
Response of the Endophytic Microbial Composition in Amaranthus Roots to Different Fertilization Treatments

Guangxi Key Laboratory of Agro-Environment and Agro-Products Safety, National Demonstration Center for Experimental Plant Science Education, Agricultural College, Guangxi University, Nanning 530004, China
*
Author to whom correspondence should be addressed.
Agronomy 2024, 14(5), 965; https://doi.org/10.3390/agronomy14050965
Submission received: 1 April 2024
/
Revised: 25 April 2024
/
Accepted: 1 May 2024
/
Published: 3 May 2024
(This article belongs to the Section Soil and Plant Nutrition)
Abstract
:To explore how endophytic microbial compositions in amaranth roots are influenced by various fertilization methods and to determine whether these microbes are associated with amaranthin formation, we conducted an analysis of the endophytic microbial community structure. The roots of amaranth plants subjected to different fertilization treatments—conventional fertilization without potassium (NP), conventional fertilization without phosphorus (NK), conventional fertilization without nitrogen (PK), and balanced fertilization (NPK)—were examined. The results showed that the proportions of Streptomyces, Actinospica, and Burkholderia-Caballeronia-Paraburkholderia in the amaranth roots under the balanced fertilization (NPK) treatment were all greater than those in the amaranth roots under the nitrogen (PK), phosphorus (NK), and potassium (NP) deficiency fertilization treatments. In contrast, the proportions of Phenylobacterium, Acrocalymma, Neocosmospora, Fusarium, Acidovorax, Gibellulopsis, Cladosporium, Dactylonectria, and Gibberella in the amaranth roots under the nutrient deficiency fertilization (NP, NK, and PK) treatments were higher than those in the amaranth roots under the balanced fertilization treatment. Additionally, a significantly positive correlation was found between Streptomyces and the amaranthin content. Furthermore, Acrocalymma, Neocosmospora, and Fusarium exhibited significantly negative correlations with the amaranthin content. The above results suggested that endophytes could easily colonize in amaranth roots as beneficial microorganisms under balanced fertilization conditions. In other words, the balanced fertilization (N, P and K fertilizers are 188.0, 53.0 and 50.0 kg·hm−2, respectively) could recruit more beneficial endogenous microorganisms in amaranth roots for improving their growth and quality.
1. Introduction
Amaranth (Amaranthus tricolor L.) is an annual vegetable renowned for its abundance of carotene, vitamins, and minerals, encompassing a diverse array of plant pigments, such as chlorophyll, amaranthin, and carotenoids [1,2,3,4]. Currently, its cultivation is expanding across Asia, North America, and Europe [5]. As an important vegetable crop in agricultural production, amaranth occupies significant portions of the cultivation area and the consumer market. However, with the modernization of agriculture and the widespread use of fertilizers, the issue of excessive fertilization has become increasingly prominent [6,7]. Excessive fertilizer application may lead to problems such as soil and water pollution, as well as impaired plant growth, thereby affecting both the yield and quality of amaranth [8,9]. Therefore, studying an appropriate fertilization scheme to enhance the health and increase the yield of amaranth is of paramount importance.
Endophytic microorganisms are pivotal contributors to plant growth and development because of their sheer abundance and diversity [10,11]. Notably, endophytic bacteria exhibit a repertoire of functions, including phosphorus solubilization, nitrogen fixation, iron carrier production, the synthesis of antibacterial agents, and the generation of plant growth hormones [12]. Their multifaceted roles include not only serving as facilitators of host plant growth but also actively contributing to nutrient accumulation and enhanced stress resistance, such as in disease, pest, or drought stress [13]. For example, endophytes can establish reciprocal interactions with host plants by exchanging nutrients and producing antibiotics and other substances to protect plants from stressful conditions [14]. The endophytic microbial composition of the plant root system is an important determinant of these symbiotic relationships, which are significantly influenced by soil type, field management (irrigation, fertilizer and pesticide application, etc.), and host genotype [15,16,17,18].
The impact of fertilization on endophytic microorganisms is intricate and profound, significantly altering the microbial composition within plant roots [19]. Research indicates that different fertilization methods can induce variations in the structure of endophytic microbial communities, thereby impacting plant growth, development, and stress resistance [20]. For instance, nitrogen, phosphorus, and potassium fertilizers can modify the pH and chemical properties of the soil environment, thus influencing the survival and activity of endophytic microorganisms [21]. Moreover, plant root nutrient secretion under various fertilization conditions can also affect the diversity and abundance of endophytic microorganisms [22]. For example, Pedraza et al. [23] observed that nitrogen fertilization markedly influenced the diversity of endophytic bacteria in plants, with greater microbial community diversity observed with moderate nitrogen fertilization. Similarly, Liu et al. [24] noted that moderate nitrogen fertilization in rice led to an increased diversity and abundance of endophytic bacteria and fungi compared to rice subjected to insufficient or excessive nitrogen inputs. Furthermore, excessive fertilization has been linked to a decreased diversity of endophytic bacteria in wheat roots, along with a reduction in the abundance of potential biocontrol bacteria, such as Bacillus, Streptomyces, and Burkholderia. Additionally, excessive fertilization suppressed antibiotic biosynthesis, exacerbating the occurrence of wheat crown rot [25]. Therefore, investigating the impact of balanced fertilization on the structure of plant endophytic microbial communities is of great significance.
Amaranthin, a pioneering natural colorant, finds application in various food systems, such as beverages, candy, cosmetics, and wine [26]. Acting as a colorant, amaranthin serves as a potent antioxidant, efficiently scavenging free radicals [27]. This dual function enhances its coloring properties and alleviates oxidative stress in plants. Antioxidants, crucial in plants, counteract harmful free radicals from physiological and environmental sources, aiding plants in combating stressors such as ultraviolet radiation, drought, and pollution, thereby enhancing overall plant health and resilience [28,29]. Previous studies have shown that endophytic microorganisms, as producers of plant hormones, can influence the synthesis of pigments (such as amaranthin) by producing hormones [30,31]. Meanwhile, betalain (amaranthin is a category of betalain) can also be produced by endophytic microorganisms, including certain fungal lineages and some nitrogen-fixing bacteria, such as basidiomycetes and Gluconacetobacter diazotrophicus [32].
Previous studies have focused mainly on the rhizosphere soil microbial composition under different fertilization treatments. Therefore, this study investigated the changes in endophytic bacterial and fungal community structures in the roots of amaranth under four different fertilization treatments. Specifically, the objectives of this study were to determine (1) the variations in the endophytic microbial community structure in amaranth roots under different fertilization treatments and (2) the correlation between endophytic microorganisms and amaranthin.
2. Materials and Methods
2.1. Field Site, Test Material, and Fertilizer Descriptions
The experiment was initiated on February 10, 2021, at the vegetable base of the Agricultural College, Guangxi University (108°17′15″ E and 22°51′02″ N). The soil type was red loam with 8.42 g·kg−1 of organic matter and a pH of 5.71. The total nitrogen, phosphorus, and potassium concentrations were 0.51 g·kg−1, 0.67 g·kg−1, and 7.21 g·kg−1, respectively. The available phosphorus, potassium, and alkali hydrolyzable nitrogen contents were 0.59 mg·kg−1, 51.01 mg·kg−1, and 13.17 mg·kg−1, respectively.
The Red Leaf 262 (Shenzhen Xiliang Seed Co., Ltd., Shenzhen, China) amaranth (Amaranthus tricolor L.) variety was used, and the seeds were purchased from the Nanning Vegetable Seed Market. Urea (46.4% N content), monoammonium phosphate (10% N content and 43% P2O5 content), and potassium sulfate (51% K2O content) were used as the N, P, and K fertilizers, respectively. They were all purchased at the Agricultural Material Market, Nanning City, from the Anhui Jinqiu Fertilizer Company (Fuyang City, Anhui Province, China).
2.2. Experimental Design
Four treatments were set up in this experiment, i.e., conventional fertilization without potassium (NP), conventional fertilization without phosphorus (NK), conventional fertilization without nitrogen (PK), and balanced fertilization (NPK), and three replications were carried out for each treatment. The experiment was carried out in pots with a 16 cm radius and a 20 cm height, and 15 kg of red loam soil was filled in each pot. In January 2021, indoor breeding was conducted as follows: Healthy, dry seeds were selected and soaked in 50 °C warm water for 20 min with continuous stirring. Subsequently, the water was cooled to 20–25 °C, and the seeds were soaked for 4–8 h. The seeds were then placed in a constant-temperature incubator at 25 °C for germination. When the germination rate reached 80%, the seedlings were transferred to nutrient-rich soil, grown until they developed 2–3 leaves, and then transplanted into potting soil. The entire amaranth cultivation process was carried out in a sunlight greenhouse. The daytime temperature in the greenhouse ranged from 25 to 30 °C, while the nighttime temperature ranged from 16 to 18 °C. The humidity level was maintained between 60% and 80%. Watering was carried out once a week, and manual weeding was performed regularly. The main diseases and pests of amaranth include stem rot and damping-off disease. A 25% carbendazim solution was used at a dilution of 600 times, typically sprayed once every 7–10 days and 2–3 consecutive times. All fertilization treatments and weeding, irrigation, and pest control measures were consistent across the experimental groups. The optimal fertilization rates for amaranth in terms of N, P, and K are 188, 53, and 50 kg/hm2, respectively [33], and all fertilizers were applied as basal fertilizers. The application rates are shown in Table 1.
2.3. Sample Collection
Sampling was conducted when the amaranth entered maturity. Briefly, a pot with an amaranth plant was turned upside down to remove the whole plant, and then the soil adhering to the roots was carefully shaken off. The roots were rinsed with sterile water several times to obtain root samples, and then the root samples were placed in sterile plastic bags filled with ice and transported to the laboratory.
In the laboratory, the root samples were rinsed with sterile water to remove impurities and adherents; subsequently, they were washed with 75% ethanol and a 1% NaClO solution for 1 min and 3 min, respectively. Then, they were washed 3 times using sterile water for 0.5 min. To determine the success of amaranth root surface sterilization, 100 µL of the last sterile aqueous wash was placed on Luria–Bertani (LB) agar plates (g/L) (NaCl-10, trypsin-5, yeast extract-5, and agar-20) and incubated for 7 days at 25 °C. As there were no colonies on the plate, disinfection was recorded as complete [34].
2.4. Analysis of the Amaranthin Content
First, 0.2 g of the leaves were weighed and ground uniformly with a small amount of methanol to a homogenous state. The precipitate was then centrifuged for 10 min (4 °C, 8000 r/min), the supernatant was discarded, and precipitates were obtained three times in succession. The precipitate was then extracted with 15 mL of pure water for 30 min and centrifuged for 15 min (4 °C, 8000 r/min). The supernatant was taken for a colorimetric analysis using a spectrophotometer (Thermo Fisher Scientific, Waltham, MA, USA) at 538 nm. The content of amaranthin was calculated according to Pitelli’s [35] method.
2.5. Analysis of the Endophytic Microbial Compositions
The total DNA was extracted according to the instructions of an E.Z.N.A. DNA Kit (Omega, Norcross, GA, USA). The DNA concentration and purity were detected with a NanoDrop2000 spectrophotometer (Thermo Fisher Scientific, USA), and the extracted genomic DNA was detected via 1% agarose gel electrophoresis. The extracted root microbial DNA was used as a template, and the primers 799F (5′-AACMGGATTAGATACCCKG-3′) and 1193R (5′-ACGTCATCCCCACCTTCC-3′) were used for the PCR amplification of the 16S rRNA V6 region of the endophytic bacteria. The primers ITS1F (5′-CTTGGTCATTTAGAGGAAGTAA-3′) and ITS2F (5′-GCTGCGTTCTTCATCATCGATFC-3′) were used for the PCR amplification of the ITS rRNA region of the endophytic fungi. The PCR apparatus used was an ABI GeneAmp® 9700(Thermo Scientific, Wilmington, DE, USA). PCR products were recovered via 2% agarose gel electrophoresis, purified with an Axy PrepDNA Gel Recovery Kit (AXYGEN)(Axygen, San Francisco, CA, USA), and eluted with Tris_HCl. The PCR products were detected and quantified using a QuantiFluor™-ST blue fluorescence quantitative system (Promega, Madison, WI, USA). According to the Illumina MiSeq platform standard operating procedures, purified amplified fragments were constructed into libraries. Sequencing was performed using the MiSeq PE300 and MiSeq PE250 platforms from Illumina (Shanghai Meggie Biomedical Technology Co., Ltd., Shanghai, China).
2.6. Statistical Analyses
The experimental data were analyzed using Excel 2019 and IBM SPSS 21. All the results are shown as the means with their standard deviation (mean ± SD). Mothur is an open-source software package used for processing, analyzing, and visualizing DNA sequence data. In this study, Mothur (version v.1.30.2, https://mothur.org/wiki/calculators/, accessed on 10 November 2023) was employed to calculate the diversity (Shannon index) and richness (ACE index) of the endophytic bacterial and fungal communities in amaranth roots. A principal coordinate analysis (PCoA) was conducted using the vegan 2.6.4 package [36] in R based on the Bray–Curtis distance. Then, partial least squares discriminant analysis (PLS-DA) was conducted using the mixOmics package in R (version 3.3.1) (http://fiehnlab.ucdavis.edu/staff/kind/Statistics/Concepts/OPLS-PLSDA, accessed on 10 November 2023). Endophytic microbial community composition and Venn diagrams were generated using the R language (version 3.3.1) tool for mapping and analysis. Linear discriminant analysis (LDA) is a technique used in multivariate statistical analyses and pattern recognition to measure the differences between data points within the same category. Linear discriminant analysis (LDA) was performed using LEfSe (http://huttenhower.sph.harvard.edu/galaxy/root?tool_id=lefse_upload, accessed on 10 November 2023) on samples, according to different grouping conditions based on taxonomic composition, to identify the differences in endophytic bacteria and fungi in the root system of amaranth. The networkx (version 1.11) tool was used to determine the correlation between microorganisms and between amaranthin and endophytic bacteria and fungi. Online data analyses were carried out using the Majorbio Cloud Platform (http://www.majorbio.com, accessed on 29 September 2022) of Majorbio Bio-Pharm Technology Co., Ltd. (Shanghai, China).
3. Results
3.1. Effect of Different Fertilization Treatments on the Amaranthin Content
As shown in Table 2, the amaranthin contents were not significantly different among the NK, PK and NPK treatments, except in the NP treatment. However, it was significantly greater in amaranth under the NPK treatment than those in amaranth under the conventional fertilization without potassium (NP). As shown in Table 2, the amaranthin contents were not significantly different among the NK, PK, and NPK treatments. However, it was significantly greater in amaranth under the NPK treatment than in amaranth under conventional fertilization without potassium (NP).
3.2. Effect of Different Fertilization Treatments on the Composition of Endophytic Bacteria in the Amaranth Root System
The Shannon index, which was used to describe the endophytic bacterial diversity, and the ACE index, which was used to describe the endophytic bacterial richness in amaranth roots, were not significantly different between the different fertilization regimens (Figure 1A,B). These results indicate that the endophytic bacterial diversity and richness in amaranth roots were not significantly affected by the application of the different fertilization treatments. In addition, to assess the similarity of the endophytic bacterial communities, we also performed principal coordinate analysis (PCoA) and partial least squares discriminant analysis (PLS−DA) at the operational taxonomic unit (OTU) level (Figure 1C,D). As shown in Figure 1C, PCoA1 and PCo2 contributed 29.98% and 17.12%, respectively, to the overall change in the endophytic bacterial structure. In addition, PLS-DA showed that the endophytic bacteria were clustered into four separate groups. The results show that the application of different fertilization treatments significantly altered the structure of the endophytic bacterial community in amaranth.
As shown in Figure 2B, at the genus level, the number of unique endophytic bacterial genera in the roots of amaranth under the NP, NK, PK, and NPK treatments was 1, 7, 6, and 7, respectively. At the OTU level, the number of unique endophytic bacterial genera in the roots of amaranth under the NP, NK, PK, and NPK treatments was 9, 28, 20, and 21, respectively (Figure 2C). The results indicate that the different fertilization treatments significantly altered the composition of the endophytic bacterial communities in the amaranth roots. Specifically, compared to the NK, PK, and NPK treatments, the number of unique endophytic bacteria enriched in amaranth roots was the lowest under the NP treatment.
At the phylum level, the compositions and proportions of the dominant endophytic bacteria (i.e., relative abundance > 1%) in the amaranth roots under the conventional fertilization without potassium (NP) treatment were Proteobacteria (69.46%), Actinobacteriota (27.85%), and other bacteria (2.30%). In contrast, the predominant taxa were Proteobacteria (73.04%), Actinobacteriota (23.49%), Myxococcota (1.14%), and others (2.32%) in the amaranth roots under the conventional fertilization without phosphorus (NK) treatment; Proteobacteria (66.07%), Actinobacteriota (29.67%), and others (3.54%) in the amaranth roots under the conventional fertilization without nitrogen (PK) treatment; and Proteobacteria (68.65%), Actinobacteriota (28.44%), and others (2.45%) in the amaranth roots under the balanced fertilization (NPK) treatment (Figure 2A).
At the genus level, 22, 23, 25, and 20 dominant endophytic bacterial genera (i.e., relative abundance > 1%) were detected in the amaranth roots under the NP, NK, PK, and NPK treatments, respectively (Figure 2D).
In comparison with those in the NP, NK, and PK treatments, the proportions of Burkholderia-Caballeronia-ParaburthoLderia and Actinospica increased, but the proportions of PhenyLobacterium, Bradyrhizobium, Sphingomonas, and Allorhizobium-Neoruizobium-Pararhixobium-Rhixobium decreased in the NPK treatment. It is worth noting that Streptomyces accounted for over 10% in the NK, PK, and NPK treatments, but only 6% in the NP treatment. In addition, Acidovorax, Lechevalieria, Mycobacterium, and unclassified_ f_ Oxalobacteraceae were the unique dominant bacterial genera in the NP, NK, and PK treatments, respectively, but they were not present in the NPK treatment. In contrast, norank_f_Micropepsaceae and Catenulispora were the unique dominant bacterial genera in the NPK treatment. Moreover, Lysobacter and unclassified_ c_Alphaproteobacteria, Ensifer, and Pseudolabry, and Nocardioides, and Phycicoccus were the unique dominant bacterial genera in the NP, NK, and PK treatments, respectively. The above results show that the application of different fertilization treatments significantly changed the community structure of endophytic bacteria in the amaranth roots.
The LEfSe analysis revealed the species with differences between groups (Figure 3A,B). At the genus level, the results showed that Longimycelium and unclassified_f_Holophagaceae were significantly enriched in the NK treatment (p < 0.05). Dokdonella, Microbacterium, and Polaromonas were significantly enriched in the PK treatment (p < 0.05). Unclassified_c_Acidimierobia, and Terrisporobacter were significantly enriched in the NPK treatment (p < 0.05). However, a significant enrichment of endophytic bacteria at the phylum or genus level was not found in the roots of the amaranth plants under the NP treatment.
At the genus level, the top 50 endophytic bacteria obtained from a network analysis showed 47 nodes and 116 edges. Among them, 95 and 21 edges were significantly positively and negatively correlated, respectively. Furthermore, Ralstonia, norank__f__67-14, Phycicoccus, unclassified__f__Oxalobacteraceae, Rhodanobacter, Acidovorax, Actinoplanes, and Bradyrhizobium were the most strongly correlated with the other endophytic bacteria (degree ≥ 8) (Figure 4).
3.3. Effects of Different Fertilization Treatments on Endophytic Fungi in Amaranth Roots
As shown in Figure 5A,B, the indices of endophytic fungal diversity (Shannon) and richness (ACE) were not significantly different among the NP, NK, PK, and NPK treatments. Taken together, these findings suggest that the endophytic fungal diversity and richness in amaranth roots were not significantly altered by the application of the different fertilization treatments. In addition, the PCoA revealed that PCoA1 and PCo2 contributed 29.98% and 17.12%, respectively, to the overall changes in the endophytic fungal structure. Meanwhile, according to the PLS−DA results, the endophytic bacteria were clearly clustered into four classes, which were distributed in the negative and positive directions of Comp2. Thus, the results show that the application of different fertilizers significantly altered the structure of the endophytic fungal community in amaranth.
Similarly, at the genus level, the total numbers of unique endophytic fungi in the roots of the amaranth plants under the NP, NK, PK, and NPK treatments were 7, 38, 16, and 18, respectively (Figure 6B). At the OTU level, the total numbers of unique endophytic fungi in the roots of the amaranth plants under the NP, NK, PK, and NPK treatments were 52, 97, 54, and 54, respectively (Figure 6C). These results suggested that different fertilization treatments could significantly alter the composition of the endophytic fungal communities in amaranth roots. Specifically, compared to the NK, PK, and NPK treatments, the number of unique endophytic fungi enriched in amaranth roots was the lowest under the NP treatment.
At the phylum level, the dominant endophytic fungi (i.e., relative abundance > 1%) in the amaranth roots were Ascomycota (91.41%), Chytridiomycota (5.27%), Mortierellomycota (1.36%), and Basidiomycota (1%) under the NP treatment. The predominant bacteria in the amaranth roots under the NK treatment were Ascomycota (90.11%), Basidiomycota (5.30%), Olpidiomycota (2.07%), and unclassified_k_Fungi (1.43%). In addition, Ascomycota (86.99%), Basidiomycota (6.08%), and Mortierellomycota (5.90%) were found in the amaranth roots under the PK treatment. In contrast, Ascomycota (87.77%), Basidiomycota (4.17%), Olpidiomycota (3.28%), unclassified_k_Fungi (2.96%), and Mortierellaomycota (1.32%) were found in the amaranth roots under the NPK treatment (Figure 6A).
At the genus level, a total of 25 dominant endophytic fungal genera were detected in the amaranth roots under the different fertilization treatments. Among them, 12, 16, 14, and 14 dominant endophytic fungal genera were found under the NP, NK, PK, and NPK treatments, respectively (Figure 6D).
Compared with those under the NP, NK, and PK treatments, the proportions of Poaceascoma, and Gibellulopsis in the amaranth roots decreased under the NPK treatment. However, Unclassified_ p_ Ascomycota was particularly enriched in the amaranth roots under the NPK treatment. Dactylonectria was an endophytic fungi unique to the NP treatment, and Gibberella, and Cladosporium were endophytic fungi unique to the NK and PK treatments. Neocosmospora and Acrocalymma were endophytic fungi unique to the NP, NK, and PK treatments. Meanwhile, the proportion of Fusarium increased in the NP treatment. All these results suggest that the application of different fertilization treatments can significantly alter the community structure of endophytic fungi in amaranth roots.
At the genus level, the results of the LEfSe analyses showed that only unclassified_ 0_ Plosporales were significantly enriched in the NPK treatment (p < 0.05). However, a significant enrichment of endophytic fungi was not found in response to different fertilization regimens consisting of diverse NP, NK, and PK treatments (Figure 7).
Similarly, the results of the network analysis also revealed 48 nodes and 101 edges (Figure 8). Among them, 83 significantly positive and 18 significantly negative correlations were found. Unclassified_f__Chaetothyriales_fam_Incertae_sedis, Acrocalymma, Purpureocillium, Sarocladium, unclassified_f__Chytridiaceae, and unclassified__o_Branch06 were the most strongly correlated with the other endophytic fungi (degree ≥ 8).
3.4. Correlation Analysis of Amaranthin with Endophytic Bacteria and Fungi
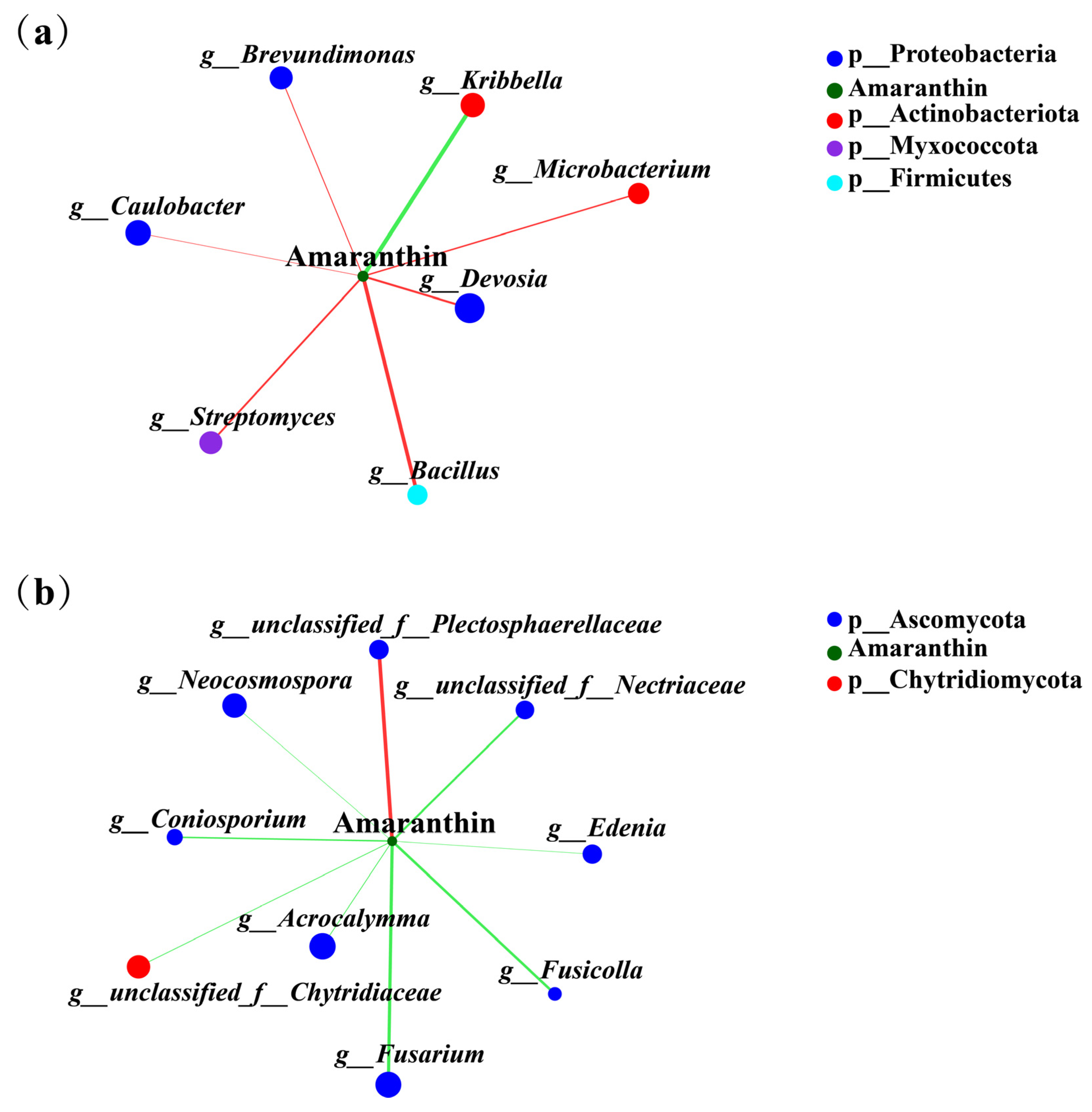
To investigate the relationship between the amaranthin and endophytic microorganisms in the roots of amaranth, Spearman’s correlation coefficient was used. The results showed that amaranthin was significantly positively correlated with Devosia, Bacillus, Streptomyces, Caulobacter, and Brevundimonas, and that it was significantly negatively correlated with Kribbella (Figure 9a). Additionally, a significant positive correlation between amaranthin and endophytic fungi, such as unclassified_f_Plectosphaerellacea, was detected. In contrast, significant negative correlations were also detected between amaranthin and endophytic fungi, such as unclassified_f_Nectriaceae, Edenia, Fusicolla, Fusarium, Acrocalymma, unclassified_f_Chytridiaceae, Coniosporium, and Neocosmospora (Figure 9b).
4. Discussion
4.1. Effects of Different Fertilization Treatments on the Amaranthin Content
Previous studies reported that the synthesis of plant pigments, such as anthocyanins, could be significantly affected by fertilization, and could be reduced under low-nitrogen conditions [37]. However, when balanced fertilizer application is applied to strawberry plants, the anthocyanin content can increase significantly [38]. Neshev et al. [39] reported that different fertilization treatments had no effect on the carotenoid content in potato leaves, but that NP fertilization significantly increased the chlorophyll content in potato leaves. Our study also revealed that the amaranthin content was affected by different fertilization regimens; among these factors, the highest amaranthin content was found in the leaves of plants under the balanced fertilization (NPK) treatment. In contrast, the lowest amaranthin content was detected in the leaves under the conventional fertilization without potassium (NP) treatment. All above results suggested that nitrogen, phosphorus and potassium were the main factors affecting amaranthin synthesis. This is consistent with previous studies [40].
4.2. Effects of Different Fertilization Treatments on the Endophytic Bacterial Community Structures in Amaranth Roots
Proteobacteria, a bioindicator of plant disease, has been reported to be the most common endophytic bacterial phylum in plant organisms [41,42,43]. Moreover, Actinobacteriota, which are producers of natural antibiotics, also widely inhabit plants [44,45]. In this experiment, Proteobacteria, and Actinobacteria were the main endophytic phyla in amaranth roots, but they exhibited significant changes under different fertilization treatments. Specifically, Proteobacteria were enriched under NK treatment, while Actinobacteria were enriched under PK treatment. All above results suggest that unbalanced fertilization may lead to amaranth producing antibiotics to protect themselves.
Streptomyces play an important role in controlling plant diseases, and many pathogens, such as ash mold, Fusarium oxysporum, anthrax of capsicum, and Aspergillus flavus, are significantly inhibited by their metabolic substances [46,47]. In addition, previous studies have shown that Streptomyces are associated with the synthesis of pigments, and gene sequencing has revealed the presence of a large number of cytochrome P450 genes in Streptomyces [48,49]. Cytochrome P450 enzymes are monooxygenases that catalyze various metabolic and biosynthetic chemical reactions [50,51]. Numerous studies have identified cytochrome P450 (CYP76AD1) as a key gene in the amaranthin metabolic pathway that plays an important role in amaranthin biosynthesis [52]. Moreover, a significant positive correlation between Streptomyces and amaranthin was found in our study. The abundance of Streptomyces was greater than 10% in the NK, PK, and NPK treatments and decreased only in the conventional fertilization without potassium (NP). This result suggested that Streptomyces was more affected by potassium than by other elements, such as nitrogen and phosphorus. It also suggested that potassium might influence the synthesis of amaranthin relating to the proportion of Streptomyces in amaranth roots.
Actinospica and Burkholderia-Caballeronia-ParaburthoLderia are beneficial bacteria for producing antibiotics and degrading pollutants [53,54]. Phenylobacterium and Acidovorax are harmful bacterial genera that can be found in the environment and cause cucumber disease [55,56]. Our results showed increased abundances of Actinospica and Burkholderia-Caballeronia-Paraburtholderia in the NPK treatment, with a decreased abundance of Phenylobacterium. Additionally, Acidovorax was enriched in amaranth roots under NP, NK, and PK treatments, but absent under NPK treatment. These results indicate that more beneficial endophytic bacteria could be enriched by balanced fertilization. In contrast, unbalanced fertilization may lead to an increase in the number of harmful endophytic bacteria in amaranth roots.
4.3. Effects of Different Fertilization Treatments on the Endophytic Fungal Community Structures in Amaranth Roots
Olpidiomycota can break down cellulose and chitin, providing more nutrients for plant growth and promoting ecosystem matter [57]. Chytidiomycota, ubiquitous fungi, are specialized parasites of many higher plant roots that act as mediators of the transmission of some soilborne viruses [58,59]. We found that in the NPK treatment, the abundance of Olpidiomycota increased, while it decreased in the unbalanced fertilization treatments (NP, NK, and PK). Furthermore, in the NPK treatment, the proportion of Chytidiomycota was the lowest, whereas it was the highest in the unbalanced fertilization treatments, especially in the NP treatment.
Acrocalymma is a type of fungus that causes crown rot [60]. However, Neocosmospora can induce black rot at the base of the plant stem and roots [61]. Many studies have shown that Fusarium species are soilborne pathogens that cause plant root rot and can secrete toxins [62,63]. In this study, we found that Fusarium, Acrocalymma, and Neocosmospora were significantly negatively correlated with amaranthin. Fusarium was the most abundant genus in the NP treatment. Neocosmospora and Acrocalymma were endophytic fungi unique to the NP, NK, and PK treatments. Previous studies have confirmed that Fusarium can produce gibberellins [64]. Moreover, the exogenous application of Gibberellin(GA3) inhibited the accumulation of amaranthin in amaranth plants [65]. These results suggest that endophytic fungi related to amaranthin formation were significantly inhibited by conventional fertilization without potassium.
Gibellulopsis is also a pathogenic fungus that induces the yellowing and wilting of sugar beet leaves [66]. Cladosporium, a mycorrhizal fungus with conidia, can easily lead to leaf mold and spot disease [67]. Dactylonectria not only produces toxins but also causes root necrosis [68,69]. Gibberella, a parasitic fungus that affects a variety of grasses, can lead to malignant seedling disease in rice, and culm rot or cob rot in maize [70]. Our study revealed that the abundance of Gibellulopsis decreased in the roots of the amaranth plants under the NPK treatment compared to those under the NP, NK, and PK treatments. In addition, Dactylonectria was the unique dominant fungal genus in the roots of the amaranth plants under the NP treatment, and Gibberella and Cladosporium were the unique dominant fungal genera in the roots of the amaranth plants under the NK and PK treatments.
All these results indicated that unbalanced fertilizations might be more suitable for the growth of pathogenic fungi in a soil environment. Consequently, it triggered more pathogenic fungi colonizing in the amaranth roots, in association with a higher incidence of diseases. Conversely, balanced fertilization might enhance the resistance of plants to adverse environmental conditions, reducing the damage from environmental stress for plants.
5. Conclusions
The above results suggest that endophytes can easily colonize amaranth roots as beneficial microorganisms under balanced fertilization conditions. However, endophytes can easily colonize amaranth roots as harmful microorganisms under unbalanced fertilization conditions. Additionally, we also observed a significant positive correlation between Streptomyces and amaranthin, as well as significant negative correlations among Acrocalymma, Neocosmospora, Fusarium, and amaranthin. All above results suggested that different fertilizations could significantly alter the endophytic microbial community structures in amaranth roots. Balanced fertilization (N, P and K fertilizers are 188.0, 53.0, and 50.0 kg·hm−2, respectively) could recruit more beneficial endogenous microorganisms in amaranth roots for improving their growth and quality.
Author Contributions
X.L.: conceptualization, methodology, software, formal analysis, data curation, writing—original draft, writing—review and editing, and visualization. D.Y.: software, data curation. X.Z.: software, data curation. X.W.: software, formal analysis. Y.X.: software, formal analysis, data curation. S.Y.: conceptualization, methodology, resources, writing—review and editing, supervision, funding acquisition. All authors have read and agreed to the published version of the manuscript.
Funding
This study was supported by the Key Research and Development Program of Guangxi (Guike AB23026076).
Informed Consent Statement
Not applicable.
Data Availability Statement
The original reads were stored in the NCBI Sequence Read Archive (SRA) database (accession number: PRJNA963072).
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Sarker, U.; Islam, M.T.; Rabbani, M.G.; Oba, S.Y. Genotypic variability for nutrient, antioxidant, yield and yield contributing traits in vegetable amaranth. J. Food Agric. Environ. 2014, 12, 168–174. Available online: https://www.researchgate.net/publication/267509472 (accessed on 10 November 2023).
- Sarker, U.; Oba, S. Nutraceuticals, antioxidant pigments, and phytochemicals in the leaves of Amaranthus spinosus and Amaranthus viridis weedy species. Sci. Rep. 2019, 9, 20413. [Google Scholar] [CrossRef] [PubMed]
- Sarker, U.; Islam, M.T.; Rabbani, M.G.; Oba, S. Variability, heritability and genetic association in vegetable amaranth (Amaranthus tricolor L.). Span. J. Agric. Res. 2015, 13, e0702. [Google Scholar] [CrossRef]
- Chakrabarty, T.; Sarker, U.; Hasan, M. Variability in mineral compositions, yield and yield contributing traits of stem amaranth (Amaranthus lividus). Genetika 2018, 50, 995–1010. [Google Scholar] [CrossRef]
- Stintzing, F.C.; Carle, R. Functional properties of anthocyanins and betalains in plants, food, and in human nutrition. Trends Food Sci. Technol. 2014, 15, 19–38. [Google Scholar] [CrossRef]
- Anton, G.; Živa, P.; Klemen, E.; Mihelič, R.; Suhadolc, M. Combined effects of long-term tillage and fertilization regimes on soil organic carbon, microbial biomass, and abundance of the total microbial communities and N-functional guilds. Appl. Soil. Ecol. 2023, 188, 104876. [Google Scholar] [CrossRef]
- Sebastiana, M.; Rafael JL, B.; Bellido, L.L.; Romero, V.M.; Moreno, F.; Murillo, J.M. Long-term effect of tillage, rotation and nitrogen fertilizer on soil quality in a Mediterranean Vertisol. Soil. Tillage Res. 2011, 114, 97–107. [Google Scholar] [CrossRef]
- Wang, L.; Wang, J.; Tang, Z.H.; Wang, J.D.; Zhang, Y.C. Long-term organic fertilization reshapes the communities of bacteria and fungi and enhances the activities of C- and P-cycling enzymes in calcareous alluvial soil. Appl. Soil. Ecol. 2024, 194, 105204. [Google Scholar] [CrossRef]
- Zhang, M.K.; Zhang, H.M. Co-transport of dissolved organic matter and heavy metals in soils induced by excessive phosphorus applications. J. Environ. Sci. 2010, 22, 598–606. [Google Scholar] [CrossRef]
- Tsavkelova, E.A.; Cherdyntseva, T.A.; Lobakova, E.S.; Kolomeĭtseva, G.L.; Netrusov, A.I. Microbiota of the orchidrhizoplane. Mikrobiologiia 2001, 70, 567–573. Available online: https://pubmed.ncbi.nlm.nih.gov/11558285/ (accessed on 10 November 2023).
- Lacava, P.T.; Bogas, A.C.; Cruz, F.P.N. Plant Growth Promotion and Biocontrol by Endophytic and Rhizospheric Microorganisms from the Tropics: A Review and Perspectives. Front. Sustain. Food Syst. 2022, 6, 796113. [Google Scholar] [CrossRef]
- Singh, C.B. Role of endophytic microorganisms in sustainable agriculture. NeBIO 2012, 3, 69–77. [Google Scholar] [CrossRef] [PubMed]
- Shankar, S.; Segaran, G.; Sathiavelu, M. Chapter 4—Antimicrobial metabolites from endophytic microorganisms and its mode of action. Biocontrol Mech. Endophytic Microorg. 2022, 75–88, 9780323884785. [Google Scholar] [CrossRef]
- Pacifico, D.; Squartini, A.; Crucitti, D.; Barizza, E.; Lo Schiavo, F.; Muresu, R.; Carimi, F.; Zottini, M. The role of the endophytic microbiome in the grapevine response to environmental triggers. Front. Plant Sci. 2019, 10, 1256. [Google Scholar] [CrossRef] [PubMed]
- Schlaeppi, K.; Dombrowski, N.; Oter, R.G.; Themaat, E.V.L.V.; Lefert, P.S. Quantitative divergence of the bacterial root microbiota in Arabidopsis thaliana relatives. Proc. Natl. Acad. Ences 2013, 111, 585–592. [Google Scholar] [CrossRef] [PubMed]
- Bulgarelli, D.; Garrido-Oter, R.; Münch, P.C.; Weiman, A.; Dröge, J.; Pan, Y.; McHardy, A.C.; Schulze-Lefert, P. Structure and Function of the Bacterial Root Microbiota in Wild and Domesticated Barley. Cell Host Microbe 2015, 17, 392–403. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.-J.; Guo, G.; Li, M.; Liang, X.-Y.; Gu, Y.-Y. Diversity of endophytic bacteria of mulberry (Morus L.) under cold conditions. Front. Microbiol. 2022, 13, 923162. [Google Scholar] [CrossRef] [PubMed]
- Xiao, J.; Chen, S.; Liang, T.; Yang, S.; Tan, H. Response of endophytic bacteria in sugarcane roots to different slow-release fertilizers with dicyandiamide (DCD) and humic acid (HA) applications. Environ. Technol. Innov. 2023, 32, 103244. [Google Scholar] [CrossRef]
- Toledo, S.; Gargaglione, V.; Peri, P.L. Mineral fertilization impacts microbial activity and endophytic fungi but not microbial biomass in semiarid grasslands. Pedobiologia 2024, 102, 150929. [Google Scholar] [CrossRef]
- Sindhu, L.; Song, Y.; Li, L.; Dong, Z.; Yang, Q.; Mangi, S.S.; Guo, W.; Yang, L.; Cui, H.; Lv, S.; et al. Meta-omics revealed that nitrogen fertilization alters the endophytic fungi divergence in maize field ecosystem. Ecol. Indic. 2024, 160, 111852. [Google Scholar] [CrossRef]
- Ma, Y.; Weisenhorn, P.; Guo, X.; Wang, D.; Yang, T.; Shi, Y.; Zhang, H.; Chu, H. Effect of long-term fertilization on bacterial communities in wheat endosphere. Pedosphere 2021, 31, 538–548. [Google Scholar] [CrossRef]
- Sun, A.; Jiao, X.-Y.; Chen, Q.; Wu, A.-L.; Zheng, Y.; Lin, Y.-X.; He, J.-Z.; Hu, H.-W. Microbial communities in crop phyllosphere and root endosphere are more resistant than soil microbiota to fertilization. Soil. Biol. Biochem. 2021, 153, 108113. [Google Scholar] [CrossRef]
- Pedraza, R.O.; Bellone, C.H.; de Bellone, S.C.; Boa Sorte, P.M.F.; dos Santos Teixeira, K.R. Azospirillum inoculation and nitrogen fertilization effect on grain yield and on the diversity of endophytic bacteria in the phyllosphere of rice rainfed crop. Eur. J. Soil. Biol. 2009, 45, 36–43. [Google Scholar] [CrossRef]
- Liu, Y.; Chu, G.; Stirling, E.; Zhang, H.; Chen, S.; Xu, C.; Zhang, X.; Ge, T.; Wang, D. Nitrogen fertilization modulates rice seed endophytic microbiomes and grain quality. Sci. Total Environ. 2023, 857 Pt 2, 159181. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Wu, Y.; Cao, C.; Han, S.; Zhao, W.; Li, Q.; Liu, X.; Kong, L. Effects of fertilizer reduction coupled with straw returning on soil fertility, wheat root endophytic bacteria, and the occurrence of wheat crown rot. Front. Microbiol. 2023, 14, 1143480. [Google Scholar] [CrossRef] [PubMed]
- Cooper-Driver, G.A. Contributions of Jeffrey Harborne and coworkers to the study of anthocyanins. Phytochemistry 2001, 56, 229–236. [Google Scholar] [CrossRef] [PubMed]
- Hazli, U.H.A.M.; Abdul-Aziz, A.; Mat-Junit, S.; Chee, C.F.; Kong, K.W. Solid-liquid extraction of bioactive compounds with antioxidant potential from Alternanthera sesillis (red) and identification of the polyphenols using UHPLC-QqQ-MS/MS. Food Res. Int. 2019, 115, 241–250. [Google Scholar] [CrossRef] [PubMed]
- Hasanuzzaman, M.; Bhuyan, M.H.M.B.; Zulfiqar, F.; Raza, A.; Mohsin, S.M.; Mahmud, J.A.; Fujita, M.; Fotopoulos, V. Reactive Oxygen Species and Antioxidant Defense in Plants under Abiotic Stress: Revisiting the Crucial Role of a Universal Defense Regulator. Antioxidants 2020, 9, 681. [Google Scholar] [CrossRef] [PubMed]
- Das, K.; Roychoudhury, A. Reactive oxygen species (ROS) and response of antioxidants as ROS-scavengers during environmental stress in plants. Front. Environ. Sci. 2014, 2, 53. [Google Scholar] [CrossRef]
- Yang, D.; Lin, X.; Zhou, X.; Li, Z.; Kurokawa, H.; Matsui, H.; Fujita, T.; Yang, S.-D. Differences in endophytic bacterial and fungal compositions in roots between red and green Amaranthus sp. S. Afr. J. Bot. 2023, 163, 275–284. [Google Scholar] [CrossRef]
- Chen, S.; Sun, Y.; Wei, Y.; Li, H.; Yang, S. Different rhizosphere soil microbes are recruited by tomatoes with different fruit color phenotypes. BMC Microbiol. 2022, 22, 210. [Google Scholar] [CrossRef] [PubMed]
- Timoneda, A.; Feng, T.; Sheehan, H.; Walker-Hale, N.; Pucker, B.; Lopez-Nieves, S.; Guo, R.; Brockington, S. The evolution of betalain biosynthesis in Caryophyllales. New Phytol. 2019, 224, 71–85. [Google Scholar] [CrossRef] [PubMed]
- Li, W.Y.; Wang, Z.Q.; Gong, W.H.; Yang, L.F. Optimum dosage test of nitrogen, phosphorus and potassium fertilizers in amaranth. Vegetables 2015, 2, 29–31. Available online: https://kns.cnki.net/kcms2/article/abstract?v=8WLnD7pOpNFi_PphG5xeLIX0HoKITDlbLr0AomwERBpbRyxl0XUeL5alpeP0ZXjKgk3w7DhWs_cRxEu_RQp1kIiNdqqRBO_CcSfnZHG4FpMtQ_4y0MUgBgZYKbOXggCQ5UCCNiUvae4FL7VkYSe9sA==&uniplatform=NZKPT&language=CHS (accessed on 10 November 2023).
- Xiao, J.; Sun, Y.; He, Y.; Tang, X.F.; Yang, S.D.; Huang, J.Y. Comparison of Rhizospheric and Endophytic Bacterial Compositions between Netted and Oriental Melons. Microbiol. Spectr. 2023, 11, e0402722. [Google Scholar] [CrossRef] [PubMed]
- Piattelli, M.; Nicola, M.; Castrogiovanni, V. The effect of kinetin on amaranthin synthesis in Amaranthus tricolor in the dark. Phytochemistry 1971, 10, 289–293. [Google Scholar] [CrossRef]
- Zhou, H.; Jia, S.; Gao, Y.; Li, X.; Lin, Y.; Yang, F.; Ni, K. Characterization of phyllosphere endophytic lactic acid bacteria reveals a potential novel route to enhance silage fermentation quality. Commun. Biol. 2024, 7, 117. [Google Scholar] [CrossRef]
- Meng, J.X.; Gao, Y.; Han, M.L. In vitro Anthocyanin Induction and Metabolite Analysis in Malus spectabilis Leaves Under Low Nitrogen Conditions. Hortic. Plant J. 2020, 6, 284–292. [Google Scholar] [CrossRef]
- Hassan, A. Effect of Nitrogen Fertilizer Levels in the Form of Organic, Inorganic and Bio fertilizer Applications on Growth, Yield and Quality of Strawberry. Middle E. J. Appl. Sci. 2015, 5, 604–617. [Google Scholar]
- Neshev, N.; Manolov, I. Influence of mineral fertilization on the leaf plastid pigment content and tuber quality of potatoes. Agrar. Nauk. 2015, 7, 35–40. [Google Scholar]
- Stobart, A.K.; Hendry GA, F.; Ei-Hussein, S.; Kinsman, L.T. The Effect of Potassium on Amaranthin Synthesis in Seedlings of Amaranthus caudatus L. Z. Für Pflanzenphysiol. 1980, 96, 217–225. [Google Scholar] [CrossRef]
- Romero, E.M.; Marina, M.; Pieckenstain, F.L. The communities of tomato (Solanum lycopersicum L.) leaf endophytic bacteria, analysed by 16S-ribosomal RNA gene pyrosequencing. Fems Microbiol. Lett. 2014, 351, 187–194. [Google Scholar] [CrossRef]
- Natacha, B.; Horton, M.W.; Joy, B. Bacterial Communities Associated with the Leaves and the Roots of Arabidopsis thaliana. PLoS ONE 2013, 8, e56329. [Google Scholar] [CrossRef]
- Bakker, M.G.; Manter, D.K.; Sheflin, A.M.; Weir, T.L.; Vivanco, J.M. Harnessing the rhizosphere microbiome through plant breeding and agricultural management. Plant Soil. 2012, 360, 1–13. [Google Scholar] [CrossRef]
- Qin, S.; Xing, K.; Jiang, J.H.; Xu, L.H.; Li, W.J. Biodiversity, bioactive natural products and biotechnological potential of plant-associated endophytic actinobacteria. Appl. Microbiol. Biotechnol. 2011, 89, 457–473. [Google Scholar] [CrossRef]
- Bérdy, J. Erratum: Thoughts and facts about antibiotics: Where we are now and where we are heading. J. Antibiot. 2012, 65, 441. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.L.; Ning, P.; Zheng, L.; Huang, J.B.; Li, G.Q.; Hsiang, T. Fumigant activity of volatiles of Strep-T. globisporus JK-1 against Penicillium italicum on Citrus microcarpa. Postharvest Biol. Technol. 2010, 58, 157–165. [Google Scholar] [CrossRef]
- Polturak, A.A. “La Vie en Rose”: Biosynthesis, sources, and applications of betalain pigments. Mol. Plant 2018, 11, 7–22. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Zheng, X.; Pan, J.; Peng, L.; Cheng, C.; Wang, X.; Zhao, C.; Zhang, Z.; Lin, Y.; Xu, X.; et al. RNA-sequencing analysis reveals betalain metabolism in the leaf of Amaranthus tricolor L. PLoS ONE 2019, 14, e0216001. [Google Scholar] [CrossRef]
- Kim, D. Streptomyces cytochrome P450 enzymes and their roles in the biosynthesis of macrolide therapeutic agents. Biomol. Ther. 2019, 27, 127–133. [Google Scholar] [CrossRef]
- Nzuza, N.; Padayachee, T.; Syed, P.R.; Kryś, J.D.; Chen, W.; Gront, D.; Nelson, D.R.; Syed, K. Ancient bacterial class Alphaproteobacteria cytochrome P450 monooxygenases can be found in other bacterial species. Int. J. Mol. Sci. 2021, 22, 5542. [Google Scholar] [CrossRef]
- Wu, Y.C.; Yuan, J.; Yao, Y.E. Effects of volatile organic compounds from Streptomyces albulus NJZJSA2 on growth of two fungal pathogens. J. Basic. Microbiol. 2015, 55, 1104–1117. [Google Scholar] [CrossRef]
- Boukaew, S.; Cheirsilp, B.; Prasertsan, P.; Yossan, S. Antifungal effect of volatile organic compounds produced by Streptomyces salmonis PSRDC-09 against the anthracnose pathogen Colle totric hum gloeos porioides PSU-03 in postharvest chili fruit. J. Appl. Microbiol. 2021, 131, 1452–1463. [Google Scholar] [CrossRef]
- Sohn, J.H.; Kwon, K.K.; Kang, J.H.; Jung, H.B.; Kim, S.J. Novosphingobium pentaromativorans sp. nov., a high-molecular-mass polycyclic aromatic hydrocarbon-degrading bacterium isolated from estuarine sediment. Int. J. Syst. Evol. Microbiol. 2004, 54 Pt 5, 1483–1487. [Google Scholar] [CrossRef] [PubMed]
- Lin, Z.; Pang, S.; Zhou, Z.; Wu, X.; Li, J.; Huang, Y.; Zhang, W.; Lei, Q.; Bhatt, P.; Mishra, S.; et al. Novel pathway of acephate degradation by the microbial consortium ZQ01 and its potential for environmental bioremediation. J. Hazard. Mater. 2022, 426, 127841. [Google Scholar] [CrossRef] [PubMed]
- Zuo, J.; Hu, L.; Shen, W. The involvement of α-proteobacteria Phenylobacterium in maintaining the dominance of toxic Microcystis blooms in Lake Taihu, China. Environ. Microbiol. 2020, 23, 1066–1078. [Google Scholar] [CrossRef]
- Bi, X.Y.; Li, X.D.; Yu, H.B. Development of a multiplex RT‒PCR assay for simultaneous detection of Cucumber green mottle mosaic virus and Acidovorax citrulli in watermelon. PeerJ 2019, 22, e7539. [Google Scholar] [CrossRef] [PubMed]
- Lay, C.Y.; Hamel, C.; St-Arnaud, M. Taxonomy and pathogenicity of Olpidium brassicae and its allied species. Fungal Biol. 2018, 122, 837–846. [Google Scholar] [CrossRef] [PubMed]
- James, T.Y.; Letcher, P.M.; Longcore, J.E.; Mozley-Standridge, S.E.; Porter, D.; Powell, M.J. A molecular phylogeny of the flagellated fungus (Chytridiomycota) and description of a new phylum (Blastocladiomycota). Mycologia 2006, 98, 860–871. [Google Scholar] [CrossRef]
- Ngoh, D.J.; Patrice, D.B.; Djongnang, G. Brown Spot and Stalk Rot Diseases of Maize (Zea Mays) and Susceptibility of Two Varieties to Physoderma maydis in Far North Cameroon. Int. J. Phytopathol. 2021, 10, 19–28. [Google Scholar] [CrossRef]
- Nikandrow, A. Acrocalymma medicaginis and Phomopsis sp. as Causal Agents of Crown Rot of Lucerne in Australia. J. Phytopathol. 2010, 130, 24–36. [Google Scholar] [CrossRef]
- Pan, R.; Deng, Q.; Deng, M.; Guan, M.; Xu, D.; Gai, Y.; Chen, W.; Yang, Y. This paper provides the first report of peanut foot rot caused by Neocosmospora vasinfecta in mainland China. Plant Pathol. 2010, 59, 1172. [Google Scholar] [CrossRef]
- Luo, H.; Hallen-Adams, H.E.; Walton, J.D. Processing of the phalloidin proprotein by prolyloligopeptidase from the mushroom conocybe albipes. J. Biol. Chem. 2009, 284, 18070–18077. [Google Scholar] [CrossRef] [PubMed]
- Nancy, J.A.; Robert, H.P.; Susan, P.M. Genes, gene clusters, and biosynthesis of trichothecenes and fumonisins in Fusarium. Toxin Rev. 2009, 28, 198–215. [Google Scholar] [CrossRef]
- Hassan, H.A.H. Gibberellin and auxin production plant root fungi and their biosynthesis under salinity-calcium interaction. Rostl. Vyrob. 2002, 48, 101–106. [Google Scholar] [CrossRef]
- Kinsman, L.T.; Pinfield, N.J.; Stobart, A.K. The hormonal control of amaranthin synthesis in Amaranthus caudatus seedlings. Planta 1975, 127, 207–212. [Google Scholar] [CrossRef]
- Zhou, Y.; Zhao, Z.; Guo, Q. This is the first report of wilt of sugar beet caused by G. nigrescens in the Xinjiang Region of China. Plant Dis. 2017, 101, 1318. [Google Scholar] [CrossRef]
- Avasthi, S.; Gautam, A.K.; Bhadauria, R. This article provides the first report of Cladosporium sphaerospermum causing leaf spot disease in Aloe vera in India. Crop Prot. 2016, 5, 649–654. [Google Scholar] [CrossRef]
- Manici, L.M.; Caboni, E.; Caputo, F.; Frattarelli, A.; Lucioli, S. Phytotoxins from Dactylonectria torresensis involved in replant disease of fruit trees. Rhizosphere 2021, 17, 100300. [Google Scholar] [CrossRef]
- Erper, I.; Ozer, G.; Alkan, M.; Zholdoshbekova, S.; Turkkan, M. First report of Dactylonectria torresensis causing black root rot in strawberry plants in Kyrgyzstan. J. Plant Pathol. 2021, 103, 379–380. [Google Scholar] [CrossRef]
- Hsuan, H.M.; Salleh, B.; Zakaria, L. Molecular Identification of Fusarium Species in Gibberella fujikuroi Species Complex from Rice, Sugarcane and Maize from Peninsular Malaysia. Int. J. Mol. Sci. 2011, 12, 6722–6732. [Google Scholar] [CrossRef]
Figure 1.
The Shannon (A) and ACE (B) indices of endophytic bacteria in amaranth roots under different fertilization treatments; PCoA (C) and PLS−DA (D) of endophytic bacteria in amaranth roots under different fertilization treatments at the OTU level.Same letters on bars within a figure indicate no significant differences in mean ranks among treatments at p > 0.05.
Figure 1.
The Shannon (A) and ACE (B) indices of endophytic bacteria in amaranth roots under different fertilization treatments; PCoA (C) and PLS−DA (D) of endophytic bacteria in amaranth roots under different fertilization treatments at the OTU level.Same letters on bars within a figure indicate no significant differences in mean ranks among treatments at p > 0.05.
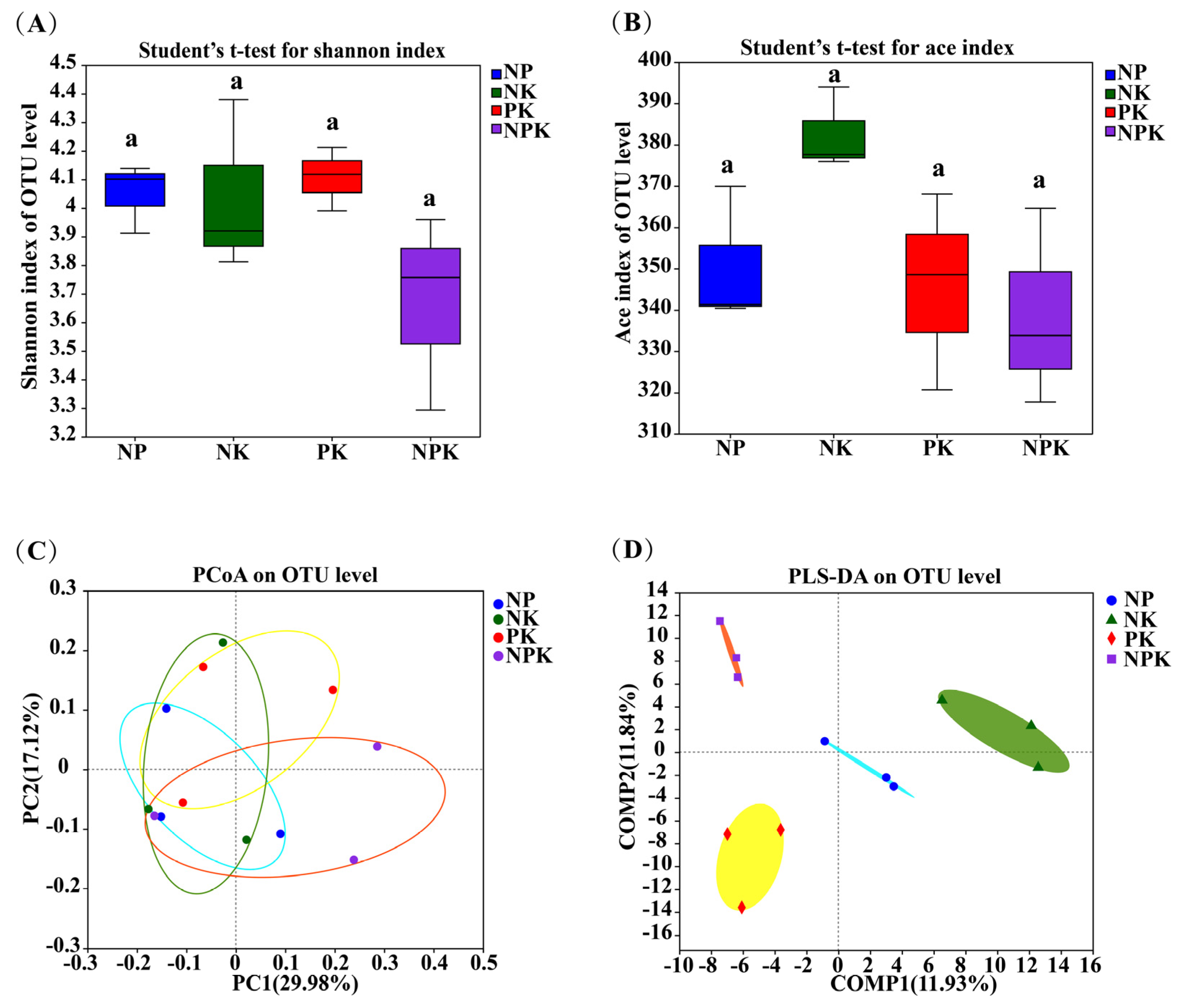
Figure 2.
(A) Compositions and proportions of endophytic bacteria in amaranth roots under different fertilization treatments at the phylum level. (B) Venn diagram of endophytic bacteria in amaranth roots at the genus level. (C) Venn diagram of endophytic bacteria in amaranth roots at the OTU level (Blue: NP; Green: NK; Red: PK; Purple: NPK). The numbers in the Venn diagram represent the number of endophytic bacteria in amaranth roots in different fertilisation treatments. (D) Compositions and proportions of endophytic bacteria in amaranth roots under different fertilization treatments at the genus level.
Figure 2.
(A) Compositions and proportions of endophytic bacteria in amaranth roots under different fertilization treatments at the phylum level. (B) Venn diagram of endophytic bacteria in amaranth roots at the genus level. (C) Venn diagram of endophytic bacteria in amaranth roots at the OTU level (Blue: NP; Green: NK; Red: PK; Purple: NPK). The numbers in the Venn diagram represent the number of endophytic bacteria in amaranth roots in different fertilisation treatments. (D) Compositions and proportions of endophytic bacteria in amaranth roots under different fertilization treatments at the genus level.
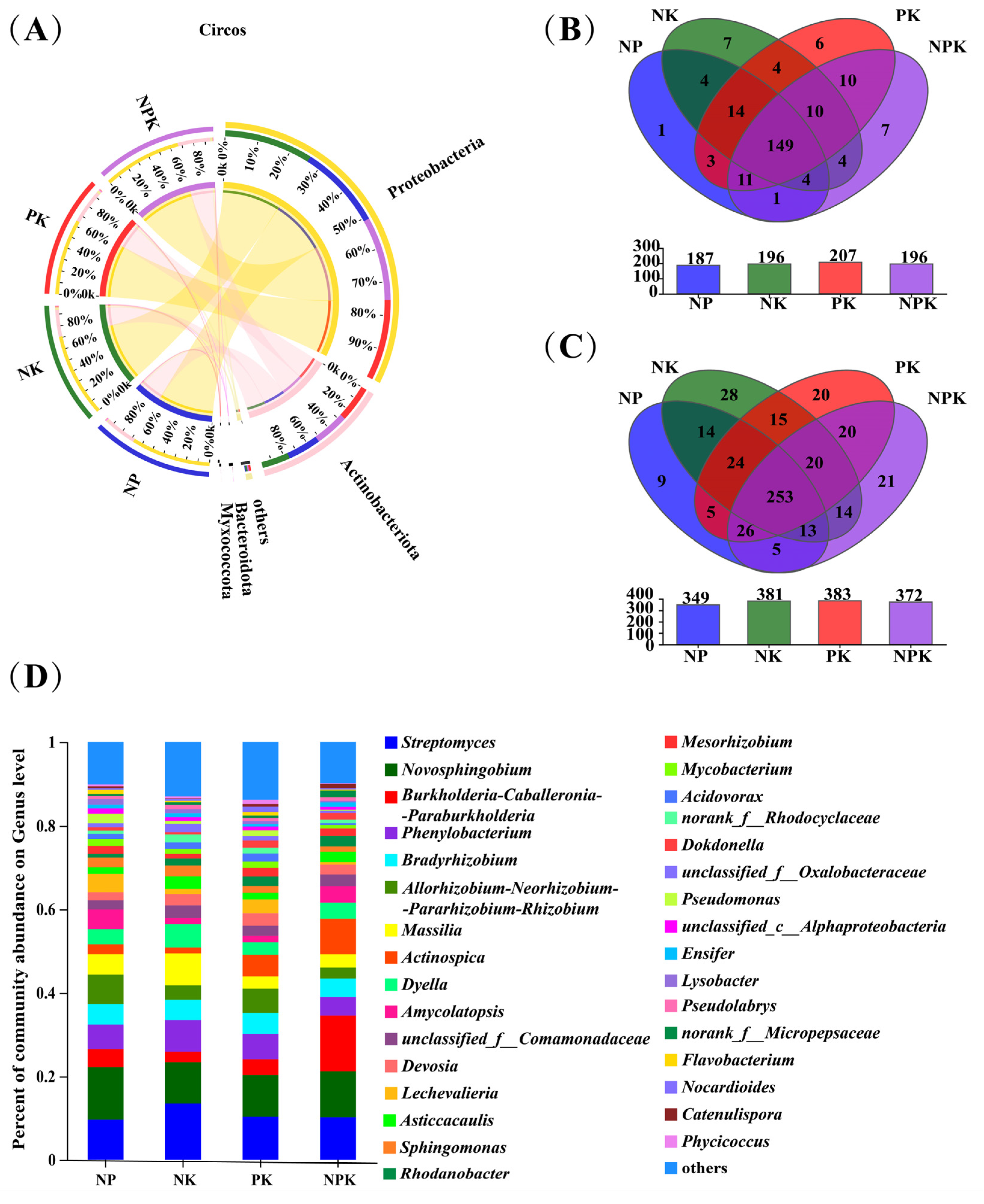
Figure 3.
Cladogram (A) and LEfSe bar (B) of the root endophytic bacterial communities under the different fertilization treatments (Blue: NP; Green: NK; Red: PK; Purple: NPK). Pathologically, nodes indicate microbial taxa that are significantly enriched in the corresponding group and have a significant effect on the differences between groups (p, phylum; C, class; 0, order; f, family; and g, genus). (p < 0.05, LDA scores ≥ 2).
Figure 3.
Cladogram (A) and LEfSe bar (B) of the root endophytic bacterial communities under the different fertilization treatments (Blue: NP; Green: NK; Red: PK; Purple: NPK). Pathologically, nodes indicate microbial taxa that are significantly enriched in the corresponding group and have a significant effect on the differences between groups (p, phylum; C, class; 0, order; f, family; and g, genus). (p < 0.05, LDA scores ≥ 2).
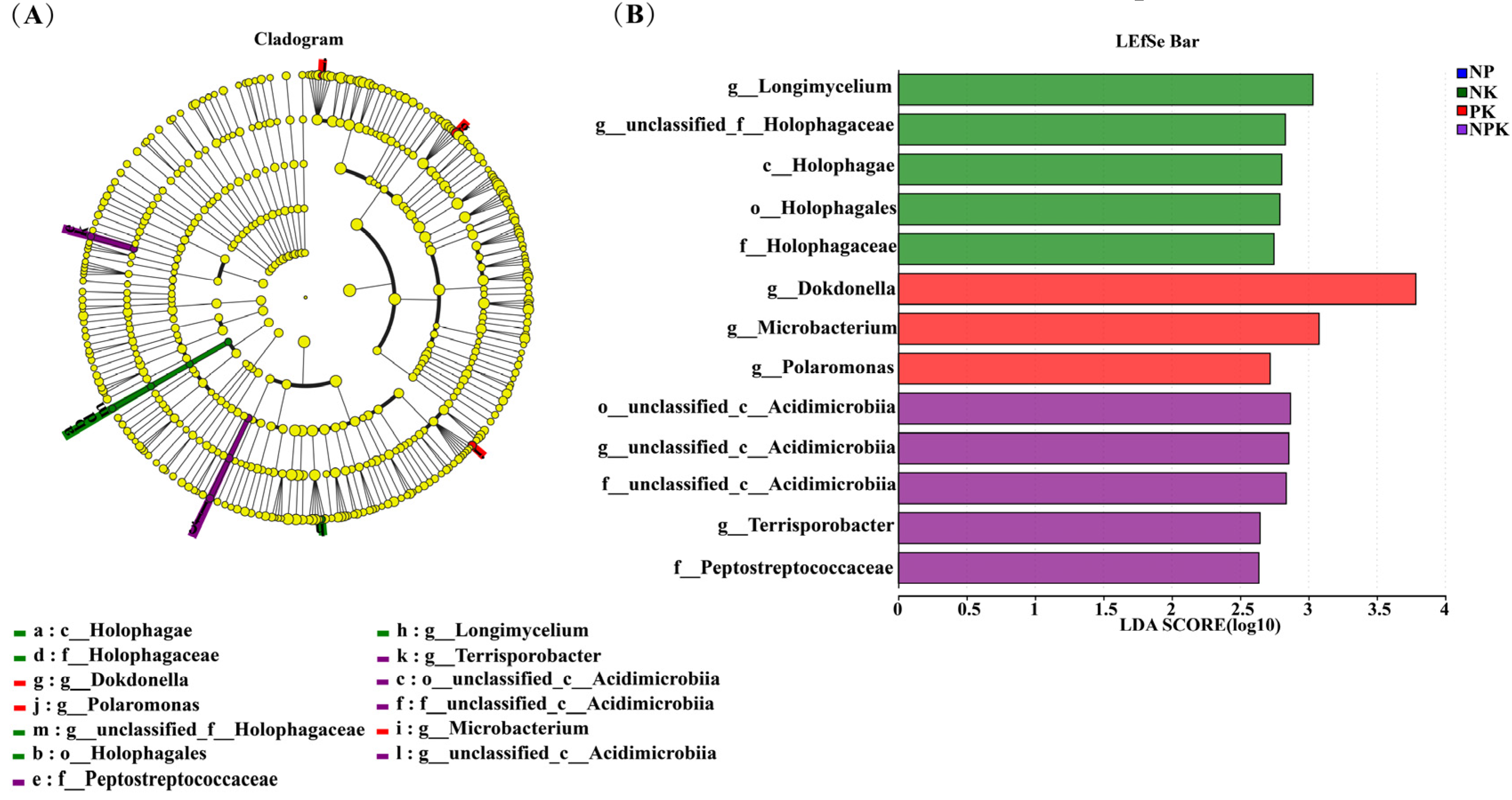
Figure 4.
Co-occurrence network analysis of endophytic bacterial communities under different fertilization treatments. Different levels are indicated by different prefixes (p, phylum; g, genus). The size of the nodes in the graph indicates the size of the species abundance, and different colors indicate different species; the colors of the connecting lines indicate positive and negative correlations, with red indicating positive correlation and green indicating negative correlation (p < 0.05); the thickness of the lines indicates the size of the correlation coefficient; the coarser the line is, the greater the correlation between the species; and the greater the number of lines is, the closer the connection between the species and other species.
Figure 4.
Co-occurrence network analysis of endophytic bacterial communities under different fertilization treatments. Different levels are indicated by different prefixes (p, phylum; g, genus). The size of the nodes in the graph indicates the size of the species abundance, and different colors indicate different species; the colors of the connecting lines indicate positive and negative correlations, with red indicating positive correlation and green indicating negative correlation (p < 0.05); the thickness of the lines indicates the size of the correlation coefficient; the coarser the line is, the greater the correlation between the species; and the greater the number of lines is, the closer the connection between the species and other species.

Figure 5.
The Shannon (A) and ACE (B) indices of endophytic fungi in amaranth roots under different fertilization treatments. PCoA (C) and PLS−DA (D) of endophytic fungi in amaranth roots under different fertilization treatments at the OTU level.Same letters on bars within a figure indicate no significant differences in mean ranks among treatments at p > 0.05.
Figure 5.
The Shannon (A) and ACE (B) indices of endophytic fungi in amaranth roots under different fertilization treatments. PCoA (C) and PLS−DA (D) of endophytic fungi in amaranth roots under different fertilization treatments at the OTU level.Same letters on bars within a figure indicate no significant differences in mean ranks among treatments at p > 0.05.
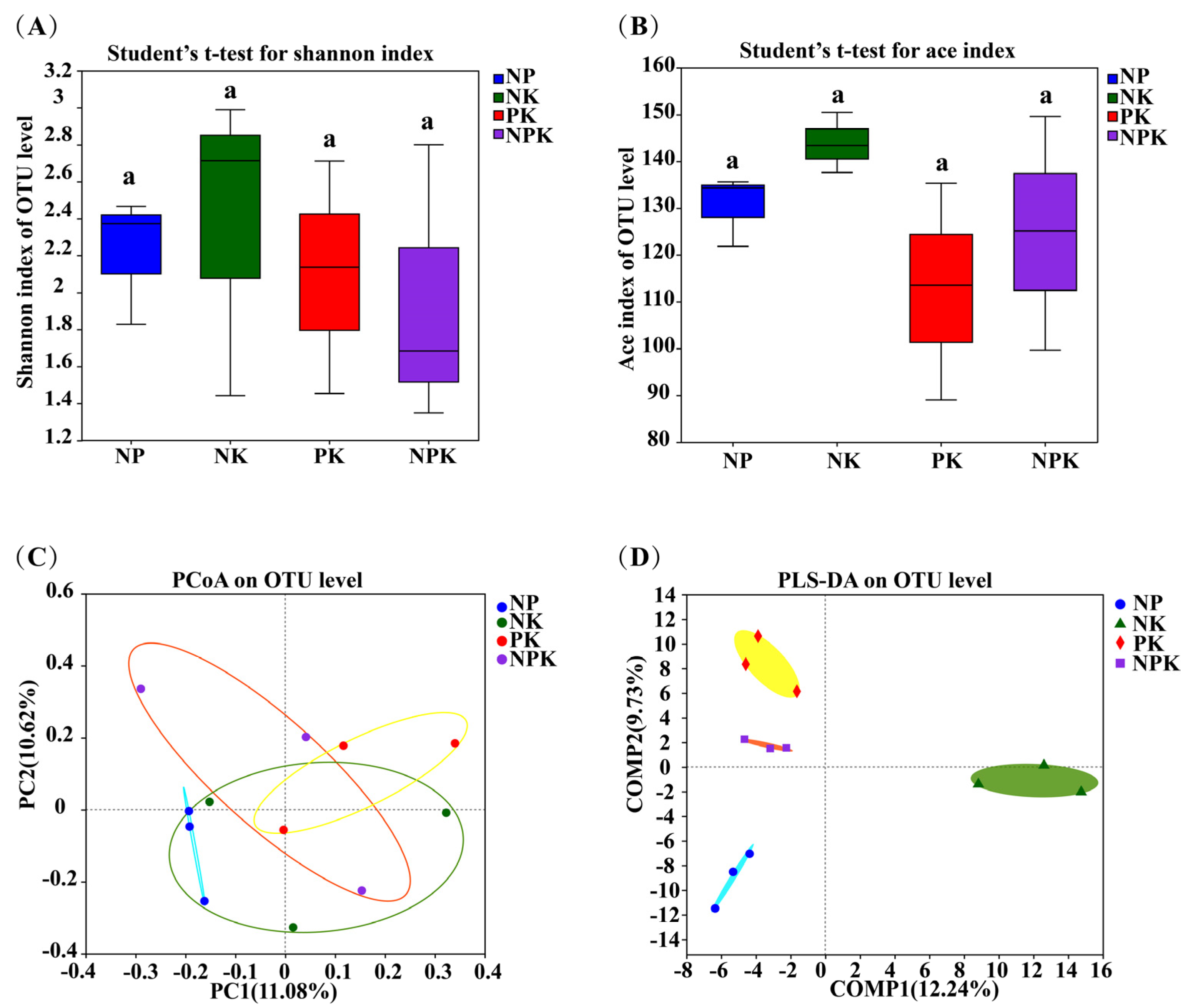
Figure 6.
(A) Compositions and proportions of endophytic fungi in amaranth roots under different fertilization treatments at the phylum level. (B) Venn diagram of endophytic fungi in amaranth roots at the genus level. (C) Venn analyses of endophytic fungi in amaranth roots at the OTU level (Blue: NP; Green: NK; Red: PK; Purple: NPK). The numbers in the Venn diagram represent the number of endophytic fungi in amaranth roots in different fertilisation treatments. (D) Compositions and proportions of endophytic fungi in amaranth roots under different fertilization treatments at the genus level.
Figure 6.
(A) Compositions and proportions of endophytic fungi in amaranth roots under different fertilization treatments at the phylum level. (B) Venn diagram of endophytic fungi in amaranth roots at the genus level. (C) Venn analyses of endophytic fungi in amaranth roots at the OTU level (Blue: NP; Green: NK; Red: PK; Purple: NPK). The numbers in the Venn diagram represent the number of endophytic fungi in amaranth roots in different fertilisation treatments. (D) Compositions and proportions of endophytic fungi in amaranth roots under different fertilization treatments at the genus level.

Figure 7.
Cladogram (A) and LEfSe bar (B) of the root endophytic fungal communities under the different fertilization treatments (Blue: NP; Green: NK; Red: PK; Purple: NPK). Pathologically, nodes indicate microbial taxa that are significantly enriched in the corresponding group and have a significant effect on the differences between groups (p, phylum; C, class; 0, order; f, family; and g, genus). (p < 0.05, LDA score ≥ 2).
Figure 7.
Cladogram (A) and LEfSe bar (B) of the root endophytic fungal communities under the different fertilization treatments (Blue: NP; Green: NK; Red: PK; Purple: NPK). Pathologically, nodes indicate microbial taxa that are significantly enriched in the corresponding group and have a significant effect on the differences between groups (p, phylum; C, class; 0, order; f, family; and g, genus). (p < 0.05, LDA score ≥ 2).
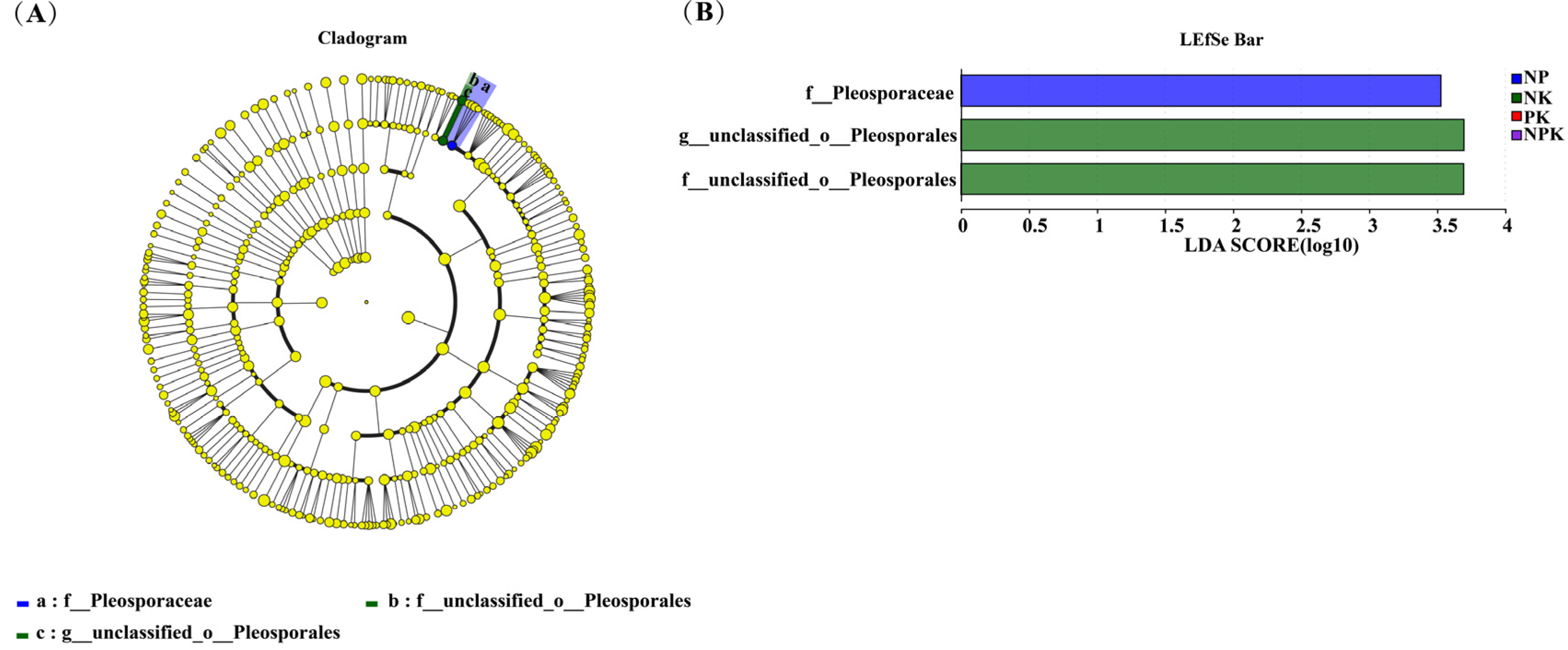
Figure 8.
Co-occurrence network analysis of endophytic fungal communities under different fertilization treatments. Different levels are indicated by different prefixes (p, phylum; g, genus). The size of the nodes in the graph indicates the size of the species abundance, and different colors indicate different species; the colors of the connecting lines indicate positive and negative correlations, with red indicating positive correlation and green indicating negative correlation (p < 0.05); the thickness of the lines indicates the size of the correlation coefficient; the coarser the line is, the greater the correlation between the species; and the greater the number of lines is, the closer the connection between the species and other species.
Figure 8.
Co-occurrence network analysis of endophytic fungal communities under different fertilization treatments. Different levels are indicated by different prefixes (p, phylum; g, genus). The size of the nodes in the graph indicates the size of the species abundance, and different colors indicate different species; the colors of the connecting lines indicate positive and negative correlations, with red indicating positive correlation and green indicating negative correlation (p < 0.05); the thickness of the lines indicates the size of the correlation coefficient; the coarser the line is, the greater the correlation between the species; and the greater the number of lines is, the closer the connection between the species and other species.
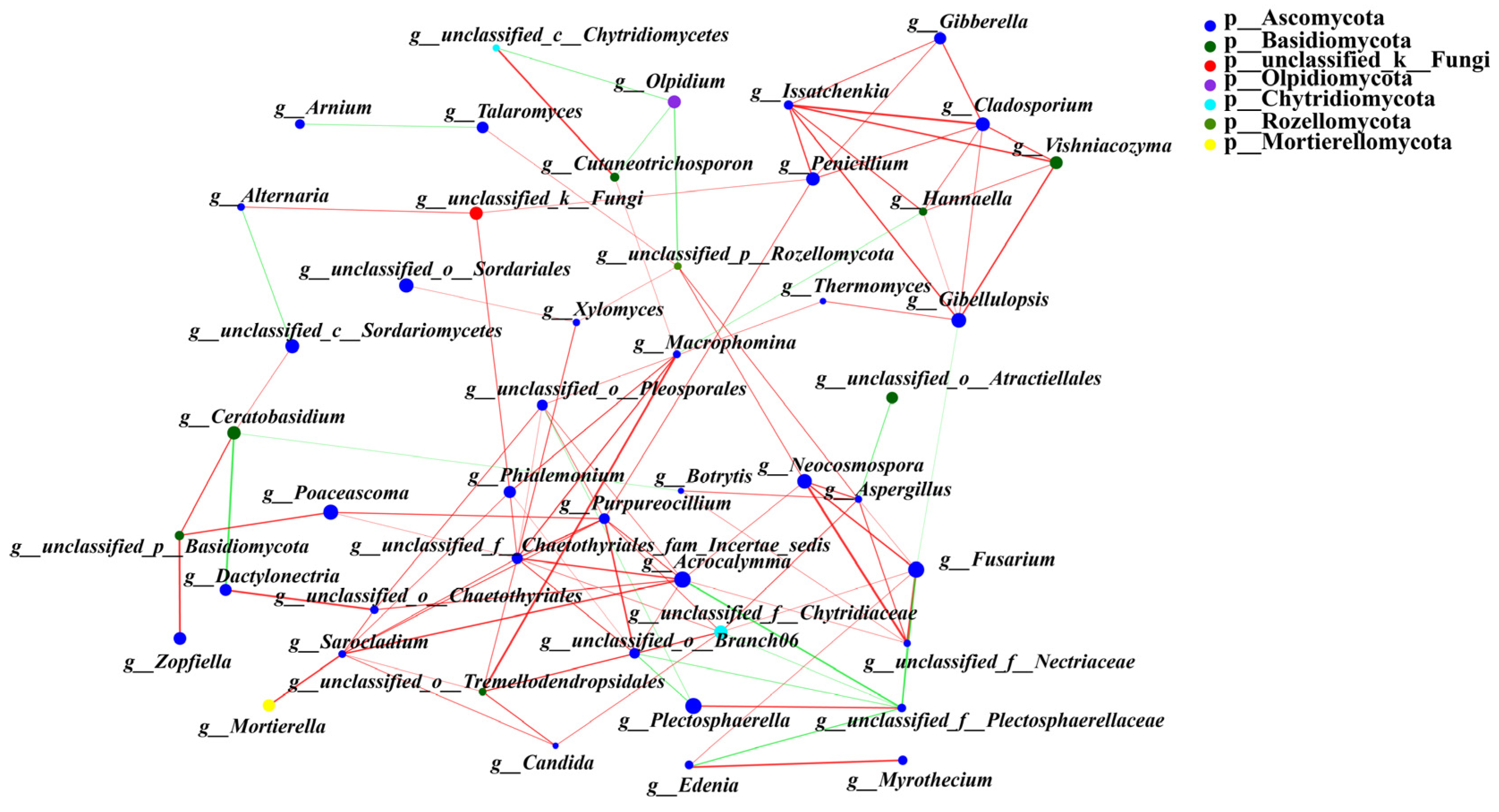
Figure 9.
Correlation analysis of amaranthin with endophytic bacteria (a) and fungi (b). Different levels are indicated by different prefixes (p, phylum; g, genus). The size of the nodes in the graph indicates the size of the species abundance, and different colors indicate different species; the colors of the connecting lines indicate positive and negative correlations, with red indicating positive correlation and green indicating negative correlation (p < 0.05); the thickness of the lines indicates the size of the correlation coefficient; the coarser the line is, the greater the correlation between the species; and the greater the number of lines is, the closer the connection between the species and other species.
Figure 9.
Correlation analysis of amaranthin with endophytic bacteria (a) and fungi (b). Different levels are indicated by different prefixes (p, phylum; g, genus). The size of the nodes in the graph indicates the size of the species abundance, and different colors indicate different species; the colors of the connecting lines indicate positive and negative correlations, with red indicating positive correlation and green indicating negative correlation (p < 0.05); the thickness of the lines indicates the size of the correlation coefficient; the coarser the line is, the greater the correlation between the species; and the greater the number of lines is, the closer the connection between the species and other species.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Amounts of N, P, and K applied in the different fertilization treatments.
| Treatment | N Fertilizer (mg/kg) | P Fertilizer (mg/kg) | K Fertilizer (mg/kg) |
|---|---|---|---|
| NP | 83.6 | 23.6 | - |
| NK | 83.6 | - | 22.2 |
| PK | - | 23.6 | 22.2 |
| NPK | 83.6 | 23.6 | 22.2 |
Table 2.
Amaranthin content in amaranth plants under different fertilization regimens.
| Treatment | Amaranthin Content (mg/g FW) |
|---|---|
| NP | 0.48 ± 0.11 b |
| NK | 0.70 ± 0.28 ab |
| PK | 0.81 ± 0.05 ab |
| NPK | 0.84 ± 0.12 a |
All the data are presented as the means ± standard deviations (SD). Different letters in the same column indicate significant differences among treatments at p < 0.05. NP: conventional fertilization without potassium; NK: conventional fertilization without phosphorus; PK: conventional fertilization without nitrogen; NPK: balanced fertilization. “mg/g FW” indicates milligrams of amaranthin per gram of fresh sample.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Lin, X.; Yang, D.; Zhou, X.; Wei, X.; Xie, Y.; Yang, S. Response of the Endophytic Microbial Composition in Amaranthus Roots to Different Fertilization Treatments. Agronomy 2024, 14, 965. https://doi.org/10.3390/agronomy14050965
AMA Style
Lin X, Yang D, Zhou X, Wei X, Xie Y, Yang S. Response of the Endophytic Microbial Composition in Amaranthus Roots to Different Fertilization Treatments. Agronomy. 2024; 14(5):965. https://doi.org/10.3390/agronomy14050965
Chicago/Turabian StyleLin, Xinru, Da Yang, Xinyan Zhou, Xun Wei, Yuanyuan Xie, and Shangdong Yang. 2024. "Response of the Endophytic Microbial Composition in Amaranthus Roots to Different Fertilization Treatments" Agronomy 14, no. 5: 965. https://doi.org/10.3390/agronomy14050965
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.