 1095–1107. [Google Scholar]
- Ndimba, B.K.; Chivasa, S.; Hamilton, J.M.; Simon, W.J.; Slabas, A.R. Proteomic analysis of changes in the extracellular matrix of Arabidopsis cell suspension cultures induced by fungal elicitors. Proteomics 2003, 3, 1047–1059. [Google Scholar] [CrossRef] [PubMed]
- Levitt, J. Responses of Plants to Environmental Stresses, 2nd ed.; Academic Press: New York, NY, USA, 1980; Volume II. [Google Scholar]
- Bray, E.A. Molecular responses to water deficit. Plant Physiol. 1993, 103, 1035–1040. [Google Scholar] [CrossRef]
- Fracasso, A.; Trindade, L.M.; Amaducci, S. Drought stress tolerance strategies revealed by RNA-seq in two sorghum genotypes with contrasting WUE. BMC Plant Biol. 2016, 16, 115. [Google Scholar] [CrossRef]
- Bray, E.A. Plant responses to water deficit. Trends Plant Sci. 1997, 2, 48–54. [Google Scholar] [CrossRef]
- Shinozaki, K.; Yamaguchi-Shinozaki, K. Molecular responses to drought and cold stress. Curr. Opin. Biotechnol. 1996, 7, 161–167. [Google Scholar] [CrossRef]
- Shinozaki, K.; Yamaguchi-Shinozaki, K. Gene expression and signal transduction in water-stress response. Plant Physiol. 1997, 115, 327–334. [Google Scholar] [CrossRef] [PubMed]
- May, M.J.; Leaver, C.J. Oxidative stimulation of glutathione synthesis in Arabidopsis thaliana suspension cultures. Plant Physiol. 1993, 103, 621–627. [Google Scholar] [CrossRef] [PubMed]
- Levine, A.; Tenhaken, R.; Dixon, R.; Lamb, C. H2O2 from the oxidative burst orchestrates the plant hypersensitive disease resistance response. Cell 1994, 79, 583–593. [Google Scholar] [CrossRef]
- Delledonne, M.; Zeier, J.; Marocco, A.; Lamb, C. Signal interactions between nitric oxide and reactive oxygen intermediates in the plant hypersensitive disease resistance response. Proc. Natl. Acad. Sci. USA 2001, 98, 13454–13459. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rajasekhar, V.K.; Lamb, C.; Dixon, R.A. Early events in the signal pathway for the oxidative burst in soybean cells exposed to avirulent Pseudomonas syringae pv glycinea. Plant Physiol. 1999, 120, 1137–1146. [Google Scholar] [CrossRef] [PubMed]
- Ndimba, B.K.; Chivasa, S.; Simon, W.J.; Slabas, A.R. Identification of Arabidopsis salt and osmotic stress responsive proteins using two-dimensional difference gel electrophoresis and mass spectrometry. Proteomics 2005, 5, 4185–4196. [Google Scholar] [CrossRef]
- Chivasa, S.; Hamilton, J.M.; Pringle, R.S.; Ndimba, B.K.; Simon, W.J.; Lindsey, K.; Slabas, A.R. Proteomic analysis of differentially expressed proteins in fungal elicitor-treated Arabidopsis cell cultures. J. Exp. Bot. 2006, 57, 1553–1562. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaffarnik, F.A.; Jones, A.M.; Rathjen, J.P.; Peck, S.C. Effector proteins of the bacterial pathogen Pseudomonas syringae alter the extracellular proteome of the host plant, Arabidopsis thaliana. Mol. Cell. Proteom. 2009, 8, 145–156. [Google Scholar] [CrossRef] [PubMed]
- Oh, I.S.; Park, A.R.; Bae, M.S.; Kwon, S.J.; Kim, Y.S.; Lee, J.E.; Kang, N.Y.; Lee, S.; Cheong, H.; Park, O.K. Secretome analysis reveals an Arabidopsis lipase involved in defense against Alternaria brassicicola. Plant Cell 2005, 17, 2832–2847. [Google Scholar] [CrossRef] [PubMed]
- Ngcala, M.G. Molecular Responses of Sorghum Cell Suspension Cultures to High Temperature Stress. Master’s Thesis, University of the Free State, Bloemfontein, South Africa, 2018. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target Gene ID a | Forward Primer | Reverse Primer |
|---|---|---|
| SORBI_3008G134700 | 5′-CCCATCCACTGGAACGAGAC-3′ | 5′-ACTGCGTCCAGATGAGGTTG-3′ |
| SORBI_3001G514200 | 5′-GGAACATCCTGGCGCATTTG-3′ | 5′-AGAGCCAGACCGACACAAAC-3′ |
| SORBI_3003G136200 | 5′-TGCCACGCACAGAATACGAG-3′ | 5′-TACACATGCCGGAGAATCGG-3′ |
| SORBI_3006G135500 | 5′-AGGGAAATGGTTGTACGGGC-3′ | 5′-GTGCTGGTGCACAAAATATATAGC-3′ |
| SORBI_3007G149600 | 5′-TGCTCTCGTGAGATCGCTTG-3′ | 5′-GCAATACCACTGCGTATCTGTTG-3′ |
| SORBI_3001G313200 | 5′-GGATCCCTGGATCTTCTGGAG-3′ | 5′-TTTGGTGGCTCTGTGGTCAG-3′ |
| SORBI_3001G073900 | 5′-CTTCTTCAACACACGCGACC-3′ | 5′-GTGTGGTGACGGATGGTAGG-3′ |
| SORBI_3003G322400 | 5′-AACGAGTATATGCATCCCCG-3′ | 5′-AGCAGGCTCTAGGGTTTTGAG-3′ |
| Sb04g003390 * | 5′-GATGAGATGCTCTCCCGTGG-3′ | 5′-TGATCTCTAGGGCCTCTGGG-3′ |
| Sb03g038910 * | 5′-TCCTGAAGCATCTTTCCCTCC-3′ | 5′-ACAGCCTGATTAGTTGGGGG-3′ |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ramulifho, E.; Goche, T.; Van As, J.; Tsilo, T.J.; Chivasa, S.; Ngara, R. Establishment and Characterization of Callus and Cell Suspension Cultures of Selected Sorghum bicolor (L.) Moench Varieties: A Resource for Gene Discovery in Plant Stress Biology. Agronomy 2019, 9, 218. https://doi.org/10.3390/agronomy9050218
Ramulifho E, Goche T, Van As J, Tsilo TJ, Chivasa S, Ngara R. Establishment and Characterization of Callus and Cell Suspension Cultures of Selected Sorghum bicolor (L.) Moench Varieties: A Resource for Gene Discovery in Plant Stress Biology. Agronomy. 2019; 9(5):218. https://doi.org/10.3390/agronomy9050218
Chicago/Turabian StyleRamulifho, Elelwani, Tatenda Goche, Johann Van As, Toi John Tsilo, Stephen Chivasa, and Rudo Ngara. 2019. "Establishment and Characterization of Callus and Cell Suspension Cultures of Selected Sorghum bicolor (L.) Moench Varieties: A Resource for Gene Discovery in Plant Stress Biology" Agronomy 9, no. 5: 218. https://doi.org/10.3390/agronomy9050218