
Mitigating Effect of Exogenous Melatonin on Salt and Drought Stress in Cyperus esculentus L. during the Tillering Stage

Agriculture Department, Shihezi University, Shihezi 832003, China
*
Author to whom correspondence should be addressed.
Agronomy 2024, 14(5), 1009; https://doi.org/10.3390/agronomy14051009
Submission received: 1 April 2024
/
Revised: 27 April 2024
/
Accepted: 8 May 2024
/
Published: 10 May 2024
(This article belongs to the Section Agroecology Innovation: Achieving System Resilience)
Abstract
:Drought and salt stress are important limiting factors that affect crop growth and yield. As a newly recognized plant hormone, melatonin can participate in the regulation of plant stress tolerance and enhance the tolerance of plants to adversity. In this study, the effects of melatonin (150 µmol·L−1) on agronomic traits, osmotic adjustment substances, antioxidant enzyme activities, and reactive oxygen species content in C. esculentus under different salt (0 and 200 mmol·L−1 NaCl), drought (70% field capacity, 50% field capacity), and salt–drought (200 mmol·L−1 NaCl + 50% field capacity) stress conditions were determined using a pot experiment. Spraying with 150 µmol·L−1 of melatonin effectively improved the plant height, number of blades, biomass, and root growth of C. esculentus seedlings under salt, drought, and combined stress. In addition, this treatment also increased the relative water content, superoxide dismutase, peroxidase, and catalase activities, and soluble sugar content of the blades and decreased the relative electroconductivity conductivity and proline, malondialdehyde, hydrogen peroxide, and superoxide anion contents. A comprehensive analysis showed that spraying the plants with exogenous melatonin could increase the activity of antioxidant enzymes and the accumulation of osmotic adjustment substances in C. esculentus blades under salt and drought stress, effectively remove excessive reactive oxygen species, alleviate oxidative damage, and enhance the ability of C. esculentus to resist salt and drought stress.
1. Introduction
As the most widely distributed abiotic stress factors in the world, drought and salinity stress can significantly inhibit plant growth, development, and crop yield, have adverse effects on global agricultural productivity, and seriously endanger the sustainable development of agriculture [1,2]. According to the statistics, about 33% of the world’s arable land is affected by periodic or unpredictable drought, and 20% of arable land is affected by salt stress, especially in arid and semi-arid areas affected by global climate change [3,4]. It is expected that the occurrence of drought and salinity stress will increase in the future [5]. In recent years, due to global climate change, the occurrence of drought stress has mostly been accompanied by the occurrence of salt stress, and the combined stressors usually cause more damage to crops than a single stressor, resulting in a serious reduction in global crop yields [6,7]. Therefore, exploring the effects of salt and drought stress on crop growth and taking certain measures to improve the tolerance of crops to drought and salinity stress has become urgent to increase crop yield.
To cope with the adverse environmental stress caused by the external environment and to maintain cell homeostasis and the normal growth and development of a plant, a series of physiological changes occur in the plant [8]. Drought stress and salt stress are two important abiotic stress factors that limit plant growth [9]. Drought and salt stress can cause a decrease in a plant’s number of blades, plant height, root growth, plants’ fresh weight, and their dry weight [10,11]. Although Cyperus esculentus L. itself has a certain level of salt and drought tolerance, stress conditions increase the threshold of reactive oxygen species, which means that C. esculentus plants use physiological, biochemical, and molecular mechanisms to cope with these pressures in the face of external drought and salinity stress [12]. In the process of salt and drought stress, all of the key processes in plant growth, such as photosynthesis, protein synthesis, energy metabolism, and osmotic regulation, are affected to varying degrees [13]. Both drought and salt stress can cause ion toxicity in cells, resulting in osmotic imbalances in plants and thus affecting plant growth and development [14]. Moreover, stress will also lead to a continuous increase in the active oxygen content in plants and a decline in their own defense mechanisms, which will cause membrane lipid peroxidation in plant cells and even apoptosis in severe cases [15]. To resist reactive oxygen species, plants begin to produce antioxidants. As a response, plant enzymes (such as superoxide dismutase (SOD), peroxidase (POD), and catalase (CAT)) and non-enzymatic defense mechanisms (such as osmotic regulators) are activated [16].
In recent years, people have begun to widely use exogenous plant hormones to improve plant stress resistance and yield, and exogenous MT has become a topic of interest in plant science research. Melatonin (N-acetyl-5-methoxytryptamine, MT) is a tryptophan-derived indole compound that was first discovered in animal tissues, and it plays an important role in many physiological processes in humans and animals. Studies on plants have found that it exists in almost all plants as a natural plant hormone [17,18], and some studies have found that melatonin is present in different parts of plants (e.g., roots, stems, blade, fruits, flowers, and seeds), and analogous to mammals, MEL levels in plants are highest in young seedlings and low in mature vegetative tissues, with concentrations increasing during seed production and maturation [19,20]. As a plant growth regulator, MT plays a protective role in the regulation of plant growth and development and in resistance to external stress, such as the occurrence of plant morphological organs, delaying blade senescence, promoting plant growth, and improving plant stress resistance [21]. It has been shown to enhance the stress resistance of maize, cotton, wheat, soybean, and rice under low temperature, drought, and salinity stress [22,23,24]. Studies have shown that MT can reduce excessive reactive oxygen species in plants by increasing both the antioxidant content and antioxidant enzyme activity and by regulating the osmotic pressure balance and the expression of related stress-resistant genes, thereby protecting plant cells from oxidative damage and alleviating stress [25]. Exogenous MT can enhance the salt tolerance of rice by scavenging excessive reactive oxygen species produced by plants under salinity stress and maintaining a dynamic balance of Na+/K+ [26]. Melatonin pretreatment can also improve the germination ability of rice seeds under drought stress and alleviate the inhibitory effect of drought stress on rice growth by improving the antioxidant system [27].
C. esculentus is a perennial herb developed from tubers and it belongs to the Cyperaceae family [28]. It is an important food crop in most parts of the world [29]. These tubers contain a variety of dietary components that are necessary for the human body, such as minerals, fibers, carbohydrates, fatty acids, proteins, and vitamins, and have a high nutritional value [30]. Compared with oil palm, cottonseed, and soybeans, the advantages of C. esculentus mainly manifest in its wide adaptability, short growth period, shorter growth cycle, and higher biomass and oil content [31]. It is a new type of economic crop used for grain, oil, feed, medicine, and other uses [28]. It grows all over the world because of its high yield and broad prospects for comprehensive utilization [32]. It has a certain level of drought tolerance and salt tolerance; thus, it is widely planted under arid saline–alkali soil conditions [33].
At present, C. esculentus is an emerging oil crop. The regulatory mechanism of exogenous MT on the growth of C. esculentus seedlings under drought and salt stress is still unclear. We previously studied the effects of exogenous MT on seed germination and the aboveground and underground growth of C. esculentus under salt and drought stress and combined stress. It was confirmed that exogenous MT promoted seed germination and the aboveground and underground growth of C. esculentus under salt and drought stress (not published). However, the physiological mechanism by which exogenous melatonin affects the growth of C. esculentus is still unclear. Therefore, in this study, 150 µmol·L−1 MT, 200 mmol·L−1 NaCl, and 50% soil water content were used to treat C. esculentus seedlings, and then, the morphological indexes, osmotic adjustment substances, antioxidant system, and enzyme activity of C. esculentus seedlings were determined. This study is based on the hypothesis that an improvement in C. esculentus’ tolerance to drought and salt stress can be achieved by applying exogenous MT. Our objectives were (1) to explore the effects of salt, drought, and salt–drought stress on the growth and biomass accumulation of C. esculentus seedlings and roots under salt and drought stress, as well as the effects of salt and drought stress on cell osmotic adjustment, membrane lipid peroxidation, and the antioxidant enzyme system of C. esculentus blades; (2) to study the effects of exogenous melatonin on the growth and development of C. esculentus under combined salt, drought, and salt–drought stresses; (3) to elucidate the physiological mechanisms by which MT regulates the growth of C. esculentus seedlings under salt and drought stresses.
2. Materials and Methods
2.1. Plant Materials and Treatments
The tests were carried out at the experimental station of the College of Agriculture at Shihezi University. Experiments used salt (NaCl, analytically pure, purchased from Shanghai McLean Biochemical Technology Co. Shanghai, China) and MT (analytically pure, purchased from Shanghai McLean Biochemical Technology Co. Shanghai, China). The experiment used a plastic flowerpot with an inner diameter of 45 cm and a height of 40 cm for outdoor potting cultivation (the soil source used in the experiment was salty natural soil from the Shihezi area), using ‘C. esculentus No. 1’ as the plant material, which was provided by the 54 regiment of the Third Division of Xinjiang Production and Construction Corps. Seeds of C. esculentus with full particles of the same size were soaked and disinfected with 5% sodium hypochlorite for 10 min, rinsed with distilled water five times, and immersed in distilled water at 35 °C in the dark for 48 h to fully imbibe the water. Filter paper was used as the germination bed. The seedlings were cultivated in the dark in a constant temperature incubator at 35 °C, and the same amount of distilled water was added every other day. All plastic flowerpots were buried in the ground to avoid direct sunlight and to maintain the soil temperature. Small holes were left at the bottom of the flowerpots, and gauze was placed over the holes to prevent the roots from growing out of the flowerpots. Sieved soil was mixed with perlite (2:1) and placed in the flowerpots, and the pots were watered. After 5 days of germination on filter paper, seeds with similar signs of germination were selected. The roots were rinsed with distilled water and the plants were moved into plastic flowerpots. There were four plants per pot, and seeds were sown at a depth of 3 cm. There were eight treatments and three repetitions of each treatment, for a total of 24 pots. The pots were watered regularly after sowing, and the weighing method was used to control the soil moisture content at 70% of the field water-holding capacity. Soil moisture was maintained to prevent soil compaction. P2O5 (2.1 g) and K2O (1.75 g) were applied as base fertilizers at the seedling stage.
Treatment began after the plants entered the tillering stage (30th day after sowing). Gradually, drought stress was initiated when the soil water content was 50%. Salinization and drought treatment began at 20:00, and after 2 days of salinity drought stress treatment, every day at 20:30, half of the seedlings were treated with 150 µmol·L−1 MT solution (Table 1). Melatonin was dissolved in an appropriate amount of absolute ethanol in the dark (anhydrous ethanol was added to melatonin continuously with constant stirring until melatonin was completely dissolved and then we stopped adding anhydrous ethanol to the melatonin) and then diluted with distilled water. Based on the results of a previous experiment (data not published), 150 µmol·L−1 MT can effectively alleviate drought and salinity stress. Spraying should be carried out in such a way that the mist vapor just beads up on the blade surface and drips down naturally. In the control group, distilled water was used instead of MT solution to perform the same operation. Applications were carried out for a total of 7 days. The stress conditions were set to medium intensity (drought: 50% soil water content, and salinity: 200 mmol·L−1 NaCl) because this level of stress was compared with each stressor alone, and the combined stress produced the same growth inhibition. Finally, photographs and measurements of related indicators were acquired.
2.2. Measurement Items and Methods
2.2.1. Determination of Plant Morphological Indicators and Biomass
For each treatment, 3 plants of essentially uniform and representative growth were selected. They were taken back to the laboratory and washed with clean water. Plant height, number of blades, and fresh weight (FW) were determined. The aboveground part of the plant was separated from the root system. An LI-3100C (LI-Cor, USA) digital blade area meter was used to measure the blade area of the plant. The roots were placed on A4 paper. After scanning, WinRHIZO (version 4.0b, Rengent Instruments Inc., Regent, Canada,) root analysis system software was used to analyze the average root diameter (ARD), total root length (RL), root surface area (RSA), root volume (RV), and other indicators. After the root structure was observed, the plant samples were placed in a 105 °C constant temperature air blast drying oven for 30 min and then dried to a constant weight at 80 °C. Finally, their dry weight (DW) was measured.
2.2.2. Indicators of Osmoregulation in Blade
The relative water content (RWC) of the blades was determined using the drying weighing method [34]. The proline (Pro) content was determined using the acid ninhydrin method [35]. The soluble sugar (SS) content was determined using the anthrone chromogenic method [36].
FW: blade fresh weight; TW: blade saturated water weight; DW: blade dry weight.
2.2.3. Antioxidant System Activity of Blade
The malondialdehyde (MDA) content was determined using the thiobarbituric acid method [35]. The relative electrical conductivity (REC) was measured using a DDS-HA conductivity meter [34]. The hydrogen peroxide (H2O2) content was determined using the potassium iodide method [35]. The superoxide anion (O2·−) content was determined following the hydroxylamine hydrochloride oxidation method [36]. The superoxide dismutase (SOD) activity was determined using the NBT photoreduction method, the peroxidase (POD) activity was determined following the guaiacol method, and the catalase (CAT) activity was determined by colorimetry [35].
S0: blank conductivity; S1: conductivity after immersion; S2: conductivity after boiling.
2.3. Statistical Analysis
Excel 2010 software was used to collate the data, and Origin 2021 was used to generate the figures. SPSS 27.0 data statistical software was used to perform significance analysis, analysis of variance, and correlation analysis, and the differences between treatments were compared using the LSD significance test method. Different letters in figures indicate significant differences at p < 0.05.
3. Results
3.1. Effects of Exogenous Melatonin on Morphological Indicators and Biomass in C. esculentus under Salt and Drought Stress
After seven days of treatment at the tillering stage, salt and drought stress significantly inhibited the growth of C. esculentus seedlings, and applying 150 μmol·L−1 MT solution to the blade significantly mitigated the inhibitory effects of the stresses on seedling growth (Table 2). Compared with the CK treatment, the plant height, number of blades, and DW of plants under CKM treatment showed no significant differences (p > 0.05), and the FW significantly increased by 12.77% (p < 0.05). Compared to the CK treatment, the plant height, number of blade, FW, and DW of plants under S treatments decreased by 22.23%, 34.56%, 34.79%, and 48.74%; under D treatments the parameters decreased by 24.23%, 33.33%, 36.72%, and 40.34%; under SD treatments the decreased by 25.16%, 29.63%, 43.95%, and 57.14%, with significant differences (p < 0.05); and there were no significant differences in plant height, number of blades, FW, or DW between the S and D treatments, and the FWs and DWs of the plants were significantly lower following the SD treatment than the S and D treatments, which indicated that salt, drought, and salt–drought stress all have significant inhibitory effects on plant growth, and the combined stresses exacerbated the inhibition of plant growth. The plant height, number of blades, FWs, and DWs of the plants under the different stress treatments significantly increased after foliar MT application. Compared with those of plants under S treatment, the plant height, number of blades, FWs, and DWs of the plants under the SM treatment increased by 14.92%, 39.62%, 24.23%, and 60.66%, respectively (p < 0.05). Compared with those of plants under the D treatment, the plant height, number of blades, FWs, and DWs of the plants under the DM treatment increased significantly by 15.19%, 55.56%, 44.10%, and 35.92%, respectively (p < 0.05), and the number of blades increased by 3.70% compared with that of the plants under the CK treatment, although the difference was not significant (p > 0.05). Compared with those of the plants under the SD treatment, the plant height, number of blades, FWs, and DWs of the plants under the SDM treatment were significantly increased by 14.67%, 14.05%, 30.88%, and 47.06% (p > 0.05); this increase was the lowest. There was no significant difference in the number of blades in the SM and DM groups compared to the CKM group, but the plant height, number of blades, FW, and DW were significantly lower in the SDM treatment group than in CKM group, and significantly lower than in the SM and DM groups, suggesting that melatonin was able to alleviate the stresses caused by salt, drought, and salt–drought stress on plant growth, but the alleviating effect was weakest on the growth of the plants affected by salt–drought stress.
The indexes of root morphogenesis also changed to different degrees after the stress treatments (Table 3). Salt and drought stress and their combination had no significant effect on the ARD of plants, and there was no significant difference in the ARD after exogenous application of MT under each stress treatment. The S, D, and SD treatments significantly reduced the RL, RSA, and RV in the root system of the plants compared to CK (p < 0.05), with SD showing the highest reductions of 45.37%, 52.16%, and 61.81%, suggesting that salt and drought stress significantly inhibited the growth of the plant root system, and the combined stress exacerbated the effects on plant root growth. Compared with CK, the RL, RSA, and RV following the CKM treatment were not significantly different. Compared with S, the RL, RSA, and RV following SM treatment increased by 18.37%, 42.12%, and 52.49%, with significant differences (p < 0.05). The RSA under SM treatment increased by 8.70% compared with that under the CK treatment, with significant differences (p < 0.05), and the RV increased by 7.82% compared with that following CK treatment. Compared with D treatment, the RL, RSA, and RV following DM treatment increased by 51.62%, 53.67%, and 54.80%, respectively, with a significant difference (p < 0.05). The RSA and RV increased by 1.76% and 1.38%, respectively, compared with under the CK treatment, but the difference was not significant (p > 0.05). Compared with the SD treatment, the RL, RSA, and RV of SDM increased significantly (p < 0.05) by 67.52%, 96.38%, and 128.51%, respectively. This shows that the application of exogenous melatonin significantly promoted root growth under salt, drought, and salt–drought stresses in C. esculentus: spraying the plants under salt stress with melatonin had the smallest effect on root growth, and spraying the plants with melatonin under salt–drought stress had the greatest effect on root growth.
3.2. Effects of Exogenous Melatonin on the Osmotic Regulation Index of Blades in C. esculentus under Salt and Drought Stress
Compared with the CK treatment, the RWC of the blades of C. esculentus seedlings under salt, drought, and salt–drought stress decreased by 14.39%, 15.37%, and 21.16%, respectively, and the difference was significant (p < 0.05) (Figure 1). There was no significant difference between the RWC of the plants following the S, D, and SD treatments, but the S treatment led to the smallest decrease and the SD treatment led to the largest decreases, suggesting that salt–drought stress exacerbates water loss from C. esculentus blades.
There was no significant difference in RWC between the CKM and CK treatments (p > 0.05). The RWC under the SM treatment increased by 12.26% compared with that under the S treatment, and the difference was significant (p < 0.05), and it recovered to 96.11% of that under CK treatment. Compared with the D treatment, the RWC under the DM treatment increased by 16.78% (p < 0.05), and it reached 98.83% of that under the CK treatment. The RWC of SDM treatment increased by 21.15% compared with that under the SD treatment, and the difference was significant (p < 0.05), which was 95.51% of that under the CK treatment. There was no significant difference in RWC among the plants that received the SM, DM, and SDM treatments, but the SDM treatment led to the highest increase in RWC, indicating that exogenous melatonin could significantly increase the RWC of C. esculentus blades under salt and drought stress, and it had the most significant effect during salt–drought stress.
Compared with the control, the Pro content increased by 18.78%, 26.45%, and 11.47%, and the SS content increased by 24.95%, 12.64%, and 16.63% under salt, drought, and salt–drought combined stress, respectively. The difference was significant (p < 0.05). Pro increased the most under the drought stress and least under the salt-drought stress; the amount of SS increased the most under salt stress and least under drought stress. This indicated that under salt stress the plant mainly resisted the stress by increasing the amount of Pro, and drought stress mainly resisted the stress by increasing the amount of SS, and the increase in the amount of SS in the C. esculentus blades was higher than the increase in the amount of Pro under salt–drought stress. The amounts of Pro and SS under the CKM treatment were not significantly different compared to those under the CK treatment (p > 0.05) (Figure 2). The amounts of Pro and SS increased to different degrees in all treatments after melatonin application, with the highest increase in Pro content being 16.96% under SDM treatment and the lowest increase being 11.88% under SM treatment, which was significant (p < 0.05); the highest increase in SS content was 36.21% under SM treatment and the lowest increase was 11.40% under SDM treatment, which was significantly different (p < 0.05). The greatest increase in Pro content was observed under the SDM treatment and the smallest increase in Pro content was observed under the SM treatment; the greatest increase in SS content was observed under the SM treatment and the smallest increase in SS content was observed under the SDM treatment. The results showed that melatonin had the most significant effect on Pro under salt–drought stress, and melatonin had the most significant effect on SS under salt stress.
3.3. Effects of Exogenous Melatonin on the Antioxidant System Activity of Blades of C. esculentus under Salt and Drought Stress
Compared with the CK treatment, the MDA content and REC under salt, drought, and salt–drought stress increased to a certain extent (Figure 3). Compared with the CK treatment, the MDA content and REC under S treatment significantly increased by 49.25% and 32.80%, respectively (p < 0.05). The MDA content and REC of the C. esculentus blades under the D treatment significantly increased by 91.35% and 25.08%, respectively (p < 0.05). The MDA content and REC of the blades under SD treatment significantly increased by 72.84% and 24.62%, respectively (p < 0.05). There was no significant difference in MDA content between the plants subjected to the D and SD treatmetns, and the MDA content following the S treatment was significantly lower than that following treatment with D and SD; there was no significant difference in REC between the S, D, and SD treatments, but S led to the highest increase. The REC and MDA content were not significantly different following the CKM treatment compared to the CK treatment (p > 0.05). Comparing the stresses, the MDA content and REC were most reduced following the DM treatment, by 55.02% and 17.71%, respectively (p < 0.05); and were least reduced following the SM treatment, by 26.05% and 10.97%, respectively (p < 0.05). It has been shown that the exogenous application of melatonin was able to significantly (p < 0.05) reduce the MDA content and the REC of blades of C. esculentus under stress conditions, and had the strongest regulatory effect on C. esculentus’ MDA and RWC under drought stress.
The H2O2 and O2·− contents in the blade of C. esculentus were increased to a certain extent under salt, drought, and salt–drought combined stress (Figure 4). Compared with the CK treatment, the H2O2 and O2·− contents significantly increased by 135.38% and 68.40%, respectively, under the S treatment, by 153.22% and 82.14%, respectively, under the D treatment, and by 109.92% and 56.91%, respectively, under the SD treatment (p < 0.05). Both H2O2 and O2·− were significantly higher following the D treatment than the S and SD treatments, and there was no significant difference between H2O2 and S, and O2·− was significantly lower than S following the SD treatment, indicating that the highest accumulation of ROS was observed in C. esculentus under drought stress, whereas the addition of salt stress to drought stress reduced the accumulation of ROS. The difference in H2O2 content and O2·− content under CKM treatment was not significant (p > 0.05) compared to that under the CK treatment. Compared to no melatonin application, both the H2O2 and O2·− contents were reduced the most under SDM treatment, by 24.02% and 58.14%, and the difference was significant (p < 0.05), while the H2O2 content was reduced the least under DM treatment, by 16.25% (p < 0.05), and the O2·− content was reduced the least under SM treatment, 54.94% (p < 0.05). It has been shown that 150 µmol·L−1 of melatonin reduced the accumulation of ROS in C. esculentus under salt, drought, and salt–drought stress, and H2O2 and O2·− responded most strongly to melatonin under salt-drought stress.
In Figure 5, compared to the CK treatment, the POD activity increased the most and the CAT activity increased the least in the C. esculentus blades following the S treatment, which significantly increased by 34.93% and 25.73% (p < 0.05), and the SOD activity significantly increased by 23.19% (p < 0.05); the POD activity increased the most and the SOD activity the least following the D treatment, which significantly increased by 59.33% and 11.59% (p < 0.05), and the CAT activity significantly increased by 23.49% (p < 0.05); and the SOD and CAT activities were significantly increased by 15.28% and 10.93% (p < 0.05) following the SD treatment. There was no significant difference in SOD activity among the S, D, and SD treatments; all POD activities were significantly different, showing that D > S > SD; and the difference in CAT activity showed that the effects of S and D were significantly higher than those of SD, and there was no significant differences between the S and D treatments, indicating that POD was more sensitive to drought stress, and CAT was sensitive to salt and drought stress. The level of difference in SOD, POD, and CAT activities during the CKM treatment compared to the CK treatment was not significant (p > 0.05). When comparing the types of stress, both the SM and DM treatments led to the most significant increases in SOD activity, 30.59% and 63.64%, and the least significant increases in CAT activity, 15.24% and 6.21%, with significant differences (p < 0.05); the activities of SOD, POD, and CAT under the SDM treatment were increased by 20.00%, 30.00%, and 16.38%, with significant differences (p < 0.05). It has been shown that 150 μmol·L−1 of exogenous melatonin significantly increased the SOD, POD, and CAT activities of C. esculentus under stress conditions, indicating that salt and drought stresses scavenge stress-generated ROS mainly by increasing SOD activity, and salt-drought stress scavenges stress-generated ROS mainly by increasing POD activity.
3.4. Evaluation of Stress Resistance of Cyperus esculentus Seedlings
3.4.1. Correlation Analysis
To explore the relationship between the growth of C. esculentus seedlings and the physiological and metabolic indexes of C. esculentus and to evaluate the resistance of C. esculentus seedlings to salt and drought stress under MT treatment, a Pearson correlation analysis was carried out among various traits (Figure 6). The correlation between the morphological indexes of C. esculentus reached a significant level (p < 0.05). The correlation between each morphological index and the RWC reached a significant level (p < 0.001), and the correlation between POD, CAT, and H2O2 reached a significant level (p < 0.001). The correlation with O2·− reached a significant level (p < 0.01). Between the indicators, a certain correlation was observed in the relevant data matrix; thus, the growth effects overlapped. Therefore, the evaluation of the sustained release effect of MT should be combined with multiple indicators for reliability.
3.4.2. Principal Component Analysis
To better analyze the relationships between the growth of C. esculentus and the physiological and metabolic indexes of C. esculentus, a principal component analysis was performed on the parameters, and two principal components with eigenvalues > 1 were extracted. The eigenvalues were 10.961 and 5.068. The proportion of variance was 60.897% and 28.156%, and the cumulative variance contribution rates reached 60.897% and 89.053%, which met the analysis requirements. As shown in Figure 7, plant height, number of blades, FW, DW, RL, RSA, RV, ARD, and RWC played a major role in the first principal component. The number of blades, RL, RSA, RV, ARD, RWC, Pro, SS, MDA, REC, SOD, CAT, and H2O2 were the main factors affecting the second principal component. The treatments were ranked by calculating the comprehensive score (F), where F = F1 × 60.897% + F2 × 28.156%. As shown in Table 4, the comprehensive scores of the effects MT on morphological and physiological metabolism indexes under salt and drought stress were 0.08, −0.19, −0.23, −0.34, 0.11, 0.21, 0.27, and 0.09, and the ranking among the treatments was as follows: DM > SM > CKM > SDM > CK > S > D > SD. According to the ranking, it can be seen that the composite scores of the treatments with exogenous melatonin were all higher than those without melatonin; this suggests that the treatments with exogenously applied were superior to the treatments without the application of melatonin when evaluated by all indicators combined. According to the ranking, MT application had a significant mitigating effect on the growth and physiological metabolism of C. esculentus under salt and drought stress.
4. Discussion
In this study, the effects of salinity, drought stress, and their combined stress on plant morphology, osmotic adjustment substances, and the activity of the antioxidant system of C. esculentus and the alleviating effect of exogenous MT on stress were investigated. Adding salt stress to drought stress exacerbates the inhibitory effects of stress on plant growth. The application of exogenous MT alleviated the growth inhibition of plants caused by salt, drought, and their combined stress by regulating osmotic balance, increasing antioxidant enzyme activity, and reducing the reactive oxygen species content in plants.
The most intuitive manifestation of the effects of salt and drought stress on plants is that growth and development are significantly inhibited [16]. The results of this study showed that drought, salt, and salt–drought combined stress had a significant inhibitory effect on the growth of C. esculentus seedlings and root growth and reduced the biomass accumulation of C. esculentus seedlings, and the combined stress led to a higher inhibition of the growth of C. esculentus than the single stresses of salt and drought. Foliar spraying with 150 μmol·L−1 MT solution effectively reduced the severity of salt and drought stress on the growth inhibition of C. esculentus seedlings, which was mainly manifested in the increase in the plant height, number of blades, FW, and DW of seedlings to varying degrees (Table 2). The results of this study are similar to those of previous studies on canola [37], maize [38], cotton [39], and strawberries [40]. The RL, RSA, RV, and ARD of plants have an important regulatory effect on the growth and development of the aboveground part [41]. In this study, compared with the control, salt, drought, and salt–drought combined stress significantly reduced the RL, RSA, and RV of C. esculentus seedlings, while exogenous MT application increased the RL, RSA, and RV of the seedlings (Table 3). This is similar to Zhu et al.’s finding in oilseed rape that stress promotes root growth through the uptake of exogenous MT [42]. The above results showed that MT application enhanced the salt tolerance and drought resistance of C. esculentus under field conditions.
For the sake of coping with physiological drought caused by abiotic stress, osmotic adjustment, a physiological mechanism to promote stress adaptation, is produced in plants. It has been shown that proline can reduce the accumulation of reactive oxygen species in plants and enhance their antioxidant enzyme activity, thus reducing the oxidative damage caused by the accumulation of reactive oxygen species in plants, etc. [43]. Therefore, proline accumulation usually enhances plants’ tolerance to osmotic stress [44]. Soluble sugar is an important nonstructural carbohydrate in plants that contributes to plant adaptation to adversity stresses [45]. As an important osmotic adjustment substance in plants, its accumulation in plants reduces the cell water potential and enhances the water-holding capacity of cells, thereby reducing the damage caused by physiological drought to plants [46]. Therefore, the content of these substances in plant cells can be used as a standard to judge the adaptations of plants that allow them to resist abiotic stress. Previous research results have shown that, under salt and drought stress conditions, the large accumulation of Pro and SS in plants increases the concentration of plant cell sap, maintains the normal turgor of plant cells, and prevents plants from experiencing excessive water loss due to physiological drought caused by an adverse external environment, thus enhancing the plant’s resistance to adversity [47]. RWC is considered a suitable indicator of blade water status, which is reduced under water stress and causes changes to cell membranes [48]. In this study, the RWC of the blades of C. esculentus seedlings under drought, salt, and salt–drought stress was significantly reduced, and the Pro and SS contents of C. esculentus seedlings under salt and drought stress were significantly increased in both. The RWC, Pro, and SS contents of C. esculentus under salt and drought stress were lower than those under salt and drought stress (Figure 1 and Figure 2), indicating that salt-drought stress had a lesser effect on the osmoregulation of C. esculentus than a single stress, simultaneous salt and drought treatments reduced the accumulation of osmoregulatory substances in C. esculentus. The osmoregulatory substances in A could be reduced by the simultaneous treatment of salt and drought. After spraying exogenous MT on the blades of C. esculentus seedlings, the RWC and SS and Pro contents significantly increased (Figure 1 and Figure 2). The increase in Pro and SS contents was helpful in reducing the cell osmotic potential, maintaining intracellular turgor, and reducing the damage caused by physiological drought caused by external abiotic stress. The increase in the RWC of the blades further confirmed the results of this study, which are similar to the conclusions drawn by Zhao et al. in their research on maize [49]. In summary, exogenous MT can maintain the cell osmotic balance by regulating the cells’ osmotic potential, thereby maintaining the stability of the osmotic potential inside and outside the cell membrane and improving the salt tolerance and drought resistance of C. esculentus seedlings.
Abiotic stress leads to oxidative stress in plants, with a high accumulation of active oxygen, resulting in lipid peroxidation, antioxidant enzyme activity is activated, and cell membrane integrity loss in plants. Plants respond to stress conditions by activating antioxidants to inhibit the ROS levels in plant cells [50]. As one of the products of membrane lipid peroxidation, MDA is usually used to judge the stability of the cell membrane and the degree of oxidative damage [8]. REC is often considered to be a good index for reflecting the permeability of the cell membrane and the integrity of the cell membrane under stress; therefore, it is easily affected by environmental pressure [51]. With the increase in drought and salt stress, the increase in lipid peroxidation in the plant blades is related to ROS production, including H2O2 and O2·− [52]. It has been reported that exogenous MT can reduce the amount of ROS produced during salt stress [53], drought stress [49,54], and senescence [55]. Improving the plant’s antioxidant capacity has always been considered the main role of MT in plant stress resistance [53]. As an antioxidant and free radical scavenger, MT can remove ROS from crops by activating antioxidant enzyme activity in plants, effectively reducing H2O2 and O2·− accumulation in blades and roots, thereby protecting membrane lipids from stress and reducing MDA levels [56]. In this study, salt and drought stress rapidly increased the MDA, H2O2, and O2·− contents in C. esculentus seedlings, and the REC was significantly higher (Figure 3 and Figure 4), thus affecting the normal growth and development of seedlings. In this study, the effect of drought stress alone on cell membrane lipid peroxidation was higher than that of salt stress alone, and the effect of combined stress was the smallest. It is speculated that the removal of active oxygen metabolism in seedlings under combined stress may be activated. However, exogenous MT application significantly reduced the ROS accumulation caused by abiotic stress and reduced the REC of the cells in the plants (Figure 3 and Figure 4). Since a decrease in ROS content can reduce the oxidative damage of the blade cell membrane of C. esculentus seedlings under MT treatment, it is speculated that the MDA content in seedlings under abiotic stress is lower than that of C. esculentus seedlings without MT treatment. To verify this, we measured the MDA content in the seedlings. The results showed that, under stress conditions, the MDA content in MT-treated seedlings was significantly lower than that in untreated seedlings (Figure 3). In summary, these findings indicate that MT can alleviate membrane damage caused by excessive ROS accumulation in plants.
To resist the oxidative stress induced by an adverse environment, a relatively complete antioxidant enzyme system has evolved in plants; this is very important for the improvement of plant tolerance to abiotic stress [57]. Antioxidant enzymes play an essential role in scavenging superoxide ions, resisting lipid peroxidation, and reducing membrane damage [47]. Studies have shown that ROS production increases the activity of antioxidant enzymes in plants under oxidative stress [58,59]. It has been confirmed that MT can enhance the activity of antioxidant enzymes in plants under abiotic stress [55,60]. The results of this study showed that the SOD activity in plants under a single stressor was lower than that under combined stress, but the POD and CAT activities in plants under a single stressor were higher than those under combined stress. This may be due to the activation of scavenging reactive oxygen species metabolism in seedlings under combined stress; therefore, ROS accumulation in a certain range did not exceed the threshold of abiotic stress-induced damage. Therefore, the SOD activity produced under combined stress was higher than that produced under a single stressor, but the sensitivities of POD and CAT to stress were higher than that of SOD. When the stress level was too high and the reactive oxygen species content in the plants exceeded the tolerance range of the plants, the activity of the protective enzymes began to decrease. After exogenous MT application, the antioxidant enzyme activity in plants increased significantly, and the activity of antioxidant enzymes under a single stressor was higher than that under combined stress (Figure 5).
The results confirmed that a decrease in membrane lipid peroxidation in seedlings under abiotic stress can be achieved by applying MT to enhance the activity of antioxidant enzymes so that cells can maintain normal metabolism and alleviate the effects of abiotic stress on plants. The above results show that MT plays an important role in protecting plants from adverse stress conditions and in preventing oxidative stress-induced damage at the cellular level [61].
The effect of melatonin on plant growth and their metabolic physiology under salt, drought, and combined salt and drought stress is not due to a single factor; it is related to several factors with varying degrees of influence. Therefore, correlation analysis and principal component analysis of multiple growth and metabolic-physiological indicators are particularly important. In this experiment, the correlation between all of the morphological indicators of C. esculentus plants was found to be at highly significant level, and a highly significant positive correlation was found between morphological indicators and RWC, suggesting that better plant growth is associated with an increase in RWC; there was a very significant negative correlation with POD, CAT, H2O2, and O2·−, which indicates that better plant growth in terms of antioxidant enzymes is associated with reduced POD and CAT activities, in terms of reactive oxygen species it is mainly related to a reduction in H2O2 and O2·− content; therefore, the above relevant indexes can be used to evaluate the effect of melatonin on salt and drought tolerance in C. esculentus. The first principal component in the principal component analysis synthesized information from the indicators of plant height, number of blades, FW, DW, RL, RSA, RV, ARD, and RWC. The second principal component synthesized the information of the indicators of Pro, SS, MDA, REC, SOD, POD, CAT, H2O2, and O2·−. Among them, plant height, number of blades, FW, DW, RL, RSA, RV, and ARD are important indicators reflecting the plant growth status, and RWC is an important indicator reflecting the degree of water deficit in the crop, which suggests that C. esculentus is mainly able to resist salt and drought stress by increasing the water content in its blades. In the second principal component, Pro, SS, and REC were shown to be osmoregulation indicators, MDA, H2O2, and O2·− were indicators of reactive oxygen content, and SOD, POD, and CAT were indicators of antioxidant enzymes, indicating that melatonin can increase osmoregulation substances, improve antioxidant enzyme activities, and reduce reactive oxygen content in C. esculentus, thereby resisting salt and drought stresses. In addition, from the results, it can be seen that melatonin application was able to mitigate the effects of salt, drought, and salt-drought stresses on the growth and physiological metabolism caused by C. esculentus.
5. Conclusions
In summary, salt, drought, and salt–drought stress all inhibited the plant growth of C. esculentus, decreased RWC, increased REC, increased the content of Pro, SS, and MDA, H2O2, O2·−, and increased the activities of SOD, POD, and CAT, indicating that the osmoregulatory substances in C. esculentus increased under the stress conditions, the cellular ROS content increased, and antioxidant enzyme activities increased, and the effects of the combined stresses on C. esculentus growth and physiological metabolism were higher than those of a single type of stress. The application of exogenous melatonin under salt stress, drought stress, and salt–drought stress can promote plant growth, increase RWC, osmoregulatory substance content, and antioxidant enzyme activity, and reduce the electrolyte extravasation rate and ROS content; therefore, the application of exogenous melatonin can alleviate the effects of salt stress, drought stress, and salt–drought stress on C. esculentus.
Author Contributions
Conceptualization, X.L., Z.W. and J.L.; methodology, N.W.; validation, N.W., X.L., Z.W. and J.L.; formal analysis, N.W.; investigation, N.W.; data curation, N.W., X.L., Z.W. and J.L.; writing—original draft preparation, N.W.; writing—review and editing, X.L., Z.W. and J.L.; visualization, N.W.; funding acquisition, J.L. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by National Key Research and Development Program of China grant number (2016YFC0501406).
Data Availability Statement
Data available on request.
Acknowledgments
Thank you to Jianguo Liu from Shihezi University for contributing to this experiment.
Conflicts of Interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
References
- Gupta, A.; Rico-Medina, A.; Caño-Delgado, A.I. The physiology of plant responses to drought. Science 2020, 368, 266–269. [Google Scholar] [CrossRef] [PubMed]
- Zelm, E.V.; Zhang, Y.; Testerink, C. Salt tolerance mechanisms of plants. Annu. Rev. Plant Biol. 2020, 71, 403–433. [Google Scholar] [CrossRef] [PubMed]
- Wicke, B.; Smeets, E.; Dornburg, V.; Vashev, B.; Gaiser, T.; Turkenburg, W.; Faaij, A. Correction: The global technical and economic potential of bioenergy from salt-affected soils. Energy Environ. Sci. 2020, 13, 2585. [Google Scholar] [CrossRef]
- Neupane, D.; Adhikari, P.; Bhattarai, D.; Rana, B.; Ahmed, Z.; Sharma, U.; Adhikari, D. Does climate change affect the yield of the top three cereals and food security in the world? Earth 2022, 3, 45–71. [Google Scholar] [CrossRef]
- Paul, K.; Pauk, J.; Kondic-Spika, A.; Grausgruber, H.; Allahverdiyev, T.; Sass, L.; Vass, I. Co-occurrence of mild salinity and drought synergistically enhances biomass and grain retardation in wheat. Front. Plant Sci. 2019, 10, 10501. [Google Scholar] [CrossRef]
- Li, P.C.; Yang, X.Y.; Wang, H.M.; Pan, T.; Yang, J.Y.; Wang, Y.Y.; Xu, Y.; Yang, Z.F.; Xu, C.W. Metabolic responses to combined water deficit and salt stress in maize primary roots. J. Integr. Agric. 2021, 20, 109–119. [Google Scholar] [CrossRef]
- Shamloo-Dashtpagerdi, R.; Aliakbari, M.; Lindlöf, A.; Tahmasebi, S. A systems biology study unveils the association between a melatonin biosynthesis gene, O-methyl transferase 1 (OMT1) and wheat (Triticum aestivum L.) combined drought and salinity stress tolerance. Planta 2022, 255, 99. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Cheng, K.; Ma, B.; Zhang, W.; Zheng, L.; Wang, Y. CaCl2 promotes the cross adaptation of Reaumuria trigyna to salt and drought by regulating Na+, ROS accumulation and programmed cell death. Plant Physiol. Biochem. 2023, 195, 214–227. [Google Scholar] [CrossRef] [PubMed]
- Abdelraheem, A.; Esmaeili, N.; O’Connell, M.; Zhang, J. Progress and perspective on drought and salt stress tolerance in cotton. Ind. Crops Prod. 2019, 130, 118–129. [Google Scholar] [CrossRef]
- Duan, W.J.; Lu, B.; Liu, L.T.; Meng, Y.J.; Ma, X.Y.; Li, J.; Zhang, K.; Sun, H.C.; Zhang, Y.J.; Dong, H.Z.; et al. Effects of Exogenous Melatonin on Root Physiology, Transcriptome and Metabolome of Cotton Seedlings under Salt Stress. Int. J. Mol. Sci. 2022, 23, 9456. [Google Scholar] [CrossRef]
- de Queiroz, G.C.M.; de Medeiros, J.F.; da Silva, R.R.; da Silva Morais, F.M.; de Sousa, L.V.; de Souza, M.V.P.; da Nóbrega Santos, E.; Ferreira, F.N.; da Silva, J.M.C.; Clemente, M.I.; et al. Growth, Solute Accumulation, and Ion Distribution in Sweet Sorghum under Salt and Drought Stresses in a Brazilian Potiguar Semiarid Area. Agriculture 2023, 13, 803. [Google Scholar] [CrossRef]
- Yang, Z.; Li, J.L.; Liu, L.N.; Xie, Q.; Sui, N. Photosynthetic regulation under salt stress and salt-tolerance mechanism of sweet sorghum. Front. Plant Sci. 2020, 10, 101722. [Google Scholar] [CrossRef] [PubMed]
- Khan, Z.; Jan, R.; Asif, S.; Farooq, M.; Jang, Y.H.; Kim, E.G.; Kim, N.; Kim, K.M. Exogenous melatonin induces salt and drought stress tolerance in rice by promoting plant growth and defense system. Sci. Rep. 2024, 14, 1214. [Google Scholar] [CrossRef] [PubMed]
- Gupta, A.; Bano, A.; Rai, S.; Mishra, R.; Singh, M.; Sharma, S.; Pathak, N. Mechanistic insights of plant-microbe interaction towards drought and salinity stress in plants for enhancing the agriculture productivity. Plant Stress 2022, 4, 100073. [Google Scholar] [CrossRef]
- Yolcu, S.; Alavilli, H.; Ganesh, P.; Panigrahy, M.; Song, K. Salt and Drought Stress Responses in Cultivated Beets (Beta vulgaris L.) and Wild Beet (Beta maritima L.). Plants 2021, 10, 1843. [Google Scholar] [CrossRef] [PubMed]
- Kaura, V.; Malhotra, P.K.; Mittal, A.; Sanghera, G.S.; Kaur, N.; Bhardwaj, R.D.; Cheema, R.S.; Kaur, G. Physiological, biochemical, and gene expression responses of sugarcane under cold, drought and salt stresses. J. Plant Growth Regul. 2023, 42, 6367–6376. [Google Scholar] [CrossRef]
- Wang, M.; Zhang, S.; Ding, F. Melatonin mitigates chilling-induced oxidative stress and photosynthesis inhibition in tomato plants. Antioxidants 2020, 9, 218. [Google Scholar] [CrossRef]
- Sun, C.; Liu, L.; Wang, L.; Li, B.; Jin, C.; Lin, X. Melatonin: A master regulator of plant development and stress responses. J. Integr. Plant Biol. 2021, 63, 126–145. [Google Scholar] [CrossRef]
- Arnao, M.B.; Hernández-Ruiz, J. Melatonin: Plant growth regulator and/or biostimulator during stress? Trends Plant Sci. 2014, 19, 789–797. [Google Scholar] [CrossRef]
- Zhang, N.; Sun, Q.; Zhang, H.; Cao, Y.; Weeda, S.; Ren, S.; Guo, Y.D. Roles of melatonin in abiotic stress resistance in plants. J. Exp. Bot. 2015, 66, 647–656. [Google Scholar] [CrossRef]
- Gao, W.; Feng, Z.; Bai, Q.; He, J.; Wang, Y. Melatonin-mediated regulation of growth and antioxidant capacity in salt-tolerant naked oat under salt stress. Int. J. Mol. Sci. 2019, 20, 1176. [Google Scholar] [CrossRef] [PubMed]
- Arnao, M.; Ruiz, J.H. Melatonin and reactive oxygen and nitrogen species: A model for the plant redox network. Melatonin Res. 2019, 2, 152–168. [Google Scholar] [CrossRef]
- Altaf, M.A.; Shahid, R.; Ren, M.X.; Mora-Poblete, F.; Arnao, M.B.; Naz, S.; Anwar, M.; Altaf, M.M.; Shahid, S.; Shakoor, A.; et al. Phytomelatonin: An overview of the importance and mediating functions of melatonin against environmental stresses. Physiol. Plant. 2021, 172, 820–846. [Google Scholar] [CrossRef] [PubMed]
- Moustafa-Farag, M.; Elkelish, A.; Dafe, M.; Khan, M.; Arnao, M.B.; Abdelhamid, M.T.; El-Ezz, A.A.; Almoneafy, A.; Mahmoud, A.; Awad, M.; et al. Role of melatonin in plant tolerance to soil stressors: Salinity, pH and heavy metals. Molecules 2020, 25, 5359. [Google Scholar] [CrossRef]
- Muhammad, I.; Yang, L.; Ahmad, S.; Farooq, S.; Khan, A.; Muhammad, N.; Ullah, S.; Adnan, M.; Ali, S.; Zhou, X.B.; et al. Melatonin-priming enhances maize seedling drought tolerance by regulating the antioxidant defense system. Plant Physiol. 2023, 191, 2301–2315. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Li, R.; Ge, J.; Liu, J.; Wang, W.; Xu, M.; Zhang, R.; Hussain, S.; Wei, H.; Dai, Q. Exogenous melatonin confers enhanced salinity tolerance in rice by blocking the ROS burst and improving Na+/K+ homeostasis. Environ. Exp. Bot. 2021, 189, 104530. [Google Scholar] [CrossRef]
- Zhang, L.; Fang, X.; Yu, N.; Chen, J.; Wang, H.; Shen, Q.; Chen, G.; Wang, Y. Melatonin Promotes Rice Seed Germination under Drought Stress by Regulating Antioxidant Capacity. Phyton 2023, 92, 1571–1587. [Google Scholar] [CrossRef]
- Guo, T.; Wan, C.; Huang, F.; Wei, C. Evaluation of quality properties and antioxidant activities of tiger nut (Cyperus esculentus L.) oil produced by mechanical expression or/with critical fluid extraction. LWT-Food Sci. Technol. 2021, 141, 110915. [Google Scholar] [CrossRef]
- Adebayo, S.F.; Arinola, S.O. Effect of germination on the nutrient and antioxidant properties of tigernut (Cyperus esculentus). J. Biol. Agric. Healthc. 2017, 7, 88–94. [Google Scholar] [CrossRef]
- Sánchez-Zapata, E.; Fernández-López, J.; Angel Pérez-Alvarez, J. Tiger nut (Cyperus esculentus) commercialization: Health aspects, composition, properties, and food applications. Compr. Rev. Food Sci. Food Saf. 2012, 11, 366–377. [Google Scholar] [CrossRef]
- Follak, S.; Belz, R.; Bohren, C.; De Castro, O.; Del Guacchio, E.; Pascual-Seva, N.; Schwarz, M.; Verloove, F.; Essl, F. Biological flora of central Europe: Cyperus esculentus L. Perspect. Plant Ecol. Evol. Syst. 2016, 23, 33–51. [Google Scholar] [CrossRef]
- Yu, Y.; Lu, X.; Zhang, T.; Zhao, C.; Guan, S.; Pu, Y.; Gao, F. Tiger nut (Cyperus esculentus L.): Nutrition, processing, function and applications. Foods 2022, 11, 601. [Google Scholar] [CrossRef] [PubMed]
- Qu, P.; Cheng, Z.; Long, C.; Su, M.; Yang, D. Comprehensive development of chufa (Cyperus esculentus L. var. sativus). China Oils Fats 2007, 09, 61–63. [Google Scholar] [CrossRef]
- Ying, Y.Q.; Song, L.L.; Jacobs, D.F.; Mei, L.; Liu, P.; Jin, S.H.; Wu, J.S. Physiological response to drought stress in Camptotheca acuminata seedlings from two provenances. Front. Plant Sci. 2015, 6, 124077. [Google Scholar] [CrossRef] [PubMed]
- Ozden, M.; Demirel, U.; Kahraman, A. Effects of proline on antioxidant system in leaves of grapevine (Vitis vinifera L.) exposed to oxidative stress by H2O2. Sci. Hortic. 2009, 119, 163–168. [Google Scholar] [CrossRef]
- Elstner, E.F.; Heupel, A. Inhibition of nitrite formation from hydroxylammoniumchloride: A simple assay for superoxide dismutase. Anal. Biochem. 1976, 70, 616–620. [Google Scholar] [CrossRef] [PubMed]
- Javeed, H.M.R.; Ali, M.; Skalicky, M.; Nawaz, F.; Qamar, R.; Rehman, A.U.; Faheem, M.; Mubeen, M.; Iqbal, M.M.; ur Ruhman, M.H.; et al. Lipoic acid combined with melatonin mitigates oxidative stress and promotes root formation and growth in salt-stressed canola seedlings (Brassica napus L.). Molecules 2021, 26, 3147. [Google Scholar] [CrossRef]
- Huang, B.; Chen, Y.E.; Zhao, Y.Q.; Ding, C.B.; Liao, J.Q.; Hu, C.; Zhou, L.J.; Zhang, Z.W.; Yuan, S.; Yuan, M. Exogenous melatonin alleviates oxidative damages and protects photosystem II in maize seedlings under drought stress. Front. Plant Sci. 2019, 10, 677. [Google Scholar] [CrossRef] [PubMed]
- Zhu, L.; Li, A.; Sun, H.; Li, P.; Liu, X.; Guo, C.; Zhang, Y.; Zhang, K.; Bai, Z.; Dong, H. The effect of exogenous melatonin on root growth and lifespan and seed cotton yield under drought stress. Ind. Crops Prod. 2023, 204, 117344. [Google Scholar] [CrossRef]
- Wu, S.; Wang, Y.; Zhang, J.; Guo, X.; Zhang, Z.; Sun, J.; Chen, X.; Wang, Y. Exogenous melatonin improves physiological characteristics and promotes growth of strawberry seedlings under cadmium stress. Hortic. Plant J. 2021, 7, 13–22. [Google Scholar] [CrossRef]
- Zhang, H.; Zhang, X.; Gao, G.; Ali, I.; Wu, X.; Tang, M.; Chen, L.; Jiang, L.; Liang, T. Effects of various seed priming on morphological, physiological, and biochemical traits of rice under chilling stress. Front. Plant Sci. 2023, 14, 1146285. [Google Scholar] [CrossRef]
- Zhu, Z.H.; Sami, A.; Xu, Q.Q.; Wu, L.L.; Zheng, W.Y.; Chen, Z.P.; Jin, X.Z.; Zhang, H.; Li, Y.; Yu, Y.; et al. Effects of seed priming treatments on the germination and development of two rapeseed (Brassica napus L.) varieties under the co-influence of low temperature and drought. PLoS ONE 2012, 16, e0257236. [Google Scholar] [CrossRef] [PubMed]
- Smirnof, N. The role of active oxygen in the response of plants towaterd eficit and desiccation. New Phytol. 1993, 125, 27–58. [Google Scholar] [CrossRef]
- Hanif, S.; Saleem, M.F.; Sarwar, M.; Irshad, M.; Shakoor, A.; Wahid, M.A.; Khan, H.Z. Biochemically triggered heat and drought stress tolerance in rice by proline application. J. Plant Growth Regul. 2021, 40, 305–312. [Google Scholar] [CrossRef]
- Piper, F.I.; Paula, S. The role of nonstructural carbohydrates storage in forest resilience under climate change. Curr. For. Rep. 2020, 6, 1–13. [Google Scholar] [CrossRef]
- Cao, X.; Shen, Q.; Ma, S.; Liu, L.; Cheng, J. Physiological andpiptranscriptional responses to progressive soil water deficit in three mulberry cultivars. Front. Plant Sci. 2020, 11, 1310. [Google Scholar] [CrossRef] [PubMed]
- Dugasa, M.T.; Cao, F.; Ibrahim, W.; Wu, F. Differences in physiological and biochemical characteristics in response to single and combined drought and salinity stresses between wheat genotypes differing in salt tolerance. Physiol. Plant. 2019, 165, 134–143. [Google Scholar] [CrossRef] [PubMed]
- Alagoz, S.M.; Hadi, H.; Toorchi, M.; Pawowski, T.A.; Lajayer, B.A.; Price, G.W.; Farooq, M.; Astatkie, T. Morpho-physiological responses and growth indices of triticale to drought and salt stresses. Sci. Rep. 2023, 13, 8896. [Google Scholar] [CrossRef]
- Zhao, C.; Yang, M.; Wu, X.; Wang, Y.; Zhang, R. Physiological and transcriptomic analyses of the effects of exogenous melatonin on drought tolerance in maize (Zea mays L.). Plant Physiol. Biochem. 2021, 168, 128–142. [Google Scholar] [CrossRef]
- Gökçe, A.F.; Gökçe ZN, Ö.; Junaid, M.D.; Chaudhry, U.K. Evaluation of biochemical and molecular response of onion breeding lines to drought and salt stresses. Sci. Hortic. 2023, 311, 111802. [Google Scholar] [CrossRef]
- Zhang, W.; Bao, G.; Tang, W.; Dai, G.; Xiao, J.; Liu, J.; Wang, Z.; Xi, J. Physiological response of barley seedlings to salinity and artemisinin combined stresses under freeze-thaw environment. Environ. Sci. Pollut. Res. Int. 2022, 29, 70552–70563. [Google Scholar] [CrossRef] [PubMed]
- Afzal, M.Z.; Jia, Q.; Ibrahim, A.K.; Niyitanga, S.; Zhang, L. Mechanisms and signaling pathways of salt tolerance in crops: Understanding from the transgenic plants. Trop. Plant Biol. 2020, 13, 297–320. [Google Scholar] [CrossRef]
- Zhan, H.; Nie, X.; Zhang, T.; Li, S.; Wang, X.; Du, X.; Tong, W.; Song, W. Melatonin: A small molecule but important for salt stress tolerance in plants. Int. J. Mol. Sci. 2019, 20, 709. [Google Scholar] [CrossRef] [PubMed]
- EL-Bauome, H.A.; Abdeldaym, E.A.; Abd El-Hady, M.A.; Darwish, D.B.E.; Alsubeie, M.S.; El-Mogy, M.M.; Basahi, M.A.; Al-Qahtani, S.M.; Al-Harbi, N.A.; Alzuaibr, F.M.; et al. Exogenous proline, methionine, and melatonin stimulate growth, quality, and drought tolerance in cauliflower plants. Agriculture 2022, 12, 1301. [Google Scholar] [CrossRef]
- Debnath, B.; Islam, W.; Li, M.; Sun, Y.; Lu, X.; Mitra, S.; Hussain, M.; Liu, S.; Qiu, D. Melatonin mediates enhancement of stress tolerance in plants. Int. J. Mol. Sci. 2019, 20, 1040. [Google Scholar] [CrossRef] [PubMed]
- Sun, C.; Gao, L.; Xu, L.; Zheng, Q.; Sun, S.; Liu, X.; Zhang, Z.; Tian, Z.; Dai, T.; Sun, J. Melatonin alleviates chromium toxicity by altering chromium subcellular distribution and enhancing antioxidant metabolism in wheat seedlings. Environ. Sci. Pollut. Res. Int. 2023, 30, 50743–50758. [Google Scholar] [CrossRef]
- Rajput, V.D.; Harish; Singh, R.K.; Verma, K.K.; Sharma, L.; Quiroz-Figueroa, F.R.; Meena, M.; Gour, V.S.; Minkina, T.; Sushkova, S.; et al. Recent developments in enzymatic antioxidant defence mechanism in plants with special reference to abiotic stress. Biology 2021, 10, 267. [Google Scholar] [CrossRef]
- Zhang, H.H.; Xu, N.; Li, X.; Han, Y.; Ren, J.W.; Li, M.B. Special issue in honour of Prof. Reto J. Strasser–Effects of exogenous abscisic acid on the photosynthetic function and reactive oxygen species metabolism of tobacco leaves under drought stress. Photosynthetica 2020, 58, 585–594. [Google Scholar] [CrossRef]
- Zhang, Z.; Cao, B.; Gao, S.; Xu, K. Grafting improves tomato drought tolerance through enhancing photosynthetic capacity and reducing ROS accumulation. Protoplasma 2019, 256, 1013–1024. [Google Scholar] [CrossRef]
- Li, J.; Liu, J.; Zhu, T.; Zhao, C.; Li, L.; Chen, M. The role of melatonin in salt stress responses. Int. J. Mol. Sci. 2019, 20, 1735. [Google Scholar] [CrossRef]
- Khan MS, S.; Ahmed, S.; ul Ikram, A.; Hannan, F.; Yasin, M.U.; Wang, J.; Zhao, B.; Islam, F.; Chen, J. Phytomelatonin: A key regulator of redox and phytohormones signaling against biotic/abiotic stresses. Redox Biol. 2023, 102805. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Effects of exogenous melatonin on blade relative water content of Cyperus esculentus seedlings under salt and drought stress. Note: Different lowercase letters indicate significant differences between treatments at p < 0.05.
Figure 1.
Effects of exogenous melatonin on blade relative water content of Cyperus esculentus seedlings under salt and drought stress. Note: Different lowercase letters indicate significant differences between treatments at p < 0.05.
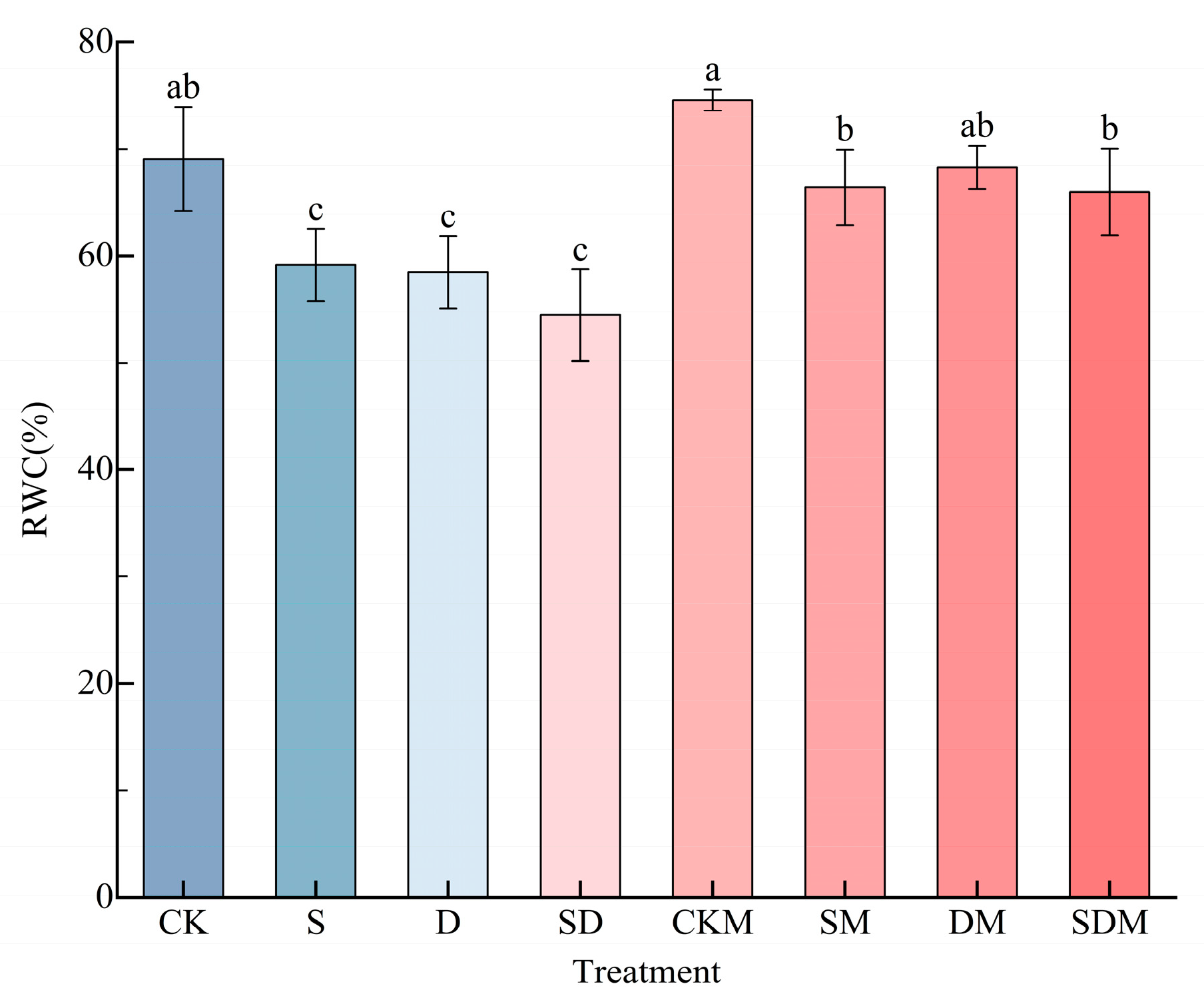
Figure 2.
Effects of exogenous melatonin on proline and soluble sugar contents in the blade of Cyperus esculentus seedlings under salt and drought stress. Note: Different lowercase letters indicate significant differences between treatments at p < 0.05.
Figure 2.
Effects of exogenous melatonin on proline and soluble sugar contents in the blade of Cyperus esculentus seedlings under salt and drought stress. Note: Different lowercase letters indicate significant differences between treatments at p < 0.05.

Figure 3.
Effects of exogenous melatonin on MDA content and relative electrical conductivity in blade of Cyperus esculentus seedlings under salt and drought stress. Note: Different lowercase letters indicate significant differences between treatments at p < 0.05.
Figure 3.
Effects of exogenous melatonin on MDA content and relative electrical conductivity in blade of Cyperus esculentus seedlings under salt and drought stress. Note: Different lowercase letters indicate significant differences between treatments at p < 0.05.
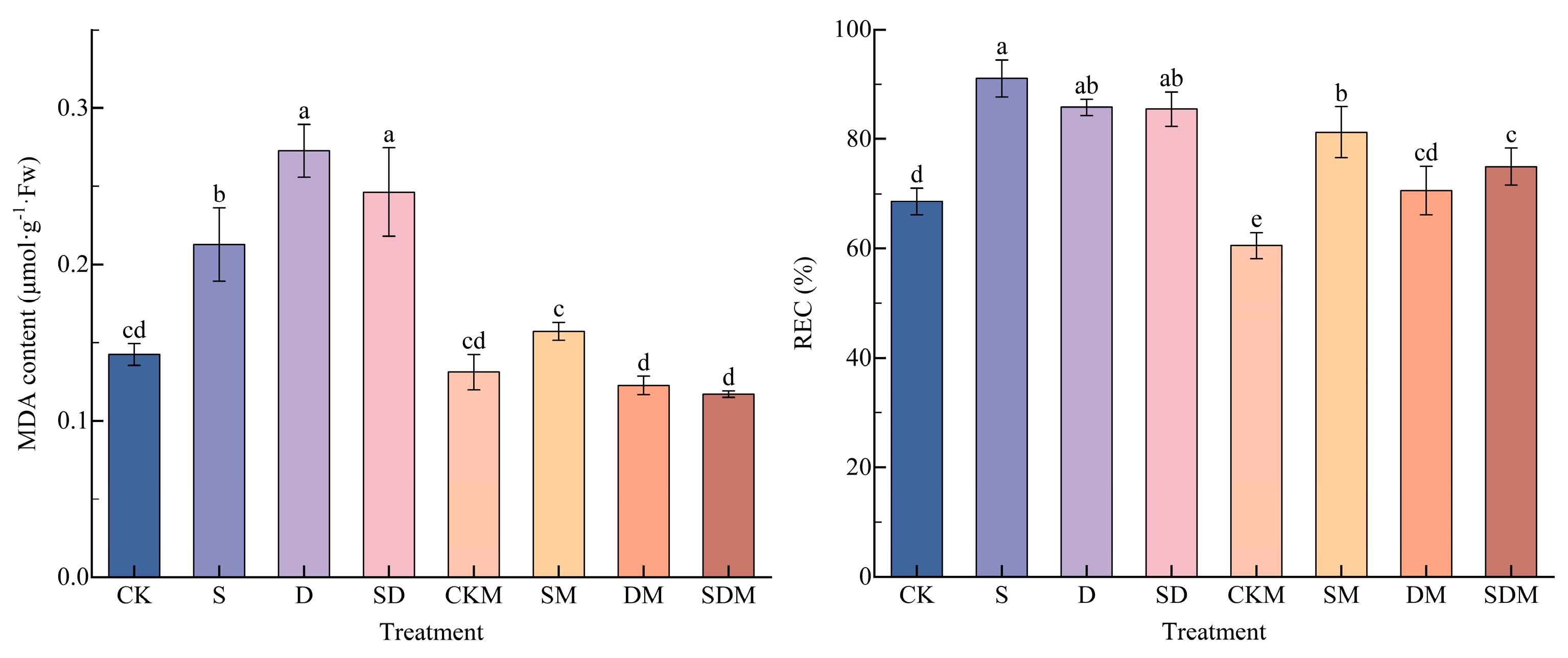
Figure 4.
Effects of exogenous melatonin on H2O2 and O2·− in the blade of Cyperus esculentus seedlings under salt and drought stress. Note: Different lowercase letters indicate significant differences between treatments at p < 0.05.
Figure 4.
Effects of exogenous melatonin on H2O2 and O2·− in the blade of Cyperus esculentus seedlings under salt and drought stress. Note: Different lowercase letters indicate significant differences between treatments at p < 0.05.
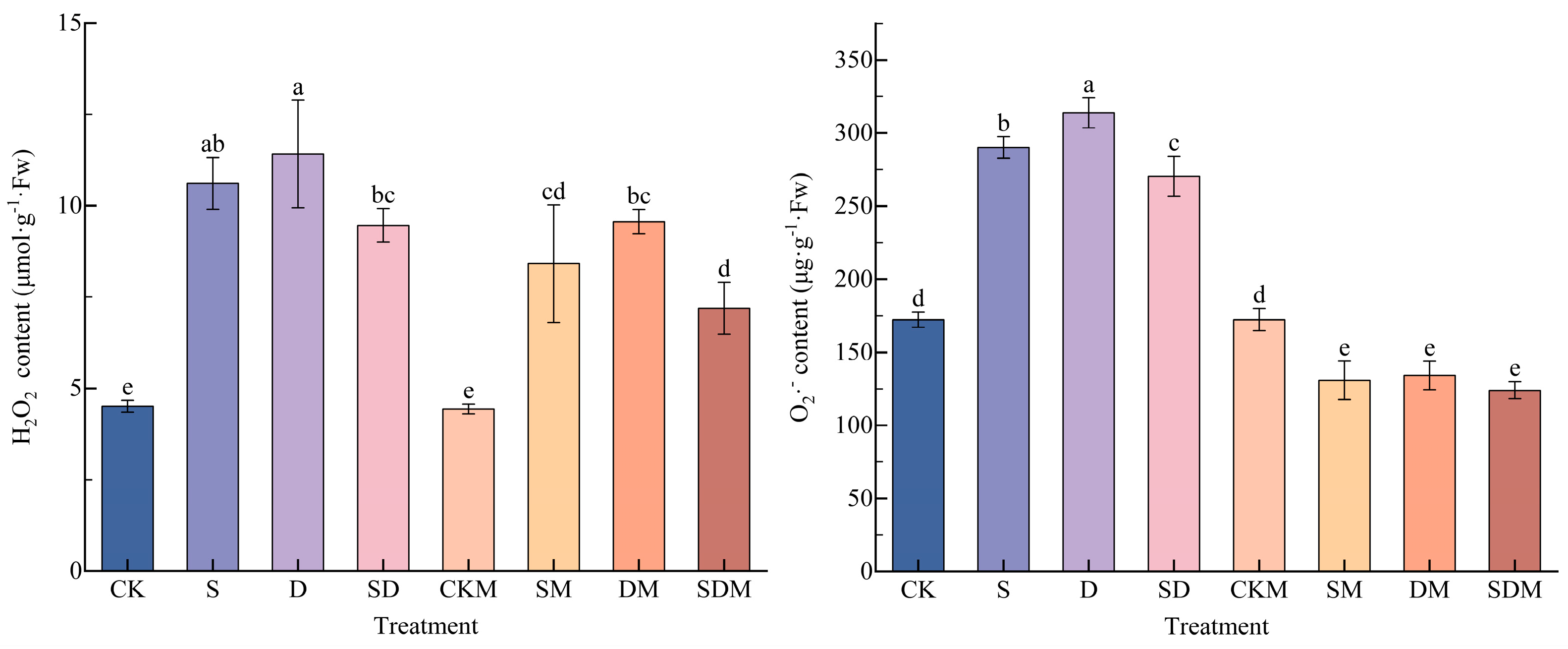
Figure 5.
Effects of exogenous melatonin on antioxidant enzyme activity in the blade of Cyperus esculentus seedlings under salt and drought stress. Note: Different lowercase letters indicate significant differences between treatments at p < 0.05.
Figure 5.
Effects of exogenous melatonin on antioxidant enzyme activity in the blade of Cyperus esculentus seedlings under salt and drought stress. Note: Different lowercase letters indicate significant differences between treatments at p < 0.05.
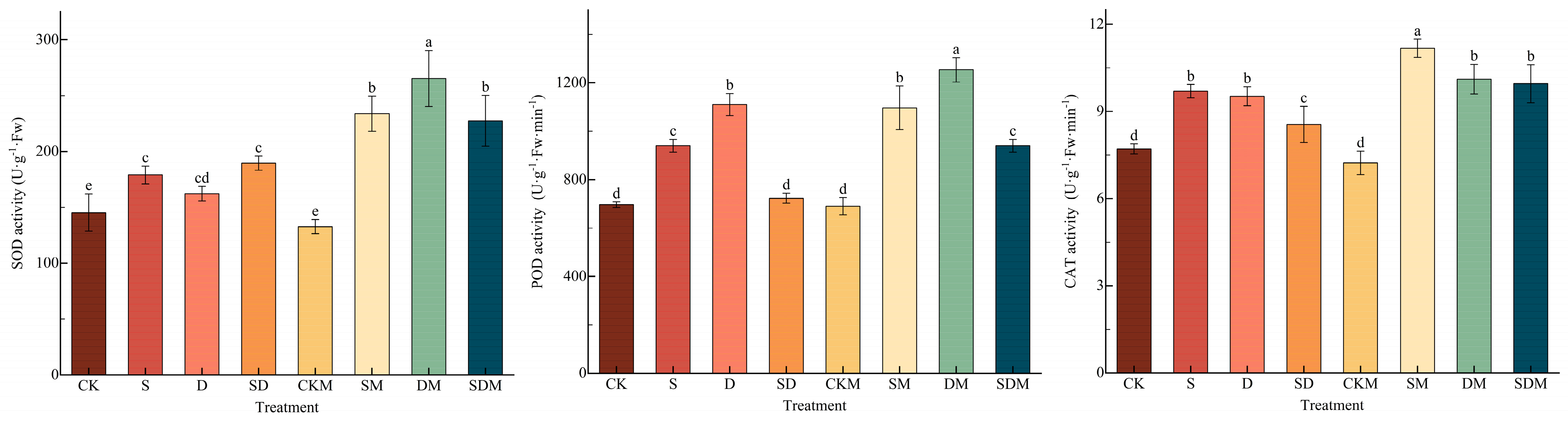
Figure 6.
Correlation analysis between morphological and physiological indexes of Cyperus esculentus L. Note: Red indicates a positive correlation; blue indicates a negative correlation. The number is the correlation coefficient. * means p < 0.05, ** means p < 0.01, *** means p < 0.001. PH: plant hight; NB: number of blades.
Figure 6.
Correlation analysis between morphological and physiological indexes of Cyperus esculentus L. Note: Red indicates a positive correlation; blue indicates a negative correlation. The number is the correlation coefficient. * means p < 0.05, ** means p < 0.01, *** means p < 0.001. PH: plant hight; NB: number of blades.
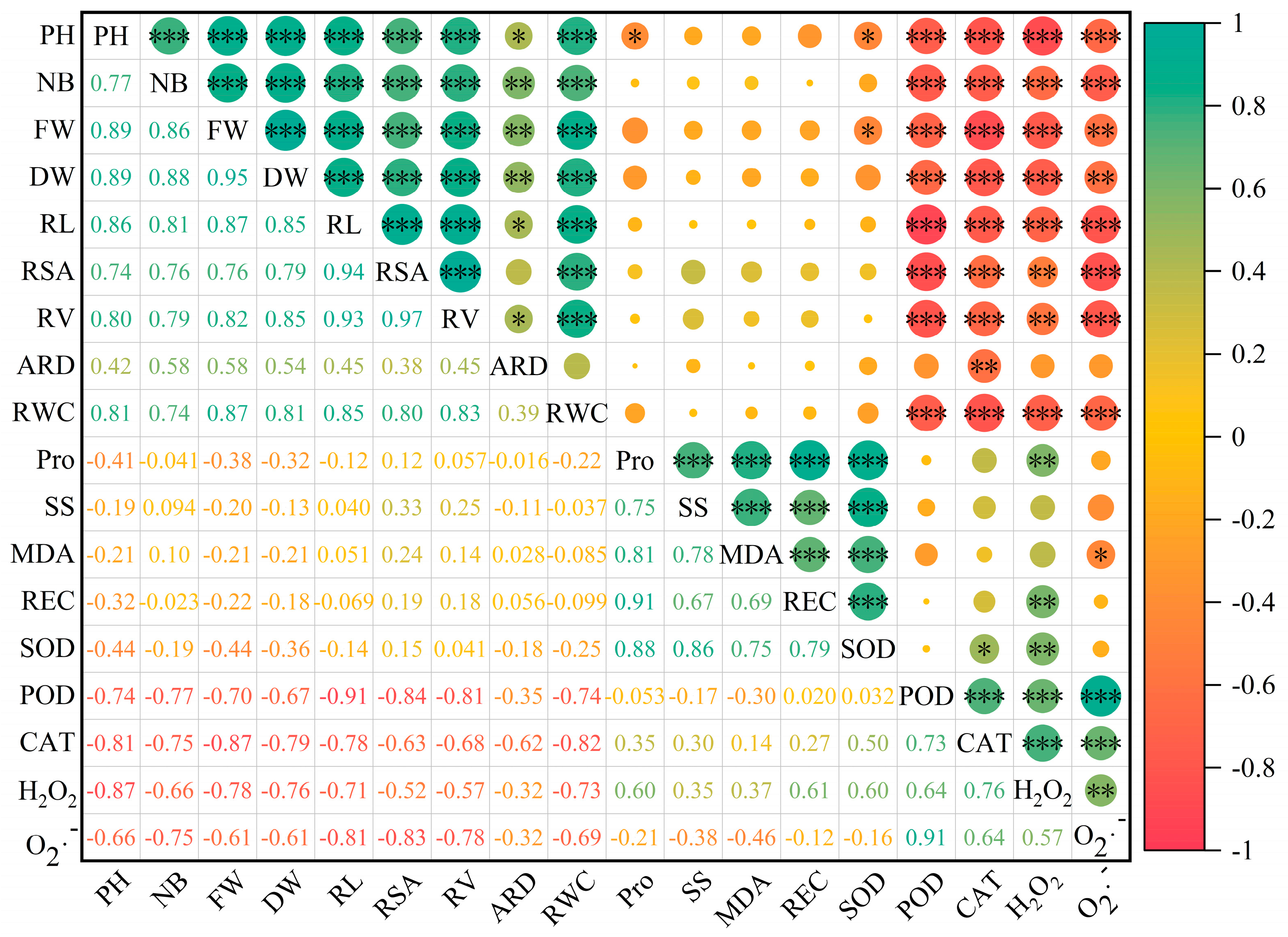
Figure 7.
Principal component analysis of morphological and physiological indexes of Cyperus esculentus.
Figure 7.
Principal component analysis of morphological and physiological indexes of Cyperus esculentus.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Combination of exogenous melatonin and salt drought stress in pot experiments.
| Treatment | Melatonin Content (μmol·L−1) | NaCl Content (mmol·L−1) | Field Capacity (%) |
|---|---|---|---|
| CK | 0 | 0 | 70 |
| S | 0 | 200 | 70 |
| D | 0 | 0 | 50 |
| SD | 0 | 200 | 50 |
| CKM | 150 | 0 | 70 |
| SM | 150 | 200 | 70 |
| DM | 150 | 0 | 50 |
| SDM | 150 | 200 | 50 |
Table 2.
Effect of melatonin on the agronomic traits of Cyperus esculentus at the tillering stage under salt and drought stress.
Table 2.
Effect of melatonin on the agronomic traits of Cyperus esculentus at the tillering stage under salt and drought stress.
| Treatment | Plant Height (cm) | Number of Blade | Fresh Weight (g) | Dry Weight (g) |
|---|---|---|---|---|
| CK | 32.07 ± 0.43 a | 27.00 ± 0.82 ab | 11.90 ± 0.54 b | 2.38 ± 0.15 a |
| S | 24.94 ± 0.46 c | 17.67 ± 2.05 d | 7.76 ± 0.15 f | 1.22 ± 0.07 d |
| D | 24.30 ± 0.93 c | 18.00 ± 0.82 d | 7.53 ± 0.40 f | 1.42 ± 0.03 cd |
| SD | 24.00 ± 1.06 c | 19.00 ± 0.82 cd | 6.67 ± 0.50 g | 1.02 ± 0.06 e |
| CKM | 31.87 ± 1.58 a | 27.33 ± 1.25 ab | 13.42 ± 0.28 a | 2.50 ± 0.16 a |
| SM | 28.66 ± 1.81 b | 24.67 ± 1.70 b | 9.64 ± 0.26 d | 1.96 ± 0.09 b |
| DM | 27.99 ± 0.61 b | 28.00 ± 1.63 a | 10.85 ± 0.21 c | 1.93 ± 0.03 b |
| SDM | 27.52 ± 0.75 b | 21.67 ± 0.47 c | 8.73 ± 0.34 e | 1.50 ± 0.06 c |
Note: Different lowercase letters indicate significant differences between treatments at p < 0.05.
Table 3.
Effect of melatonin on root morphogenesis of Cyperus esculentus at the tillering stage under salt and drought stress.
Table 3.
Effect of melatonin on root morphogenesis of Cyperus esculentus at the tillering stage under salt and drought stress.
| Treatment | RL (cm) | RSA (cm2) | RV (cm3) | ARD (mm) |
|---|---|---|---|---|
| CK | 1312.11 ± 47.72 a | 307.64 ± 15.06 b | 6.52 ± 0.48 a | 0.81 ± 0.08 a |
| S | 997.71 ± 47.02 c | 235.28 ± 8.83 c | 4.61 ± 0.59 c | 0.76 ± 0.04 a |
| D | 812.93 ± 46.90 d | 203.72 ± 10.67 d | 4.27 ± 0.69 c | 0.78 ± 0.08 a |
| SD | 716.84 ± 69.56 e | 147.16 ± 11.13 e | 2.49 ± 0.32 d | 0.77 ± 0.07 a |
| CKM | 1338.24 ± 52.85 a | 307.61 ± 12.24 b | 6.95 ± 0.47 a | 0.81 ± 0.06 a |
| SM | 1180.98 ± 52.16 b | 334.39 ± 21.48 a | 7.03 ± 0.55 a | 0.78 ± 0.04 a |
| DM | 1232.57 ± 56.32 b | 313.05 ± 28.15 ab | 6.61 ± 0.47 a | 0.82 ± 0.07 a |
| SDM | 1200.82 ± 65.05 b | 288.99 ± 27.19 b | 5.69 ± 0.67 b | 0.78 ± 0.04 a |
Note: Different lowercase letters indicate significant differences between treatments at p < 0.05. RL: root length; RSA: root surface area; RV: root volume; ARD: average root diameter.
Table 4.
Comprehensive score and ranking of melatonin under salt and drought stress.
| Treatment | Principal Component Score | Comprehensive Score (F) | Comprehensive Score Ranking | |
|---|---|---|---|---|
| PC1 (F1) | PC2 (F2) | |||
| CK | 0.32 | −0.43 | 0.08 | 5 |
| S | −0.29 | −0.05 | −0.19 | 6 |
| D | −0.33 | −0.11 | −0.23 | 7 |
| SD | −0.38 | −0.37 | −0.34 | 8 |
| CKM | 0.41 | −0.5 | 0.11 | 3 |
| SM | 0.07 | 0.6 | 0.21 | 2 |
| DM | 0.16 | 0.61 | 0.27 | 1 |
| SDM | 0.03 | 0.26 | 0.09 | 4 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Wang, N.; Luo, X.; Wang, Z.; Liu, J. Mitigating Effect of Exogenous Melatonin on Salt and Drought Stress in Cyperus esculentus L. during the Tillering Stage. Agronomy 2024, 14, 1009. https://doi.org/10.3390/agronomy14051009
AMA Style
Wang N, Luo X, Wang Z, Liu J. Mitigating Effect of Exogenous Melatonin on Salt and Drought Stress in Cyperus esculentus L. during the Tillering Stage. Agronomy. 2024; 14(5):1009. https://doi.org/10.3390/agronomy14051009
Chicago/Turabian StyleWang, Ningning, Xuemei Luo, Zhen Wang, and Jianguo Liu. 2024. "Mitigating Effect of Exogenous Melatonin on Salt and Drought Stress in Cyperus esculentus L. during the Tillering Stage" Agronomy 14, no. 5: 1009. https://doi.org/10.3390/agronomy14051009
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.