

Effect of Varied Salinity on Marigold Flowers: Reduced Size and Quantity Despite Enhanced Antioxidant Activity


1
Estacion Biológica La Ceiba, Chisec 16013, Guatemala
2
Forest Research Center (CEF), Associate Laboratory TERRA, School of Agriculture (ISA), University of Lisbon (UL), Tapada da Ajuda, 1349-017 Lisbon, Portugal
*
Author to whom correspondence should be addressed.
Agronomy 2023, 13(12), 3076; https://doi.org/10.3390/agronomy13123076
Submission received: 16 November 2023
/
Revised: 6 December 2023
/
Accepted: 11 December 2023
/
Published: 17 December 2023
(This article belongs to the Special Issue Selected Papers from the 3rd International Electronic Conference on Agronomy (IECAG 2023))
Abstract
:Salinity is an increasing problem worldwide that limits crop production. The cultivation of salt-tolerant flowers is a potential sustainable strategy for the exploitation of saline soils while avoiding the use of freshwater resources. However, our understanding of how flowers can adapt to salinity is still limited. In this study, we investigated the effects of salinity on a widely-used ornamental plant (Tagetes patula) by submitting three cultivars (cv. Aurora Orange, Fireball, Safari Scarlet) to different salinity levels (0, 50, 100, 300 mM). The plants were grown under controlled conditions and followed over two weeks. We tested the effect of salinity on plant growth and flower production, as well as on the levels of total carotenoids, polyphenols, and flavonoids, and the activities of antioxidant enzymes (catalase, peroxidase activity, ascorbate peroxidase). The findings indicate a substantial decline in both plant growth and flower production under saline conditions. Overall, plant height was ¼ lower, and there were 1/3 less flowers under salinity. Additionally, there was a consistent rise in antioxidant compounds, highlighting the swift response of defense mechanisms, both enzymatic and non-enzymatic, to combat stress. The high levels of antioxidant compounds sustain the use of marigold flowers as a new source of nutritional compounds, with enriched nutritional contents. Yet, cultivation of these plants in saline conditions should carefully consider the pronounced adverse effects observed at high salinity levels (e.g., 100 and 300 mM) on both plant and flower production.
1. Introduction
Soil salinity stands out as one of the most severe abiotic stresses, since it causes major reductions in cultivated land area, as well as in crop yield and quality [1]. High soil salinity causes ion toxicity, osmotic stress, nutrient deficiency, and oxidative stress in most plants. This stress condition also leads to oxidative stress, resulting in the overproduction of reactive oxygen species (ROS) [2,3]. To counterbalance these effects, plants activate a complex detoxification system through the actions of enzymatic antioxidants, including superoxide dismutase (SOD), catalase (CAT), ascorbate peroxidase (APX), glutathione reductase (GR), peroxidase (POX), and non-enzymatic antioxidants such as vitamins, flavonoids, and carotenoids, to reduce the excess ROS [4,5,6,7]. At the same time, plant antioxidants play a potential role in human diets, and can be used for many purposes, including serving as new sources of bioactive and nutritional compounds [8]. Thus, exploring the role of these antioxidants not only provides important information on plant adaptation mechanisms to harsh environmental conditions but also identifies functional compounds with benefits for human health.
In response to high salinity, salt-tolerant species elevate antioxidant levels to balance ROS, coordinating cellular processes such as water uptake and ion regulation [6,9,10,11]. However, most plants are not tolerant to salinity, and a high degree of responses can even be found among sensitive and tolerant species. These responses also vary within the same species, being also dependent on the duration and the level of the stress [10,12,13,14]. For instance, salinity delays flowering in some species such as rice [15] and Arabidopsis [16], while in others, namely salt-tolerant species, flowering occurs earlier and faster [17]. But even in salt-tolerant plants, there can be a reduction in pollen fertility, seed set, and seed viability in the presence of salinity [18]. Ultimately, the combined effects of osmotic stress and ion toxicity triggered by salinity generate a secondary stress that affects germination, growth, and overall plant development [6,19].
Understanding salt tolerance in ornamental plants also holds global significance as it could contribute to the conservation of freshwater resources in floriculture [20]. Although some research has advanced our understanding of salt tolerance of ornamental plants, the underlying mechanisms are complex as they vary widely between plants. For instance, some ornamental plants, such as snapdragon (Antirrhinum majus), petunia (Petunia × hybrida), verbena (Verbena × hybrida), coleus (Solenostemon scutellarioides), and begonia (Begonia hiemalis) can tolerate moderate levels of salinity (7.0–14.2 dS m−1 NaCl for 5 weeks) without a significant reduction in plant growth [21]. Other ornamental plants, such as Zinnia, are highly affected by salinity, even at low concentrations [22], while others, such as periwinkle (Catharanthus roseus), may show some tolerance, but only under low salinity levels (up to 3 dS m−1 [23]). Thus, to breed new salt-tolerant plants, it is important to understand the tolerance threshold where salinity does not compromise physiological processes, resulting in decreased quality or reduced yield.
Marigold species (Tagetes L.) demonstrate potential for cultivation under specific levels of salinity and are highly demanded in the flavor, fragrance, food, medicinal, and floricultural industries [24]. Marigolds contain a variety of bioactive components, including carotenoids, flavonoids, and phenolic acids, making their flowers valuable in the search for antioxidant compounds [25]. Pigments such as zeaxanthin and lutein from marigold flowers have also been used as food coloring agents [26,27]. Nevertheless, most marigold plants are highly affected by abiotic factors such as drought and temperature [28,29,30]. However, some marigold species and varieties are viewed as promising cash crops that can grow in saline environments, although with significant challenges. For instance, exposure to short-duration salinity levels (10 days at 50, and 100 nM NaCl) of T. patula L. (French marigold) plants triggered an accumulation of antioxidants and minerals in its flowers, but also led to a decrease in plant weight and the marketability of post-harvested flowers [24]. The growth of T. erecta L. (Aztec or Mexican marigold) cv. Sumo orange plants under 0, 50, 100, 150, and 200 mM NaCl for 25 days was also affected by increased salinity [31]. As salinity stress increased, Ca2+, Mg2+, and Na+ concentrations also increased, while the K+ concentration decreased in the roots and shoots. Additionally, the K+/Na+ and Ca2+/Na+ ratios of the roots and shoots were significantly lower in all salinity treatments, even though plants received a salt pre-treatment (except controls) to avoid a salt stress shock [31]. Seed germination of T. minuta L. (Mint marigold) cv. Him Swarnima grown under 0, 50, 100, 150, and 200 mM of soluble salts (CaCl2, KCl, MgCl2, Na2SO4, and NaCl) decreased significantly with increased levels of salinity under all salts, especially after 100 mM [32].
In this study, we investigated the salt-tolerance mechanisms of three French marigold cultivars (cv. Aurora Orange, Fireball, Safari Scarlet) under varying NaCl concentrations (0, 50, 100, and 300 mM). Our aim was to understand the extent to which flowers can tolerate salinity. We studied the effects of salinity on overall plant growth and flower production, examining how enzymatic and non-enzymatic components reacted to salinity. Despite the positive impacts from an edible point of view, salinity may impose constraints on plant development. While marigolds have been reported to tolerate salinity, being a source of edible flowers and nutritional compounds, the level of salinity and the cultivars used play a crucial role [24]. Thus, we specifically aim to (1) unravel the impacts of different salinity levels on French marigold flowers, (2) understand the antioxidant response mechanisms of flowers to salt stress, (3) detect the occurrence of intraspecific tolerance variability to salinity, and (4) test the salt-tolerance trade-off between the level of beneficial antioxidant compounds versus plant growth and flower production.
2. Materials and Methods
2.1. Plant Material and Experimental Conditions
Seeds from three Tagetes patula widely sold cultivars (cv. Aurora Orange, Fireball, Safari Scarlet) were chosen for this study since they are recommended for use in gardens, including city and courtyard gardens, and coastal sites as they are thought to be resistant to abiotic stresses. Yet, the effects of stresses have not been tested in these varieties. Thus, initially seeds were grown in 2 L capacity pots (50% peat, 25% perlite, and 25% vermiculite following [30]) in a nursery (T = 20.5–25.3 °C; RH = 72–76%, light and dark period = 16:8 h) for 3 weeks to ensure plant uniformity at the stage of two-true leaves. Plants were watered (50 mL per pot) twice a week using Hoagland nutritive solution (Na2Fe·EDTA 20 mg/L, H3BO3 2.86 mg/L, MnSO4·4H2O 2.13 mg/L, ZnSO4·7H2O 0.22 mg/L, CuSO4·7H2O 0.08 mg/L, (NH4)6Mo7O24 0.02 mg/L).
Seedlings were then transplanted to 200 L capacity tanks and transferred to a controlled environmental chamber with the following conditions: long-day photoperiod (16 h light), temperature of 23 and 19 °C during the light and dark periods, respectively, and relative humidity of 72–76%, with an irradiance of 700–800 μmol m−2 s−1 (Tenney WIT-983; New Columbia, PN, USA). Each drained pot contained only one plant being supported by a polyethylene tray. Plants were irrigated with sterile water (200 mL per pot) twice a week and allowed to acclimatize for one month, before the imposition of stress. The optimal pH and EC of the NS were 5.8 and 2.1 mS cm−1, respectively.
After one month, plants started to flower and were exposed to the following salinity treatments: 0, 50, 100, and 300 mM NaCl levels for two weeks following previous stress experiments [24]. Each treatment consisted of 10 biological replications, totaling 40 plants (biological replicates) per cultivar. The regular electrical conductivity (EC) of the nutrient solution was 2.0 mS cm−1 for the control treatment (0 mM NaCl), 6.0 mS cm−1 for the salinity of 50 mM NaCl, 10.0 mS cm−1 for the salinity of 100 mM NaCl, and 28.0 mS cm−1 for the salinity of 300 mM NaCl, being constant during salinity stress treatments. At the end of the experiment, the following traits were measured: plant height (cm), number of flowers produced, and size of flowers, e.g., its diameter (cm). Flowers were then harvested and dried using a thermo-ventilated oven at 65 °C.
2.2. Determination of Total Carotenoid, Polyphenol, and Flavonoid Content
The total carotenoid content was based on 500 mg of fresh flowers, which were extracted with 30 mL of 80% ice-cold acetone before being vortexed and centrifuged. The supernatant was separated, and its absorbance was measured at 480 nm, as described by [24]. The following equation was used for quantification: Carotenoids (μg) = 4 × A480 × volume (mL). Results were converted and expressed as mg of carotenoids per gram of DW.
The total polyphenol content of flowers was determined based on extracts prepared from 500 mg of fresh flowers following repeated (four-fold) addition of 2.5 mL 50% (v/v) methanol under ultrasonication (MSE Sonicprep 150 ultrasonic disintegrator; Sanyo, Loughborough, UK). The aliquot was collected and centrifugated for 5 min at 4 °C at 3000× g (Harrier 18/80, Sanyo Scientific, Torrance, CA, USA). From the supernatant, we collected and transferred to a new Eppendorf tube 125 μL, adding 1.5 mL of Milli-Q water, 125 μL of FolinCiocalteu reagent (Sigma Aldrich, Poole, UK), and 1.25 mL of 7% (w/v) sodium carbonate. The reaction mix (3 mL) was incubated in the dark for 90 min. The absorbance was then measured at 755 nm (Genesys 10 Vis, ThermoSpectronic, Rochester, NY, USA) according to [24]. Results were expressed in terms of μmol of gallic acid equivalents (GAE; Sigma Aldrich, Poole, UK) per gram of DW.
The total flavonoid content was extracted from 500 mg of fresh flowers, following [30], using 4 mL of Milli-Q water and 0.3 mL of sodium nitrite solution (5% concentration). The reaction mix was incubated for 5 min, after which we added 0.3 mL of aluminium chloride (10% concentration). The sample was left to incubate for 6 min, after which we added 2 mL of sodium hydroxide (1 M). Immediately after that, the final volume was completed and mixed with water (Milli-Q) until 10 mL. The absorbance was measured at 510 nm. Results were expressed as mg of catechin equivalents (CE) per gram of DW.
2.3. Antioxidative Enzyme Activities
Maximal cellular enzyme activities were assayed using 500 mg fresh weight (FW) of flower tissues, pooled from 3 flowers per plant and treatment. After that, samples were homogeneized under 1 mL of buffer containing 200 mM Tris-HCl (pH 8), 10 mM MgCl2 6H2O, 30 mM β-mercaptoethanol, 4 mM DTT, 2% Triton X-100, “Complete cocktail EDTA” (2 pills), and 10% glycerol, adding 1% (1 mL) of polyvinylpolypyrrolidone (PVPP) to each sample in the homogenization phase. The samples were centrifuged (13,000× g, 20 min, 4 °C), and the supernatant was used to quantify enzymatic activities.
The activity of catalase (CAT) was determined in a 1.5 mL reaction mixture containing 10 mM H2O2, 50 mM phosphate buffer (pH 7.0), and the enzyme extract. The activity was evaluated following [24] through the rate of H2O2 consumption at 240 nm. Results were expressed in CAT units mg−1 of protein DW (1 unit = 1 mM of H2O2 reduction per min).
Peroxidase activity (POX) was determined as described by [33], by measuring the absorbance at 430 nm, and using an extinction coefficient of 2.47 mM−1 cm−1 for calculations. Results were expressed in POX units/mg of protein. One POX unit was defined as the amount of enzyme to decompose 1 μmol of H2O2 per minute. Results were expressed in units mg−1 of protein DW.
Ascorbate peroxidase (APX) was determined according to [34]. Each reaction mixture contained 20 mM ascorbate and 0.1 mM H2O2 in 50 mM phosphate buffer (pH 7.8) and 10 μL of the enzyme extract in a total volume of 1 mL. The sample reaction was assessed through the H2O2-dependent oxidation of ascorbate at 290 nm, using an extinction coefficient of 2.8 mM−1 cm−1 for calculations. Results were expressed in APX units mg−1 of protein DW.
2.4. Statistical Analysis
Mean values (±SE) were calculated from the 10 replicates per cultivar using IBM SPSS v.22. To analyze the effects of salinity within each cultivar, we used a multivariate ANOVA at the 5% significance level after checking the homogeneity of variance using Levene’s Test for Equality of Variances. Additionally, a second ANOVA was performed to test for significant differences between cultivars within the same salinity level. Significant differences between means were also followed by Tukey’s test for post-hoc comparisons at the 5% significance level. The Pearson correlation coefficient was used to test the relationship between plant height, the number of flowers produced, size, and the production of antioxidant compounds (carotenoids, polyphenols, and flavonoids) and the activity of antioxidative enzymes (CAT, POX, APX).
3. Results
3.1. Influence of Salinity on Plant Growth and Flower Production
Salinity significantly decreased the plant height of T. patula cv. Aurora Orange (F3,14 = 10.035; p = 0.001), cv. Fireball (F3,14 = 52.518; p = 0.001), and cv. Safari Scarlet (F3,14= 5.788; p= 0.011). Overall, salinity reduced plant height by 1/4 in comparison to non-saline conditions. In cv. Aurora Orange, plant height was significantly reduced under 100 mM and 300 mM NaCl, while no significant differences were found between control and 50 mM NaCl conditions (Figure 1A). In contrast, plant height already decreased in the other two cultivars under the lowest salinity level (Figure 1A).
Salinity also affected the number of flowers produced (Figure 1B), since a significant decrease of almost 1/3 was reported with increased salinity levels, either in cv. Aurora Orange (F3,14 = 9.331; p = 0.002), cv. Fireball (F3,14 = 37.500; p = 0.001), or cv. Safari Scarlet (F3,14 = 10.008; p= 0.001). The average number of flowers decreased from 32 and 21 in cv. Aurora Orange, 30 and 20 in cv. Fireball, and from 24 and 18 in cv. Safari Scarlet, as salinity increased. This negative effect was especially felt under 300 mM NaCl in all cultivars, where the lowest number of flowers was recorded due to the abortion of several flower buds (Figure 1B).
Likewise, salinity also affected the size of flowers, becoming smaller with the increase in salinity (Figure 1C). The average flower size decreased from 4.46 to 3.58 in cv. Aurora Orange, 3.56 to 3.20 in cv. Fireball, and from 3.54 to 2.52 in cv. Safari Scarlet as salinity increased. Significant differences were recorded either in cv. Aurora Orange (F3,14 = 8.461; p = 0.003), cv. Fireball (F3,14 = 2.716; p = 0.032), or cv. Safari Scarlet (F3,14 = 7.310; p = 0.005). Some intraspecific variability was found since the size of flowers in cvs. Aurora Orange and Fireball was only significantly affected by the highest salinity levels (100 and 300 mM NaCl) while cv. Safari Scarlet showed the lowest size only under 300 mM NaCl (Figure 1C).
3.2. Determination of Total Carotenoid, Polyphenol, and Flavonoid Content
Carotenoids varied between 1.23 and 3.44 in cv. Aurora Orange, 1.28 and 3.19 in cv. Fireball, and between 1.27 and 3.74 in cv. Safari Scarlet (Table 1). They significantly increased with salinity, almost triplicating the values under the highest concentration (F3,14 = 12.370; p = 0.0002). The levels of polyphenols also increased as salinity increased (F3,16 = 9.561; p = 0.0002). Polyphenols varied between 23.21 and 55.45 in cv. Aurora Orange, 22.25 and 55.66 in cv. Fireball, and 24.21 and 58.11 in cv. Safari Scarlet, and doubled under 300 mM NaCl (Table 1). Likewise, salinity increased the levels of flavonoids, more than doubling under the highest salinity concentration (F3,11 = 11.221; p = 0.0003): 4.31 and 10.41 in cv. Aurora Orange, 4.23 and 11.23 in cv. Fireball, and 6.29 and 14.74 in cv. Safari Scarlet (Table 1). The highest levels were reported under 300 mM NaCl in all cultivars (Table 1). Significant differences were found between cultivars, being the levels of carotenoids, polyphenols, and flavonoids higher in cv. Safari Scarlet than in the other cultivars (F2,11 = 4.291; p = 0.0001, F2,10 = 5.506; p = 0.0001 and F2,8 = 3.101; p = 0.0001, respectively). Also, in the case of cv. Aurora Orange, the levels of carotenoids only increased under 100 and 300 mM NaCl in comparison to control conditions (Table 1).
3.3. Antioxidative Enzyme Activities
Enzyme activities increased significantly with enhanced levels of salinity, considering CAT (F3,14 = 8.221; p = 0.0002), POX (F3,11 = 6.734; p = 0.002), or APX (F3,16 = 6.356; p = 0.001) (Table 2). The activity of CAT more than doubled with salinity, varying from 1.23 to 3.44 in cv. Aurora Orange, 1.28 to 3.19 in cv. Fireball, and 1.37 to 3.74 in cv. Safari Scarlet (Table 2). In cv. Aurora Orange, CAT activity only increased at the highest salinity levels (100 and 300 mM NaCl), while in the other cultivars, the activity of this enzyme already increased at 50 mM NaCl. The activity of POX also more than doubled with the increase in salinity levels. POX activity varied between 23.21 and 55.45 in cv. Aurora Orange, 22.25 to 55.66 in cv. Fireball, and 24.21 to 58.11 in cv. Safari Scarlet (Table 2). A significant variation in the activity of enzymes was found between cultivars, with enzymatic levels usually higher in cv. Safari Scarlet than in the other two cultivars (Table 2). Nevertheless, the highest level of activity was recorded at 300 mM NaCl in all cultivars, and considering all enzymes tested.
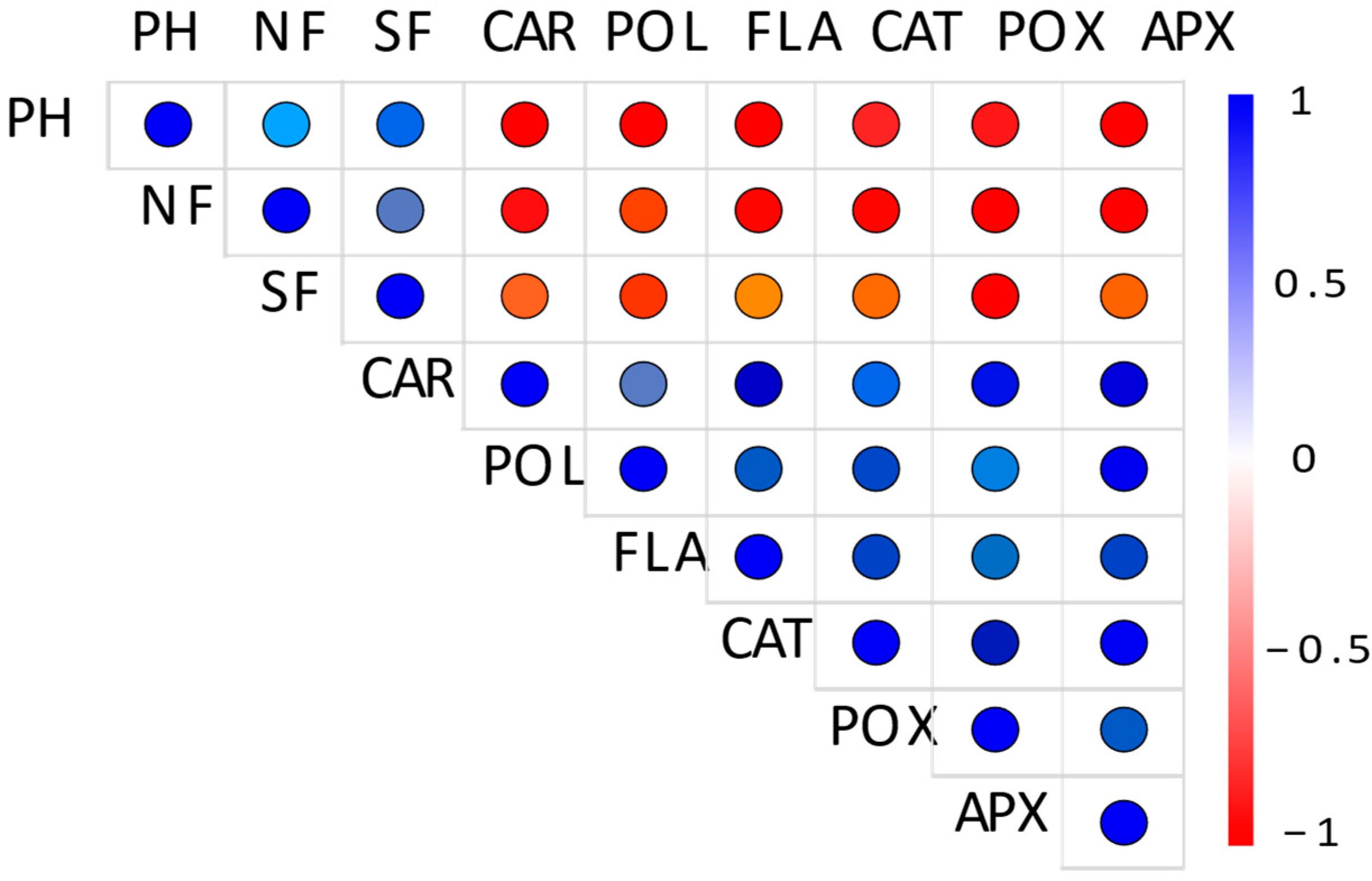
Overall, considering all cultivars together, a significant negative correlation was found between plant traits (plant height, number of flowers, and their sizes) and the activity of antioxidant compounds and enzymes, as salinity increased (Figure 2; p < 0.05).
4. Discussion
4.1. Effects of Salinity on Plant Height and Flower Production
Salinity significantly reduced plant height, the number of flowers, and their size in all cultivars studied (Figure 1), likely due to the stress triggered by salinity [35]. This oxidative, osmotic, and toxic stress affects the direction of cell division, the formation of phragmoplasts, and the structural organization of cells, often leading to significant damage in the cytoskeleton [36,37,38]. Consequently, these processes may lead to the death of cells and/or changes in plant morphology and growth [36,37], as reported here.
Flowering and the underlying processes of the development of flower reproductive structures are also disrupted by adverse conditions. For instance, the number of inflorescences in the halophyte Crithmum maritimum decreased significantly as salinity increased, at least in some genotypes [39]. Here, we also found a decrease in the number of flowers with the increase in salinity levels. Likewise, flower abortion, reduction of pollen number, and viability have often been reported to be high in other species [40], being limiting factors in the floriculture industry [35]. In addition, some morphological changes were already seen in some flowers, especially under 300 mM NaCl, together with a high number of aborted buds as reported here. The expression of some MADS-box transcription factors, which regulate flower development suffer changes under salinity [41,42] and may be responsible for the levels of aborted flowers, although this needs to be further studied.
4.2. High Levels of Antioxidant Molecules on Flowers
To balance the levels of ROS produced by saline stress, plants trigger several antioxidant compounds, such as the ones reported here. As such, carotenoids increased as salinity increased in all cultivars, except in Aurora Orange, where no differences were found between control conditions and the lowest saline level (Table 1). This differs from a previous study involving other T. patula cultivars, where an increase in carotenoid content was reported under saline conditions, although with higher levels under 50 mM NaCl and decreasing under 100 mM NaCl [24]. Carotenoids are involved in quenching 1O2 and peroxyl radicals generated during excess excitation of the chlorophyll, helping to protect the photosynthetic apparatus and avoiding plant damage [43]. In salt-tolerant plants, salinity triggers carotenoid biosynthesis and increases carotenoid concentration, as reported in the taproots of carrots [44] or the leaves of Thellungiella [45], being key for achieving tolerance under saline conditions [46]. Thus, the increased levels of carotenoids under saline conditions seem important to maintain the quality of marigold flowers. Carotenoids are an important contributor to the petal colors of marigolds, and during flower development, carotenoid biosynthesis genes are upregulated [26].
As reported here, total polyphenol and flavonoid content also plays a role in Tagetes responses, since their levels increase with increased salinity levels in all cultivars. Increases in total phenols have also been reported in the flowers of other T. patula cultivars, while the levels of anthocyanins decrease under salinity stress [24]. Water-soluble polyphenols are a major group of antioxidants in many species, showing a rapid ability to scavenge O2·–, OH, H2O2, and 1O2 compared with the lipid-soluble carotenoids [47]. Among phenolic compounds, flavonoids are the largest and best-studied group, playing a major role under abiotic stresses [47]. Increased flavonoid levels are crucial for balancing Na+/K+ ions, reducing the effects of stress through the regulation of transcriptional and hormonal mechanisms involving ABA hormone signaling [48]. Thus, altogether, the three components—carotenoids, polyphenols, and flavonoids—were highly responsive to saline stress in French marigold flowers.
4.3. The Role of Antioxidant Enzymatic Machinery on Flowers
Together with non-enzymatic components, the high activity of the enzymes CAT, POX, and APX suggests a fast response of marigold flowers to balance the oxidative stress imposed by salinity in all studied cultivars. CATs are primary scavenging enzymes that directly dismutate H2O2 and reduce the levels of ROS, especially under drought and salt stress because of their critical involvement in photorespiration [49]. POXs are a family of isoenzymes, also capable of scavenging H2O2, mainly in the apoplastic space [50]. Another major hydrogen peroxide detoxifying system under abiotic stresses is the ascorbate-glutathione cycle, where APX enzymes catalyze the conversion of H2O2 into H2O, using ascorbate as a specific electron donor [51]. Therefore, the increased activity of these enzymes in the stressed French marigold plants studied here allows for the removal of the H2O2 produced by salinity. The activity of CAT, SOD, APX, and POD has been found to increase in the flowers of other T. patula cultivars, albeit only under 50 mM NaCl and decreasing under 100 mM NaCl [24], contrary to our results. This highlights the need to study more cultivars, since responses to stresses are very dependent on the cultivar used, as reported here and in other studies [24,28,30,31,52,53].
4.4. Usefulness of Saline Conditions for French Marigold Plants: Implications for the Floriculture Industry
Ornamental plants are usually irrigated with high-quality water. Yet, as freshwater resources become scarcer and soil salinity becomes higher, the cultivation of plants that can tolerate (at least some) degree of salinity becomes a helpful solution. Ornamental plants that can grow and survive under saline levels at or over 200 mM NaCl are considered salt-tolerant, providing a beneficial resource for use in saline or degraded soils [54]. However, few plants can tolerate high levels of salinity [21]. It is, thus, interesting to find tolerant T. patula cultivars, at least to some levels of salinity, as reported in this study. In fact, after the end of the experiment, the plants returned to non-saline conditions and no cases of senescence or plant mortality were recorded, suggesting that these plants could recover from short-term salinity exposures. Nonetheless, since reproductive stages are usually highly sensitive to stresses [55], further evaluation is needed to understand the effects of salinity on flower development and reproductive processes, including under higher periods of exposure. The significant negative correlations already found in this study between plant height, the number and sizes of flowers, and the activity of antioxidant compounds and enzymes might change under different salinity exposures. Tagetes patula flowers are also viewed as nutritional or functional foods, with potential economic value [24]. There is a growing interest in the antioxidant compounds, carotenoids, flavonoids, and polyphenols produced by plants since they may contribute to human health. An ongoing study suggests that these cultivars have high levels of total phenolic content and proteins, while the level of fatty acids is relatively low, supporting the presence of nutritional compounds in these Tagetes cultivars (unpub. results).
Based on our results, farmers who are considering the use of saline conditions to produce flowers with nutritional compounds could start to employ low levels of salinity. However, due to the negative effects of salinity on French marigold flowers, we recommend the production of T. patula flowers under short exposure to salinity (of up to 50 mM NaCl) to achieve high production of nutritional compounds without compromising flower production. Higher values may be beneficial to produce antioxidants and minerals, but they significantly compromise the quantity of flowers produced.
5. Conclusions
Salinity significantly reduced plant height, the number of flowers, and their size in all cultivars studied, despite the increase in antioxidant compounds. These compounds are beneficial for human consumption and increased at the lowest salinity level (50 mM). This stress also had minimal effects on plant growth and flower production, suggesting the possibility of using saline irrigation water to enhance useful antioxidant compounds. Thus, these cultivars can be used in saline soils, but only under small levels of salinity (up to 50mM NaCl). This contrasts sharply with the adverse impacts observed at higher salinity levels (100 and 300 mM NaCl). Future studies should investigate the effects of varying salt levels over different durations on flower production to provide a more comprehensive understanding. Exploring additional methodological approaches, such as exogenous foliar application, may prove beneficial for enhancing salt tolerance in these plants, as suggested by previous studies [56,57]. Using saline water holds significance for sustainable agricultural practices by: (1) providing an alternative to farmers beyond traditional glycophyte crops, (2) helping diversify the agricultural market, and (3) contributing to the sustainable use of scarce freshwater resources. Thus, based on our results, salinity triggered an increase in important health antioxidant compounds that can be used as a new source of nutritional food, despite the negative impacts on plant height and flower production.
Author Contributions
Conceptualization, I.M.; methodology, I.M.; formal analysis, M.R.G.; investigation, M.R.G.; data curation, M.R.G.; writing—original draft preparation, M.R.G. and I.M; writing—review and editing, M.R.G. and I.M.; supervision, I.M.; project administration, I.M.; funding acquisition, I.M. All authors have read and agreed to the published version of the manuscript.
Funding
This research received national funds through the FCT—Fundação para a Ciência e a Tecnologia, I.P., Portugal through the UIDB/00239/2020 (CEF), the associate laboratory TERRA (LA/P/0092/2020), and the Scientific Employment Stimulus—Individual Call (CEEC Individual)—2021.01107.CEECIND/CP1689/CT0001 (IM).
Data Availability Statement
The data presented in this study are available in the manuscript itself.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Yamaguchi, T.; Blumwald, E. Developing salt-tolerant crop plants: Challenges and opportunities. Trends Plant Sci. 2005, 10, 615–620. [Google Scholar] [CrossRef] [PubMed]
- Hasanuzzaman, M.; Raihan, M.R.H.; Masud, A.A.C.; Rahman, K.; Nowroz, F.; Rahman, M.; Nahar, K.; Fujita, M. Regulation of reactive oxygen species and antioxidant defense in plants under salinity. Int. J. Mol. Sci. 2021, 22, 9326. [Google Scholar] [CrossRef] [PubMed]
- Kamran, M.; Parveen, A.; Ahmar, S.; Malik, Z.; Hussain, S.; Chattha, M.S.; Saleem, M.H.; Adil, M.; Heidari, P.; Chen, J.T. An overview of hazardous impacts of soil salinity in crops, tolerance mechanisms, and amelioration through selenium supplementation. Int. J. Mol. Sci. 2020, 21, 148. [Google Scholar] [CrossRef] [PubMed]
- Arif, Y.; Singh, P.; Siddiqui, H.; Bajguz, A.; Hayat, S. Salinity induced physiological and biochemical changes in plants: An omic approach towards salt stress tolerance. Plant Physiol. Biochem. 2020, 156, 64–77. [Google Scholar] [CrossRef] [PubMed]
- Vinci, G.; Marques, I.; Rodrigues, A.P.; Martins, S.; Leitão, A.E.; Semedo, M.C.; Silva, M.J.; Lidon, F.C.; DaMatta, F.M.; Ribeiro-Barros, A.I.; et al. Protective Responses at the Biochemical and Molecular Level Differ between a Coffea arabica L. Hybrid and Its Parental Genotypes to Supra-Optimal Temperatures and Elevated Air [CO2]. Plants 2022, 11, 2702. [Google Scholar] [CrossRef] [PubMed]
- Laamari, I.; Marques, I.; Ribeiro-Barros, A.I.; Zoubeir, B.; Abassi, M. Can saline preconditioning enhance plant survival in degraded soils? Physiological, biochemical, and molecular responses in Casuarina glauca saplings. Plant Ecol. 2023, 224, 905–919. [Google Scholar] [CrossRef]
- Gill, S.S.; Tuteja, N. Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol. Biochem. 2010, 48, 909–930. [Google Scholar] [CrossRef] [PubMed]
- Loizzo, M.R.; Tundis, R. Plant antioxidant for application in food and nutraceutical industries. Antioxidants 2019, 8, 453. [Google Scholar] [CrossRef]
- Hajhashemi, V.; Vaseghi, G.; Pourfarzam, M.; Abdollahi, A. Are antioxidants helpful for disease prevention? Res. Pharm. Sci. 2010, 5, 5–12. [Google Scholar]
- Al Kharusi, L.; Al Yahyai, R.; Yaish, M.W. Antioxidant response to salinity in salt-tolerant and salt-susceptible cultivars of date palm. Agriculture 2019, 9, 8. [Google Scholar] [CrossRef]
- Fernandes, I.; Paulo, O.S.; Marques, I.; Sarjkar, I.; Sen, A.; Graça, I.; Pawlowski, K.; Ramalho, J.C.; Ribeiro-Barros, A.I. Salt Stress Tolerance in Casuarina glauca: Insights from the Branchlets Transcriptome. Plants 2022, 11, 2942. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.; Zheng, J.; Zhang, X.; Hu, Q.; Qian, R. Salicylic acid alleviates the adverse effects of salt stress on Dianthus superbus (Caryophyllaceae) by activating photosynthesis, protecting morphological structure, and enhancing the antioxidant system. Front. Plant Sci. 2017, 8, 243823. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Fujita, M. Plant Responses and Tolerance to Salt Stress: Physiological and Molecular Interventions. Int. J. Mol. Sci. 2022, 23, 4810. [Google Scholar] [CrossRef]
- Tahjib-Ul-Arif, M.; Hasan, M.T.; Rahman, M.A.; Nuruzzaman, M.; Rahman, A.M.S.; Hasanuzzaman, M.; Haque, M.R.; Hossain, M.A.; Abdel Latef, A.A.H.; Murata, Y.; et al. Plant response to combined salinity and waterlogging stress: Current research progress and future prospects. Plant Stress 2023, 7, 100137. [Google Scholar] [CrossRef]
- Lutts, S.; Kinet, J.M.; Bouharmont, J. Changes in plant response to NaCl during development of rice (Oryza sativa L.) varieties differing in salinity resistance. J. Exp. Bot. 1995, 46, 1843–1852. [Google Scholar] [CrossRef]
- Park, H.J.; Kim, W.-Y.; Yun, D.-J. A New Insight of Salt Stress Signaling in Plant. Mol. Cells 2016, 39, 447–459. [Google Scholar] [CrossRef]
- Oh, D.-H.; Hong, H.; Lee, S.Y.; Yun, D.-J.; Bohnert, H.J.; Dassanayake, M. Genome Structures and Transcriptomes Signify Niche Adaptation for the Multiple-Ion-Tolerant Extremophyte Schrenkiella parvula. Plant Physiol. 2014, 164, 2123–2138. [Google Scholar] [CrossRef]
- Boscaiu, M.; Estrelles, E.; Soriano, P.; Vicente, Ó. Effects of salt stress on the reproductive biology of the halophyte Plantago crassifolia. Biol. Plant. 2005, 49, 141–143. [Google Scholar] [CrossRef]
- Liang, W.; Ma, X.; Wan, P.; Liu, L. Plant salt-tolerance mechanism: A review. Biochem. Biophys. Res. Commun. 2018, 495, 286–291. [Google Scholar] [CrossRef]
- Cassaniti, C.; Romano, D.; Hop, M.E.C.M.; Flowers, T.J. Growing floricultural crops with brackish water. Environ. Exp. Bot. 2013, 92, 165–175. [Google Scholar] [CrossRef]
- Villarino, G.H.; Mattson, N.S. Assessing tolerance to sodium chloride salinity in fourteen floriculture species. Horttechnology 2011, 21, 539–545. [Google Scholar] [CrossRef]
- Niu, G.; Wang, M.; Rodriguez, D.; Zhang, D. Response of Zinnia plants to saline water irrigation. HortScience 2012, 47, 793–797. [Google Scholar] [CrossRef]
- Marković, M.; Šoštarić, J.; Kojić, A.; Popović, B.; Bubalo, A.; Bošnjak, D.; Stanisavljević, A. Zinnia (Zinnia elegans L.) and Periwinkle (Catharanthus roseus (L.) G. Don) Responses to Salinity Stress. Water 2022, 14, 1066. [Google Scholar] [CrossRef]
- Chrysargyris, A.; Tzionis, A.; Xylia, P.; Tzortzakis, N. Effects of salinity on tagetes growth, physiology, and shelf life of edible flowers stored in passive modified atmosphere packaging or treated with ethanol. Front. Plant Sci. 2018, 871, 1765. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Gao, W.; Yun, X.; Qing, Z.; Zeng, J. Effect of Natural Antioxidants from Marigolds (Tagetes erecta L.) on the Oxidative Stability of Soybean Oil. Molecules 2022, 27, 2865. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Zhang, S.; Zhang, H.; Chen, X.; Liang, F.; Qin, H.; Zhang, Y.; Cong, R.; Xin, H.; Zhang, Z. Carotenoid metabolite and transcriptome dynamics underlying flower color in marigold (Tagetes erecta L.). Sci. Rep. 2020, 10, 16835. [Google Scholar] [CrossRef] [PubMed]
- Burlec, A.F.; Pecio, Ł.; Kozachok, S.; Mircea, C.; Corciovă, A.; Vereştiuc, L.; Cioancă, O.; Oleszek, W.; Hăncianu, M. Phytochemical profile, antioxidant activity, and cytotoxicity assessment of Tagetes erecta L. flowers. Molecules 2021, 26, 1201. [Google Scholar] [CrossRef]
- Kumar, A.; Gautam, R.D.; Kumar, A.; Singh, S.; Singh, S. Understanding the Effect of Different Abiotic Stresses on Wild Marigold (Tagetes minuta L.) and Role of Breeding Strategies for Developing Tolerant Lines. Front. Plant Sci. 2022, 12, 3332. [Google Scholar] [CrossRef]
- Cicevan, R.; Sestras, A.F.; Plazas, M.; Boscaiu, M.; Vilanova, S.; Gramazio, P.; Vicente, O.; Prohens, J.; Sestras, R.E. Biological Traits and Genetic Relationships Amongst Cultivars of Three Species of Tagetes (Asteraceae). Plants 2022, 11, 760. [Google Scholar] [CrossRef]
- Cicevan, R.; Al Hassan, M.; Sestras, A.F.; Prohens, J.; Vicente, O.; Sestras, R.E.; Boscaiu, M. Screening for drought tolerance in cultivars of the ornamental genus Tagetes (Asteraceae). PeerJ 2016, 2016, e2133. [Google Scholar] [CrossRef]
- Koksal, N.; Alkan-Torun, A.; Kulahlioglu, I.; Ertargin, E.; Karalar, E. Ion uptake of marigold under saline growth conditions. Springerplus 2016, 5, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Gautam, R.D.; Singh, S.; Chauhan, R.; Kumar, A.; Singh, S. Comparative study of the effects of different soluble salts on seed germination of wild marigold (Tagetes minuta L.). J. Appl. Res. Med. Aromat. Plants 2022, 31, 100421. [Google Scholar] [CrossRef]
- Tarchoune, I.; Sgherri, C.; Izzo, R.; Lachaal, M.; Ouerghi, Z.; Navari-Izzo, F. Antioxidative responses of Ocimum basilicum to sodium chloride or sodium sulphate salinization. Plant Physiol. Biochem. PPB 2010, 48, 772–777. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Z.; Wei, G.; Li, J.; Qian, Q.; Yu, J. Silicon alleviates salt stress and increases antioxidant enzymes activity in leaves of salt-stressed cucumber (Cucumis sativus L.). Plant Sci. 2004, 167, 527–533. [Google Scholar] [CrossRef]
- García-Caparrós, P.; Lao, M.T. The effects of salt stress on ornamental plants and integrative cultivation practices. Sci. Hortic. 2018, 240, 430–439. [Google Scholar] [CrossRef]
- Baranova, E.N.; Gulevich, A.A. Asymmetry of Plant Cell Divisions under Salt Stress. Symmetry 2021, 13, 1811. [Google Scholar] [CrossRef]
- Bano, C.; Amist, N.; Singh, N.B. Morphological and Anatomical Modifications of Plants for Environmental Stresses. Mol. Plant Abiotic Stress Biol. Biotechnol. 2019, 29–44. [Google Scholar] [CrossRef]
- Zagoskina, N.V.; Zubova, M.Y.; Nechaeva, T.L.; Kazantseva, V.V.; Goncharuk, E.A.; Katanskaya, V.M.; Baranova, E.N.; Aksenova, M.A. Polyphenols in Plants: Structure, Biosynthesis, Abiotic Stress Regulation, and Practical Applications (Review). Int. J. Mol. Sci. 2023, 24, 13874. [Google Scholar] [CrossRef]
- Ventura, Y.; Myrzabayeva, M.; Alikulov, Z.; Omarov, R.; Khozin-Goldberg, I.; Sagi, M. Effects of salinity on flowering, morphology, biomass accumulation and leaf metabolites in an edible halophyte. AoB Plants 2014, 6, plu053. [Google Scholar] [CrossRef]
- Ghanem, M.E.; Van Elteren, J.; Albacete, A.; Quinet, M.; Martínez-Andújar, C.; Kinet, J.M.; Pérez-Alfocea, F.; Lutts, S. Impact of salinity on early reproductive physiology of tomato (Solanum lycopersicum) in relation to a heterogeneous distribution of toxic ions in flower organs. Funct. Plant Biol. 2009, 36, 125–136. [Google Scholar] [CrossRef]
- Kaashyap, M.; Ford, R.; Mann, A.; Varshney, R.K.; Siddique, K.H.M.; Mantri, N. Comparative Flower Transcriptome Network Analysis Reveals DEGs Involved in Chickpea Reproductive Success during Salinity. Plants 2022, 11, 434. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Fatima, M.; Zhou, P.; Ma, Q.; Ming, R. Analysis of MADS-box genes revealed modified flowering gene network and diurnal expression in pineapple. BMC Genom. 2020, 21, 8. [Google Scholar] [CrossRef] [PubMed]
- Giuliano, G.; Al-Babili, S.; Von Lintig, J. Carotenoid oxygenases: Cleave it or leave it. Trends Plant Sci. 2003, 8, 145–149. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.H.; Deng, Y.J.; Wang, Y.H.; Lou, Y.R.; He, L.F.; Liu, H.; Li, T.; Yan, Z.M.; Zhuang, J.; Xiong, A.S. Changes in Carotenoid Concentration and Expression of Carotenoid Biosynthesis Genes in Daucus carota Taproots in Response to Increased Salinity. Horticulturae 2022, 8, 650. [Google Scholar] [CrossRef]
- Stepien, P.; Johnson, G.N. Contrasting Responses of Photosynthesis to Salt Stress in the Glycophyte Arabidopsis and the Halophyte Thellungiella: Role of the Plastid Terminal Oxidase as an Alternative Electron Sink. Plant Physiol. 2009, 149, 1154–1165. [Google Scholar] [CrossRef] [PubMed]
- Bose, J.; Rodrigo-Moreno, A.; Shabala, S. ROS homeostasis in halophytes in the context of salinity stress tolerance. J. Exp. Bot. 2014, 65, 1241–1257. [Google Scholar] [CrossRef] [PubMed]
- Frary, A.; Göl, D.; Keleş, D.; Ökmen, B.; Pinar, H.; Şiǧva, H.T.; Yemenicioǧlu, A.; Doǧanlar, S. Salt tolerance in Solanum pennellii: Antioxidant response and related QTL. BMC Plant Biol. 2010, 10, 58. [Google Scholar] [CrossRef]
- Jan, R.; Kim, N.; Lee, S.H.; Khan, M.A.; Asaf, S.; Lubna; Park, J.R.; Asif, S.; Lee, I.J.; Kim, K.M. Enhanced Flavonoid Accumulation Reduces Combined Salt and Heat Stress Through Regulation of Transcriptional and Hormonal Mechanisms. Front. Plant Sci. 2021, 12, 796956. [Google Scholar] [CrossRef]
- Mishra, N.; Jiang, C.; Chen, L.; Paul, A.; Chatterjee, A.; Shen, G. Achieving abiotic stress tolerance in plants through antioxidative defense mechanisms. Front. Plant Sci. 2023, 14, 1110622. [Google Scholar] [CrossRef]
- Zandi, P.; Schnug, E. Reactive Oxygen Species, Antioxidant Responses and Implications from a Microbial Modulation Perspective. Biology 2022, 11, 155. [Google Scholar] [CrossRef]
- Sofo, A.; Scopa, A.; Nuzzaci, M.; Vitti, A. Ascorbate Peroxidase and Catalase Activities and Their Genetic Regulation in Plants Subjected to Drought and Salinity Stresses. Int. J. Mol. Sci. 2015, 16, 13561–13578. [Google Scholar] [CrossRef] [PubMed]
- Moliner, C.; Barros, L.; Dias, M.I.; López, V.; Langa, E.; Ferreira, I.C.F.R.; Gómez-Rincón, C. Edible flowers of tagetes erecta l. As functional ingredients: Phenolic composition, antioxidant and protective effects on Caenorhabditis elegans. Nutrients 2018, 10, 2002. [Google Scholar] [CrossRef] [PubMed]
- Chisowa, E.H.; Hall, D.R.; Farman, D.I. Chemical composition of the essential oil of Tagetes minuta L. from Zambia. J. Essent. Oil Res. 1998, 10, 183–184. [Google Scholar] [CrossRef]
- Guo, J.; Shan, C.; Zhang, Y.; Wang, X.; Tian, H.; Han, G.; Zhang, Y.; Wang, B. Mechanisms of Salt Tolerance and Molecular Breeding of Salt-Tolerant Ornamental Plants. Front. Plant Sci. 2022, 13, 854116. [Google Scholar] [CrossRef]
- Kazan, K.; Lyons, R. The link between flowering time and stress tolerance. J. Exp. Bot. 2016, 67, 47–60. [Google Scholar] [CrossRef]
- Zrig, A.; AbdElgawad, H.; Touneckti, T.; Ben Mohamed, H.; Hamouda, F.; Khemira, H. Potassium and calcium improve salt tolerance of Thymus vulgaris by activating the antioxidant systems. Sci. Hortic. 2021, 277, 109812. [Google Scholar] [CrossRef]
- Jahan, M.S.; Li, G.; Xie, D.; Farag, R.; Hasan, M.M.; Alabdallah, N.M.; Al-Harbi, N.A.; Al-Qahtani, S.M.; Zeeshan, M.; Nasar, J.; et al. Melatonin Mitigates Salt-Induced Growth Inhibition Through the Regulation of Carbohydrate and Nitrogen Metabolism in Tomato Seedlings. J. Soil Sci. Plant Nutr. 2023, 23, 4290–4308. [Google Scholar] [CrossRef]
Figure 1.
Impact of salinity levels (0, 50, 100, and 300 mM NaCl) on plant height, (A) the number of flowers produced, (B) and the size of flowers, (C) considering Tagetes patula cultivars cv. Aurora Orange, Fireball, and Safari Scarlet. Each box plot represents the minimum, lower quartile, median, upper quartile, and maximum values. Different superscripts indicate significant differences between salinity levels for the same cultivar (ANOVA followed by a Tukey test at p < 0.05).
Figure 1.
Impact of salinity levels (0, 50, 100, and 300 mM NaCl) on plant height, (A) the number of flowers produced, (B) and the size of flowers, (C) considering Tagetes patula cultivars cv. Aurora Orange, Fireball, and Safari Scarlet. Each box plot represents the minimum, lower quartile, median, upper quartile, and maximum values. Different superscripts indicate significant differences between salinity levels for the same cultivar (ANOVA followed by a Tukey test at p < 0.05).

Figure 2.
Pearson’s correlation among the different morphological and physiological parameters considering Tagetes patula cultivars exposed to different salt stress levels (0, 50, 100, and 300 mM NaCl). Each circle indicates the Pearson’s correlation coefficient between a pair of parameters. PH; plant height, NF; number of flowers, SF; size of flowers, CAR; carotenoids, POL; total polyphenols, FLA; flavonoids, CAT; catalase, POX; peroxidase, and APX; ascorbate peroxidase. In all cases, the correlation was significant at 0.05.
Figure 2.
Pearson’s correlation among the different morphological and physiological parameters considering Tagetes patula cultivars exposed to different salt stress levels (0, 50, 100, and 300 mM NaCl). Each circle indicates the Pearson’s correlation coefficient between a pair of parameters. PH; plant height, NF; number of flowers, SF; size of flowers, CAR; carotenoids, POL; total polyphenols, FLA; flavonoids, CAT; catalase, POX; peroxidase, and APX; ascorbate peroxidase. In all cases, the correlation was significant at 0.05.

{kind=link}
{kind=link}
Table 1.
Impact of salinity levels (0, 50, 100, and 300 mM NaCl) on total carotenoids, polyphenols, and flavonoids on the flowers of three Tagetes patula cultivars (cv. Aurora Orange, Fireball, and Safari Scarlet). Results are expressed as mean values ± SE (n = 10). Different superscripts indicate significant differences between salinity levels for the same cultivar (ANOVA followed by a Tukey test at p < 0.05).
Table 1.
Impact of salinity levels (0, 50, 100, and 300 mM NaCl) on total carotenoids, polyphenols, and flavonoids on the flowers of three Tagetes patula cultivars (cv. Aurora Orange, Fireball, and Safari Scarlet). Results are expressed as mean values ± SE (n = 10). Different superscripts indicate significant differences between salinity levels for the same cultivar (ANOVA followed by a Tukey test at p < 0.05).
| 0 mM NaCl | 50 mM NaCl | 100 mM NaCl | 300 mM NaCl | ||
|---|---|---|---|---|---|
| Carotenoids | Aurora Orange | 1.23 ± 0.10 c | 1.29 ± 0.15 c | 2.64 ± 0.71 b | 3.44 ± 0.91 a |
| Fireball | 1.28 ± 0.12 d | 1.42 ± 0.19 c | 2.59 ± 0.81 b | 3.19 ± 0.95 a | |
| Safari Scarlet | 1.27 ± 0.14 d | 1.57 ± 0.21 c | 3.11 ± 0.90 b | 3.74 ± 0.92 a | |
| Polyphenols | Aurora Orange | 23.21 ± 0.66 d | 33.99 ± 0.55 c | 44.61 ± 0.51 b | 55.45 ± 1.01 a |
| Fireball | 22.25 ± 0.55 d | 33.98 ± 0.66 c | 44.50 ± 0.44 b | 55.66 ± 0.99 a | |
| Safari Scarlet | 24.21 ± 0.77 d | 34.99 ± 0.88 c | 45.66 ± 0.90 b | 58.11 ± 1.75 a | |
| Flavonoids | Aurora Orange | 4.31 ± 0.34 d | 6.32 ± 0.44 c | 8.11 ± 0.61 b | 10.41 ± 1.92 a |
| Fireball | 4.23 ± 0.31 d | 6.23 ± 0.55 c | 8.59 ± 0.55 b | 11.23 ± 1.95 a | |
| Safari Scarlet | 6.29 ± 0.55 d | 7.44 ± 0.61 c | 9.11 ± 1.78 b | 14.74 ± 2.11 a |
Table 2.
Impact of salinity levels (0, 50, 100, and 300 mM NaCl) on enzyme activities of catalase (CAT), peroxidase (POX), and ascorbate peroxidase (APX), expressed as units/mg of protein DW on the flowers of three Tagetes patula cultivars (cv. Aurora Orange, Fireball, and Safari Scarlet). Results are expressed as mean values ± SE (n = 10). Different superscripts indicate significant differences between salinity levels for the same cultivar (ANOVA followed by a Tukey test at p < 0.05).
Table 2.
Impact of salinity levels (0, 50, 100, and 300 mM NaCl) on enzyme activities of catalase (CAT), peroxidase (POX), and ascorbate peroxidase (APX), expressed as units/mg of protein DW on the flowers of three Tagetes patula cultivars (cv. Aurora Orange, Fireball, and Safari Scarlet). Results are expressed as mean values ± SE (n = 10). Different superscripts indicate significant differences between salinity levels for the same cultivar (ANOVA followed by a Tukey test at p < 0.05).
| 0 mM NaCl | 50 mM NaCl | 100 mM NaCl | 300 mM NaCl | ||
|---|---|---|---|---|---|
| Aurora Orange | 1.23 ± 0.10 c | 1.29 ± 0.25 c | 2.64 ± 0.71 b | 3.44 ± 0.91 a | |
| CAT | Fireball | 1.28 ± 0.12 d | 1.42 ± 0.19 c | 2.59 ± 0.81 b | 3.19 ± 0.95 a |
| Safari Scarlet | 1.37 ± 0.14 d | 1.57 ± 0.21 c | 3.11 ± 0.90 b | 3.74 ± 0.92 a | |
| POX | Aurora Orange | 23.21 ± 0.66 d | 33.99 ± 0.55 c | 44.61 ± 0.51 b | 55.45 ± 1.01 a |
| Fireball | 22.25 ± 0.55 d | 33.98 ± 0.66 c | 44.50 ± 0.44 b | 55.66 ± 0.99 a | |
| Safari Scarlet | 24.21 ± 0.77 d | 34.99 ± 0.88 c | 45.66 ± 0.90 b | 58.11 ± 1.75 a | |
| APX | Aurora Orange | 4.31 ± 0.34 d | 6.32 ± 0.44 c | 8.11 ± 0.61 b | 10.41 ± 1.92 a |
| Fireball | 4.23 ± 0.31 d | 6.23 ± 0.55 c | 8.59 ± 0.55 b | 11.23 ± 1.95 a | |
| Safari Scarlet | 6.29 ± 0.55 d | 7.44 ± 0.61 c | 9.11 ± 1.78 b | 14.74 ± 2.11 a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Guzman, M.R.; Marques, I. Effect of Varied Salinity on Marigold Flowers: Reduced Size and Quantity Despite Enhanced Antioxidant Activity. Agronomy 2023, 13, 3076. https://doi.org/10.3390/agronomy13123076
AMA Style
Guzman MR, Marques I. Effect of Varied Salinity on Marigold Flowers: Reduced Size and Quantity Despite Enhanced Antioxidant Activity. Agronomy. 2023; 13(12):3076. https://doi.org/10.3390/agronomy13123076
Chicago/Turabian StyleGuzman, María Rita, and Isabel Marques. 2023. "Effect of Varied Salinity on Marigold Flowers: Reduced Size and Quantity Despite Enhanced Antioxidant Activity" Agronomy 13, no. 12: 3076. https://doi.org/10.3390/agronomy13123076
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.