Soil Amendment with Raw Garlic Stalk: A Novel Strategy to Stimulate Growth and the Antioxidative Defense System in Monocropped Eggplant in the North of China


 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Site Description
2.2. Preparation of Raw Garlic Stalk (RGS)
2.3. Plant Bioassays
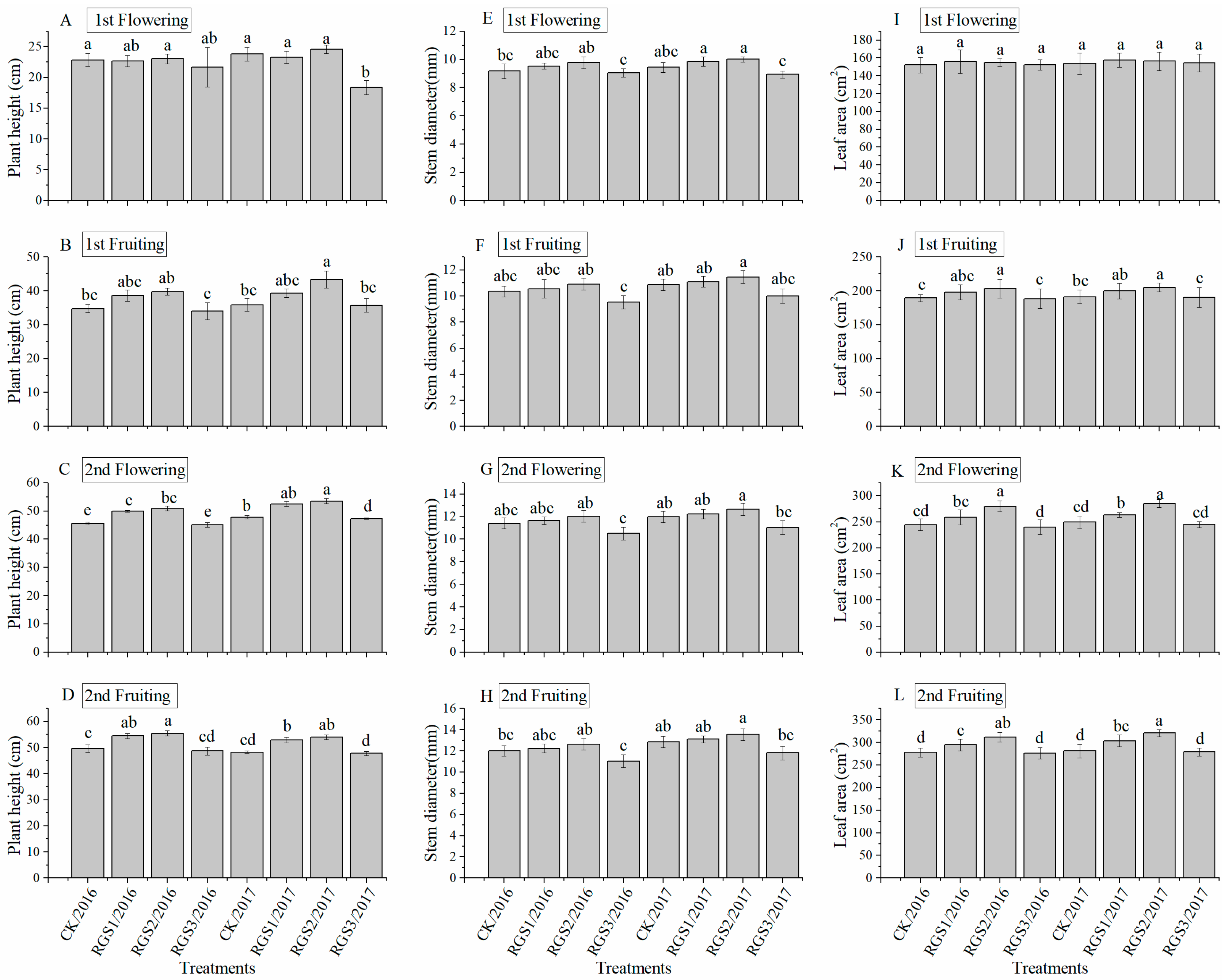
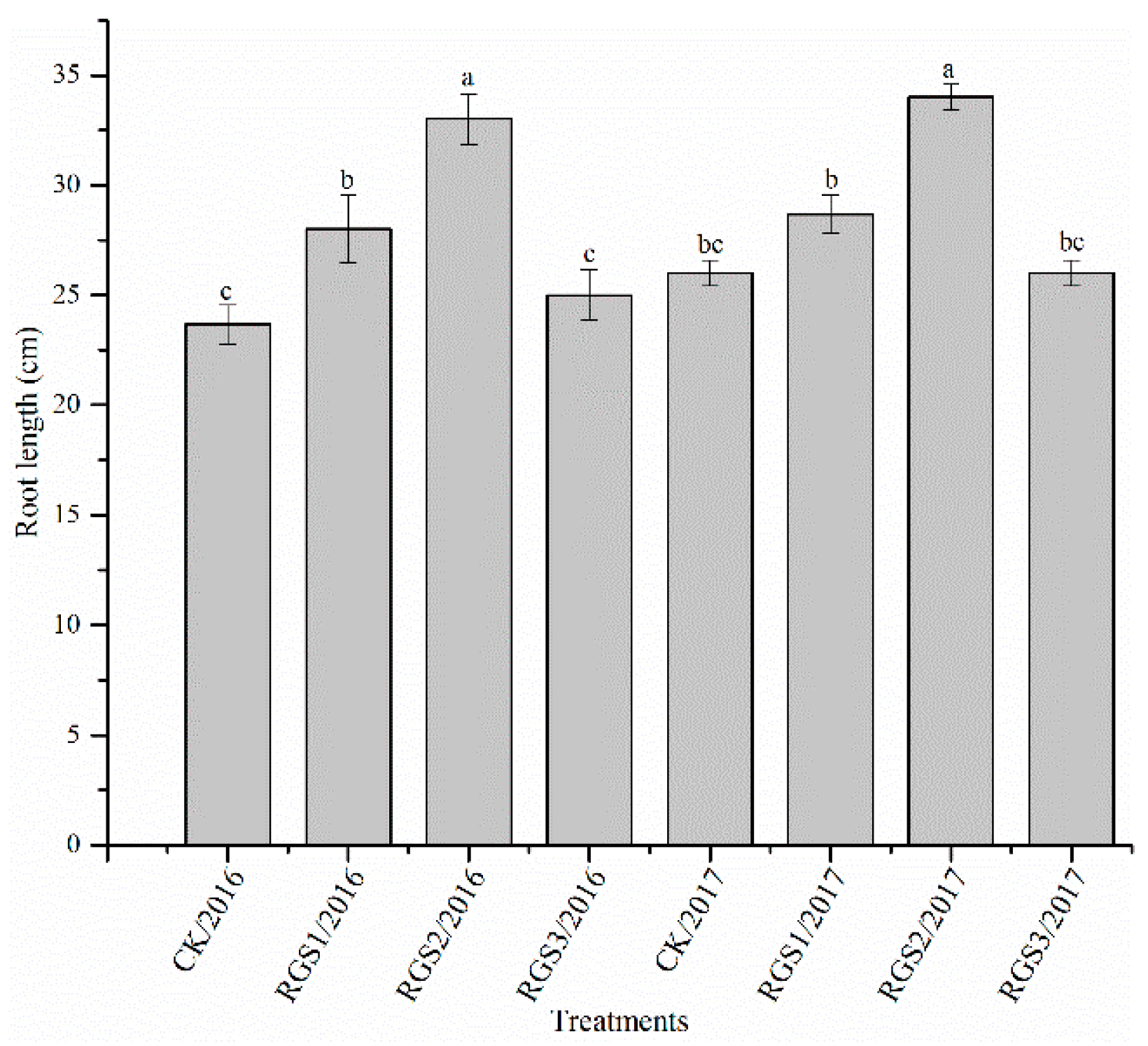
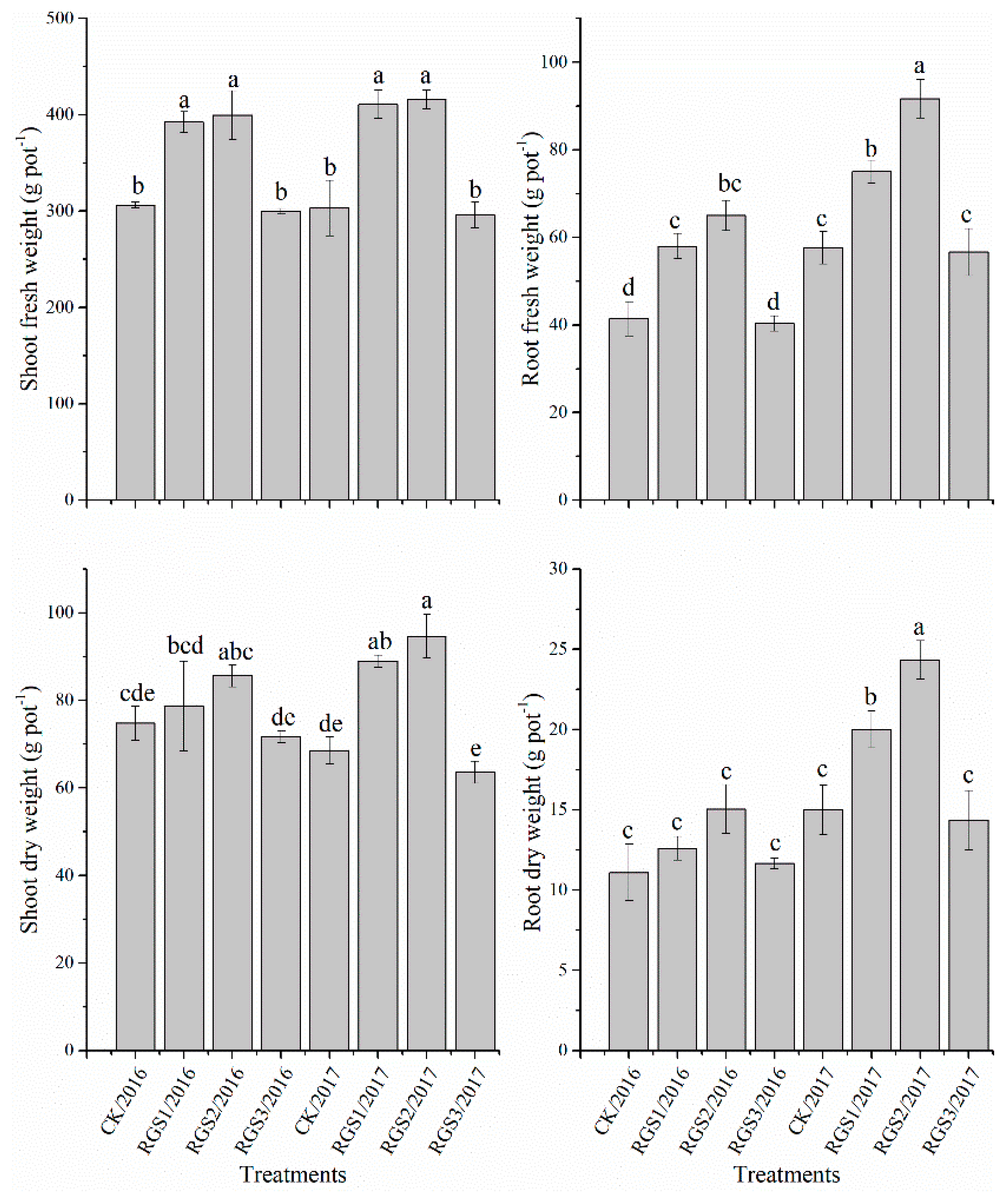
2.4. Measurement of Growth Parameters Based on Morphology
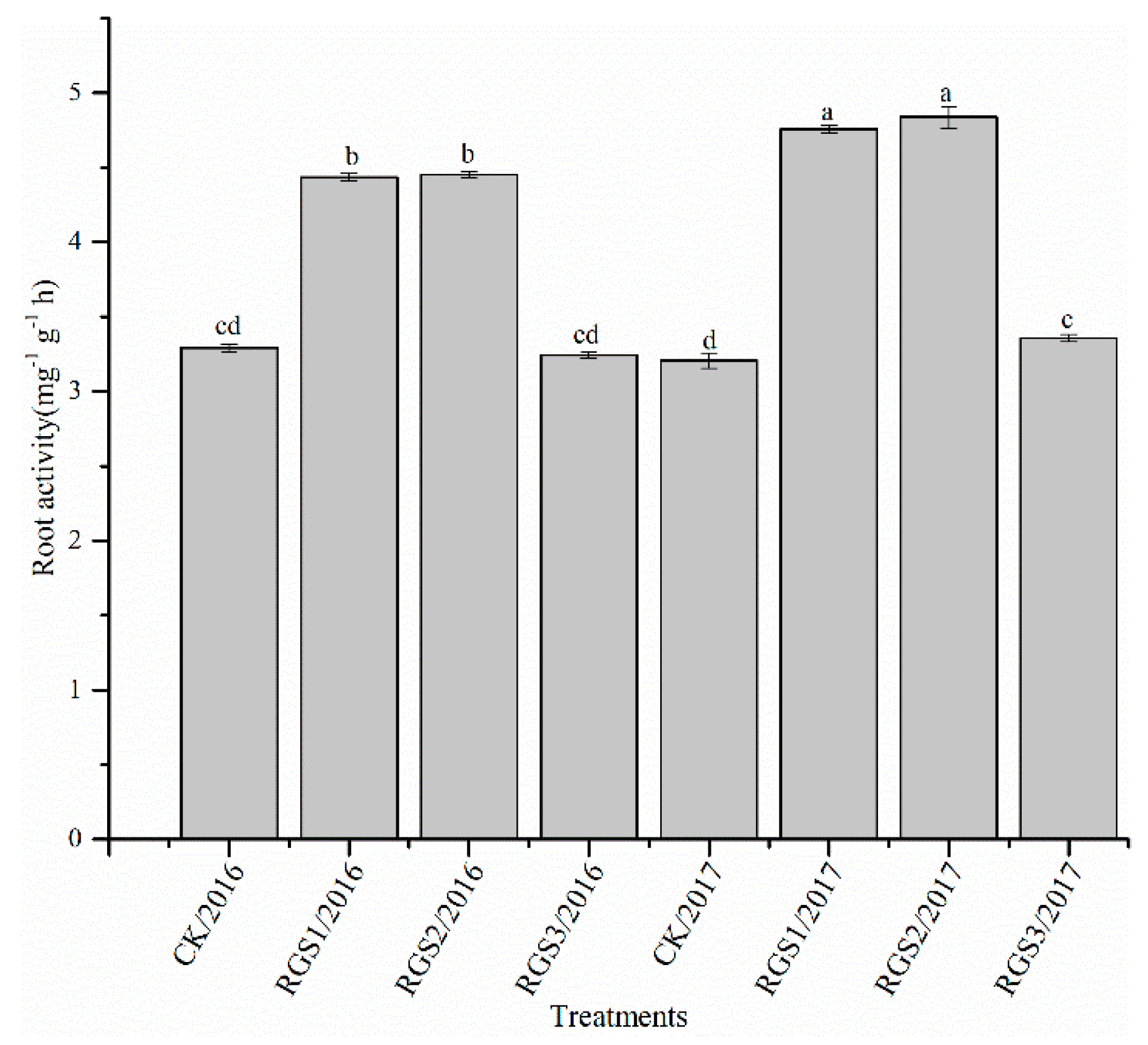
2.5. Measurement of Root Activity
2.6. Determination of Physiological Indices
2.6.1. Chlorophyll and Carotenoid Contents
2.6.2. Estimation of Plant Gas Exchange Parameters
2.7. Preparation of Leaf Extract for Malondialdehyde (MDA) and Antioxidative Enzymes Activity Assays
2.7.1. Quantification of H2O2 and MDA Contents
2.7.2. Antioxidative Enzyme Assays
2.8. Estimation of the Fusarium wilt Disease Control Index and Eggplant Yield
2.9. Statistical Analysis
3. Results
3.1. Effect of RGS on Morphological Growth Parameters and Root Activity
3.2. Effect of RGS on Chlorophyll Contents and Carotenoids in Leaves
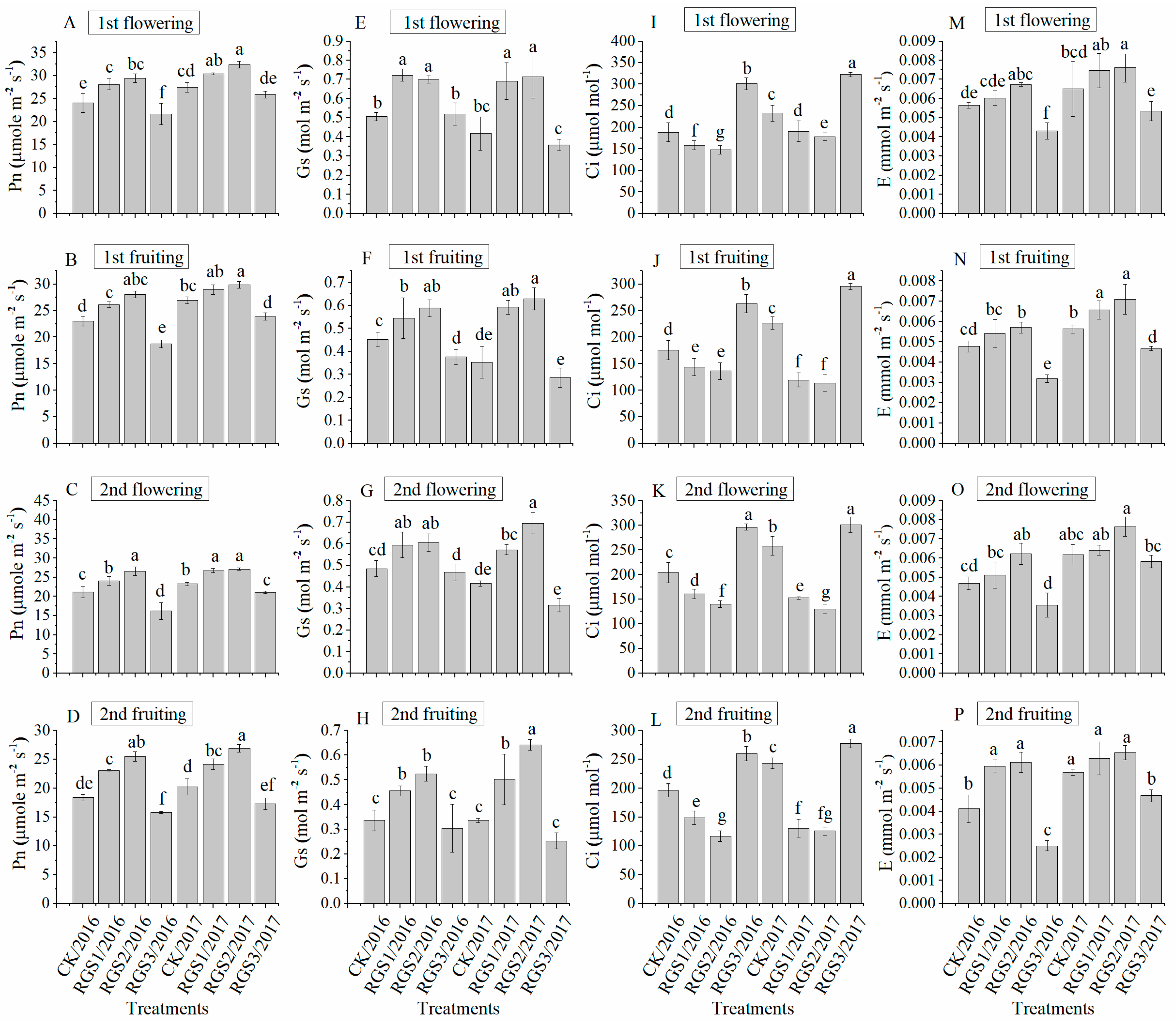
3.3. Effect of RGS on Gas Exchange Parameters
3.4. Effect of RGS on H2O2 and MDA Contents
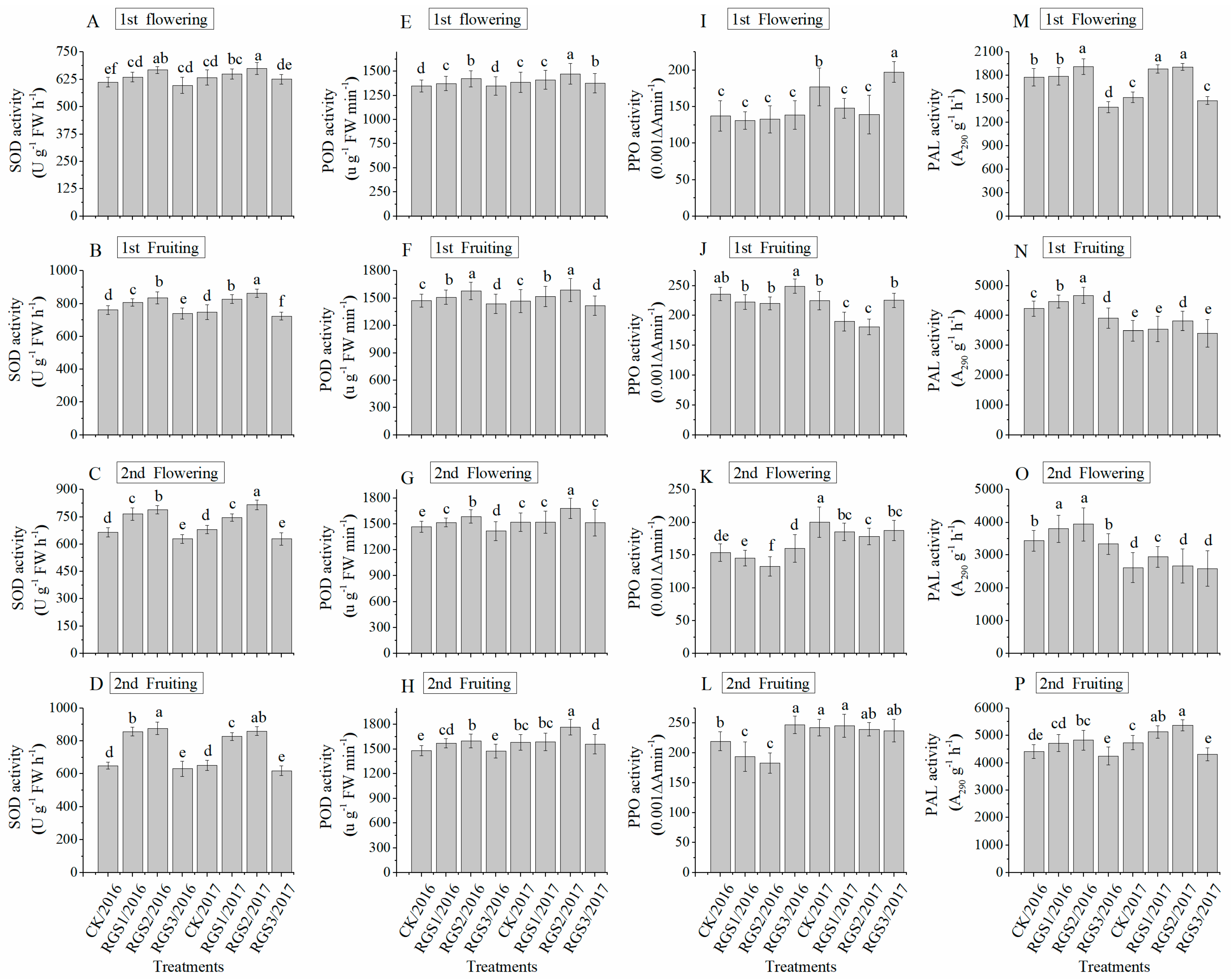
3.5. Antioxidants and Plant Enzyme Activities
3.6. Effect of Different RGS Rates on Fusarium wilt and Yield of Eggplant
4. Discussion
4.1. Effect of RGS on Plant Growth
4.2. RGS as an Antioxidative Defense Regulator
4.3. Effect of RGS on Fusarium wilt and Yield of Eggplant
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Sarrantonio, M.; Gallandt, E. The role of cover crops in North American cropping systems. J. Crop Prod. 2003, 8, 53–74. [Google Scholar] [CrossRef]
- Gupta, R.; Ladha, J. Placement effects on rice residue decomposition and nutrient dynamics on two soil types during wheat cropping in rice-wheat system in northwestern India. Nutr. Cycl. Agroecosyst. 2010, 88, 471–480. [Google Scholar]
- Ilieva-Makulec, K.; Olejniczak, I.; Szanser, M. Response of soil micro-and mesofauna to diversity and quality of plant litter. Eur. J. Soil Biol. 2006, 42, S244–S249. [Google Scholar] [CrossRef]
- DuPont, S.T.; Ferris, H.; Van Horn, M. Effects of cover crop quality and quantity on nematode-based soil food webs and nutrient cycling. Appl. Soil Ecol. 2009, 41, 157–167. [Google Scholar] [CrossRef]
- Tian, Y.; Liu, J.; Zhang, X.; Gao, L. Effects of summer catch crop, residue management, soil temperature and water on the succeeding cucumber rhizosphere nitrogen mineralization in intensive production systems. Nutr. Cycl. Agroecosyst. 2010, 88, 429–446. [Google Scholar] [CrossRef]
- Xiao, X.; Cheng, Z.; Meng, H.; Khan, M.A.; Li, H. Intercropping with garlic alleviated continuous cropping obstacle of cucumber in plastic tunnel. Acta Agric. Scand. B Soil Plant Sci. 2012, 62, 696–705. [Google Scholar] [CrossRef]
- Wang, M.; Wu, C.; Cheng, Z.; Meng, H. Growth and physiological changes in continuously cropped eggplant (Solanum melongena L.) upon relay intercropping with garlic (Allium sativum L.). Front. Plant Sci. 2015, 6, 262. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Zhou, B.; Lin, S.; Li, X.; Ye, X. Accumulation of cinnamic acid and vanillin in eggplant root exudates and the relationship with continuous cropping obstacle. Afr. J. Biotechnol. 2011, 10, 2659–2665. [Google Scholar]
- Safikhani, N.; Morid, B.; Zamanizadeh, H. First report of Fusarium wilt of eggplant caused by Fusarium oxysporum f. sp. melongenae in Iran. New Dis. Rep. 2013, 28. [Google Scholar] [CrossRef]
- Asada, K. Production and scavenging of reactive oxygen species in chloroplasts and their functions. Plant Physiol. 2006, 141, 391–396. [Google Scholar] [CrossRef]
- Sharma, P.; Jha, A.B.; Dubey, R.S.; Pessarakli, M. Reactive oxygen species, oxidative damage, and antioxidative defense mechanism in plants under stressful conditions. J. Bot. 2012, 2012. [Google Scholar] [CrossRef]
- Noctor, G.; Veljovic-Jovanovic, S.; Driscoll, S.; Novitskaya, L.; Foyer, C.H. Drought and oxidative load in the leaves of C3 plants: A predominant role for photorespiration? Ann. Bot. 2002, 89, 841–850. [Google Scholar] [CrossRef] [PubMed]
- Tanou, G.; Molassiotis, A.; Diamantidis, G. Hydrogen peroxide-and nitric oxide-induced systemic antioxidant prime-like activity under NaCl-stress and stress-free conditions in citrus plants. J. Plant Physiol. 2009, 166, 1904–1913. [Google Scholar] [CrossRef] [PubMed]
- Bright, J.; Desikan, R.; Hancock, J.T.; Weir, I.S.; Neill, S.J. ABA-induced NO generation and stomatal closure in Arabidopsis are dependent on H2O2 synthesis. Plant J. 2006, 45, 113–122. [Google Scholar] [CrossRef] [PubMed]
- Peng, C.; Ou, Z.; Liu, N.; Lin, G. Response to high temperature in flag leaves of super high-yielding rice Pei’ai 64S/E32 and Liangyoupeijiu. Rice Sci. 2005, 12, 179–186. [Google Scholar]
- Yin, X.M.; Huang, L.F.; Zhang, X.; Wang, M.L.; Xu, G.Y.; Xia, X.J. OsCML4 improves drought tolerance through scavenging of reactive oxygen species in rice. J. Plant Biol. 2015, 58, 68–73. [Google Scholar] [CrossRef]
- Suleman, P.; Redha, A.; Afzal, M.; Al-Hasan, R. Temperature-induced changes of malondialdehyde, heat-shock proteins in relation to chlorophyll fluorescence and photosynthesis in Conocarpus lancifolius (Engl.). Acta Physiol. Plant. 2013, 35, 1223–1231. [Google Scholar] [CrossRef]
- Meloni, D.A.; Oliva, M.A.; Martinez, C.A.; Cambraia, J. Photosynthesis and activity of superoxide dismutase, peroxidase and glutathione reductase in cotton under salt stress. Environ. Exp. Bot. 2003, 49, 69–76. [Google Scholar] [CrossRef]
- Choi, I.-H.; Shin, S.-C.; Park, I.-K. Nematicidal activity of onion (Allium cepa) oil and its components against the pine wood nematode (Bursaphelenchus xylophilus). Nematology 2007, 9, 231–235. [Google Scholar]
- Wang, M.; Wu, C.; Cheng, Z.; Meng, H.; Zhang, M.; Zhang, H. Soil chemical property changes in eggplant/garlic relay intercropping systems under continuous cropping. PLoS ONE 2014, 9, e111040. [Google Scholar] [CrossRef]
- Cheng, Z.-H.; Jin, R.; Tong, F. Study on Allelopathic Effect of Garlic Plant Volatile on Cucumber Seedling Growth. Acta Agric. Boreali-Occident. Sin. 2007, 3, 037. [Google Scholar]
- Abràmoff, M.D.; Magalhães, P.J.; Ram, S.J. Image processing with ImageJ. Biophotonics Int. 2004, 11, 36–42. [Google Scholar]
- Lariguet, P.; Ranocha, P.; De Meyer, M.; Barbier, O.; Penel, C.; Dunand, C. Identification of a hydrogen peroxide signalling pathway in the control of light-dependent germination in Arabidopsis. Planta 2013, 238, 381–395. [Google Scholar] [CrossRef] [PubMed]
- Arnon, D.I. Copper enzymes in isolated chloroplasts. Polyphenoloxidase in Beta vulgaris. Plant Physiol. 1949, 24, 1. [Google Scholar] [CrossRef]
- Distefano, G.; La Malfa, S.; Vitale, A.; Lorito, M.; Deng, Z.; Gentile, A. Defence-related gene expression in transgenic lemon plants producing an antimicrobial Trichoderma harzianum endochitinase during fungal infection. Transgenic Res. 2008, 17, 873–879. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Qu, W. The Guidance of Plant Physiology Experiments; Chinese Agricultural Science and Technology Press: Beijing, China, 2004. [Google Scholar]
- Gao, J. Experimental Guidance for Plant Physiology; China Higher Education Press: Beijing, China, 2006. [Google Scholar]
- Bestwick, C.S.; Brown, I.R.; Mansfield, J.W. Localized changes in peroxidase activity accompany hydrogen peroxide generation during the development of a nonhost hypersensitive reaction in lettuce. Plant Physiol. 1998, 118, 1067–1078. [Google Scholar] [CrossRef] [PubMed]
- Zheng, L.; Zhong, G.; Zhang, S. PPO activity determination and anti-browning measure for amorphophallus albus. Chin. J. Southwest Univ. (Nat. Sci. Ed.) 2007, 29, 118–121. [Google Scholar]
- Wang, W.; Wang, H.; Feng, Y.; Wang, L.; Xiao, X.; Xi, Y.; Luo, X.; Sun, R.; Ye, X.; Huang, Y. Consistent responses of the microbial community structure to organic farming along the middle and lower reaches of the Yangtze River. Sci. Rep. 2016, 6, 35046. [Google Scholar] [CrossRef] [Green Version]
- Corzo-Martínez, M.; Corzo, N.; Villamiel, M. Biological properties of onions and garlic. Trends Food Sci. Technol. 2007, 18, 609–625. [Google Scholar] [CrossRef]
- Hafez, O.M.; Saleh, M.A.; El-Lethy, S.R. Response of some seedlings olive cultivars to foliar spray of yeast and garlic extracts with or without vascular arbuscular mycorrhizal fungi. J. World Appl. Sci. 2013, 24, 1119–1129. [Google Scholar]
- Lanzotti, V. The analysis of onion and garlic. J. Chromatogr. A 2006, 1112, 3–22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gruhlke, M.C.; Hemmis, B.; Noll, U.; Wagner, R.; Lühring, H.; Slusarenko, A.J. The defense substance allicin from garlic permeabilizes membranes of Beta vulgaris, Rhoeo discolor, Chara corallina and artificial lipid bilayers. Biochim. Biophys. Acta (BBA)-Gen. Subj. 2015, 1850, 602–611. [Google Scholar] [CrossRef] [PubMed]
- Oracz, K.; Voegele, A.; Tarkowská, D.; Jacquemoud, D.; Turečková, V.; Urbanová, T.; Strnad, M.; Sliwinska, E.; Leubner-Metzger, G. Myrigalone A inhibits Lepidium sativum seed germination by interference with gibberellin metabolism and apoplastic superoxide production required for embryo extension growth and endosperm rupture. Plant Cell Physiol. 2011, 53, 81–95. [Google Scholar] [CrossRef] [PubMed]
- Han, X.; Cheng, Z.; Meng, H.; Yang, X.; Ahmad, I. Allelopathic effect of decomposed garlic (Allium sativum L.) stalk on lettuce (L. sativa var. crispa L.). Pak. J. Bot. 2013, 45, 225–233. [Google Scholar]
- Masclaux, C.; Valadier, M.-H.; Brugière, N.; Morot-Gaudry, J.-F.; Hirel, B. Characterization of the sink/source transition in tobacco (Nicotiana tabacum L.) shoots in relation to nitrogen management and leaf senescence. Planta 2000, 211, 510–518. [Google Scholar] [CrossRef] [PubMed]
- Borlinghaus, J.; Albrecht, F.; Gruhlke, M.C.; Nwachukwu, I.D.; Slusarenko, A.J. Allicin: Chemistry and biological properties. Molecules 2014, 19, 12591–12618. [Google Scholar] [CrossRef] [PubMed]
- Cheng, F.; Cheng, Z.; Meng, H.; Tang, X. The garlic allelochemical diallyl disulfide affects tomato root growth by influencing cell division, phytohormone balance and expansin gene expression. Front. Plant Sci. 2016, 7, 1199. [Google Scholar] [CrossRef]
- Yu, B.; Zhang, D.; Huang, C.; Qian, M.; Zheng, X.; Teng, Y.; Su, J.; Shu, Q. Isolation of anthocyanin biosynthetic genes in red Chinese sand pear (Pyrus pyrifolia Nakai) and their expression as affected by organ/tissue, cultivar, bagging and fruit side. Sci. Hortic. 2012, 136, 29–37. [Google Scholar] [CrossRef]
- Yoshii, H.; Yoshii, Y.; Asai, T.; Furukawa, T.; Takaichi, S.; Fujibayashi, Y. Photo-excitation of carotenoids causes cytotoxicity via singlet oxygen production. Biochem. Biophys. Res. Commun. 2012, 417, 640–645. [Google Scholar] [CrossRef] [Green Version]
- Krasylenko, Y.A.; Yemets, A.I.; Sheremet, Y.A.; Blume, Y.B. Nitric oxide as a critical factor for perception of UV-B irradiation by microtubules in Arabidopsis. Physiol. Plant. 2012, 145, 505–515. [Google Scholar] [CrossRef]
- Yang, Y.; Han, X.; Liang, Y.; Ghosh, A.; Chen, J.; Tang, M. The combined effects of arbuscular mycorrhizal fungi (AMF) and lead (Pb) stress on Pb accumulation, plant growth parameters, photosynthesis, and antioxidant enzymes in Robinia pseudoacacia L. PLoS ONE 2015, 10, e0145726. [Google Scholar] [CrossRef] [PubMed]
- Das, K.; Roychoudhury, A. Reactive oxygen species (ROS) and response of antioxidants as ROS-scavengers during environmental stress in plants. Front. Environ. Sci. 2014, 2, 53. [Google Scholar] [CrossRef]
- Liu, Z.; Wang, X. Changes in color, antioxidant, and free radical scavenging enzyme activity of mushrooms under high oxygen modified atmospheres. Postharvest Biol. Technol. 2012, 69, 1–6. [Google Scholar] [CrossRef]
- Rai, A.C.; Singh, M.; Shah, K. Effect of water withdrawal on formation of free radical, proline accumulation and activities of antioxidant enzymes in ZAT12-transformed transgenic tomato plants. Plant Physiol. Biochem. 2012, 61, 108–114. [Google Scholar]
- Hasan, S.A.; Fariduddin, Q.; Ali, B.; Hayat, S.; Ahmad, A. Cadmium: Toxicity and tolerance in plants. J. Environ. Biol. 2009, 30, 165–174. [Google Scholar] [PubMed]
- Li, Y.; Zhang, S.; Jiang, W.; Liu, D. Cadmium accumulation, activities of antioxidant enzymes, and malondialdehyde (MDA) content in Pistia stratiotes L. Environ. Sci. Pollut. Res. 2013, 20, 1117–1123. [Google Scholar] [CrossRef] [PubMed]
- Baxter, A.; Mittler, R.; Suzuki, N. ROS as key players in plant stress signalling. J. Exp. Bot. 2013, 65, 1229–1240. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Del Rio, D.; Stewart, A.J.; Pellegrini, N. A review of recent studies on malondialdehyde as toxic molecule and biological marker of oxidative stress. Nutr. Metab. Cardiovasc. Dis. 2005, 15, 316–328. [Google Scholar] [CrossRef]
- Tommasino, E.; Griffa, S.; Grunberg, K.; Ribotta, A.; Colomba, E.L.; Carloni, E.; Quiroga, M.; Luna, C.M. Malondialdehyde content as a potential biochemical indicator of tolerant Cenchrus ciliaris L. genotypes under heat stress treatment. Grass Forage Sci. 2012, 67, 456–459. [Google Scholar] [CrossRef]
- Huang, G.-T.; Ma, S.-L.; Bai, L.-P.; Zhang, L.; Ma, H.; Jia, P.; Liu, J.; Zhong, M.; Guo, Z.-F. Signal transduction during cold, salt, and drought stresses in plants. Mol. Biol. Rep. 2012, 39, 969–987. [Google Scholar] [CrossRef]
- Gill, S.S.; Tuteja, N. Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol. Biochem. 2010, 48, 909–930. [Google Scholar] [CrossRef]
- Hirt, H.; Shinozaki, K. Plant Responses to Abiotic Stress; Springer Science & Business Media: Berlin, Germany, 2003; Volume 4. [Google Scholar]
- Yiu, J.-C.; Tseng, M.-J. Manipulation of superoxide dismutase and catalase to enhance sulfur dioxide tolerance in transgenic Chinese cabbage. In II International Symposium on Biotechnology of Tropical and Subtropical Species 692; International Society for Horticultural Science: Taipei, Taiwan, 2001; pp. 91–100. [Google Scholar]
- Shafi, A.; Chauhan, R.; Gill, T.; Swarnkar, M.K.; Sreenivasulu, Y.; Kumar, S.; Kumar, N.; Shankar, R.; Ahuja, P.S.; Singh, A.K. Expression of SOD and APX genes positively regulates secondary cell wall biosynthesis and promotes plant growth and yield in Arabidopsis under salt stress. Plant Mol. Biol. 2015, 87, 615–631. [Google Scholar] [CrossRef] [PubMed]
- Tomás-Barberán, F.A.; Espín, J.C. Phenolic compounds and related enzymes as determinants of quality in fruits and vegetables. J. Sci. Food Agric. 2001, 81, 853–876. [Google Scholar] [CrossRef]
- Criado, M.; Civera, M.; Martínez, A.; Rodrigo, D. Use of Weibull distribution to quantify the antioxidant effect of Stevia rebaudiana on oxidative enzymes. LWT-Food Sci. Technol. 2015, 60, 985–989. [Google Scholar] [CrossRef]
- Constabel, C.P.; Barbehenn, R. Defensive roles of polyphenol oxidase in plants. In Induced Plant Resistance to Herbivory; Springer: Berlin, Germany, 2008; pp. 253–270. [Google Scholar]
- Thilagavathi, R.; Saravanakumar, D.; Ragupathi, N.; Samiyappan, R. A combination of biocontrol agents improves the management of dry root rot (Macrophomina phaseolina) in greengram. Phytopathol. Mediterr. 2007, 46, 157–167. [Google Scholar]
- Tarchoune, I.; Sgherri, C.; Izzo, R.; Lachaal, M.; Ouerghi, Z.; Navari-Izzo, F. Antioxidative responses of Ocimum basilicum to sodium chloride or sodium sulphate salinization. Plant Physiol. Biochem. 2010, 48, 772–777. [Google Scholar] [CrossRef]
- Xue, Y.J.; Tao, L.; Yang, Z.M. Aluminum-induced cell wall peroxidase activity and lignin synthesis are differentially regulated by jasmonate and nitric oxide. J. Agric. Food Chem. 2008, 56, 9676–9684. [Google Scholar] [CrossRef] [PubMed]
- Shinshi, H.; Noguchi, M. Relationships between peroxidase, IAA oxidase and polyphenol oxidase. Phytochemistry 1975, 14, 1255–1258. [Google Scholar] [CrossRef]
- Möller, R.; Koch, G.; Nanayakkara, B.; Schmitt, U. Lignification in cell cultures of Pinus radiata: Activities of enzymes and lignin topochemistry. Tree Physiol. 2006, 26, 201–210. [Google Scholar] [CrossRef]
- Chakraborty, M.; Chatterjee, N.; Quimio, T. Integrated management of fusarial wilt of eggplant (Solanum melongena) with soil solarization. Micol. Apl. Int. 2009, 21, 23–26. [Google Scholar]
- Oka, Y. Mechanisms of nematode suppression by organic soil amendments—A review. Appl. Soil Ecol. 2010, 44, 101–115. [Google Scholar] [CrossRef]
- Alabouvette, C.; Olivain, C.; Steinberg, C. Biological control of plant diseases: The European situation. Eur. J. Plant Pathol. 2006, 114, 329–341. [Google Scholar] [CrossRef]
- Gong, B.; Bloszies, S.; Li, X.; Wei, M.; Yang, F.; Shi, Q.; Wang, X. Efficacy of garlic straw application against root-knot nematodes on tomato. Sci. Hortic. 2013, 161, 49–57. [Google Scholar] [CrossRef]
- Sial, T.A.; Khan, M.N.; Lan, Z.; Kumbhar, F.; Zhao, Y.; Zhang, J.; Sun, D.; Xiu, L. Contrasting effects of banana peels waste and its biochar on greenhouse gas emissions and soil biochemical properties. Process Saf. Environ. Prot. 2018, 122, 366–377. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Chlorophyll a (mg g−1 FW) | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Treatment | 1st Flowering | 1st Fruiting | 2nd Flowering | 2nd Fruiting | ||||||||
| 2016 | 2017 | Means | 2016 | 2017 | Means | 2016 | 2017 | Means | 2016 | 2017 | Means | |
| CK | 13.71 e | 15.46 d | 14.59 c | 12.94 | 10.76 | 11.85 c | 10.68 d | 9.96 e | 10.32 c | 9.49 | 8.58 | 9.03 |
| RGS1 | 17.87 c | 20.51 ab | 19.19 b | 15.59 | 13.34 | 14.46 b | 11.01 cd | 11.55 c | 11.28 b | 10.36 | 9.97 | 10.16 |
| RGS2 | 19.55 b | 20.88 a | 20.22 a | 17.06 | 14.19 | 15.62 a | 13.83 a | 12.37 c | 13.10 a | 11.49 | 10.42 | 10.96 |
| RGS3 | 12.97 e | 13.08 e | 13.02 d | 11.85 | 8.99 | 10.42 d | 9.81 e | 9.15 f | 9.48 d | 8.48 | 7.80 | 8.14 |
| Year means | 16.03 | 17.48 | 14.36 | 11.82 | 11.33 | 10.76 | 9.96 | 9.19 | ||||
| LSD-test | Treatment | Year | Interaction | Treatment | Year | Interaction | Treatment | Year | Interaction | Treatment | Year | Interaction |
| *** | *** | ** | *** | *** | NS | *** | *** | *** | NS | *** | NS | |
| Chlorophyll b (mg g−1 FW) | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Treatment | 1st Flowering | 1st Fruiting | 2nd Flowering | 2nd Fruiting | ||||||||
| 2016 | 2017 | Means | 2016 | 2017 | Means | 2016 | 2017 | Means | 2016 | 2017 | Means | |
| CK | 5.54 | 4.70 | 5.12 c | 3.44 | 3.79 | 3.61 b | 3.57 | 2.44 | 3.00 bc | 2.37 | 2.90 | 2.64 |
| RGS1 | 6.06 | 5.78 | 5.92 b | 4.23 | 4.45 | 4.34 a | 3.76 | 2.67 | 3.21 ab | 2.43 | 2.96 | 2.69 |
| RGS2 | 7.74 | 7.38 | 7.57 a | 4.23 | 4.35 | 4.29 a | 3.87 | 2.77 | 3.32 a | 2.58 | 3.27 | 2.93 |
| RGS3 | 5.36 | 4.64 | 5.00 c | 3.39 | 3.55 | 3.47 b | 3.53 | 2.15 | 2.83 c | 2.67 | 2.87 | 2.57 |
| Year means | 6.18 | 5.62 | 3.82 | 4.03 | 3.68 | 2.51 | 2.41 | 3.00 | ||||
| LSD-test | Treatment | Year | Interaction | Treatment | Year | Interaction | Treatment | Year | Interaction | Treatment | Year | Interaction |
| *** | ** | NS | *** | ** | NS | ** | *** | NS | NS | *** | NS | |
| Chlorophyll ab (mg g−1 FW) | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Treatment | 1st Flowering | 1st Fruiting | 2nd Flowering | 2nd Fruiting | ||||||||
| 2016 | 2017 | Means | 2016 | Treatment | 2016 | 2017 | Means | 2016 | Treatment | 2016 | 2017 | |
| CK | 19.26 de | 20.17 d | 19.71 c | 16.38 | 14.55 | 15.6 c | 14.25 c | 12.39 e | 13.32 c | 11.86 | 11.48 | 11.67 c |
| RGS1 | 23.94 c | 26.29 b | 25.11 b | 19.83 | 17.78 | 18.80 b | 14.77 bc | 14.22 c | 14.49 b | 12.79 | 12.93 | 12.86 b |
| RGS2 | 27.30 ab | 28.27 a | 27.78 a | 21.29 | 18.55 | 19.91 a | 17.70 a | 15.13 b | 16.42 a | 14.08 | 13.69 | 13.88 a |
| RGS3 | 18.3 ef | 17.72 f | 18.02 d | 15.24 | 12.55 | 13.89 d | 1.33 d | 11.30 f | 12.31 d | 10.75 | 10.67 | 10.71 d |
| Year means | 22.21 | 2.11 | 18.18 | 15.86 | 15.02 | 13.26 | 12.36 | 12.19 | ||||
| LSD-test | Treatment | Year | Interaction | Treatment | Year | Interaction | Treatment | Year | Interaction | Treatment | Year | Interaction |
| *** | ** | * | *** | *** | NS | *** | *** | ** | *** | NS | NS | |
| Carotenoids (mg g−1 FW) | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Treatment | 1st Flowering | 1st Fruiting | 2nd Flowering | 2nd Fruiting | ||||||||
| 2016 | 2017 | Means | 2016 | Treatment | 2016 | 2017 | Means | 2016 | Treatment | 2016 | 2017 | |
| CK | 3.36 e | 4.69 c | 4.02 b | 2.92 c | 3.05 c | 2.99 b | 1.36 d | 3.51 b | 2.43 b | 1.71 | 2.21 | 1.97 b |
| RGS1 | 3.73 de | 6.39 a | 5.06 a | 4.78 a | 3.89 b | 4.33 a | 1.57 d | 3.91 b | 2.74 a | 2.01 | 2.53 | 2.27 a |
| RGS2 | 4.04 d | 5.92 b | 4.98 a | 3.92 b | 4.29 ab | 4.10 a | 1.63 d | 4.34 a | 2.99 a | 2.05 | 2.49 | 2.27 a |
| RGS3 | 3.34 e | 3.70 de | 3.52 c | 2.31 d | 2.40 d | 2.36 c | 1.28 d | 3.02 c | 2.15 b | 1.29 | 2.16 | 1.73 c |
| Year means | 3.62 | 5.18 | 3.48 | 3.41 | 1.46 | 3.69 | 1.77 | 2.35 | ||||
| LSD-test | Treatment | Year | Interaction | Treatment | Year | Interaction | Treatment | Year | Interaction | Treatment | Year | Interaction |
| *** | *** | *** | *** | NS | ** | *** | *** | * | *** | *** | NS | |
| Number of Infected Plants | Disease Incidence (%) | Disease Severity Index | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Treatment | 2016 | 2017 | Means | 2016 | 2017 | Means | 2016 | 2017 | Means |
| CK | 8.00 b | 12.01 a | 10.00 | 26.67 b | 40.06 a | 33.36 | 50.33 a | 51.00 a | 50.67 |
| RGS1 | 7.17 b | 7.00 b | 7.08 | 23.89 bc | 20.67 c | 22.28 | 38.00 b | 39.00 b | 38.50 |
| RGS2 | 5.00 c | 4.00 cd | 4.50 | 16.67 d | 13.33 de | 15.00 | 37.67 b | 32.00 c | 34.83 |
| RGS3 | 3.00 de | 2.00 e | 2.50 | 10.00 ef | 6.67 f | 8.33 | 33.00 c | 31.00 c | 32.00 |
| Year means | 5.79 | 6.25 | 19.31 | 20.18 | 39.75 | 38.25 | |||
| LSD-test | Treatment | Year | Interaction | Treatment | Year | Interaction | Treatment | Year | Interaction |
| *** | NS | *** | *** | NS | *** | *** | * | * | |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ghani, M.I.; Ali, A.; Atif, M.J.; Ali, M.; Amin, B.; Anees, M.; Cheng, Z. Soil Amendment with Raw Garlic Stalk: A Novel Strategy to Stimulate Growth and the Antioxidative Defense System in Monocropped Eggplant in the North of China. Agronomy 2019, 9, 89. https://doi.org/10.3390/agronomy9020089
Ghani MI, Ali A, Atif MJ, Ali M, Amin B, Anees M, Cheng Z. Soil Amendment with Raw Garlic Stalk: A Novel Strategy to Stimulate Growth and the Antioxidative Defense System in Monocropped Eggplant in the North of China. Agronomy. 2019; 9(2):89. https://doi.org/10.3390/agronomy9020089
Chicago/Turabian StyleGhani, Muhammad Imran, Ahmad Ali, Muhammad Jawaad Atif, Muhammad Ali, Bakht Amin, Muhammad Anees, and Zhihui Cheng. 2019. "Soil Amendment with Raw Garlic Stalk: A Novel Strategy to Stimulate Growth and the Antioxidative Defense System in Monocropped Eggplant in the North of China" Agronomy 9, no. 2: 89. https://doi.org/10.3390/agronomy9020089