
Effects of Different Conditioners on Soil Microbial Community and Labile Organic Carbon Fractions under the Combined Application of Swine Manure and Straw in Black Soil
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Cite
2.2. Experimental Design
2.3. Soil Sampling and Analysis
2.4. Analysis Methods
2.5. Statistical Analysis
3. Results
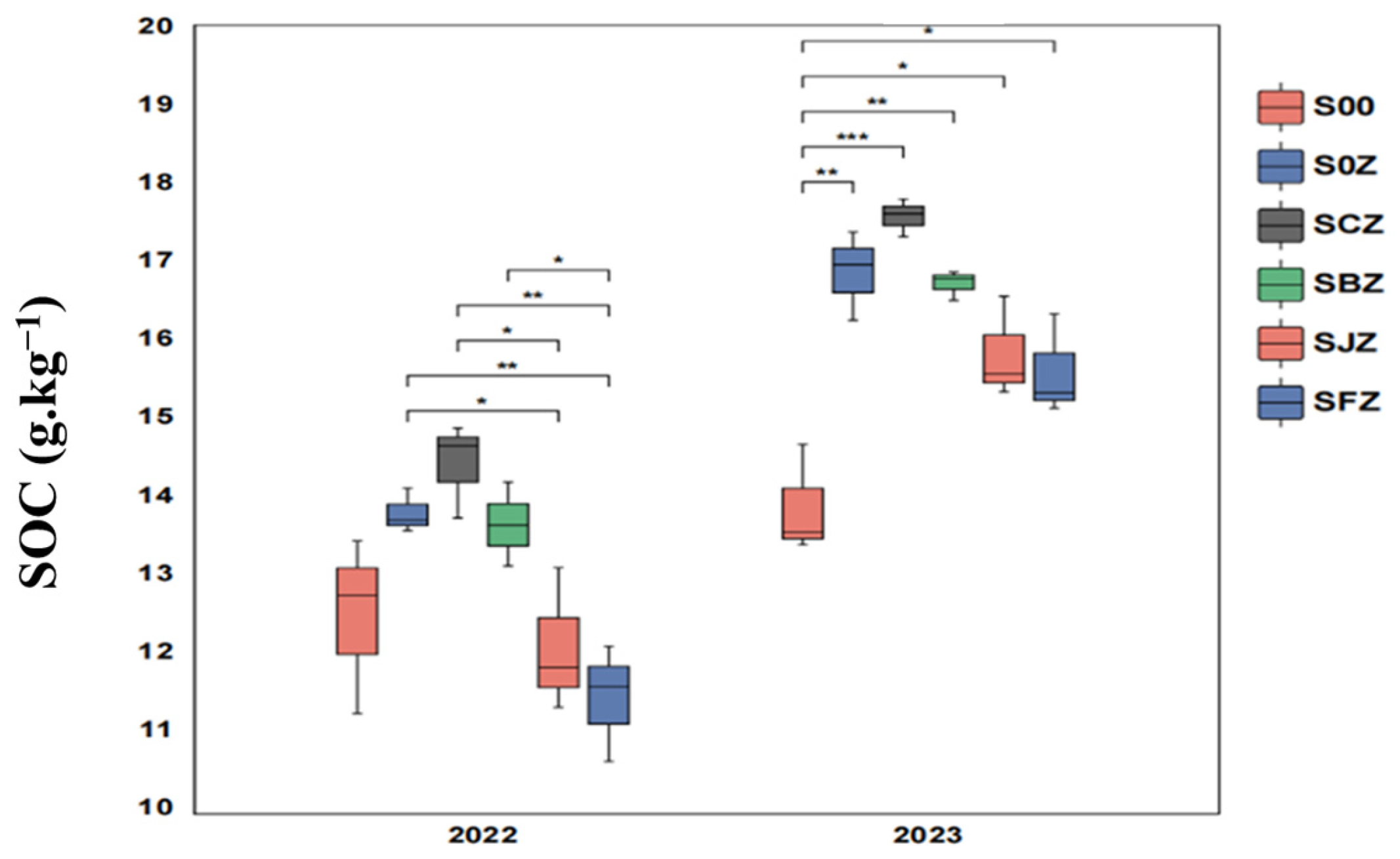
3.1. SOC Content
3.2. LOC Fractions
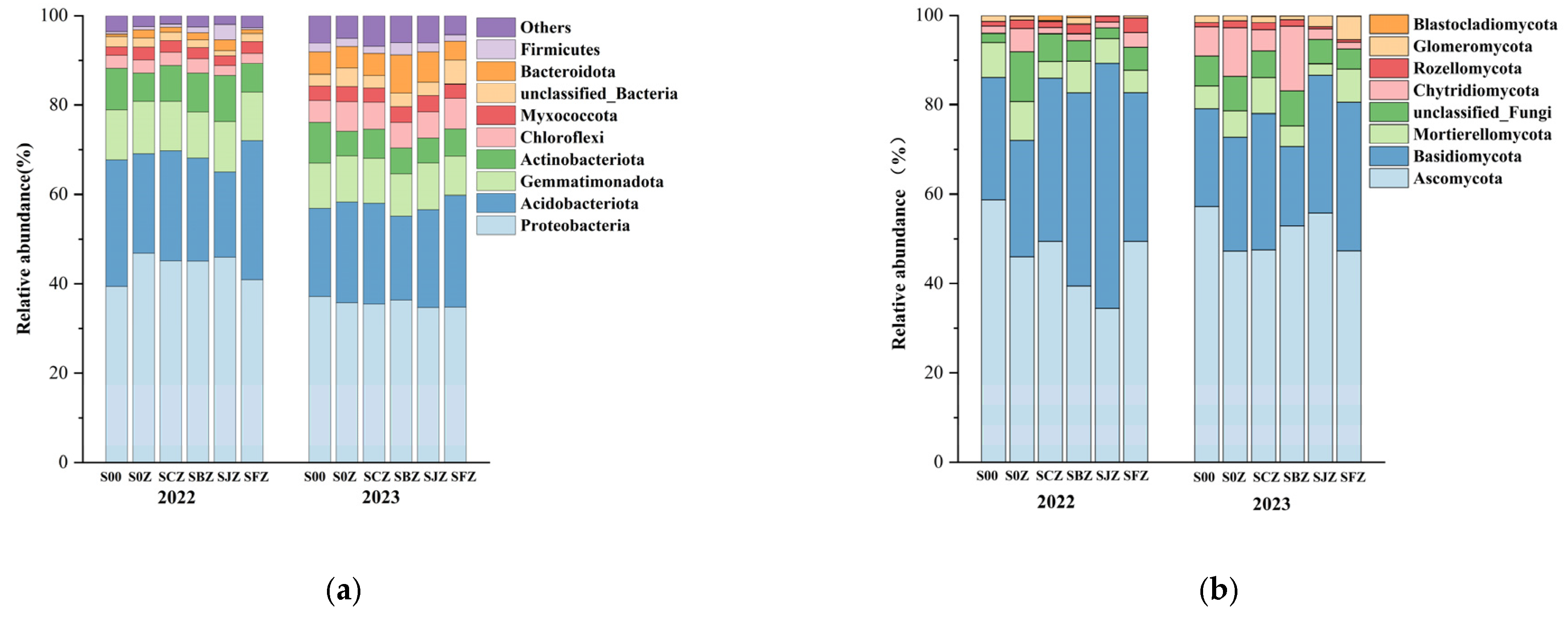
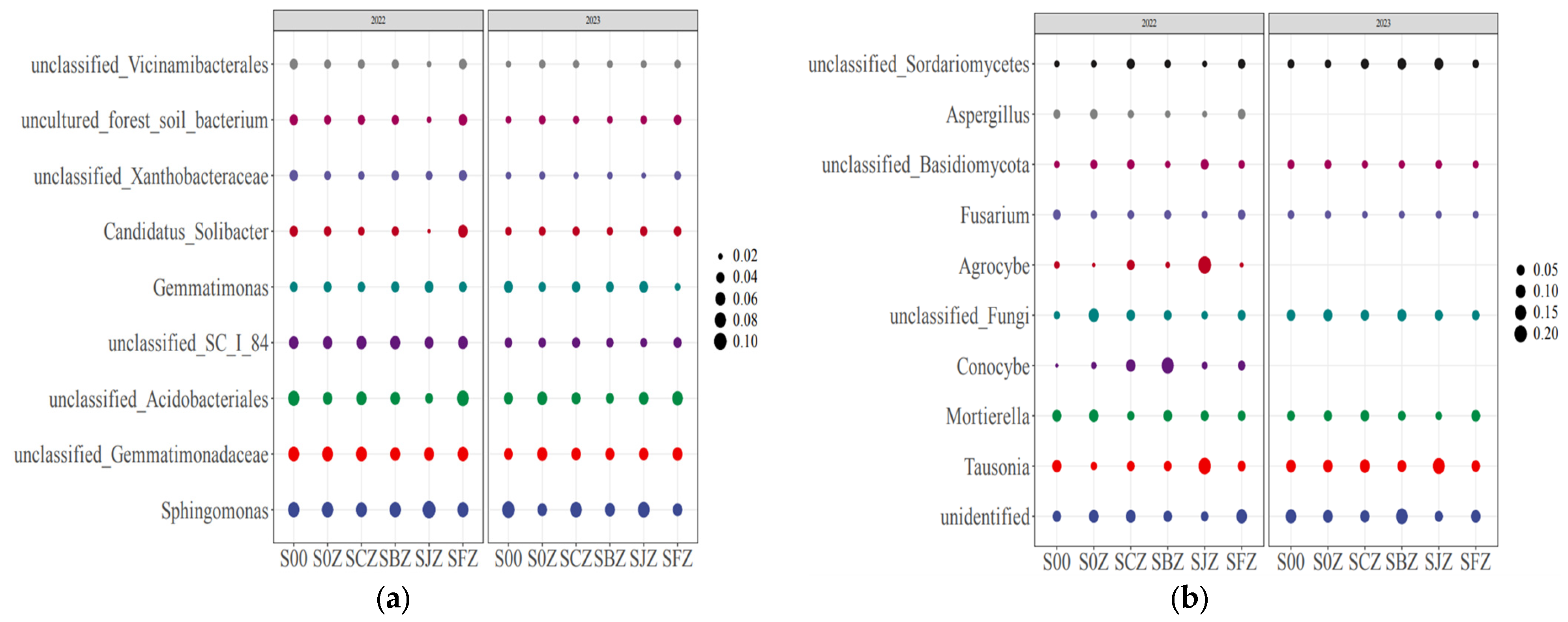
3.3. Soil Microbial Community Structure
3.4. Relationships between Microbial Community and Organic Carbon Fractions
4. Discussion
4.1. Effects of the Application of Manure and Straw on SOC Fractions
4.2. Effects of the Application of Manure and Straw on Soil Microbial Diversity
4.3. Relationship between Soil Microbial Communities and Soil Organic Carbon Components
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Bernhardt, E.S.; Schlesinger, W.H. The Carbon Cycle of Terrestrial Ecosystems, Biogeochemistry, 4th ed.; Elsevier Inc.: New York, NY, USA, 2020; pp. 141–182. [Google Scholar]
- Hou, G.L.; Delang, C.O.; Lu, X.X.; Gao, L. A meta-analysis of changes in soil organic carbon stocks after afforestation with deciduous broadleaved, sempervirent broadleaved, and conifer tree species. Ann. For. Sci. 2020, 77, 92. [Google Scholar] [CrossRef]
- Zhao, Y.; Wang, M.; Hu, S.; Zhang, X.; Shi, X. Economics- and policy-driven organic carbon input enhancement dominates soil organic carbon accumulation in Chinese croplands. Proc. Natl. Acad. Sci. USA 2018, 115, 4045–4050. [Google Scholar] [CrossRef] [PubMed]
- Mclauchlan, K.K.; Hobbie, S.E. Comparison of Labile Soil Organic Matter Fractionation Techniques. Soil Sci. Soc. Am. J. 2004, 68, 1616–1625. [Google Scholar] [CrossRef]
- Lan, J. Responses of soil organic carbon components and their sensitivity to karst rocky desertification control measures in Southwest China. J. Soils Sediments 2021, 21, 978–989. [Google Scholar] [CrossRef]
- Plaza-Bonilla, D.; álvaro-Fuentes, J.; Cantero-Martínez, C. Identifying soil organic carbon fractions sensitive to agricultural management practices. Soil Tillage Res. 2014, 139, 19–22. [Google Scholar] [CrossRef]
- Zhang, K.L.; Maltais-Landry, G.; Liao, H.L. How soil biota regulate C cycling and soil C pools in diversified crop rotations. Soil Boil. Biochem. 2021, 156, 108219. [Google Scholar] [CrossRef]
- Bitas, V.; Kim, H.S.; Bennett, J.W.; Kang, S. Sniffing on microbes: Diverse roles of microbial volatile organic compounds in plant health. Mol. Plant-Microbe Interact. 2013, 26, 835–843. [Google Scholar] [CrossRef] [PubMed]
- Kravchenko, I.; Tikhonova, E.; Semenov, V.M. Temperature Sensitivity of Litter and Soil Organic Matter Decomposition: Perspective of Soil Microbial Community Structure and Function; Elsevier eBooks: Amsterdam, The Netherlands, 2021; pp. 1–43. [Google Scholar]
- Song, Y.; Zhai, J.; Zhang, J.; Qiao, L.; Xue, S. Forest management practices of Pinus tabulaeformis plantations alter soil organic carbon stability by adjusting microbial characteristics on the Loess Plateau of China. Sci. Total Environ. 2020, 766, 144209. [Google Scholar] [CrossRef] [PubMed]
- Zhao, F.Z.; Ren, C.J.; Zhang, L.; Han, X.H.; Wang, J. Changes in soil microbial community are linked to soil carbon fractions after afforestation. Eur. J. Soil Sci. 2018, 69, 370–379. [Google Scholar] [CrossRef]
- Li, Y.; Sun, Y.; Kuramae, E.E.; Zhang, S.; Wang, E.; Yao, Z.; Nasir, F.; Tian, L.; Gao, Q.; Tian, C. Carbohydrate Metabolism Bacteria Positive Effect Determines the Increasing Soil Organic Carbon during Long-Term Straw Fertilization Returning. Pedosphere 2023, in press. [Google Scholar] [CrossRef]
- Gravuer, K.; Gennet, S.; Throop, H.L. Organic amendment additions to rangelands: A meta-analysis of multiple ecosystem outcomes. GCB Bioenergy 2019, 25, 1152–1170. [Google Scholar] [CrossRef] [PubMed]
- Carlson, J.; Saxena, J.; Basta, N.; Hundal, L.; Busalacchi, D.; Dick, R.P. Application of organic amendments to restore degraded soil: Effects on soil microbial properties. Environ. Monit. Assess. 2015, 187, 109. [Google Scholar] [CrossRef]
- Li, B.; Song, H.; Cao, W.; Wang, Y.; Chen, J.; Guo, J. Responses of soil organic carbon stock to animal manure application: A new global synthesis integrating the impacts of agricultural managements and environmental conditions. GCB Bioenergy 2021, 27, 5356–5367. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Liang, F.; Wang, Y.; Cao, W.; Song, H.; Chen, J.; Guo, J. Magnitude and efficiency of straw return in building up soil organic carbon: A global synthesis integrating the impacts of agricultural managements and environmental conditions. Sci. Total Environ. 2023, 875, 162670. [Google Scholar] [CrossRef] [PubMed]
- Rumpel, C.; Amiraslani, F.; Bossio, D.; Chenu, C.; Cardenas, M.G.; Henry, B.; Espinoza, A.F.; Koutika, L.S.; Ladha, J.; Madari, B.E. Studies from global regions indicate promising avenues for maintaining and increasing soil organic carbon stocks. Reg. Environ. Chang. 2023, 23, 8. [Google Scholar] [CrossRef]
- Marousek, J.; Minofar, B.; Marouskova, A.; Strunecky, O.; Gavurova, B. Environmental and economic advantages of production and application of digestate biochar. Environ. Sci Technol. 2023, 30, 103109. [Google Scholar] [CrossRef]
- Chen, L.; Sun, S.; Zhou, Y.; Zhang, B.; Peng, Y.; Zhuo, Y.; Ai, W.; Gao, C.; Wu, B.; Liu, D.; et al. Straw and straw biochar differently affect fractions of soil organic carbon and microorganisms in farmland soil under different water regimes. Environ. Technol. Innov. 2023, 32, 103412. [Google Scholar] [CrossRef]
- Yuan, G.; Huan, W.; Song, H.; Lu, D.; Zhou, J. Effects of straw incorporation and potassium fertilizer on crop yields, soil organic carbon, and active carbon in the rice–wheat system. Soil Tillage Res. 2021, 209, 104958. [Google Scholar] [CrossRef]
- Salehi, A.; Fallah, S.; Sourki, A.A. Organic and inorganic fertilizer effect on soil CO2 flux, microbial biomass, and growth of Nigella sativa L. Int. Agrophysics 2017, 31, 103–116. [Google Scholar] [CrossRef]
- Sedlar, O.; Balik, J.; Cerny, J.; Kulhanek, M.; Smatanova, M. Long-Term Application of Organic Fertilizers in Relation to Soil Organic Matter Quality. Agronomy 2023, 13, 175. [Google Scholar] [CrossRef]
- Guo, Z.; Han, J.; Li, J.; Xu, Y.; Wang, X.; Singer, A.C. Effects of long-term fertilization on soil organic carbon mineralization and microbial community structure. PLoS ONE 2019, 14, e0211163. [Google Scholar]
- Wang, L.W.; Ok, Y.S.; Tsang, D.C.W.; Shen, Z.; Hou, D. Biochar Aging: Mechanisms, Physicochemical Changes, Assessment, And Implications for Field Applications. Environ. Sci. Technol. 2020, 54, 14797–14814. [Google Scholar] [CrossRef] [PubMed]
- Chen, K.; Li, N.; Zhang, S.; Liu, N.; Yang, J.; Zhan, X.; Han, X. Biochar-induced changes in the soil diazotroph community abundance and structure in a peanut field trial. Biochar 2022, 4, 412–426. [Google Scholar] [CrossRef]
- Khan, N.; Bolan, N.; Jospeh, S.; Anh, M.T.L.; Meier, S.; Kookana, R.; Borchard, N.; Sánchez-Monedero, M.A.; Jindo, K.; Solaiman, Z.M.; et al. Complementing compost with biochar for agriculture, soil remediation and climate mitigation. Adv. Agron. 2023, 179, 1–90. [Google Scholar]
- Wang, X.; Wang, X.; Geng, P.; Yang, Q.; Han, X. Effects of different returning method combined with decomposer on decomposition of organic components of straw and soil fertility. Sci. Rep. 2021, 11, 15495. [Google Scholar] [CrossRef] [PubMed]
- Huang, D.Z.; Zhang, M.B.; An, P.P.; Zhang, M.; Zhang, X.; Li, Z.B.; Li, C.H. Effect of application amount of boron fertilizer on growth, development and grain yield of maize. J. Maize Sci. 2018, 26, 118–124. [Google Scholar]
- Xu, Z.; Sun, R.; He, T.; Sun, Y.; Wu, M.; Xue, Y.; Meng, F.; Wang, J. Disentangling the impact of straw incorporation on soil microbial communities: Enhanced network complexity and ecological stochasticity. Sci. Total Environ. 2023, 863, 160918. [Google Scholar] [CrossRef] [PubMed]
- Tong, L.; Zhu, L.; Lv, Y.; Zhu, K.; Liu, X.; Zhao, R. Response of organic carbon fractions and microbial community composition of soil aggregates to long-term fertilizations in an intensive greenhouse system. J. Soils Sediments 2020, 20, 641–652. [Google Scholar] [CrossRef]
- Ramirez, K.S.; Lauber, C.L.; Knight, R.; Bradford, M.A.; Fierer, N. Consistent effects of nitrogen fertilization on soil bacterial communities in contrasting systems. Ecology 2010, 91, 3463–3470. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Lu, M.; Cui, J.; And, B.L.; Fang, C. Effects of straw carbon input on carbon dynamics in agricultural soils: A meta-analysis. GCB Bioenergy 2014, 20, 1366–1381. [Google Scholar] [CrossRef] [PubMed]
- Jensen, M.B.; Jonge, N.D.; Dolriis, M.D.; Kragelund, C.; Fischer, C.H.; Eskesen, M.R.; Noer, K.; Mller, H.B.; Ottosen, L.D.M.; Nielsen, J.L. Cellulolytic and Xylanolytic Microbial Communities Associated with Lignocellulose-Rich Wheat Straw Degradation in Anaerobic Digestion. Front. Microbiol. 2021, 12, 645174. [Google Scholar] [CrossRef] [PubMed]
- Che, W.K.; Piao, J.L.; Gao, Q.; Li, X.B.; Li, X.; Jin, F. Response of Soil Physicochemical Properties, Soil Nutrients, Enzyme Activity and Rice Yield to Rice Straw Returning in Highly Saline-Alkali Paddy Soils. J. Soil Sci. Plant Nutr. 2023, 23, 4396–4411. [Google Scholar] [CrossRef]
- Zhou, G.P.; Gao, S.J.; Lu, Y.H.; Liao, Y.L.; Nie, J.; Cao, W.D. Co-incorporation of green manure and rice straw improves rice production, soil chemical, biochemical and microbiological properties in a typical paddy field in southern China. Soil Tillage Res. 2020, 197, 104499. [Google Scholar] [CrossRef]
- Sun, L.; Sun, Z.; Hu, J.; Opoku-Kwanowaa, Y.; Wu, J. Combined application of animal manure and stover enhance labile organic carbon and benefit the microbial community in dark brown soil. Arch. Agron. Soil Sci. 2023, 69, 519–532. [Google Scholar] [CrossRef]
- Sun, R.; Dsouza, M.; Gilbert, J.A.; Guo, X.; Wang, D.; Guo, Z.; Ni, Y.; Chu, H. Fungal community composition in soils subjected to long-term chemical fertilization is most influenced by the type of organic matter. Environ. Microbiol. 2016, 18, 5137–5150. [Google Scholar] [CrossRef]
- Li, X.; Song, Y.; Wang, F.; Bian, Y.; Jiang, X. Combined effects of maize straw biochar and oxalic acid on the dissipation of polycyclic aromatic hydrocarbons and microbial community structures in soil: A mechanistic study. J. Hazard. Mater. 2018, 364, 325–331. [Google Scholar] [CrossRef]
- Breulmann, M.; Masyutenko, N.P.; Kogut, B.M.; Schroll, R.; Doerfler, U.; Buscot, F.; Schulz, E. Short-term bioavailability of carbon in soil organic matter fractions of different particle sizes and densities in grassland ecosystems. Sci. Total Environ. 2014, 497–498, 29–37. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Ding, W.; Yu, H.; He, X. Linking organic carbon accumulation to microbial community dynamics in a sandy loam soil: Result of 20 years compost and inorganic fertilizers repeated application experiment. Biol. Fertil. Soils 2015, 51, 137–150. [Google Scholar] [CrossRef]
- Morugan-Coronado, A.; Perez-Rodriguez, P.; Insolia, E.; Soto-Gomez, D.; Fernandez-Calvino, D.; Zornoza, R. The impact of crop diversification, tillage and fertilization type on soil total microbial, fungal and bacterial abundance: A worldwide meta-analysis of agricultural sites. Agric. Ecosyst. Environ. 2022, 329, 107867. [Google Scholar] [CrossRef]
- Blair, G.; Lefroy, R.; Lisle, L. Soil carbon fractions based on their degree of oxidation, and the development of a carbon management index for agricultural systems. Aust. J. Agric. Res. 1995, 46, 393–406. [Google Scholar] [CrossRef]
- Ouyang, W.; Shan, Y.; Hao, F.; Chen, S.; Pu, X.; Wang, M.K. The effect on soil nutrients resulting from land use transformations in a freeze-thaw agricultural ecosystem. Soil Tillage Res. 2013, 132, 30–38. [Google Scholar] [CrossRef]
- Jiang, P.K.; Xu, Q.F.; Xu, Z.H.; Cao, Z.H. Seasonal changes in soil labile organic carbon pools within a Phyllostachys praecox stand under high rate fertilization and winter mulch in subtropical China. For. Ecol. Manag. 2006, 236, 30–36. [Google Scholar] [CrossRef]
- Vance, E.D.; Brookes, P.C.; Jenkinson, D.S. An extraction method for measuring soil microbial biomass C. Soil Biol. Biochem. 1987, 19, 703–707. [Google Scholar] [CrossRef]
- Ma, Q.; Wen, Y.; Wang, D.; Sun, X.; Hill, P.W.; Macdonald, A.; Chadwick, D.R.; Wu, L.; Jones, D.L. Farmyard Manure Applications Stimulate Soil Carbon and Nitrogen Cycling by Boosting Microbial Biomass Rather than Changing Its Community Composition. Soil Biol. Biochem. 2020, 144, 107760. [Google Scholar] [CrossRef]
- Yu, Q.; Hu, X.; Ma, J.; Ye, J.; Sun, W.; Wang, Q.; Lin, H. Effects of long-term organic material applications on soil carbon and nitrogen fractions in paddy fields. Soil Tillage Res. 2020, 196, 104483. [Google Scholar] [CrossRef]
- Li, H.; Wang, Y.; Qin, L.; He, H.; Zhang, T.; Wang, J.; Zheng, X. Effects of different slopes and fertilizer types on the grey water footprint of maize production in the black soil region of China. J. Clean. Prod. 2020, 246, 119077. [Google Scholar] [CrossRef]
- Novak, J.M.; Busscher, W.J.; Laird, D.L.; Ahmedna, M.; Watts, D.W.; Niandou, M. Impact of Biochar Amendment on Fertility of a Southeastern Coastal Plain Soil. Soil Sci. 2009, 174, 105–112. [Google Scholar] [CrossRef]
- Lal, R. Soil carbon sequestration impacts on global climate change and food security. Science 2004, 304, 1623–1627. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Li, H.; Yao, T.; Su, M.; Gun, S. Effects of microbial inoculation on enzyme activity, available nitrogen content, and bacterial succession during swine manure composting. Bioresour. Technol. 2020, 306, 123167. [Google Scholar] [CrossRef] [PubMed]
- Yang, F.; Xu, Z.; Huang, Y.; Tsang, D.C.W.; Ok, Y.S.; Zhao, L.; Qiu, H.; Xu, X.; Cao, X. Stabilization of Dissolvable Biochar by Soil Minerals: Release Reduction and Organo-Mineral Complexes Formation. J. Hazard. Mater. 2021, 412, 125213. [Google Scholar] [CrossRef] [PubMed]
- Blanco-Canqui, H. Biochar and Soil Physical Properties. Soil Sci. Soc. Am. J. 2017, 81, 687–711. [Google Scholar] [CrossRef]
- Zimmerman, A.R.; Gao, B.; Ahn, M.Y. Positive and negative carbon mineralization priming effects among a variety of biochar-amended soils. Soil Boil. Biochem. 2011, 43, 1169–1179. [Google Scholar] [CrossRef]
- Burrell, L.D.; Zehetner, F.; Rampazzo, N.; Wimmer, B.; Soja, G. Long-term effects of biochar on soil physical properties. Geoderma 2016, 282, 96–102. [Google Scholar] [CrossRef]
- Pokharel, P.; Ma, Z.; Chang, S.X. Biochar increases soil microbial biomass with changes in extra- and intracellular enzyme activities: A global meta-analysis. Biochar 2021, 2, 65–79. [Google Scholar] [CrossRef]
- Joseph, S.; Graber, E.R.; Chia, C.; Munroe, P.; Donne, S.; Thomas, T.; Nielsen, S.; Marjo, C.; Rutlidge, H.; Pan, G.X. Shifting Paradigms: Development of High-Efficiency Biochar Fertilizers Based on Nano-Structures and Soluble Components. Carbon Manag. 2013, 4, 323–343. [Google Scholar] [CrossRef]
- Wang, H. Biochar and Organic Fertilizer Applications Enhance Soil Functional Microbial Abundance and Agroecosystem Multifunctionality. Biochar 2024, 6, 3. [Google Scholar]
- Khan, M.A.; Kim, K.W.; Mingzhi, W.; Lim, B.K.; Lee, W.H.; Lee, J.Y. Nutrient-Impregnated Charcoal: An Environmentally Friendly Slow-Release Fertilizer. Environmentalist 2008, 28, 231–235. [Google Scholar] [CrossRef]
- Singh, B.P.; Hatton, B.J.; Balwant Singh, B.S.; Cowie, A.L. The Role of Biochar in Reducing Nitrous Oxide Emissions and Nitrogen Leaching from Soil. In Proceedings of the 19th World Congress of Soil Science: Soil Solutions for a Changing World, Brisbane, Australia, 1–6 August 2010; pp. 257–259. [Google Scholar] [CrossRef]
- Craig, A.; Leo, M.C.; Tim, J.C.; Mark, F.; Alison, S.; Robert, A.H.; Robert, R.S. Biochar Induced Soil Microbial Community Change: Implications for Biogeochemical Cycling of Carbon, Nitrogen and Phosphorus. Pedobiologia 2011, 54, 309–320. [Google Scholar]
- Dempster, D.N.; Gleeson, D.B.; Solaiman, Z.M. Decreased soil microbial biomass and nitrogen mineralisation with Eucalyptus biochar addition to a coarse textured soil. Plant Soil 2012, 354, 311–324. [Google Scholar] [CrossRef]
- Xu, X.K.; Luo, X.B.; Xu, J.Z. Biodegradation of dissolved organic carbon in soil extracts and leachates from a temperate forest stand and its relationship to ultraviolet absorbance. Chin. Sci. Bull. 2012, 57, 912–920. [Google Scholar] [CrossRef]
- Liu, F.; Chen, L.; Zhang, B.; Wang, G.; Qin, S.; Yang, Y. Ultraviolet radiation rather than inorganic nitrogen increases dissolved organic carbon biodegradability in a typical thermo-erosion gully on the Tibetan Plateau. Sci. Total Environ. 2018, 627, 1276–1284. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Gong, J.R.; Liu, M.; Luo, Q.; Xu, S.; Pan, Y.; Zhai, Z. Effects of land use and precipitation on above- and below-ground litter decomposition in a semi-arid temperate steppe in Inner Mongolia, China. Appl. Soil Ecol. 2015, 96, 183–191. [Google Scholar] [CrossRef]
- Zhu, Z.; Bai, Y.; Lv, M.; Tian, G.; Ge, S. Soil Fertility, Microbial Biomass, and Microbial Functional Diversity Responses to Four Years Fertilization in an Apple Orchard in North China. Hortic. Plant J. 2020, 6, 223–230. [Google Scholar] [CrossRef]
- Philippot, L.; Raaijmakers, J.M.; Lemanceau, P.; van der Putten, W.H. Going back to the roots: The microbial ecology of the rhizosphere. Nat. Rev. Microbiol. 2013, 11, 789–799. [Google Scholar] [CrossRef] [PubMed]
- Chakraborty, A.; Chakrabarti, K.; Chakraborty, A.; Ghosh, S. Effect of long-term fertilizers and manure application on microbial biomass and microbial activity of a tropical agricultural soil. Biol. Fertil. Soils 2011, 47, 227–233. [Google Scholar] [CrossRef]
- Melero, S.; López-Garrido, R.; Murillo, J.M.; Moreno, F. Conservation tillage: Short- and long-term effects on soil carbon fractions and enzymatic activities under Mediterranean conditions. Soil Tillage Res. 2009, 104, 292–298. [Google Scholar] [CrossRef]
- Liang, Y.; Al-Kaisi, M.; Yuan, J.; Liu, J.; Zhang, H.; Wang, L.; Cai, H.; Ren, J. Effect of chemical fertilizer and straw-derived organic amendments on continuous maize yield, soil carbon sequestration and soil quality in a Chinese Mollisol. Agric. Ecosyst. Environ. 2021, 314, 107403. [Google Scholar] [CrossRef]
- Lützow, M.; Kögel-Knabner, I.; Ekschmitt, K.; Flessa, H.; Guggenberger, G.; Matzner, E.; Marschner, B. SOM fractionation methods: Relevance to functional pools and to stabilization mechanisms. Soil Biol. Biochem. 2007, 39, 2183–2207. [Google Scholar] [CrossRef]
- Luan, H.A.; Gao, W.; Huang, S.W.; Tang, J.W.; Li, M.Y. Partial substitution of chemical fertilizer with organic amendments affects soil organic carbon composition and stability in a greenhouse vegetable production system. Soil Tillage Res. 2019, 191, 185–196. [Google Scholar] [CrossRef]
- Shi, S.; Zhang, Q.; Lou, Y.; Du, Z.; Wang, Q.; Hu, N.; Wang, Y.; Gunina, A.; Song, J. Soil organic and inorganic carbon sequestration by consecutive biochar application: Results from a decade field experiment. Soil Use Manag. 2021, 37, 95–103. [Google Scholar] [CrossRef]
- Jones, R.T.; Robeson, M.S.; Lauber, C.L.; Hamady, M.; Knight, R.; Fierer, N. A comprehensive survey of soil acidobacterial diversity using pyrosequencing and clone library analyses. Isme J. 2009, 3, 442. [Google Scholar] [CrossRef] [PubMed]
- Núria, M.; Mustapha, B.; Jordi, E.; Florence, L.; Gaetan, L.; Catherine, D.; Jean, J.L.; Claire, C.; Yvon, B.; Joël, D.; et al. Early-Life Establishment of the Swine Gut Microbiome and Impact on Host Phenotypes. Environ. Microbiol. Rep. 2015, 7, 554–569. [Google Scholar]
- Zhang, X.; Zhao, L.; Xu, S.; Liu, Y.; Liu, H.; Cheng, G. Soil moisture effect on bacterial and fungal community in Beilu River (Tibetan Plateau) permafrost soils with different vegetation types. J. Appl. Microbiol. 2013, 114, 1054–1065. [Google Scholar] [CrossRef] [PubMed]
- Yelle, D.J.; Ralph, J.; Lu, F.; Hammel, K.E. Evidence for cleavage of lignin by a brown rot basidiomycete. Environ. Microbiol. 2008, 10, 1844–1849. [Google Scholar] [CrossRef] [PubMed]
- Durling, M.B.; Sterkenburg, E.; Bahr, A.; Lindahl, B.D.; Clemmensen, K.E. Changes in fungal communities along a boreal forest soil fertility gradient. New Phytol. 2015, 207, 1145–1158. [Google Scholar]
- Tuulia, M.; Jaana, K.; Firoz, S.; Kristina, L.T.; Daniel, C. Interactions affect hyphal growth and enzyme profiles in combinations of coniferous wood-decaying fungi of Agaricomycetes. PLoS ONE 2017, 12, e0185171. [Google Scholar]
- Blackwood, C.B.; Waldrop, M.P.; Zak, D.R.; Sinsabaugh, R.L. Molecular analysis of fungal communities and laccase genes in decomposing litter reveals differences among forest types but no impact of nitrogen deposition. Environ. Microbiol. 2007, 9, 1306–1316. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Kong, Z.; Wang, Y.; Wang, M.; Shen, Q. Insights into the functionality of fungal community during the large scale aerobic co-composting process of swine manure and rice straw. J. Environ. Manag. 2020, 270, 110958. [Google Scholar] [CrossRef] [PubMed]
- Shen, Z.; Penton, C.R.; Lv, N.; Xue, C.; Yuan, X.; Ruan, Y.; Li, R.; Shen, Q. Banana Fusarium Wilt Disease Incidence Is Influenced by Shifts of Soil Microbial Communities Under Different Monoculture Spans. Microb. Ecol. 2017, 75, 739–750. [Google Scholar] [CrossRef]
- Wang, C.; Chen, D.; Shen, J.L.; Yuan, Q.; Fan, F.L.; Wei, W.X.; Li, Y.; Wu, J.S. Biochar alters soil microbial communities and potential functions 3–4 years after amendment in a double rice cropping system. Agric. Ecosyst. Environ. 2021, 311, 107291. [Google Scholar] [CrossRef]
- Ning, L.; Chen, Z.; Chao, X.; Huan, C.; Duan, Y.H. Insight into how organic amendments can shape the soil microbiome in long-term field experiments as revealed by network analysis. Soil Biol. Biochem. 2016, 99, 137–149. [Google Scholar]
- Sun, R.; Zhang, X.X.; Guo, X.; Wang, D.; Chu, H. Bacterial diversity in soils subjected to long-term chemical fertilization can be more stably maintained with the addition of livestock manure than wheat straw. Soil Biol. Biochem. 2015, 88, 9–18. [Google Scholar] [CrossRef]
- Liu, X.; Li, Y.J.; Ren, X.J.; Chen, B.H.; Zhang, Y.; Shen, C.W.; Wang, F.; Wu, D.F. Long-Term Greenhouse Cucumber Production Alters Soil Bacterial Community Structure. J. Soil Sci. Plant Nutr. 2020, 2, 306–321. [Google Scholar] [CrossRef]
- Fierer, N.; Bradford, M.A.; Jackson, R.B. Toward an ecological classification of soil bacteria. Ecology 2007, 88, 1354–1364. [Google Scholar] [CrossRef] [PubMed]
- Ren, C.; Zhao, F.; Kang, D.; Yang, G.; Han, X.; Tong, X.; Feng, Y.; Ren, G. Linkages of C:N:P stoichiometry and bacterial community in soil following afforestation of former farmland. For. Ecol Manag. 2016, 376, 59–66. [Google Scholar] [CrossRef]
- Fierer, N.; Strickland, M.S.; Liptzin, D.; Bradford, M.A.; Cleveland, C.C. Global patterns in belowground communities. Ecol. Lett. 2009, 12, 1238–1249. [Google Scholar] [CrossRef] [PubMed]
- Brundrett, M.C.; Ashwath, N. Glomeromycotan mycorrhizal fungi from tropical Australia III. Measuring diversity in natural and disturbed habitats. Plant Soil 2013, 370, 419–433. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Materials | Total Organic Carbon (g·kg−1) | Total Nitrogen (g·kg−1) | Total Phosphorous (g·kg−1) | Total Potassium (g·kg−1) | pH | EC (μs·cm−1) |
|---|---|---|---|---|---|---|
| Maize straw | 424.03 | 8.46 | 1.18 | 10.27 | 7.02 | 1.7 |
| Biochar | 330.87 | 8.9 | 15.96 | 2.18 | 7.07 | 1.3 |
| Swine manure | 280.52 | 20.04 | 0.9 | 22.15 | 7.04 | 3.89 |
| Biological-organic fertilizer | 255.63 | 17.08 | 7.82 | 14.11 | 8.06 | 3.7 |
| Biological agent | 446.15 | 8.37 | 4.27 | 12.32 | 6.51 | ---- |
| Boron slag | ---- | ---- | ---- | ---- | 3.66 | ---- |
| Time (Year) | Treatments | Bacterial Community | Fungi Community | ||
|---|---|---|---|---|---|
| Simpson Index | Chao1 Index | Simpson Index | Chao1 Index | ||
| 2022 | S00 | 0.994 abcd | 697 c | 0.945 a | 298 c |
| S0Z | 0.994 cd | 680 c | 0.982 a | 314 b | |
| SCZ | 0.991 d | 600 c | 0.982 a | 278 c | |
| SBZ | 0.993 abc | 583 c | 0.8 a | 245 c | |
| SJZ | 0.993 bcd | 696 c | 0.984 b | 413 c | |
| SFZ | 0.993 abcd | 626 c | 0.967 a | 412 b | |
| 2023 | S00 | 0.994 abcd | 1775 ab | 0.982 a | 462 b |
| S0Z | 0.996 ab | 1881 ab | 0.978 a | 505 ab | |
| SCZ | 0.995 abc | 1978 ab | 0.965 a | 531 a | |
| SBZ | 0.996 a | 2061 a | 0.962 a | 494 a | |
| SJZ | 0.996 abc | 1838 ab | 0.887 ab | 537 a | |
| SFZ | 0.996 abc | 1612 b | 0.959 a | 479 ab | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zheng, S.; Wu, J.; Sun, L. Effects of Different Conditioners on Soil Microbial Community and Labile Organic Carbon Fractions under the Combined Application of Swine Manure and Straw in Black Soil. Agronomy 2024, 14, 879. https://doi.org/10.3390/agronomy14050879
Zheng S, Wu J, Sun L. Effects of Different Conditioners on Soil Microbial Community and Labile Organic Carbon Fractions under the Combined Application of Swine Manure and Straw in Black Soil. Agronomy. 2024; 14(5):879. https://doi.org/10.3390/agronomy14050879
Chicago/Turabian StyleZheng, Shuang, Jinggui Wu, and Liming Sun. 2024. "Effects of Different Conditioners on Soil Microbial Community and Labile Organic Carbon Fractions under the Combined Application of Swine Manure and Straw in Black Soil" Agronomy 14, no. 5: 879. https://doi.org/10.3390/agronomy14050879