Pre-Sowing Irrigation Plus Surface Fertilization Improves Morpho-Physiological Traits and Sustaining Water-Nitrogen Productivity of Cotton



Abstract
:1. Introduction
2. Materials and Methods
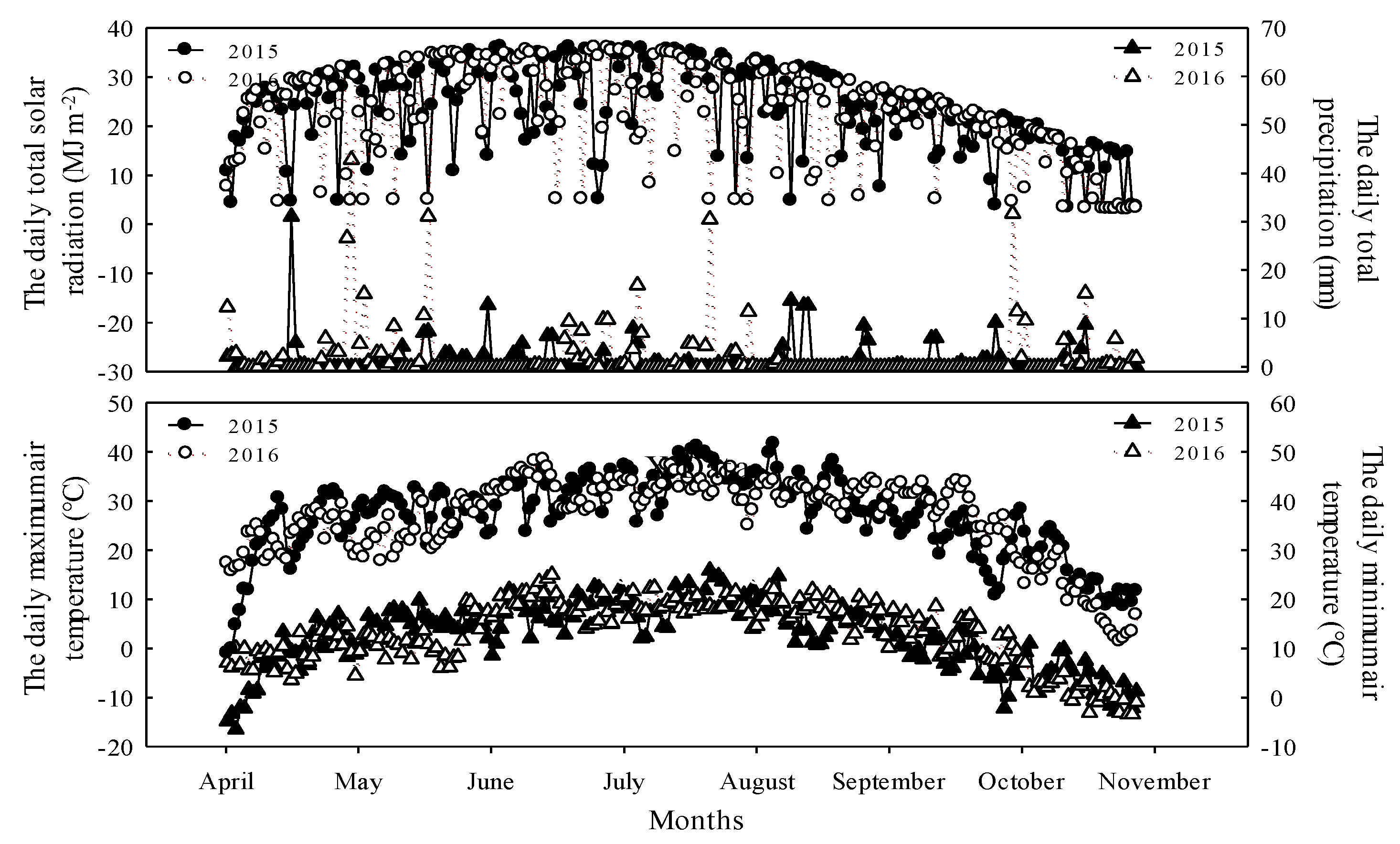
2.1. Details of Experimental Site
2.2. Experimental Design and Crop Management
2.3. Observations
2.4. Soil Water Content and Available Nitrogen
2.5. Root Growth Traits
2.6. Root Nitrate Reductase Activity (NR)
2.7. Biomass Accumulation
2.8. Water-Nitrogen Productivity
2.9. Statistical Analysis
3. Results
3.1. Soil Water Content and Available Nitrogen
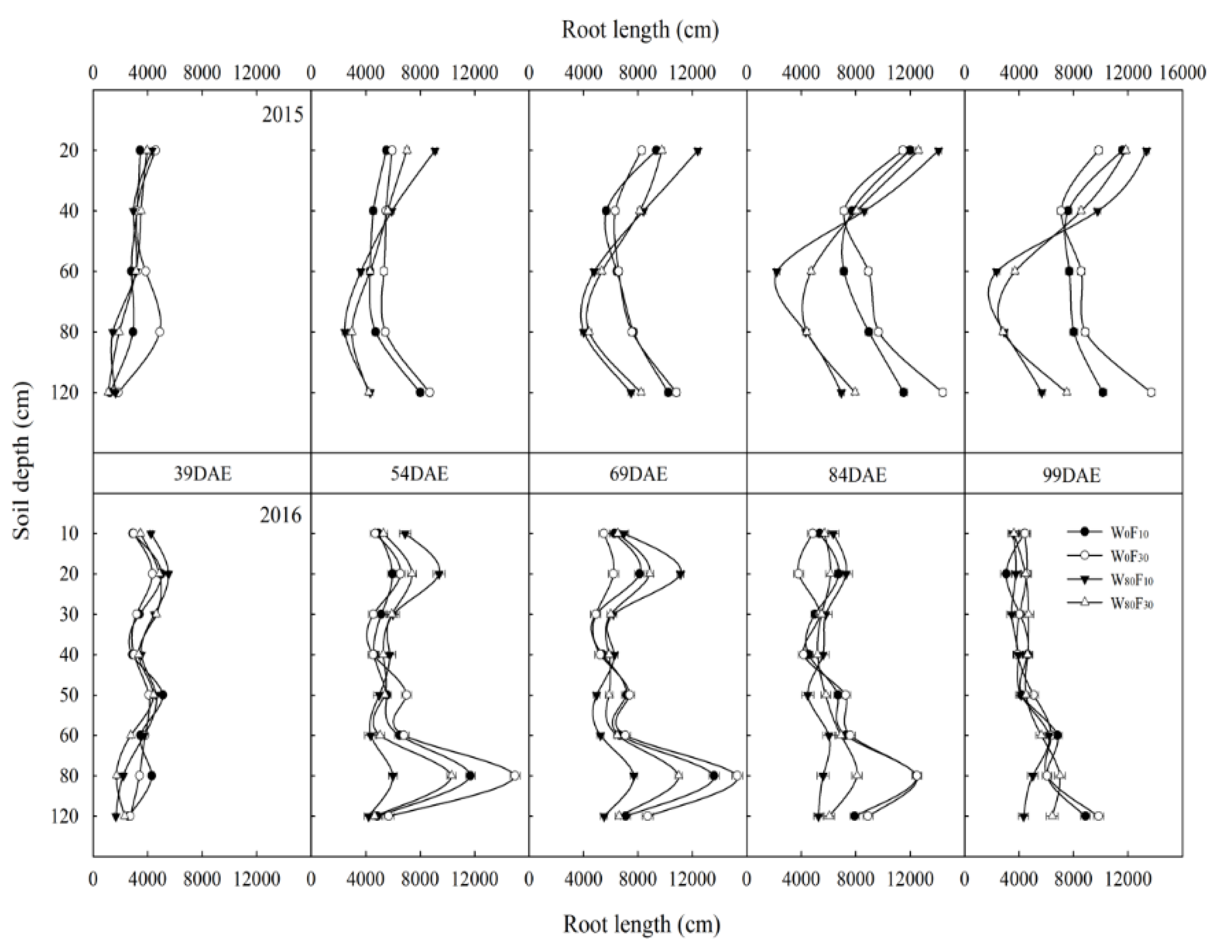
3.2. Root Length
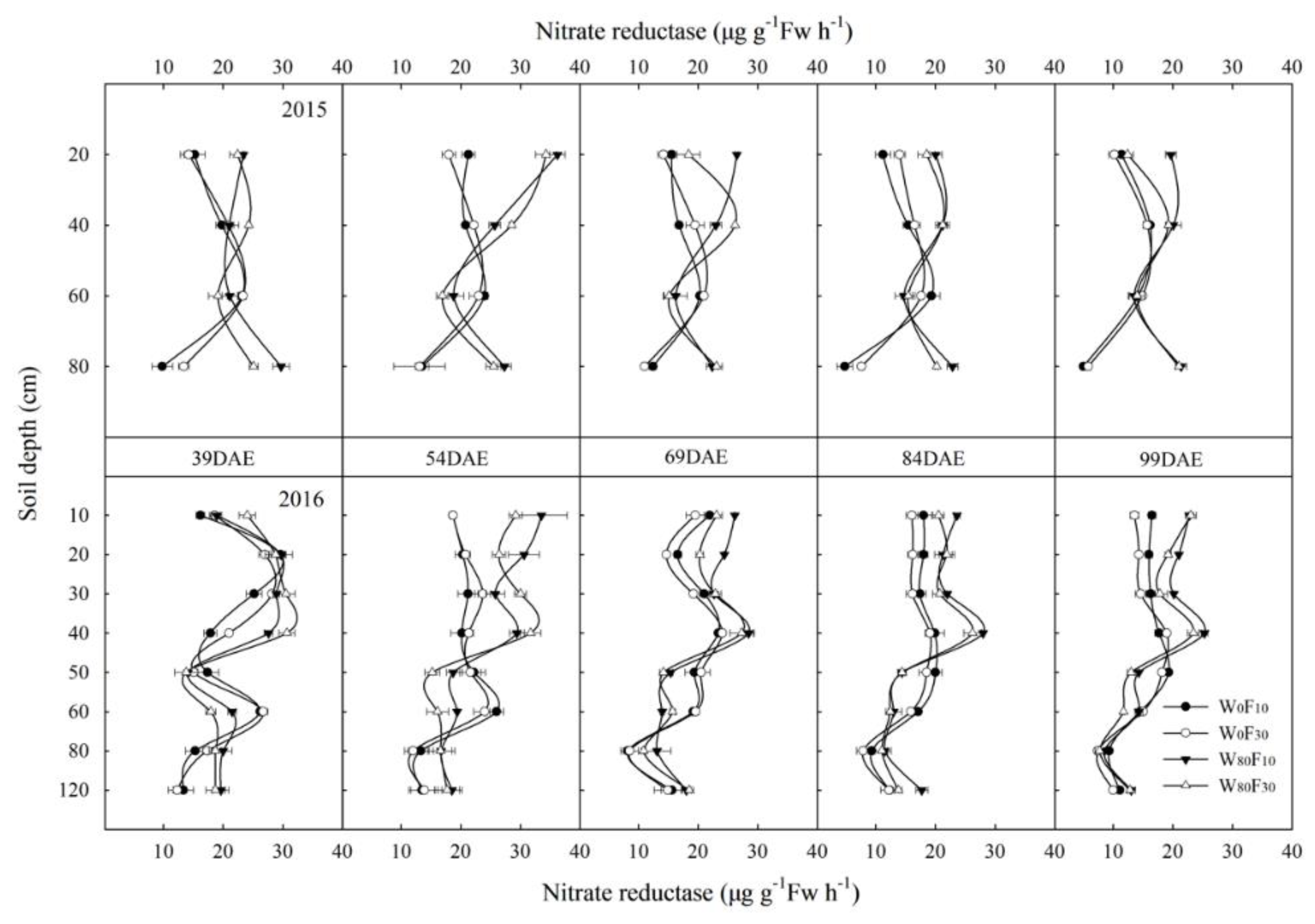
3.3. Root Nitrate Reductase Activity
3.4. Cotton Plant Biomass Accumulation
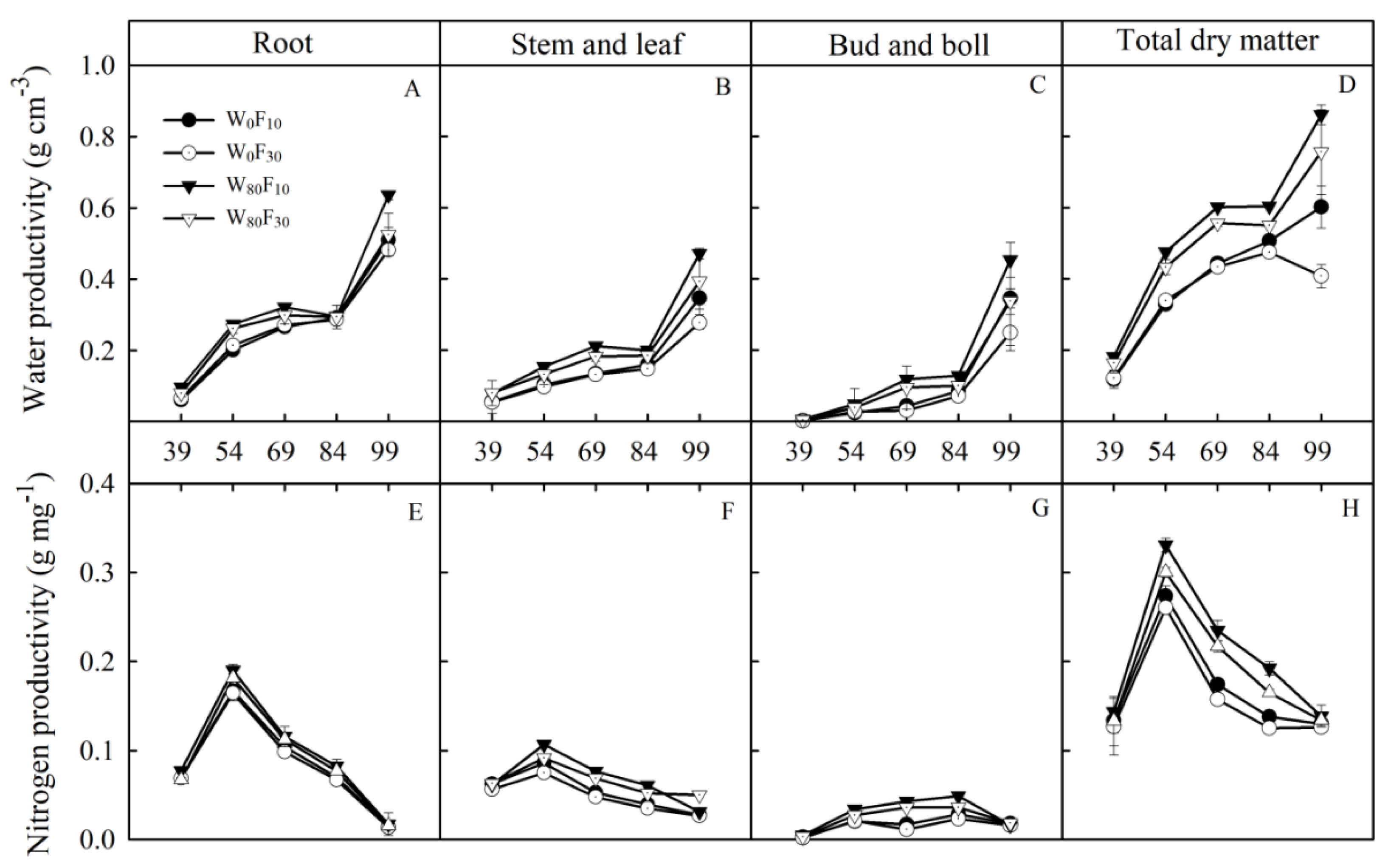
3.5. Water-Nitrogen Productivity
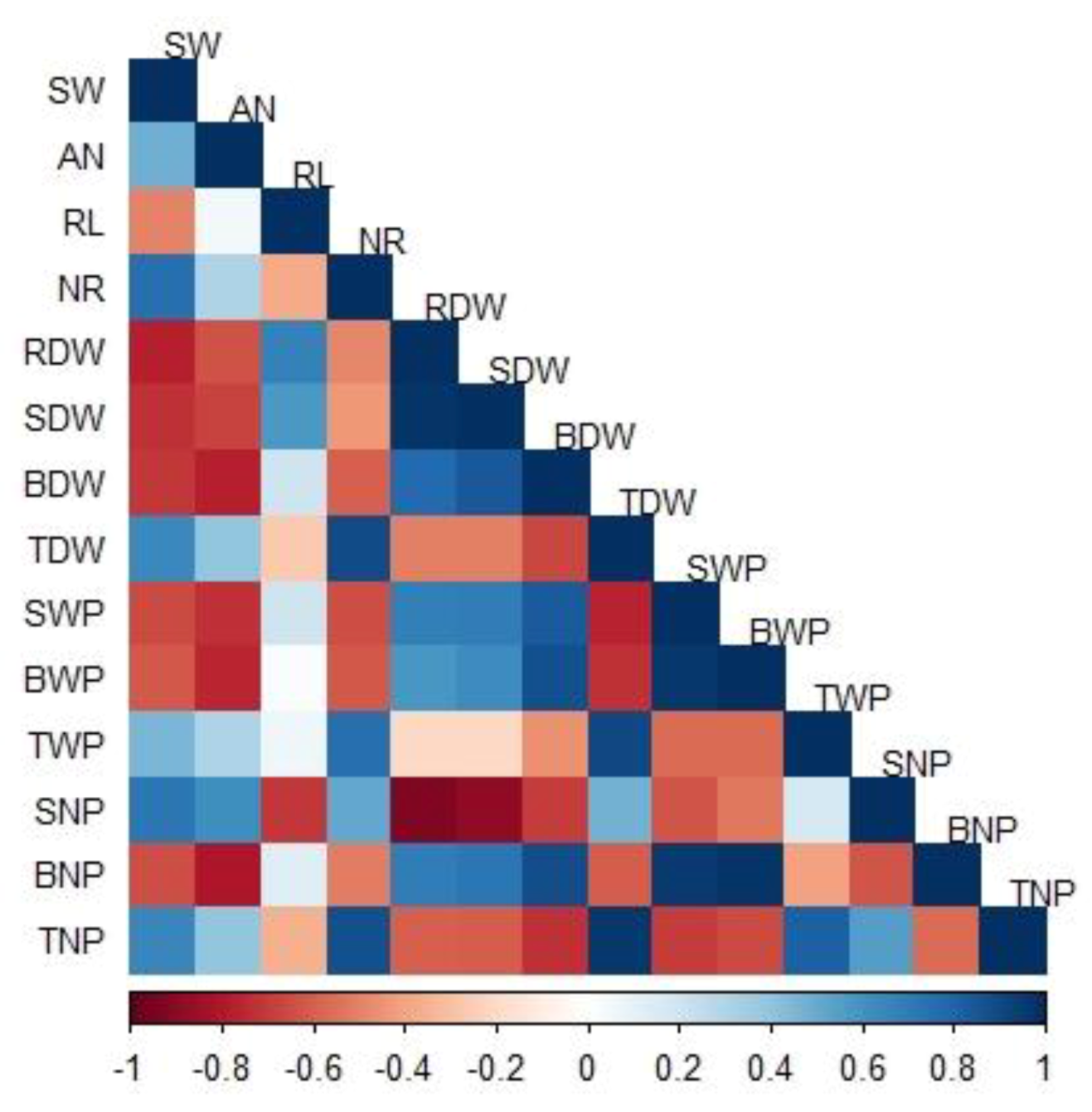
3.6. Factors Affecting Productivity
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Constable, G.A.; Bange, M.P. The yield potential of cotton (Gossypium hirsutum L.). Field Crops Res. 2015, 182, 98–106. [Google Scholar] [CrossRef]
- Khan, A.; Tan, D.K.Y.; Afridi, M.Z.; Luo, H.; Tung, S.A.; Ajab, M.; Fahad, S. Nitrogen fertility and abiotic stresses management in cotton crop: A review. Environ. Sci. Pollut. R 2017, 24, 14551–14566. [Google Scholar] [CrossRef] [PubMed]
- Khan, A.; Pan, X.D.; Najeeb, U.; Tan, D.K.Y.; Fahad, S.; Zahoor, R.; Luo, H.H. Coping with drought: Stress and adaptive mechanisms, and management through cultural and molecular alternatives in cotton as vital constituents for plant stress resilience and fitness. Biol. Res. 2018, 51, 47. [Google Scholar] [CrossRef] [PubMed]
- Berry, P.; Ramirezvillegas, J.; Bramley, H.; Mgonja, M.A.; Mohanty, S.; Jackson, M.; Fordlloyd, B.; Parry, M. Regional impacts of climate change on agriculture and the role of adaptation. In Plant Genetic Resources and Climate Change; CABI: London, UK, 2014; Volume 2, pp. 78–97. [Google Scholar]
- Zhang, X.Y.; Chen, S.Y.; Sun, H.Y.; Wang, Y.M.; Shao, L.W. Root size, distribution and soil water depletion as affected by cultivars and environmental factors. Field Crops Res. 2009, 114, 75–83. [Google Scholar] [CrossRef]
- Bouman, B.A.M. A conceptual framework for the improvement of crop water productivity at different spatial scales. Agric. Syst. 2007, 93, 43–60. [Google Scholar] [CrossRef]
- Afridi, M.Z.; Jan, M.T.; Arif, M.; Jan, A. Wheat yielding components response to different levels of fertilizer-N application time and decapitation stress. Sarhad J. Agric. 2010, 26, 499–506. [Google Scholar]
- Liao, N.; Hou, Z.; Qi, L.I.; Ru, S.; Huijuan, B.O. Increase effect of biochar on cotton yield and nitrogen use efficiency under different nitrogen application levels. Plant Nutr. Fertil. Sci. 2015, 21, 782–791. [Google Scholar]
- Dai, J.L.; Li, W.J.; Tang, W.; Zhang, D.M.; Li, Z.H.; Lu, H.Q.; Eneji, A.E.; Dong, H.Z. Manipulation of dry matter accumulation and partitioning with plant density in relation to yield stability of cotton under intensive management. Field Crops Res. 2015, 180, 207–215. [Google Scholar] [CrossRef]
- Rao, S.S.; Tanwar, S.P.S.; Regar, P.L. Effect of deficit irrigation, phosphorous inoculation and cycocel spray on root growth, seed cotton yield and water productivity of drip irrigated cotton in arid environment. Agric. Water Manag. 2016, 169, 14–25. [Google Scholar] [CrossRef]
- Han, H.M.; Tian, Z.W.; Fan, Y.H.; Cui, Y.K.; Cai, J.; Jiang, D.; Cao, W.X.; Dai, T.B. Water-deficit treatment followed by re-watering stimulates seminal root growth associated with hormone balance and photosynthesis in wheat (Triticum aestivum L.) seedlings. Plant Growth Regul. 2015, 77, 201–210. [Google Scholar] [CrossRef]
- Song, H.; Feng, B.L.; Gao, X.L.; Dai, H.P.; Zhang, P.P. Relationship between root activity and leaf Senescence in different adzuki bean cultivars (lines). Acta Bot. Boreali-Occident. Sin. 2011, 31, 2270–2275. [Google Scholar]
- Amanullah. Effects of NPK source on the dry matter partitioning in cool season C3-cereals (wheat, rye, barley, and oats) at various growth stages. J. Plant Nutr. 2017, 40, 352–364. [Google Scholar] [CrossRef]
- Flexas, J.; Bota, J.; Galmes, J.; Medrano, H.; Ribas-Carbo, M. Keeping a positive carbon balance under adverse conditions: Responses of photosynthesis and respiration to water stress. Physiol. Plant. 2006, 127, 343–352. [Google Scholar] [CrossRef]
- Li, K.F.; Zhang, F.C.; Qi, Y.L. Effects of water and fertility spatial coupling in root zone soil of winter wheat on root growth and activity. Agric. Res. Arid Areas 2009, 27, 48–52. [Google Scholar]
- Wasson, A.P.; Richards, R.A.; Chatrath, R.; Misra, S.C.; Prasad, S.V.; Rebetzke, G.J.; Kirkegaard, J.A.; Christopher, J.; Watt, M. Traits and selection strategies to improve root systems and water uptake in water-limited wheat crops. J. Exp. Bot. 2012, 63, 3485–3498. [Google Scholar] [CrossRef]
- Liu, R.X.; Zhou, Z.G.; Guo, W.Q. Effects of N fertilization on root development and activity of water-stressed cotton (Gossypium hirsutum L.) plants. Agric. Water Manag. 2008, 95, 1261–1270. [Google Scholar] [CrossRef]
- Chen, Y.L.; Zhang, J.; Li, Q.; He, X.L.; Su, X.P.; Chen, F.J.; Yuan, L.X.; Mi, G.H. Effects of nitrogen application on post-silking root senescence and yield of maize. Agron. J. 2015, 107, 835–842. [Google Scholar] [CrossRef]
- Klimesova, J.; Nobis, M.P.; Herben, T. Senescence, ageing and death of the whole plant: Morphological prerequisites and constraints of plant immortality. New Phytol. 2015, 206, 14–18. [Google Scholar] [CrossRef]
- Luo, H.; Zhang, H.; Han, H.; Hu, Y.; Zhang, Y.; Zhang, W. Effects of water storage in deeper soil layers on growth, yield, and water productivity of cotton (Gossypium Hirsutum L.) in arid areas of northwestern China. Irrig. Drain. 2014, 63, 59–70. [Google Scholar] [CrossRef]
- Lu, X.R.; Jia, X.Y.; Niu, J.H. The present situation and prospects of cotton industry development in China. Sci. Agric. Sin. 2018, 51, 26–36. [Google Scholar]
- Chen, Z.K.; Niu, Y.P.; Ma, H.; Hafeez, A.; Luo, H.H.; Zhang, W.F. Photosynthesis and biomass allocation of cotton as affected by deep-layer water and fertilizer application depth. Photosynthetica 2017, 55, 638–647. [Google Scholar] [CrossRef]
- Shen, Y.F.; Li, S.Q. Effects of the spatial coupling of water and fertilizer on the chlorophyll fluorescence parameters of winter wheat leaves. Agric. Sci. Chin. 2011, 10, 1923–1931. [Google Scholar] [CrossRef]
- Garrido-Lestache, E.; Lopez-Bellido, R.J.; Lopez-Bellido, L. Effect of N rate, timing and splitting and N type on bread-making quality in hard red spring wheat under rainfed Mediterranean conditions. Field Crops Res. 2004, 85, 213–236. [Google Scholar] [CrossRef]
- Baethgen, W.E.; Alley, M.M. Optimizing soil and fertilizer nitrogen use by intensively managed winter wheat. I. crop nitrogen uptake. Agron. J. 1989, 81, 120–125. [Google Scholar] [CrossRef]
- Gwenzi, W.; Veneklaas, E.J.; Holmes, K.W.; Bleby, T.M.; Phillips, I.R.; Hinz, C. Spatial analysis of fine root distribution on a recently constructed ecosystem in a water-limited environment. Plant Soil 2011, 344, 255–272. [Google Scholar] [CrossRef]
- Zhou, Z.; He, H.; Ma, L.; Yu, X.; Mi, Q.; Pang, J.; Tang, G.; Liu, B. Overexpression of a GmCnx1 gene enhanced activity of nitrate reductase and aldehyde oxidase, and boosted mosaic virus resistance in soybean. PLoS ONE 2015, 10, e0124273. [Google Scholar] [CrossRef]
- Yang, G.Z.; Tang, H.Y.; Nie, Y.C.; Zhang, X.L. Responses of cotton growth, yield, and biomass to nitrogen split application ratio. Eur. J. Agron. 2011, 35, 164–170. [Google Scholar] [CrossRef]
- Wang, Z.S.; Min, X.U.; Liu, R.X.; Xiao-Dong, W.U.; Zhu, H.; Chen, B.L.; Zhou, Z.G. Effects of nitrogen rates on biomass and nitrogen accumulation of cotton with different varieties in growth duration. Cotton Sci. 2011, 23, 537–544. [Google Scholar]
- Xu, G.H.; Fan, X.R.; Miller, A.J. Plant nitrogen assimilation and use efficiency. Annu. Rev. Plant Biol. 2012, 63, 153–182. [Google Scholar] [CrossRef]
- Hu, X.T.; Chen, H.; Wang, J.; Meng, X.B.; Chen, F.H. Effects of soil water content on cotton root growth and distribution under mulched drip irrigation. Agric. Sci. Chin. 2009, 8, 709–716. [Google Scholar] [CrossRef]
- Palta, J.A.; Chen, X.; Milroy, S.P.; Rebetzke, G.J.; Dreccer, M.F.; Watt, M. Large root systems: Are they useful in adapting wheat to dry environments? Funct. Plant Biol. 2011, 38, 347–354. [Google Scholar] [CrossRef]
- Lynch, J.P. Steep, cheap and deep: An ideotype to optimize water and N acquisition by maize root systems. Ann. Bot. 2013, 112, 347–357. [Google Scholar] [CrossRef] [PubMed]
- Zwart, S.J.; Bastiaanssen, W.G.M. Review of measured crop water productivity values for irrigated wheat, rice, cotton and maize. Agric. Water Manag. 2004, 69, 115–133. [Google Scholar] [CrossRef]
- Osaki, M.; Shinano, T.; Matsumoto, M.; Zheng, T.; Tadano, T. A root-shoot interaction hypothesis for high productivity of field crops. Soil Sci. Plant Nutr. 1997, 43, 445–454. [Google Scholar] [CrossRef]
- Yang, L.; Zheng, B.; Mao, C.; Qi, X.; Liu, F.; Wu, P. Analysis of transcripts that are differentially expressed in three sectors of the rice root system under water deficit. Mol. Genet. Genom. 2004, 272, 433–442. [Google Scholar] [CrossRef]
- Wu, H.X.; Ma, Y.Z.; Xiao, J.P.; Zhang, Z.H.; Shi, Z.H. Photosynthesis and root characteristics of rice (Oryza sativa L.) in floating culture. Photosynthetica 2013, 51, 231–237. [Google Scholar] [CrossRef]
- Blum, A. Drought resistance, water-use efficiency, and yield potential—Are they compatible, dissonant, or mutually exclusive? Aust. J. Agric. Res. 2005, 56, 1159–1168. [Google Scholar] [CrossRef]
- Lilley, J.M.; Kirkegaard, J.A. Benefits of increased soil exploration by wheat roots. Field Crops Res. 2011, 122, 118–130. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Years | Treatments | Root Dry Matter (g plant−1) | Stem and Leaf Dry Matter (g plant−1) | Bud and Boll Dry Matter (g plant−1) | The Total Dry Matter (g plant−1) |
|---|---|---|---|---|---|
| 2015 | W0F10 | 19.8 ± 0.42 c | 19.6 ± 0.05 c | 17.4 ± 0.59 c | 62.06 ± 0.48 b |
| W0F30 | 19.9 ± 0.29 c | 16.2 ± 0.27 d | 13.1 ± 0.9 d | 53.47 ± 1.53 c | |
| W80F10 | 23.2 ± 0.19 a | 22.1 ± 0.10 a | 23.9 ± 0.01 a | 70.02 ± 0.82 a | |
| W80F30 | 21.1 ± 0.31 b | 24.6 ± 0.04 b | 21.3 ± 1.37 b | 71.4 ± 1.49 a | |
| 2016 | W0F10 | 21.5 ± 0.79 c | 12.2 ± 0.09 c | 12.2 ± 0.04 c | 45.87 ± 0.92 c |
| W0F30 | 21.0 ± 0.61 c | 10.0 ± 0.07 d | 9.0 ± 0.04 d | 40.106 ± 0.83 d | |
| W80F10 | 26.5 ± 0.95 a | 16.9 ± 0.02 a | 16.2 ± 0.04 a | 59.54 ± 1.90 a | |
| W80F30 | 24.1 ± 0.82 b | 15.2 ± 0.06 b | 13.0 ± 0.04 b | 52.28 ± 0.716 b |
| Treatments | R2 | t1 (DAE) | t2 (DAE) | T (DAE) | tm (DAE) | Vm (g plant−1 d−1) | Vt (g plant−1 d−1) | W0 (g plant−1) | |
|---|---|---|---|---|---|---|---|---|---|
| Total Biomass of Cotton Plant | W0F10 | 0.975 | 32.32 | 62.11 | 29.79 | 46.62 | 0.97 | 0.85 | 45.47 |
| W0F30 | 0.966 | 30.18 | 58.64 | 28.45 | 43.84 | 0.87 | 0.77 | 39.31 | |
| W80F10 | 0.99 | 34.20 | 65.46 | 31.26 | 49.21 | 1.23 | 1.08 | 60.54 | |
| W80F30 | 0.998 | 34.04 | 64.55 | 30.51 | 48.69 | 1.08 | 0.96 | 52.25 | |
| Aerial Part Biomass of Cotton Plant | W0F10 | 0.986 | 35.92 | 75.49 | 39.57 | 54.91 | 0.42 | 0.37 | 26.4 |
| W0F30 | 0.962 | 32.25 | 68.94 | 36.70 | 49.86 | 0.41 | 0.36 | 23.9 | |
| W80F10 | 0.987 | 42.34 | 86.00 | 43.66 | 63.30 | 0.51 | 0.46 | 35.50 | |
| W80F30 | 0.996 | 40.75 | 82.26 | 41.51 | 60.68 | 0.46 | 0.40 | 30.00 | |
| Cotton Bud and Boll Biomass | W0F10 | 0.954 | 30.19 | 73.85 | 43.66 | 51.14 | 0.35 | 0.31 | 24.09 |
| W0F30 | 0.984 | 30.43 | 74.86 | 44.43 | 51.76 | 0.29 | 0.25 | 20.17 | |
| W80F10 | 0.997 | 28.46 | 69.3 | 40.84 | 48.07 | 0.47 | 0.41 | 30.22 | |
| W80F30 | 0.997 | 28.84 | 70.35 | 41.51 | 48.76 | 0.40 | 0.36 | 26.33 | |
| Cotton Stem and Leaf Biomass | W0F10 | 0.999 | 28.96 | 66.20 | 37.24 | 46.84 | 0.52 | 0.46 | 30.63 |
| W0F30 | 0.988 | 34.17 | 70.87 | 36.70 | 51.79 | 0.36 | 0.32 | 20.70 | |
| W80F10 | 0.997 | 27.37 | 66.94 | 39.57 | 46.36 | 0.59 | 0.52 | 36.84 | |
| W80F30 | 0.996 | 25.85 | 54.96 | 29.11 | 39.83 | 0.59 | 0.52 | 26.90 | |
| Cotton Root Biomass | W0F10 | 0.997 | 30.86 | 43.24 | 12.38 | 58.70 | 0.29 | 0.21 | 22.03 |
| W0F30 | 0.999 | 29.58 | 38.96 | 9.38 | 61.30 | 0.28 | 0.17 | 20.80 | |
| W80F10 | 0.99 | 26.57 | 34.42 | 7.86 | 62.88 | 0.33 | 0.18 | 25.56 | |
| W80F30 | 0.999 | 23.97 | 30.69 | 6.72 | 63.92 | 0.30 | 0.14 | 23.81 |
| Year | Treatments | Total Moisture Consumption Rate (cm3) | Total Moisture Consumption Ratio | Water Productivity of Root (g DM cm−3) | Water Productivity of Stem and Leaf (g DM cm−3) | Water Productivity of Bud and Boll (g DM cm−3) | Water Productivity of Total Dry Matter (g DM cm−3) |
|---|---|---|---|---|---|---|---|
| 2015 | W0F10 | 799.2 ± 14.62 b | 0.62 ± 0.01 b | 0.33 ± 0.02 a | 0.26 ± 0.00 c | 0.23 ± 0.01 b | 0.60 ± 0.03 c |
| W0F30 | 867.5 ± 13.98 a | 0.69 ± 0.01 a | 0.29 ± 0.01 b | 0.19 ± 0.01 d | 0.15 ± 0.01 c | 0.42 ± 0.03 b | |
| W80F10 | 846.1 ± 16.39 a | 0.46 ± 0.01 d | 0.33 ± 0.01 a | 0.31 ± 0.02 a | 0.33 ± 0.01 a | 0.76 ± 0.03 a | |
| W80F30 | 859.1 ± 19.91 a | 0.55 ± 0.01 c | 0.34 ± 0.01 a | 0.28 ± 0.01 b | 0.25 ± 0.02 b | 0.66 ± 0.032 b | |
| 2016 | W0F10 | 762.1 ± 13.42 b | 0.65 ± 0.01 b | 0.27 ± 0.01 bc | 0.15 ± 0.00 c | 0.15 ± 0.00 b | 0.57 ± 0.01 c |
| W0F30 | 849.1 ± 28.84 a | 0.71 ± 0.02 a | 0.24 ± 0.00 c | 0.12 ± 0.00 d | 0.11 ± 0.00 c | 0.46 ± 0.02 d | |
| W80F10 | 822.7 ± 37.77 a | 0.54 ± 0.00 c | 0.31 ± 0.00 a | 0.20 ± 0.00 a | 0.19 ± 0.01 a | 0.69 ± 0.02 a | |
| W80F30 | 872.2 ± 36.93 a | 0.54 ± 0.02 c | 0.29 ± 0.00 ab | 0.18 ± 0.01 b | 0.16 ± 0.01 b | 0.62 ± 0.03 b |
| Year | Treatments | Nitrogen Productivity of Root (g DM mg−1) | Nitrogen Productivity of Stem and Leaf (g DM mg−1) | Nitrogen Productivity of Bud and Boll (g DM mg−1) | Nitrogen Productivity of Total Dry Matter (g DM mg−1) |
|---|---|---|---|---|---|
| 2015 | W0F10 | 0.11 ± 0.004 b | 0.09 ± 0.000 b | 0.08 ± 0.003 c | 0.28 ± 0.007 c |
| W0F30 | 0.11 ± 0.002 b | 0.07 ± 0.001 c | 0.06 ± 0.004 d | 0.24 ± 0.007 d | |
| W80F10 | 0.13 ± 0.000 a | 0.10 ± 0.005 a | 0.11 ± 0.001 a | 0.33 ± 0.005 a | |
| W80F30 | 0.11 ± 0.001 b | 0.11 ± 0.001 a | 0.09 ± 0.006 b | 0.31 ± 0.009 b | |
| 2016 | W0F10 | 0.10 ± 0.003 b | 0.06 ± 0.000 c | 0.06 ± 0.000 c | 0.21 ± 0.004 c |
| W0F30 | 0.09 ± 0.003 b | 0.05 ± 0.000 d | 0.04 ± 0.000 d | 0.18 ± 0.004 d | |
| W80F10 | 0.12 ± 0.00 a | 0.07 ± 0.000 a | 0.07 ± 0.000 a | 0.26 ± 0.008 a | |
| W80F30 | 0.10 ± 0.003 b | 0.07 ± 0.000 b | 0.07 ± 0.000 b | 0.23± 0.004 b |
| Square and Boll Water Productivity | Square and Boll Nitrogen Productivity | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| x1-1 | x2 | x3 | x4 | x5 | x6 | x1-2 | x2 | x3 | x4 | x5 | x6 | |
| Soil moisture content (x1-1) | 0.058 | 0.41 | 0.455 | −0.183 | 0.943 | 0.121 | - | - | - | - | - | - |
| Available nitrogen (x1-2) | - | - | - | - | - | - | −0.027 | 0.211 | 0.375 | −0.947 | −1.221 | −0.062 |
| Root length (x2) | −0.177 | 0.382 | −0.128 | 0.735 | 1.548 | 0.031 | −0.013 | 0.312 | −0.117 | 0.616 | 0.638 | 0.274 |
| Nitrate reductase (x3) | 0.37 | 0.031 | 0.339 | 0.25 | 1.18 | −0.037 | −0.069 | −0.073 | 0.572 | 0.732 | 0.791 | −0.007 |
| Root dry matter (x4) | −0.277 | −0.056 | −0.165 | 0.564 | 1.633 | 0.226 | 0.143 | 0.13 | 0.179 | 0.201 | 1.765 | 0.055 |
| Stem and leaf dry matter (x5) | −0.259 | −0.047 | −0.149 | 0.512 | 2.688 | 0.033 | 0.154 | 0.111 | −0.151 | 0.471 | 1.801 | 0.152 |
| Bud and boll dry matter (x6) | −0.254 | −0.017 | −0.2 | 0.183 | 0.27 | 0.585 | 0.177 | 0.039 | −0.138 | 0.161 | 0.221 | 0.364 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, Z.; Gao, H.; Hou, F.; Khan, A.; Luo, H. Pre-Sowing Irrigation Plus Surface Fertilization Improves Morpho-Physiological Traits and Sustaining Water-Nitrogen Productivity of Cotton. Agronomy 2019, 9, 772. https://doi.org/10.3390/agronomy9110772
Chen Z, Gao H, Hou F, Khan A, Luo H. Pre-Sowing Irrigation Plus Surface Fertilization Improves Morpho-Physiological Traits and Sustaining Water-Nitrogen Productivity of Cotton. Agronomy. 2019; 9(11):772. https://doi.org/10.3390/agronomy9110772
Chicago/Turabian StyleChen, Zongkui, Hongyun Gao, Fei Hou, Aziz Khan, and Honghai Luo. 2019. "Pre-Sowing Irrigation Plus Surface Fertilization Improves Morpho-Physiological Traits and Sustaining Water-Nitrogen Productivity of Cotton" Agronomy 9, no. 11: 772. https://doi.org/10.3390/agronomy9110772