Micro-Level Management of Agricultural Inputs: Emerging Approaches

{kind=link}
Abstract
:1. Introduction
1.1. Ecosystem Degradation and Resource Depletion
1.2. Attaining Sustainability
2. A Brief History of Agriculture Intensification
2.1. The Emergence of Agriculture as a Science
2.2. The Agricultural Green Revolution
2.3. Traditional Approaches to Agriculture Management
3. Modern Approaches to Agriculture Management—Utilization of Plant Growth Promoting Microorganisms (PGPMs)
4. Micro-Level Agriculture Management: Opportunities and Challenges
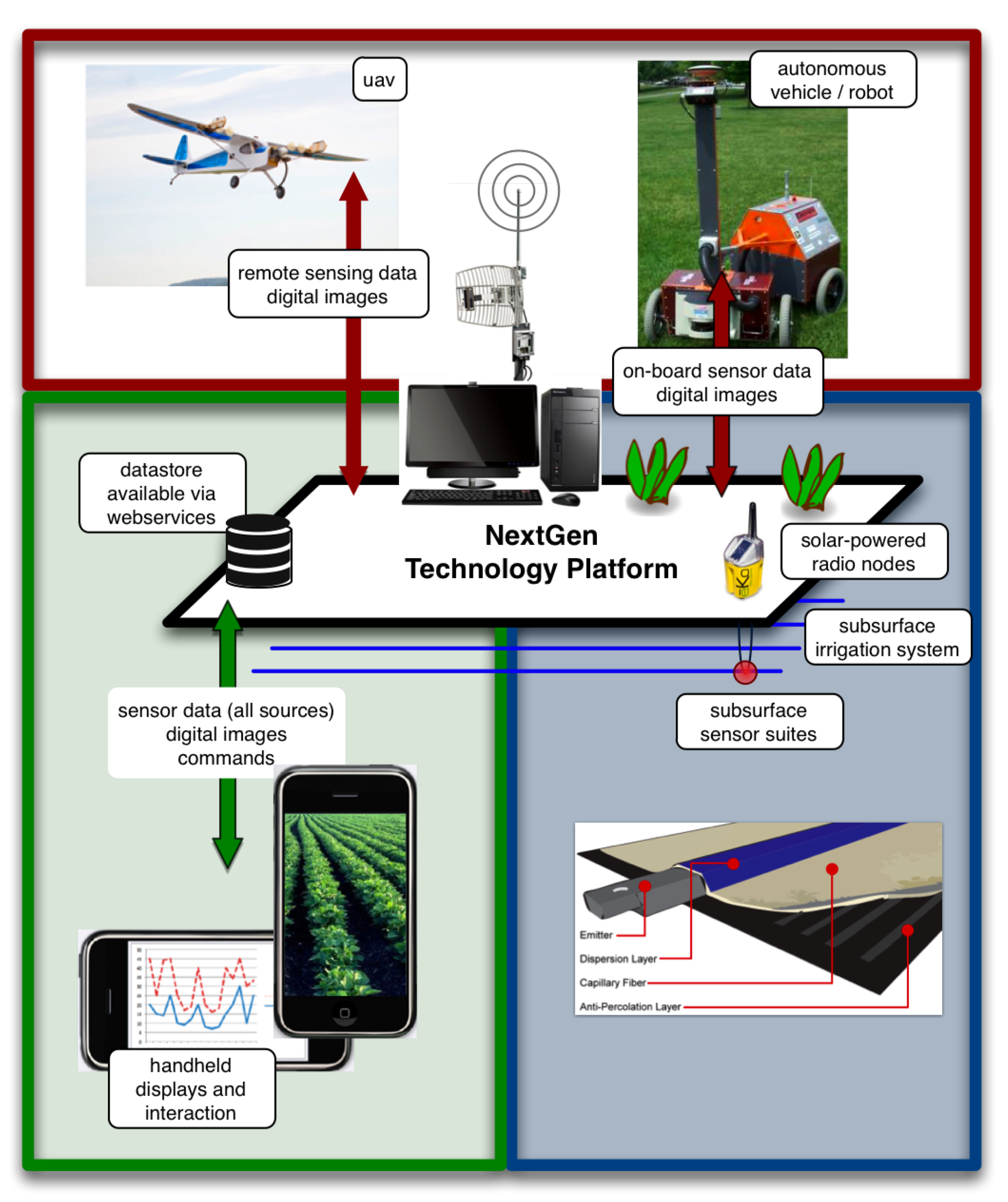
4.1. Smart Field Technology Platform: An Introduction
4.2. Smart Field Technology Platform: Applications
4.3. Smart Field Technology Platform: Interdisciplinary Opportunities

5. Concluding Remarks
Acknowledgments
References
- Eswaran, H.; Lal, R.; Reich, P.F. Land degradation: An overview. In Responses to Land Degradation; Bridges, E.M., Hannam, I.D., Oldeman, L.R., Pening de Vries, F.W.T., Scherr, S.J., Sompatpanit, S., Eds.; Science Publishers Inc.: Enfield, NH, USA, 2001; pp. 20–35. [Google Scholar]
- Smith, P.; Martino, D.; Cai, Z.; Gwary, D.; Janzen, H.; Kumar, P.; McCarl, B.; Ogle, S.; O’Mara, F.; Rice, C.; et al. Greenhouse gas mitigation in agriculture. Philos. Trans. R. Soc. B Biol. Sci. 2008, 363, 789–813. [Google Scholar] [CrossRef]
- Tilman, D.; Cassman, K.G.; Matson, P.A.; Naylor, R.; Polasky, S. Agricultural sustainability and intensive production practices. Nature 2002, 418, 671–677. [Google Scholar] [CrossRef]
- Sutton, M.A.; Oenema, O.; Erisman, J.W.; Leip, A.; van Grinsven, H.; Winiwarter, W. Too much of a good thing. Nature 2011, 472, 159–161. [Google Scholar]
- Matson, P.A.; Naylor, R.; Ortiz-Monasterio, I. Integration of environmental, agronomic, and economic aspects of fertilizer management. Science 1998, 280, 112–115. [Google Scholar] [CrossRef]
- Postel, S.L. Water for food production: Will there be enough in 2025? Bioscience 1998, 48, 629. [Google Scholar] [CrossRef]
- Cassman, K.G. Ecological intensification of cereal production systems: Yield potential, soil quality, and precision agriculture. Proc. Natl. Acad. Sci. USA 1999, 96, 5952–5959. [Google Scholar] [CrossRef]
- Smith, K.A.; Ball, T.; Conen, F.; Dobbie, K.E.; Massheder, J.; Rey, A. Exchange of greenhouse gases between soil and atmosphere: Interactions of soil physical factors and biological processes. Eur. J. Soil Sci. 2003, 54, 779–791. [Google Scholar] [CrossRef]
- Galloway, J.N.; Townsend, A.R.; Erisman, J.W.; Bekunda, M.; Cai, Z.C.; Freney, J.R.; Martinelli, L.A.; Seitzinger, S.P.; Sutton, M.A. Transformation of the nitrogen cycle: Recent trends, questions, and potential solutions. Science 2008, 320, 889–892. [Google Scholar]
- McKenzie, V.J.; Townsend, A.R. Parasitic and infectious disease responses to changing global nutrient cycles. EcoHealth 2007, 4, 384–396. [Google Scholar]
- Lafferty, K.D. How should environmental stress affect the population dynamics of disease? Ecol. Lett. 2003, 6, 654–664. [Google Scholar] [CrossRef]
- Howarth, R.W.; Boyer, E.W.; Pabich, W.J.; Galloway, J.N. Nitrogen use in the united states from 1961 to 2000 and potential future trends. Ambio 2002, 31, 88–96. [Google Scholar]
- Tilman, D.; Fargione, J.; Wolff, B.; D’Antonio, C.; Dobson, A.; Howarth, R.; Schindler, D.; Schlesinger, W.H.; Simberloff, D.; Swackhamer, D. Forecasting agriculturally driven global environmental change. Science 2001, 292, 281–284. [Google Scholar]
- Sutton, M.A.; Howard, C.M.; Erisman, J.W.; Billen, G.; Bleeker, A.; Grennfelt, P.; van Grinsven, H.; Grizzetti, B. The European Nitrogen Assessment; Cambridge University Press: Cambridge, UK, 2011; p. 664. [Google Scholar]
- Day, W. Engineering advances for input reduction and systems management to meet the challenges of global food and farming futures. J. Agric. Sci. 2011, 149, 55–61. [Google Scholar] [CrossRef]
- Cole, C.V.; Duxbury, J.; Freney, J.; Heinemeyer, O.; Minami, K.; Mosier, A.; Paustian, K.; Rosenberg, N.; Sampson, N.; Sauerbeck, D.; et al. Global estimates of potential mitigation of greenhouse gas emissions by agriculture. Nutr. Cycl. Agroecosys. 1997, 49, 221–228. [Google Scholar] [CrossRef]
- Hou, A.X.; Chen, G.X.; Wang, Z.P.; van Cleemput, O.; Patrick, W.H. Methane and nitrous oxide emissions from a rice field in relation to soil redox and microbiological processes. Soil Sci. Soc. Am. J. 2000, 64, 2180–2186. [Google Scholar] [CrossRef]
- United States Environmental Protection Agency. Nitrous Oxide Emissions. Available online: http://www.epa.gov/nitrousoxide/scientific.html (accessed on 1 October 2012).
- Forster, P.; Ramaswamy, V.; Artaxo, P.; Berntsen, T.; Betts, R.; Fahey, D.; Haywood, J.; Lean, J.; Lowe, D.; Myhre, G.; et al. Changes in atmospheric constituents and in radiative forcing. In Climate Change 2007: The Physical Science Basis: Contribution of Working Group I to the Fourth Assessment Report of the Intergovernmental Panel on Climate Change; Solomon, S., Qin, D., Manning, M., Chen, Z., Marquis, M., Averyt, K., Tignor, M., Miller, H., Eds.; Cambridge University Press: Cambridge, UK, 2007; pp. 129–234. [Google Scholar]
- Dalal, R.C.; Wang, W.; Robertson, G.P.; Parton, W.J. Nitrous oxide emission from Australian agricultural lands and mitigation options: A review. Soil Res. 2003, 41, 165–195. [Google Scholar] [CrossRef]
- Daily, G.C. Ecology: The value of nature and the nature of value. Science 2000, 289, 395–396. [Google Scholar] [CrossRef]
- Khan, M.S.; Zaidi, A.; Wani, P.A. Role of phosphate solubilizing microorganisms in sustainable agriculture—A review. In Sustainable Agriculture; Lichtfouse, E., Navarrete, M., Debaeke, P., Véronique, S., Alberola, C., Eds.; Springer: Amsterdam, The Netherlands, 2009; pp. 551–570. [Google Scholar]
- Postel, S.L.; Daily, G.C.; Ehrlich, P.R. Human appropriation of renewable fresh water. Science 1996, 271, 785–788. [Google Scholar]
- Gordon, L.J.; Steffen, W.; Jonsson, B.F.; Folke, C.; Falkenmark, M.; Johannessen, A. Human modification of global water vapor flows from the land surface. Proc. Natl. Acad. Sci. USA 2005, 102, 7612–7617. [Google Scholar] [CrossRef]
- Wallace, J.S. Increasing agricultural water use efficiency to meet future food production. Agric. Ecosyst. Environ. 2000, 82, 105–119. [Google Scholar] [CrossRef]
- Alizadeh, A. Status of Agricultural Water Use in Iran. In Water Conservation, Reuse, and Recycling: Proceeding of an Iranian American Workshop; National Academies Press: Washington, DC, USA, 2005; pp. 94–105. [Google Scholar]
- Barnett, T.P.; Pierce, D.W. When will lake mead go dry? Water Resour. Res. 2008, 44, W3201. [Google Scholar] [CrossRef]
- Oki, T.; Kanae, S. Global hydrological cycles and world water resources. Science 2006, 313, 1068–1072. [Google Scholar] [CrossRef]
- Walker, W.R. Surface Irrigation: Theory and Practice; Prentice-Hall: Englewood Cliffs, NJ, USA, 1987; p. 386. [Google Scholar]
- Kay, M. Surface Irrigation: Systems and Practice; Cranfield Press: Cranfield, UK, 1986; p. 142. [Google Scholar]
- Foley, J.A.; DeFries, R.; Asner, G.P.; Barford, C.; Bonan, G.; Carpenter, S.R.; Chapin, F.S.; Coe, M.T.; Daily, G.C.; Gibbs, H.K.; et al. Global consequences of land use. Science 2005, 309, 570–574. [Google Scholar]
- Lymphatic Filariasis Elimination Program. The Cater Center Web site. Available online: http://www.cartercenter.org/health/lf/index.html (accessed on 25 September 2012).
- Malaria Control Program. The Cater Center Web site. Available online: http://www.cartercenter.org/health/malaria_control/index.html (accessed on 25 September 2012).
- Schistosomiasis Control Program. The Cater Center Web site. Available online: http://www.cartercenter.org/health/schistosomiasis/index.html (accessed on 25 September 2012).
- Ward, F.A.; Pulido-Velazquez, M. Water conservation in irrigation can increase water use. Proc. Natl. Acad. Sci. USA 2008, 105, 18215–18220. [Google Scholar] [CrossRef]
- Erisman, J.W. How a century of ammonia synthesis changed the world. Nat. Geosci. 2008, 1, 636–639. [Google Scholar] [CrossRef]
- Food and Agriculture Organization of the United Nations Web site. Available online: http://faostat.fao.org/ (accessed on 17 September 2012).
- Mazoyer, M.; Roudart, L. A History of World Agriculture from the Neolithic Age to the Current Crisis; Monthly Review Press: New York, NY, USA, 2006; p. 528. [Google Scholar]
- Biography: Luther Burbank, 1849–1926. Western Sonoma County Historical Society Web site. Available online: http://www.wschsgrf.org/articles/biographylutherburbank1849-1926 (accessed on 4 October 2012).
- Jethro Tull (1674–1741). BBC History Web site. Available online: http://www.bbc.co.uk/history/historic_figures/tull_jethro.shtml (accessed on 4 October 2012).
- Welbaum, G.E.; Sturz, A.V.; Dong, Z.M.; Nowak, J. Managing soil microorganisms to improve productivity of agro-ecosystems. Crit. Rev. Plant Sci. 2004, 23, 175–193. [Google Scholar] [CrossRef]
- Schils, R. How James Watt Invented the Copier: Forgotten Inventions of Our Great Scientists; Springer: New York, NY, USA, 2011; pp. 63–69. [Google Scholar]
- Twigg, C.A.; Twigg, M.V. Centenary of the death of Justus von Liebig. J. Chem. Educ. 1973, 50, 273. [Google Scholar] [CrossRef]
- McNeill, J.R.; Winiwarter, V. Breaking the sod: Humankind, history, and soil. Science 2004, 304, 1627–1629. [Google Scholar] [CrossRef]
- Egli, T.; Zinn, M. The concept of multiple-nutrient-limited growth of microorganisms and its application in biotechnological processes. Biotechnol. Adv. 2003, 22, 35–43. [Google Scholar] [CrossRef]
- Bongaarts, J. Human population growth and the demographic transition. Philos. Trans. R. Soc. B Biol. Sci. 2009, 364, 2985–2990. [Google Scholar] [CrossRef]
- De Fraiture, C.; Molden, D.; Wichelns, D. Investing in water for food, ecosystems, and livelihoods: An overview of the comprehensive assessment of water management in agriculture. Agric. Water Manag. 2010, 97, 495–501. [Google Scholar] [CrossRef]
- Rudel, T.K.; Schneider, L.; Uriarte, M.; Turner, B.L.; DeFries, R.; Lawrence, D.; Geoghegan, J.; Hecht, S.; Ickowitz, A.; Lambin, E.F.; et al. Agricultural intensification and changes in cultivated areas, 1970–2005. Proc. Natl. Acad. Sci. USA 2009, 106, 20675–20680. [Google Scholar]
- World Population Prospects: The 2010 Revision, Press Release 3 May 2011. Available online: http://esa.un.org/wpp/Documentation/publications.htm (accessed on 15 August 2012).
- Tilman, D.; Balzer, C.; Hill, J.; Befort, B.L. Global food demand and the sustainable intensification of agriculture. Proc. Natl. Acad. Sci. USA 2011, 108, 20260–20264. [Google Scholar]
- McNeill, J.R. Something New under the Sun: An Environmental History of the Twentieth-Century World, 1st ed; Norton: New York, NY, USA, 2000; p. 421. [Google Scholar]
- Pardey, P.G.; Beintema, N.M. Slow Magic: Agricultural R&D a Century after Mendel; International Food Policy Research Institute: Washington, DC, USA, 2001; pp. 11–12. [Google Scholar]
- Cook, R.J. Toward cropping systems that enhance productivity and sustainability. Proc. Natl. Acad. Sci. USA 2006, 103, 18389–18394. [Google Scholar] [CrossRef]
- Srinivasan, A. Handbook of Precision Agriculture: Principles and Applications; Food Products Press: New York, NY, USA, 2006; p. 683. [Google Scholar]
- Robert, P.C. Precision agriculture: A challenge for crop nutrition management. Plant Soil 2002, 247, 143–149. [Google Scholar] [CrossRef]
- Virginia Tech Soil Testing Lab Web site. Available online: http://www.soiltest.vt.edu/ (accessed on 25 September 2012).
- Christy, C.D. Real-time measurement of soil attributes using on-the-go near infrared reflectance spectroscopy. Comput. Electron. Agric. 2008, 61, 10–19. [Google Scholar]
- LSU AgCenter Web site. Available online: www.LSUAgCenter.com (accessed on 11 October 2011).
- Geonics Web site. Available online: http://www.geonics.com (accessed on 11 October 2011).
- Sturz, A.V.; Christie, B.R.; Nowak, J. Bacterial endophytes: Potential role in developing sustainable systems of crop production. Crit. Rev. Plant Sci. 2000, 19, 1–30. [Google Scholar] [CrossRef]
- Irrigation & Water Technologies Pty. Ltd. Web site. Available online: www.kisss.com.au (accessed on 8 December 2012).
- Irrigro Web site. Available online: www.irrigro.com (accessed on 8 December 2012).
- Gabbard, J.; Gabbard, C.; Nowak, J. Enabling Sustainable Agriculture and Natural Resource Research and Education by Integrating Engineering Technology into Sustainable Practices; Annual Report—Project #117789; Virginia Tech Institute for Critical Technology and Applied Science (ICTAS): Blacksburg, VA, USA, 2012; Unpublished work. [Google Scholar]
- Sturz, A.V.; Nowak, J. Endophytic communities of rhizobacteria and the strategies required to create yield enhancing associations with crops. Appl. Soil Ecol. 2000, 15, 183–190. [Google Scholar] [CrossRef]
- Frommel, M.I.; Nowak, J.; Lazarovits, G. Growth enhancement and developmental modifications of in vitro grown potato (Solanum tuberosum spp. Tuberosum) as affected by a nonfluorescent Pseudomonas sp. Plant Physiol. 1991, 96, 928–936. [Google Scholar] [CrossRef]
- Gochnauer, M.B.; McCully, M.E.; Labbe, H. Different populations of bacteria associated with sheathed and bare regions of roots of field-grown maize. Plant Soil 1989, 114, 107–120. [Google Scholar] [CrossRef]
- Vessey, J.K. Plant growth promoting rhizobacteria as biofertilizers. Plant Soil 2003, 255, 571–586. [Google Scholar]
- Wilson, D. Endophyte—The evolution of a term, and clarification of its use and definition. Oikos 1995, 73, 274–276. [Google Scholar] [CrossRef]
- Bell, C.R.; Dickie, G.A.; Harvey, W.L.G.; Chan, J. Endophytic bacteria in grapevine. Can. J. Microbiol. 1995, 41, 46–53. [Google Scholar]
- Kloepper, J.W. Plant growth-promoting rhizobacteria as biological control agents. In Soil Microbial Ecology: Applications in Agriculture and Environmental Management; Metting, F.B., Jr., Ed.; Marcel Dekker Inc.: New York, NY, USA, 1992; pp. 255–274. [Google Scholar]
- Kim, S.; Lowman, S.; Hou, G.; Nowak, J.; Flinn, B.; Mei, C. Growth promotion and colonization of switchgrass (Panicum virgatum) cv. Alamo by bacterial endophyte Burkholderia phytofirmans strain PsJN. Biotechnol. Biofuels 2012, 5, 37. [Google Scholar] [CrossRef]
- Compant, S.; Mitter, B.; Colli-Mull, J.; Gangl, H.; Sessitsch, A. Endophytes of grapevine flowers, berries, and seeds: Identification of cultivable bacteria, comparison with other plant parts, and visualization of niches of colonization. Microb. Ecol. 2011, 62, 188–197. [Google Scholar] [CrossRef]
- Zilber-Rosenberg, I.; Rosenberg, E. Role of microorganisms in the evolution of animals and plants: The hologenome theory of evolution. FEMS Microbiol. Rev. 2008, 32, 723–735. [Google Scholar] [CrossRef]
- Nowak, J.; Asiedu, S.K.; Bensalim, S.; Richards, J.; Stewart, A.; Smith, C.; Stevens, D.; Sturz, A.V. From laboratory to applications: Challenges and progress with in vitro dual cultures of potato and beneficial bacteria. Plant Cell Tissue Organ Cult. 1998, 52, 97–103. [Google Scholar] [CrossRef]
- Da, K.; Nowak, J.; Flinn, B. Potato cytosine methylation and gene expression changes induced by a beneficial bacterial endophyte, Burkholderia phytofirmans strain PsJN. Plant Physiol. Biochem. 2012, 50, 24–34. [Google Scholar] [CrossRef]
- Nowak, J. Benefits of in vitro “biotization” of plant tissue cultures with microbial inoculants. In Vitro Cell. Dev. Biol. Plant 1998, 34, 122–130. [Google Scholar] [CrossRef]
- Galleguillos, C.; Aguirre, C.; Barea, J.M.; Azcon, R. Growth promoting effect of two Sinorhizobium meliloti strains (a wild type and its genetically modified derivative) on a non-legume plant species in specific interaction with two arbuscular mycorrhizal fungi. Plant Sci. 2000, 159, 57–63. [Google Scholar] [CrossRef]
- Lazarovits, G.; Nowak, J. Rhizobacteria for improvement of plant growth and establishment. Hortscience 1997, 32, 188–192. [Google Scholar]
- Fallik, E.; Sarig, S.; Okon, Y. Morphology and physiology of plant roots associated with Azospirillum. In Azospirillum/Plant Associations; Okon, Y., Ed.; CRC Press Inc.: Boca Raton, FL, USA, 1994; pp. 77–83. [Google Scholar]
- Nowak, J.; Asiedu, S.K.; Lazarovits, G.; Pillay, V.; Stewart, A.; Smith, C.; Liu, Z. Enhancement of in Vitro Growth and Transplant Stress Tolerance of Potato and Vegetable Plantlets Co-cultured with a Plant Growth Promoting Pseudomonad Bacterium. In Proceedings of Ecophysiology and Photosynthetic in Vitro Cultures, Aix en Provence, France, 1–3 December 1995; Carre, F., Chagvardieff, P., Eds.; CEA: Aix en Provence, France, 1995; pp. 173–180. [Google Scholar]
- Nowak, J.; Bensalim, S.; Smith, C.; Dunbar, C.; Asiedu, S.; Madani, A.; Lazarovits, G.; Northcott, D.; Sturz, A. Behaviour of plant material issued from in vitro tuberization. Potato Res. 1999, 42, 505–519. [Google Scholar] [CrossRef]
- Bensalim, S.; Nowak, J.; Asiedu, S.K. A plant growth promoting rhizobacterium and temperature effects on performance of 18 clones of potato. Am. J. Potato Res. 1998, 75, 145–152. [Google Scholar] [CrossRef]
- Pillay, V.K.; Nowak, J. Inoculum density, temperature, and genotype effects onin vitro growth promotion and epiphytic and endophytic colonization of tomato (Lycopersicon esculentum L.) seedlings inoculated with a pseudomonad bacterium. Can. J. Microbiol. 1997, 43, 354–361. [Google Scholar] [CrossRef]
- Blom, D.; Fabbri, C.; Connor, E.C.; Schiestl, F.P.; Klauser, D.R.; Boller, T.; Eberl, L.; Weisskopf, L. Production of plant growth modulating volatiles is widespread among rhizosphere bacteria and strongly depends on culture conditions. Environ. Microbiol. 2011, 13, 3047–3058. [Google Scholar]
- Compant, S.; Nowak, J.; Coenye, T.; Clément, C.; Ait Barka, E. Diversity and occurrence of Burkholderia spp. in the natural environment. FEMS Microbiol. Rev. 2008, 32, 607–626. [Google Scholar] [CrossRef]
- Compant, S.; Clement, C.; Sessitsch, A. Plant growth-promoting bacteria in the rhizo- and endosphere of plants: Their role, colonization, mechanisms involved and prospects for utilization. Soil Biol. Biochem. 2010, 42, 669–678. [Google Scholar] [CrossRef] [Green Version]
- Ma, W.; Zalec, K.; Glick, B.R. Biological activity and colonization pattern of the bioluminescence-labeled plant growth-promoting bacterium Kluyvera ascorbata SUD165/26. FEMS Microbiol. Ecol. 2001, 35, 137–144. [Google Scholar] [CrossRef]
- Bonkowski, M. Protozoa and plant growth: The microbial loop in soil revisited. New Phytol. 2004, 162, 617–631. [Google Scholar]
- Bais, H.P. The role of root exudates in rhizosphere interactions with plants and other organisms. Annu. Rev. Plant Biol. 2006, 57, 233–266. [Google Scholar] [CrossRef]
- Hartmann, A.; Schmid, M.; van Tuinen, D.; Berg, G. Plant-driven selection of microbes. Plant Soil 2009, 321, 235–257. [Google Scholar] [CrossRef]
- Topp, G.C.; Watt, M.; Hayhoe, H.N. Point specific measurement and monitoring of soil water content with an emphasis on TDR. Can. J. Soil Sci. 1996, 76, 307–316. [Google Scholar] [CrossRef]
- Watt, M.; Silk, W.K.; Passioura, J.B. Rates of root and organism growth, soil conditions, and temporal and spatial development of the rhizosphere. Ann. Bot. 2006, 97, 839–855. [Google Scholar] [CrossRef]
- Jones, D.L.; Nguyen, C.; Finlay, R.D. Carbon flow in the rhizosphere: Carbon trading at the soil-root interface. Plant Soil 2009, 321, 5–33. [Google Scholar]
- Richardson, A.; Barea, J.-M.; McNeill, A.; Prigent-Combaret, C. Acquisition of phosphorus and nitrogen in the rhizosphere and plant growth promotion by microorganisms. Plant Soil 2009, 321, 305–339. [Google Scholar] [CrossRef]
- Bloemberg, G.V.; Lugtenberg, B.J.J. Molecular basis of plant growth promotion and biocontrol by rhizobacteria. Curr. Opin. Plant Biol. 2001, 4, 343–350. [Google Scholar] [CrossRef]
- Sturz, A.V.; Christie, B.R.; Matheson, B.G.; Nowak, J. Biodiversity of endophytic bacteria which colonize red clover nodules, roots, stems and foliage and their influence on host growth. Biol. Fertil. Soils 1997, 25, 13–19. [Google Scholar] [CrossRef]
- Haichar, F.E.; Marol, C.; Berge, O.; Rangel-Castro, J.I.; Prosser, J.I.; Balesdent, J.; Heulin, T.; Achouak, W. Plant host habitat and root exudates shape soil bacterial community structure. ISME J. 2008, 2, 1221–1230. [Google Scholar]
- Rudrappa, T.; Czymmek, K.J.; Pare, P.W.; Bais, H.P. Root-secreted malic acid recruits beneficial soil bacteria. Plant Physiol. 2008, 148, 1547–1556. [Google Scholar] [CrossRef]
- Van West, P.; Morris, B.M.; Reid, B.; Appiah, A.A.; Osborne, M.C.; Campbell, T.A.; Shepherd, S.J.; Gow, N.A.R. Oomycete plant pathogens use electric fields to target roots. Mol. Plant-Microbe Interact. 2002, 15, 790–798. [Google Scholar] [CrossRef]
- Schulz, S.; Dickschat, J.S. Bacterial volatiles: The smell of small organisms. Nat. Prod. Rep. 2007, 24, 814–842. [Google Scholar]
- Raaijmakers, J.M.; Paulitz, T.C.; Steinberg, C.; Alabouvette, C.; Moenne-Loccoz, Y. The rhizosphere: A playground and battlefield for soilborne pathogens and beneficial microorganisms. Plant Soil 2009, 321, 341–361. [Google Scholar] [CrossRef]
- Compant, S.; Duffy, B.; Nowak, J.; Clement, C.; Barka, E.A. Use of plant growth-promoting bacteria for biocontrol of plant diseases: Principles, mechanisms of action, and future prospects. Appl. Environ. Microbiol. 2005, 71, 4951–4959. [Google Scholar] [CrossRef]
- Burr, T.J.; Schroth, M.N.; Suslow, T. Increased potato yields by treatment of seed-pieces with specific strains of Pseudomonas fluorescens and P. Putida. Phytopathology 1978, 68, 1377–1383. [Google Scholar] [CrossRef]
- Brown, M.E. Plant growth substances produced by micro-organisms of soil and rhizosphere. J. Appl. Microbiol. 1972, 35, 443–451. [Google Scholar] [CrossRef]
- Sitnikov, D.M.; Schineller, J.B.; Baldwin, T.O. Transcriptional regulation of bioluminesence genes from Vibrio fischeri. Mol. Microbiol. 1995, 17, 801–812. [Google Scholar]
- Boller, T. Mechanisms involved in non-target effects of pesticides on soil-borne pathogens. In Pesticide Interactions in Crop Production: Beneficial and Deleterious Effects; Altman, J., Ed.; CRC Press: Boca Raton, FL, USA, 1993; pp. 281–301. [Google Scholar]
- Mark, G.L. Transcriptome profiling of bacterial responses to root exudates identifies genes involved in microbe-plant interactions. Proc. Natl. Acad. Sci. USA 2005, 102, 17454–17459. [Google Scholar] [CrossRef]
- Berg, G.; Smalla, K. Plant species and soil type cooperatively shape the structure and function of microbial communities in the rhizosphere. FEMS Microbiol. Ecol. 2009, 68, 1–13. [Google Scholar]
- Fierer, N.; Jackson, R.B. The diversity and biogeography of soil bacterial communities. Proc. Natl. Acad. Sci. USA 2006, 103, 626–631. [Google Scholar] [CrossRef]
- Revsbech, N.P.; Pedersen, O.; Reichardt, W.; Briones, A. Microsensor analysis of oxygen and pH in the rice rhizosphere under field and laboratory conditions. Biol. Fertil. Soils 1999, 29, 379–385. [Google Scholar] [CrossRef]
- Xu, H. Soil-root interface water potential in sweet corn as affected by organic fertilizer and a microbial inoculant. In Nature Farming and Microbial Applications; Xu, H., Parr, J.F., Umemura, H., Eds.; Food Products Press: Binghamton, NY, USA, 2000; pp. 139–156. [Google Scholar]
- Tavaria, F.K.; Zuberer, D.A. Effect of low pO2 on colonization of maize roots by a genetically altered Pseudomonas putida [PH6(L1019)]. Biol. Fertil. Soils 1997, 26, 43–49. [Google Scholar] [CrossRef]
- Wu, T.; Chellemi, D.O.; Graham, J.H.; Martin, K.J.; Rosskopf, E.N. Comparison of soil bacterial communities under diverse agricultural land management and crop production practices. Microb. Ecol. 2008, 55, 293–310. [Google Scholar]
- Carney, K.M.; Matson, P.A. The influence of tropical plant diversity and composition on soil microbial communities. Microb. Ecol. 2006, 52, 226–238. [Google Scholar] [CrossRef]
- Jangid, K.; Williams, M.A.; Franzluebbers, A.J.; Schmidt, T.M.; Coleman, D.C.; Whitman, W.B. Land-use history has a stronger impact on soil microbial community composition than aboveground vegetation and soil properties. Soil Biol. Biochem. 2011, 43, 2184–2193. [Google Scholar] [CrossRef]
- Bloemberg, G.V. Simultaneous imaging of WCS365 populations expressing three different autofluorescent proteins in the rhizosphere: New perspectives for studying microbial communities. Mol. Plant-Microbe Interact. 2000, 13, 1170–1176. [Google Scholar] [CrossRef]
- Marschner, P.; Yang, C.H.; Lieberei, R.; Crowley, D.E. Soil and plant specific effects on bacterial community composition in the rhizosphere. Soil Biol. Biochem. 2001, 33, 1437–1445. [Google Scholar] [CrossRef]
- Berg, G. Plant-microbe interactions promoting plant growth and health: Perspectives for controlled use of microorganisms in agriculture. Appl. Microbiol. Biotechnol. 2009, 84, 11–18. [Google Scholar]
- Andre, H.M.; Ducarme, X.; Lebrun, P. Soil biodiversity: Myth, reality or conning? Oikos 2002, 96, 3–24. [Google Scholar] [CrossRef]
- Odonnell, A.G.; Goodfellow, M.; Hawksworth, D.L. Theoretical and practical aspects of the quantification of biodiversity among microorganisms. Philos. Trans. R. Soc. Lond. Ser. B Biol. Sci. 1994, 345, 65–73. [Google Scholar]
- Smith, K.P.; Handelsman, J.; Goodman, R.M. Genetic basis in plants for interactions with disease-suppressive bacteria. Proc. Natl. Acad. Sci. USA 1999, 96, 4786–4790. [Google Scholar] [CrossRef]
- Loper, J.E.; Kobayashi, D.Y.; Paulsen, I.T. The genomic sequence of Pseudomonas fluorescens Pf-5: Insights into biological control. Phytopathology 2007, 97, 233–238. [Google Scholar] [CrossRef]
- Jacobsen, B.J.; Zidack, N.K.; Larson, B.J. The role of bacillus-based biological control agents in integrated pest management systems: Plant diseases. Phytopathology 2004, 94, 1272–1275. [Google Scholar] [CrossRef]
- Long, S.R. Genes and signals in the rhizobium-legume symbiosis. Plant Physiol. 2001, 125, 69–72. [Google Scholar]
- Harman, G.E.; Howell, C.R.; Viterbo, A.; Chet, I.; Lorito, M. Trichoderma species—Opportunistic, avirulent plant symbionts. Nat. Rev. Microbiol. 2004, 2, 43–56. [Google Scholar] [CrossRef]
- Conn, K.L.; Nowak, J.; Lazarovits, G. A gnotobiotic bioassay for studying interactions between potatoes and plant growth-promoting rhizobacteria. Can. J. Microbiol. 1997, 43, 801–808. [Google Scholar] [CrossRef]
- Liu, Z.; Pillay, V.; Nowak, J. In vitro culture of watermelon and cantaloupe with and without beneficial bacterium. Acta Hortic. 1995, 402, 58–60. [Google Scholar]
- Wang, B.; Qiu, Y.L. Phylogenetic distribution and evolution of mycorrhizas in land plants. Mycorrhiza 2006, 16, 299–363. [Google Scholar]
- Nagarajan, S. Abiotic tolerance and crop improvement. In Abiotic Stress Adaptation in Plants: Physiological, Molecular and Genomic Foundation; Pareek, A., Sopory, S.K., Bohnert, H.J., Govindjee, Eds.; Springer: Dordrecht, The Netherlands, 2010; pp. 1–11. [Google Scholar]
- Smith, S.E.; Read, D.J. Mycorrhizal Symbiosis, 3rd ed; Academic Press: New York, NY, USA, 2008; p. 523. [Google Scholar]
- Buscot, F. What are soils? In Microorganisms in Soils: Roles in Genesis and Functions; Buscot, F., Varma, A., Eds.; Springer: New York, NY, UAS, 2005; pp. 3–16. [Google Scholar]
- Nowak, J.; Shulaev, V. Priming for transplant stress resistance in in vitro propagation. In Vitro Cell. Dev. Biol. Plant 2003, 39, 107–124. [Google Scholar]
- Harwood, R. A history of sustainable agriculture. In Sustainable Agricultural Systems; Edwards, C.A., Lal, R., Madden, P., Miller, R.H., House, G., Eds.; Soil and Water Conservation Society: Ankeny, IA, USA, 1990; pp. 3–19. [Google Scholar]
- Shane, M.W.; Lambers, H. Cluster roots: A curiosity in context. Plant Soil 2005, 274, 101–125. [Google Scholar] [CrossRef]
- Jakobsen, I.; Abbott, L.K.; Robson, A.D. External hyphae of vesicular-arbuscular mycorrhizal fungi associated with Trifolium subterraneum L. New Phytol. 1992, 120, 371–380. [Google Scholar] [CrossRef]
- Barea, J.M.; Pozo, M.J.; Azcon, R.; Azcon-Aguilar, C. Microbial co-operation in the rhizosphere. J. Exp. Bot. 2005, 56, 1761–1778. [Google Scholar]
- Bonfante, P. Plants, mycorrhizal fungi and endobacteria: A dialog among cells and genomes. Biol. Bull. 2003, 204, 215–220. [Google Scholar] [CrossRef]
- Toljander, J.F.; Lindahl, B.D.; Paul, L.R.; Elfstrand, M.; Finlay, R.D. Influence of arbuscular mycorrhizal mycelial exudates on soil bacterial growth and community structure. FEMS Microbiol. Ecol. 2007, 61, 295–304. [Google Scholar] [CrossRef]
- Garbaye, J. Helper bacteria—A new dimension to the mycorrhizal symbiosis. New Phytol. 1994, 128, 197–210. [Google Scholar]
- Alphei, J.; Bonkowski, M.; Scheu, S. Protozoa, nematoda and lumbricidae in the rhizosphere of Hordelymus europeaus (poaceae): Faunal interactions, response of microorganisms and effects on plant growth. Oecologia 1996, 106, 111–126. [Google Scholar]
- Kuikman, P.J.; Jansen, A.G.; Vanveen, J.A. 15N-nitrogen mineralization from bacteria by protozoan grazing at different soil-moisture regimes. Soil Biol. Biochem. 1991, 23, 193–200. [Google Scholar]
- Spiertz, J.H.J. Nitrogen, sustainable agriculture and food security: A review. Agron. Sustain. Dev. 2010, 30, 43–55. [Google Scholar] [CrossRef]
- Adesemoye, A.; Kloepper, J. Plant–microbes interactions in enhanced fertilizer-use efficiency. Appl. Microbiol. Biotechnol. 2009, 85, 1–12. [Google Scholar] [CrossRef]
- Brady, N.C.; Weil, R.R. The Nature and Properties of Soils, 14th ed; Pearson Education: Upper Saddle River, NJ, USA, 2008; p. 965. [Google Scholar]
- Ogunseitan, O. Microbial Diversity: Form and Function in Prokaryotes; Blackwell Publishing: Malden, MA, USA, 2005; p. 292. [Google Scholar]
- Hernandez, G. Biological nitrogen fixation and sustainable agriculture. In Nitrogen Fixation: Global Perspectives; Finan, T.M., O’Brian, M.R., Layzell, D.B., Vessey, J.K., Newton, W., Eds.; CABI Publishing: New York, NY, USA, 2002; pp. 339–340. [Google Scholar]
- Malik, K.A.; Zafar, Y.; Bilal, R.; Azam, E. Use of 15N isotope dilution for quantification of N2 fixation associated with roots of kallar grass Leptochloa fusca (L.). Biol. Fertil. Soils 1987, 4, 103–108. [Google Scholar]
- Franche, C.; Lindstrom, K.; Elmerich, C. Nitrogen-fixing bacteria associated with leguminous and non-leguminous plants. Plant Soil 2009, 321, 35–59. [Google Scholar]
- Okon, Y. Azospirillum as a potential inoculant for agriculture. Trends Biotechnol. 1985, 3, 223–228. [Google Scholar] [CrossRef]
- Van Berkum, P.; Bohlool, B.B. Evaluation of nitrogen fixation by bacteria in association with roots of tropical grasses. Microbiol. Rev. 1980, 44, 491–517. [Google Scholar]
- Kouas, S.; Labidi, N.; Debez, A.; Abdelly, C. Effect of P on nodule formation and N fixation in bean. Agron. Sustain. Dev. 2005, 25, 389–393. [Google Scholar] [CrossRef]
- Chen, Y.P.; Rekha, P.D.; Arun, A.B.; Shen, F.T.; Lai, W.A.; Young, C.C. Phosphate solubilizing bacteria from subtropical soil and their tricalcium phosphate solubilizing abilities. Appl. Soil Ecol. 2006, 34, 33–41. [Google Scholar] [CrossRef]
- Smith, S.E.; Smith, F.A.; Jakobsen, I. Functional diversity in arbuscular mycorrhizal (AM) symbioses: The contribution of the mycorrhizal P uptake pathway is not correlated with mycorrhizal responses in growth or total P uptake. New Phytol. 2004, 162, 511–524. [Google Scholar]
- Goldstein, A.H.; Rogers, R.D.; Mead, G. Mining by microbe. Nat. Biotechnol. 1993, 11, 1250–1254. [Google Scholar]
- Unno, Y.; Okubo, K.; Wasaki, J.; Shinano, T.; Osaki, M. Plant growth promotion abilities and microscale bacterial dynamics in the rhizosphere of lupin analysed by phytate utilization ability. Environ. Microbiol. 2005, 7, 396–404. [Google Scholar] [CrossRef]
- Tinker, P.B.; Nye, P.H. Solute Movement in the Rhizosphere; Oxford University Press: New York, NY, USA, 2000. [Google Scholar]
- Jones, D.L.; Darrah, P.R. Role of root derived organic-acids in the mobilization of nutrients from the rhizosphere. Plant Soil 1994, 166, 247–257. [Google Scholar] [CrossRef]
- Wallace, R.J. Antimicrobial properties of plant secondary metabolites. Proc. Nutr. Soc. 2004, 63, 621–629. [Google Scholar]
- Sakakibara, H. Cytokinins: Activity, biosynthesis, and translocation. Annu. Rev. Plant Biol. 2006, 57, 431–449. [Google Scholar] [CrossRef]
- Barazani, O.; Friedman, J. Is IAA the major root growth factor secreted from plant-growth-mediating bacteria? J. Chem. Ecol. 1999, 25, 2397–2406. [Google Scholar] [CrossRef]
- Patten, C.L.; Glick, B.R. Bacterial biosynthesis of indole-3-acetic acid. Can. J. Microbiol. 1996, 42, 207–220. [Google Scholar]
- Leveau, J.H.J.; Gerards, S. Discovery of a bacterial gene cluster for catabolism of the plant hormone indole 3-acetic acid. FEMS Microbiol. Ecol. 2008, 65, 238–250. [Google Scholar] [CrossRef]
- Hartmann, H.T. Plant Propagation: Principles and Practices, 6th ed; Prentice Hall: Upper Saddle River, NJ, USA, 1997; p. 770. [Google Scholar]
- Mañero, F.J.G.; Acero, N.; Lucas, J.A.; Probanza, A. The infuence of native rhizobacteria on European alder (Alnus glutinosa (L.) Gaertn.) growth. Plant Soil 1996, 182, 67–74. [Google Scholar] [CrossRef]
- Schmulling, T. New insights into the functions of cytokinins in plant development. J. Plant Growth Regul. 2002, 21, 40–49. [Google Scholar]
- Glick, B.R. Modulation of plant ethylene levels by the bacterial enzyme ACC deaminase. FEMS Microbiol. Lett. 2005, 251, 1–7. [Google Scholar]
- Arshad, M. Ethylene: Agricultural Sources and Applications; Kluwer Academic/Plenum Publishers: New York, NY, USA, 2002. [Google Scholar]
- Shah, S.; Li, J.P.; Moffatt, B.A.; Glick, B.R. Isolation and characterization of ACC deaminase genes from two different plant growth-promoting rhizobacteria. Can. J. Microbiol. 1998, 44, 833–843. [Google Scholar]
- Campbell, B.G.; Thomson, J.A. 1-aminocyclopropane-1-carboxylate deaminase genes from Pseudomonas strains. FEMS Microbiol. Lett. 1996, 138, 207–210. [Google Scholar] [CrossRef]
- Wang, C.X.; Knill, E.; Glick, B.R.; Defago, G. Effect of transferring 1-aminocyclopropane-1-carboxylic acid (ACC) deaminase genes into Pseudomonas fluorescens strain CHA0 and its gaca derivative CHA96 on their growth-promoting and disease-suppressive capacities. Can. J. Microbiol. 2000, 46, 898–907. [Google Scholar]
- Hao, Y.; Charles, T.C.; Glick, B.R. ACC deaminase from plant growth-promoting bacteria affects crown gall development. Can. J. Microbiol. 2007, 53, 1291–1299. [Google Scholar]
- Toklikishvili, N.; Dandurishvili, N.; Vainstein, A.; Tediashvili, M.; Giorgobiani, N.; Lurie, S.; Szegedi, E.; Glick, B.; Chernin, L. Inhibitory effect of ACC deaminase-producing bacteria on crown gall formation in tomato plants infected by agrobacterium tumefaciens or A. vitis. Plant Pathol. 2010, 59, 1023–1030. [Google Scholar] [CrossRef]
- Cheng, Z.Y.; Park, E.; Glick, B.R. 1-aminocyclopropane-1-carboxylate deaminase from Pseudomonas putida UW4 facilitates the growth of canola in the presence of salt. Can. J. Microbiol. 2007, 53, 912–918. [Google Scholar] [CrossRef]
- Farwell, A.J.; Vesely, S.; Nero, V.; Rodriguez, H.; McCormack, K.; Shah, S.; Dixon, D.G.; Glick, B.R. Tolerance of transgenic canola plants (Brassica napus) amended with plant growth-promoting bacteria to flooding stress at a metal-contaminated field site. Environ. Pollut. 2007, 147, 540–545. [Google Scholar] [CrossRef]
- Bonaterra, A.; Ruz, L.; Badosa, E.; Pinochet, J.; Montesinos, E. Growth promotion of Prunus rootstocks by root treatment with specific bacterial strains. Plant Soil 2003, 255, 555–569. [Google Scholar] [CrossRef]
- Cook, R.J.; Baker, K.F. The Nature and Practice of Biological Control of Plant Pathogens; American Phytopathological Society: St. Paul, MN, USA, 1983; p. 539. [Google Scholar]
- Lockwood, J.L. Biological control of soil-borne plant pathogens. In Relation of Energy Stress to Behaviour of Soil-borne Plant Pathogens and to Disease Development; Hornby, D., Ed.; CAB International: Wallingford, UK, 1990; pp. 197–214. [Google Scholar]
- Duijff, B.J.; Meijer, J.W.; Bakker, P.; Schippers, B. Siderophoree-mediated competition for iron and induced resistance in the suppression of fusarium-wilt of carnation by fluorescent Pseudomonas spp. Neth. J. Plant Pathol. 1993, 99, 277–289. [Google Scholar] [CrossRef]
- Van Loon, L. Plant responses to plant growth-promoting rhizobacteria. Eur. J. Plant Pathol. 2007, 119, 243–254. [Google Scholar] [CrossRef]
- Chen, C.; Bauske, E.M.; Musson, G.; Rodriguezkabana, R.; Kloepper, J.W. Biological-control of fusarium-wilt on cotton by use of endophytic bacteria. Biol. Control 1995, 5, 83–91. [Google Scholar] [CrossRef]
- Maurhofer, M.; Keel, C.; Schnider, U.; Voisard, C.; Haas, D.; Defago, G. Influence of enhanced antibiotic production in Pseudomonas fluorescens strain-CHA0 on its disease suppressive capacity. Phytopathology 1992, 82, 190–195. [Google Scholar] [CrossRef]
- Eriksson, A.; Anfora, G.; Lucchi, A.; Lanzo, F.; Virant-Doberlet, M.; Mazzoni, V. Exploitation of insect vibrational signals reveals a new method of pest management. PLoS ONE 2012, 7, e32954. [Google Scholar]
- Bruinsma, M.; Kowalchuk, G.A.; van Veen, J.A. Effects of genetically modified plants on microbial communities and processes in soil. Biol. Fertil. Soils 2003, 37, 329–337. [Google Scholar]
- Kowalchuk, G.A.; Bruinsma, M.; van Veen, J.A. Assessing responses of soil microorganisms to GM plants. Trends Ecol. Evol. 2003, 18, 403–410. [Google Scholar] [CrossRef]
- Fred, E.B.; Baldwin, I.L.; McCoy, E. Root Nodule Bacteria and Leguminous Plants; University of Wisconsin Madison: Madison, WI, USA, 1932; Volume 5, p. 343. [Google Scholar]
- Phan Thi, C.; Tran Dang, D.; Tran Minh, H.; Nguyen Thanh, H.; Choudhury, A.T.M.A.; Kecskés, M.L.; Kennedy, I.R. Inoculant plant growth-promoting microorganisms enhance utilisation of urea-N and grain yield of paddy rice in southern vietnam. Eur. J. Soil Biol. 2009, 45, 52–61. [Google Scholar] [CrossRef]
- Kennedy, I.R.; Choudhury, A.T.M.A.; Kecskés, M.L.; Rose, M.T. (Eds.) Efficient Nutrient Use in Rice Production in Vietname Achieved Using Inoculant Biofertilisers. In Proceedings of a Project (SMCN/2002/073) Workshop, Hanoi, Vietnam, 12–13 October 2007; ACIAR Proceedings: Hanoi, Vietnam, 2007; 130.
- Whipps, J.M.; McQuilken, M.P. Aspects of biocontrol of fungal plant pathogens. In Exploitation of Microorganisms; Jones, D.G., Ed.; Chapman & Hall: London, UK, 1993; pp. 45–79. [Google Scholar]
- Schroth, M.N.; Becker, J.O. Concepts of ecological and physiological activities of rhizobacteria related to biological control and plant growth promotion. In Biological Control of Soil-Borne Plant Pathogens; Hornby, D., Ed.; CAB International: Wallingford, UK, 1990; pp. 389–414. [Google Scholar]
- Mark, G.L.; Murphy, J.; Cassells, A.C. Microbial characterisation and preparation of inoculum for in vitro mycorrhization of strawberry in autotophic culture. In Pathogen and Microbial Contamination Management in Micropropagation; Cassells, A.C., Ed.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 1997; Volume 12, pp. 345–350. [Google Scholar]
- Sturz, A.V. The role of endophytic bacteria during seed piece decay and potato tuberization. Plant Soil 1995, 175, 257–263. [Google Scholar]
- Yachi, S.; Loreau, M. Biodiversity and ecosystem productivity in a fluctuating environment: The insurance hypothesis. Proc. Natil. Acad. Sci. USA 1999, 96, 1463–1468. [Google Scholar] [CrossRef]
- Lal, R. Soils and sustainable agriculture: A review. Agron. Sustain. Dev. 2008, 28, 57–64. [Google Scholar]
- Irrigation & Water Technologies Pty. Ltd. Web site. Available online: http://www.kisss.com.au/ (accessed on 8 December 2012).
- Lamm, F.R. Advantages and Disadvantages of Subsurface Drip Irrigation. In Proceedings of International Meeting on Advances in Drip/Micro Irrigation, Puerto de La Cruz, Tenerife, Canary Islands, Spain, 2–5 December 2002; Instituto Canario de Investigaciones Agrarias: Canary Islands, Spain, 2002. [Google Scholar]
- Mazzei Web site. Available online: www.mazzei.net (accessed on 8 December 2012).
- Bhattarai, S.P.; Pendergast, L.; Midmore, D.J. Root aeration improves yield and water use efficiency of tomato in heavy clay and saline soils. Sci. Hortic. 2006, 108, 278–288. [Google Scholar]
- Rose, M.; Deaker, R.; Potard, S.; Tran, C.; Vu, N.; Kennedy, I. The survival of plant growth promoting microorganisms in peat inoculant as measured by selective plate counting and enzyme-linked immunoassay. World J. Microbiol. Biotechnol. 2011, 27, 1649–1659. [Google Scholar] [CrossRef]
- Bécard, G.; Piché, Y. Fungal growth stimulation by CO2 and root exudates in vesicular-arbuscular mycorrhizal symbiosis. Appl. Environ. Microbiol. 1989, 55, 2320–2325. [Google Scholar]
- Welbaum, G.E.; Shen, Z.X.; Watkinson, J.I.; Wang, C.L.; Nowak, J. Priming soilless growing medium with disaccharides stimulated microbial biofilm formation, and increased particle aggregation and moisture retention during muskmelon transplant production. J. Am. Soc. Hortic. Sci. 2009, 134, 387–395. [Google Scholar]
- Chanway, C.P. Inoculation of tree roots with plant growth promoting soil bacteria: An emerging technology for reforestation. For. Sci. 1997, 43, 99–112. [Google Scholar]
- Sticher, L.; MauchMani, B.; Metraux, J.P. Systemic acquired resistance. Annu. Rev. Phytopathol. 1997, 35, 235–270. [Google Scholar]
- Benhamou, N.; Kloepper, J.W.; QuadtHallman, A.; Tuzun, S. Induction of defense-related ultrastructural modifications in pea root tissues inoculated with endophytic bacteria. Plant Physiol. 1996, 112, 919–929. [Google Scholar]
- Sharma, V.K.; Nowak, J. Enhancement of verticillium wilt resistance in tomato transplants by in vitro co-culture of seedlings with a plant growth promoting rhizobacterium (Pseudomonas sp. strain PsJN). Can. J. Microbiol. 1998, 44, 528–536. [Google Scholar]
- Fernandez, O.; Theocharis, A.; Bordiec, S.; Feil, R.; Jacquens, L.; Clément, C.; Fontaine, F.; Barka, E.A. Burkholderia phytofirmans PsJN acclimates grapevine to cold by modulating carbohydrate metabolism. Mol. Plant-Microbe Interact. 2012, 25, 496–504. [Google Scholar] [CrossRef]
- Fernandez, O.; Vandesteene, L.; Feil, R.; Baillieul, F.; Lunn, J.; Clément, C. Trehalose metabolism is activated upon chilling in grapevine and might participate in Burkholderia phytofirmans induced chilling tolerance. Planta 2012, 236, 355–369. [Google Scholar] [CrossRef]
- Weller, D.M.; Raaijmakers, J.M.; Gardener, B.B.M.; Thomashow, L.S. Microbial populations responsible for specific soil suppressiveness to plant pathogens. Annu. Rev. Phytopathol. 2002, 40, 309–348. [Google Scholar] [CrossRef]
- Coombs, A. Seeds Inherit Memories of Enemies. The Scientist 2012. [Google Scholar]
- Techy, L.; Woolsey, C.A.; Schmale, D.G. Monitoring the spread of a plant pathogen in the lower atmosphere using unmanned aerial vehicles and a bouyancy-controlled weather balloon. In Aerospace Technology Conference and Exposition; SAE: Seattle, WA, USA, 2009. [Google Scholar]
- Murakami, T.; Yui, M.; Amaha, K. Canopy height measurement by photogrammetric analysis of aerial images: Application to buckwheat (Fagopyrum esculentum Moench) lodging evaluation. Comput. Electron. Agric. 2012, 89, 70–75. [Google Scholar] [CrossRef]
- Luquet, D.; Bégué, A.; Vidal, A.; Clouvel, P.; Dauzat, J.; Olioso, A.; Gu, X.F.; Tao, Y. Using multidirectional thermography to characterize water status of cotton. Remote Sens. Environ. 2003, 84, 411–421. [Google Scholar] [CrossRef]
- Grace, J.; Nichol, C.; Disney, M.; Lewis, P.; Quaife, T.; Bowyer, P. Can we measure terrestrial photosynthesis from space directly, using spectral reflectance and fluorescence? Glob. Change Biol. 2007, 13, 1484–1497. [Google Scholar] [CrossRef]
- Rascher, U.; Nichol, C.J.; Small, C.; Hendricks, L. Monitoring spatio-temporal dynamics of photosynthesis with a portable hyperspectral imaging system. Photogramm. Eng. Remote Sens. 2007, 73, 45–56. [Google Scholar]
- Scharte, J.; Schon, H.; Weis, E. Photosynthesis and carbohydrate metabolism in tobacco leaves during an incompatible interaction with Phytophthora nicotianae. Plant Cell Environ. 2005, 28, 1421–1435. [Google Scholar] [CrossRef]
- Wen, C.; Guyer, D. Image-based orchard insect automated identification and classification method. Comput. Electron. Agric. 2012, 89, 110–115. [Google Scholar] [CrossRef]
- Lindenthal, M.; Steiner, U.; Dehne, H.W.; Oerke, E.C. Effect of downy mildew development on transpiration of cucumber leaves visualized by digital infrared thermography. Phytopathology 2005, 95, 233–240. [Google Scholar] [CrossRef]
- Zhao, Y.; He, Y.; Xu, X. A novel algorithm for damage recognition on pest-infested oilseed rape leaves. Comput. Electron. Agric. 2012, 89, 41–50. [Google Scholar]
- Sessitsch, A.; Weilharter, A.; Gerzabek, M.H.; Kirchmann, H.; Kandeler, E. Microbial population structures in soil particle size fractions of a long-term fertilizer field experiment. Appl. Environ. Microbiol. 2001, 67, 4215–4224. [Google Scholar] [CrossRef]
- Sessitsch, A.; Howieson, J.G.; Perret, X.; Antoun, H.; Martínez-Romero, E. Advances in Rhizobium research. Crit. Rev. Plant Sci. 2002, 21, 323–378. [Google Scholar] [CrossRef]
- Ogata, K. System Dynamics, 4th ed; Pearsons Education, Inc.: Englewood Cliffs, NJ, USA, 2004; p. 768. [Google Scholar]
- Hay, R.; Porter, J. The Physiology of Crop Yield, 2nd ed; Blackwell Publishing Ltd.: Oxford, UK, 2006; p. 314. [Google Scholar]
- Yin, X.; Struik, P.C. Modelling the crop: From system dynamics to systems biology. J. Exp. Bot. 2010, 61, 2171–2183. [Google Scholar] [CrossRef]
- Yin, X.; Struik, P.C. Applying modelling experiences from the past to shape crop systems biology: The need to converge crop physiology and functional genomics. New Phytol. 2008, 179, 629–642. [Google Scholar] [CrossRef]
- Masuka, B.; Araus, J.L.; Das, B.; Sonder, K.; Cairns, J.E. Phenotyping for abiotic stress tolerance in miaze. J. Integr. Plant Biol. 2012, 54, 238–249. [Google Scholar]
- Wang, S.; Min, J.; Yi, B. Location Based Services for Mobiles: Technologies and Standards. In Proceedings of IEEE International Conference on Communication (ICC), Beijing, China, 19–23 May 2008; pp. 35–38.
- Moxley, T. Technology Takes Root at Virginia Tech’s Catawba Sustainability Center “Smart Farm”. The Roanoke Times 2011. [Google Scholar]
- Sij, J.W.; Bordovsky, D.G.; Jones, D.L.; Slosser, J.E. Utilizing subsurface drip irrigation and conservation tillage in cotton production systems. SRX Agric. 2010, 2010, 1–6. [Google Scholar]
- Camp, C.; Lamm, F.; Evans, R.; Phene, C. Subsurface Drip Irrigation—Past, Present, and Future. In Proceedings of the 4th Decennial National Irrigation Symposium, Phoenix, AZ, USA, 14–16 November 2000; pp. 14–16.
- Metzker, M.L. Sequencing technologies—The next generation. Nat. Rev. Genet. 2009, 11, 31–46. [Google Scholar] [CrossRef]
© 2012 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Weekley, J.; Gabbard, J.; Nowak, J. Micro-Level Management of Agricultural Inputs: Emerging Approaches. Agronomy 2012, 2, 321-357. https://doi.org/10.3390/agronomy2040321
Weekley J, Gabbard J, Nowak J. Micro-Level Management of Agricultural Inputs: Emerging Approaches. Agronomy. 2012; 2(4):321-357. https://doi.org/10.3390/agronomy2040321
Chicago/Turabian StyleWeekley, Jonathan, Joseph Gabbard, and Jerzy Nowak. 2012. "Micro-Level Management of Agricultural Inputs: Emerging Approaches" Agronomy 2, no. 4: 321-357. https://doi.org/10.3390/agronomy2040321