Abstract
Cowpea is a nutrient-rich staple legume and climate-resilient crop for vulnerable agroecosystems. However, the crop still remains underutilized, mainly due to its narrow genetic base, and the production is often ravaged by aphid infestation outbreaks. Thus, genetic diversity assessment and the detection of defense-related alleles are fundamental to germplasm management and utilization in breeding strategies to support food safety in climate change times. A germplasm collection of 87 cowpea landraces sourced from Greece was subjected to seed phenotyping, SSR genotyping and to screening for the presence of aphid-resistance-conferring alleles. Significant diversity in the species’ local germplasm was revealed. The landraces were grouped in metapopulations based on their broader geographical origin. High amounts of variation and statistically significant differences were detected among the landraces regarding the seed morphological traits, the seed color and eye color according to MANOVA (Wilk’s λ = 0.2, p < 0.01) and significant correlations were revealed among these features according to Pearson’s test (p < 0.05). High levels of genetic polymorphism were detected for the metapopulations, ranging from 59% (VuPop3) to 82% (VuPop4). The AMOVA revealed that 93% of the molecular diversity was distributed among the landraces of each metapopulation. Further population structure analysis presumed the existence of two inferred populations, where in population A, 79% of the landraces have a cream/cream-brown seed coat, whereas in population B, 94% of the landraces are brown-ochre to black-seeded. Molecular screening for alleles conferring aphid resistance revealed the correspondence of 12 landraces to the resistant genotype of TVu-2876. The study highlights the importance of cowpea germplasm collection genetic diversity, as a source of important agronomic traits, to support breeding efforts and expand cowpea cultivation to foster food security and agriculture sustainability and diversification in climate change.
Keywords:
cowpea; germplasm; seed; phenotyping; diversity; molecular markers; aphid resistance; breeding 1. Introduction
Ongoing human population growth places great pressure on the production of agronomic resources, threatening food security, that is exacerbated by quickly escalating climate change. Furthermore, the recent pandemic crisis had an indirect impact in intensifying food insecurity, affecting disproportionally susceptible groups of the population [1]. In order to encounter undernourishment and the risk of a potential food crisis in vulnerable ecosystems, such as the Mediterranean, viable staple food production schemes and agriculture diversification are essential. Pulses have a key role in agriculture sustainability, diversification and food security.
Grain legumes, commonly known as pulses, are protein-rich crops cultivated in rainfed and low-input farming systems, promoting sustainable farming and enhancing soil nutrient composition and structure via biological nitrogen fixation [2]. Among pulses, cowpea holds an exceptional position due to its adaptability to sandy soils and drought, which have made it an important crop in semiarid regions. Cowpea (Vigna unguiculata L. Walp.) is a diploid (2n = 22) herbaceous, warm-season, predominantly self-pollinated (approx. 95%) annual crop of the Leguminosae family, domesticated in Western and Eastern Africa. Recent studies indicated that by 400 BCE, cowpea cultivation spread throughout the Mediterranean basin, sub-Saharan Africa and Southern Asia, and was transferred to the New World during colonization [3,4]. Its continuous cultivation in the Mediterranean region since antiquity is closely linked to the its socio-economic and cultural value for local communities [3,5], as depicted also by the broad phenotypic and genotypic diversity in the local germplasm [6].
Cowpea is cultivated in Southern Europe, subtropical Asia and America, while most of the world’s grain production share comes from sub-Saharan Africa (>95%). Its grains consist of up to 33% protein, 63% carbohydrates, 4.6% lipids, minerals, vitamins and fiber [7,8] and it constitutes a cheap protein source of energy for food and fodder, hence, demonstrating enormous potential for mitigating malnutrition [9] and supporting a plant protein diet. However, it is considered an underutilized crop, being inferior in yield, compared to other legumes that are cultivated in intensive farming schemes. Globally, dry seed yield remains below the potential of the crop (8.9 M tons in 2020), with an average yield of 5912 hg/ha in 2020, as it is grown predominantly by smallholder farmers in non-intensive, resource-poor traditional systems [10,11]. On the other hand, farmers prefer higher-yielding crops that provide increased income, such as dry peas (20,364 hg/ha), green peas (78,477 hg/ha), chickpeas (10,163 hg/ha), lentils (13,049 hg/ha) and soybeans (278,42 hg/ha) (FAOSTAT, 2020) [12]. Compared to other legumes, cowpea thrives in a range of soil types, pH and temperatures, while it is well adapted to drought-related conditions; therefore, it represents a promising climate-smart crop [13]. Thus, among other orphan crops, cowpea has lately gained researchers’ interest and efforts for the promotion of its cultivation in many countries. Yet, extensive breeding remains crucial to improve the crop’s yield and the food-value chain in a changing environment. Evaluation of the genetic diversity in the cowpea germplasm is a precondition for constructive breeding of the species. Cowpea’s molecular genetic diversity has already been examined with various marker systems, indicatively RAPDs, AFLPs, ISSRs, SNPs [14] and SSRs [15,16,17,18]. Among them, microsatellites comprise a robust genotyping method, extensively used in population genetic studies in various plant species. SSRs are multi-allelic, co-dominant, highly reproducible markers, with coverage extending along the entire genome. This marker system is equally proposed for genotype identification, pedigree analysis, genetic mapping of qualitative and quantitative traits, as well as for marker-assisted selection breeding strategies [19,20,21]. Taking into account the scarcity of genomic information on Greek cowpea accessions, SSR molecular markers constitute a functional approach for a thorough evaluation of untapped diversity.
Climate change and global warming adversely affect legume cropping systems, especially in drought-prone regions in the temperate zone, including the Mediterranean basin. Increased temperatures and altered precipitation patterns have significant impacts on cowpea production and on insect pests and pathogens. Specifically, changes in climate can result in an increased number of aphid generations, increased survival during overwintering, expansion of their geographic distribution, emergence of new biotypes, increased incidence of aphid-transmitted plant diseases and reduced effectiveness of biological control, especially natural enemies [22]. Globally, up to 20 viruses have been reported to infect cowpea. Many of them are seed borne, such as cowpea mosaic virus (CMV), and occur in several host species. Insect-borne viral transmission from diseased to healthy plants within fields and across neighboring fields may result in severe epidemics and crop damage. Among them, cowpea aphid-borne mosaic virus (CABMV) is responsible for crop losses of up to 87%, depending on the cultivar and the developmental stage of the plant [23]. Thus far, aphid population management relies mostly on prophylactic application of insecticides to properly secure the cowpea crop production. However, global warming drivers accounting for increased temperatures and a decrease in relative humidity may cause synthetic insecticides and management approaches to be less effective. Further, the use of synthetic pesticides further exacerbates the carbon and energy footprint and the environmental impact, raising concerns over field-evolved resistance of aphid populations to chemical formulations [24,25]. Therefore, breeding for aphid resistance is the most effective approach to support cowpea production in a climate-changing environment.
In this respect, evaluation of species genetic diversity in the local germplasm could further the discovery of valuable genes/alleles that contribute to yield and crop tolerance. Landraces constitute a major part of the local germplasm, encompassing substantial diversity levels, conserved both naturally and by farmer’s selection [26,27]. In Greece, cowpea comprises a valuable pulse crop of marginal lands and an important genetic pool of local landraces still in use for plant protein production and forage [28]. So far, molecular genetic characterization of the European cowpea germplasm is scarce and limited to Spanish and Italian accessions [29,30,31]. On the other hand, the Balkan peninsula located at the crossroads of three continents, Africa, Asia and Europe, is considered a diversity hotspot for several species, including cowpea; however, reports on cowpeas’ local germplasm molecular diversity are scarce. To address this knowledge gap, the present study aimed to explore the genetic diversity of 87 local cowpea accessions, through the assessment of important seed morphological traits and imaging as well as genotyping with SSR molecular markers. Additionally, the presence of aphid-resistance-conferring alleles was also investigated using linked molecular markers for further breeding strategies.
2. Materials and Methods
2.1. Plant Material
A germplasm collection of 87 landraces provided by the Institute of Plant Breeding and Genetic Resources of the Hellenic Agricultural Organization-DEMETER was collected from local farmers’ fields in Greece (Figure 1), where the seed progeny was recycled for every consecutive growing season. Details on plant material are demonstrated in Supplementary Table S1, according to IPGRB [32].
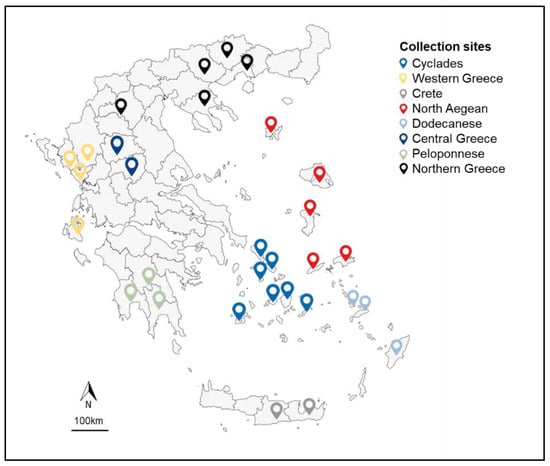
Figure 1.
Map of the collection sites of 87 V. unguiculata landraces, grouped in 8 metapopulations as to geographic origin.
2.2. Genetic Diversity Assessment
2.2.1. Seed Morphological Diversity Analysis
We estimated the Hundred-Seed Weight (HSW) by weighing 100 seeds in three independent replicates. The seed morphological diversity was assessed by subjecting 10 seeds of every sample to visual examination, according to IBPGR 1981 standard criteria, applying image analysis, as previously described [33]. Multivariate Analysis of Variance (MANOVA) was employed to detect statistically significant differences among the measured variables, according to Wilks’ Lambda test. Multiple-Factor Analysis (MFA) was used to cluster the 87 accessions in groups according to the Gower’s distance. The UPGMA dendrogram distinguished the accessions in different clusters. The aforementioned analyses were performed with R software version 4.2 [34].
2.2.2. Molecular Genetic Diversity Analysis
For every accession, the molecular genetic diversity was assessed on the bulked samples of 10 individuals. Genomic DNA was extracted from seeds, applying the QIAGEN DNeasy Plant Pro Kit (Qiagen, Germany). Qbit 4 Fluorometer (Thermo Fisher Scientific, USA) was used in order to estimate the yield of the extracted DNA for every sample, which was further normalized to 10 ng/μL for downstream applications. For the examination of the genetic diversity, we initially used 20 SSR molecular markers that were previously found to be polymorphic in cowpea population genetic studies [20,35]. In this germplasm collection, four of them (VM9, VM35, VM74, VM78) were not capable of amplifying all the accessions, while two (VM37, VM71) of them were monomorphic. Thus, those markers were excluded from the analysis (online Supplementary Table S2). We also screened the accessions with molecular markers KAD-61 and CP-171-172, in order to investigate the presence of alleles linked to aphid resistance [36]. PCR amplification reactions were carried out on a Veriti™ 96-Well Thermal Cycler, using KAPA Taq ReadyMix PCR Kit, in a total reaction volume of 15 μL, containing 1X KAPA Taq ReadyMix (1.5 mM MgCl2), 0.2 μM of each primer, PCR-grade H2O and 10 ng genomic DNA template.
The PCR amplification program constitutes an initial denaturation step at 94 °C/4 min, followed by 35 cycles of denaturation at 94 °C/30 s, primer annealing at the optimal Ta for each marker, extension at 72 °C/40 s and final extension at 72 °C/4 min. The amplified products of KAD-61 and CP-171-172 (5 μL of each PCR) were electrophoresed onto 2% agarose gel in 1x TAE buffer, stained with ethidium bromide and visualized by illumination with ultraviolet light. The amplified products of 14 SSR markers were further subjected to fragment analysis (CeMIA SA, Larissa, Greece) and data were analyzed using OSIRIS software [37].
For every sample, the SSR amplicons were classified as present (1) or absent (0) and were further used for the construction of a binary matrix. Landraces are cultivated germplasms, continually evolving in agricultural environments within a defined ecogeographical area and under the influence of local human culture [38]. Based on their geographical origin, the accessions were grouped into 8 metapopulations, in order to assess the genetic diversity in the cowpea collection [39]: Cyclades (VuPop1), Western Greece (VuPop2), Crete (VuPop3), North Aegean (VuPop4), Dodecanese (VuPop5), Central Greece (VuPop6), Peloponnese (VuPop7) and Northern Greece (VuPop8); they were subjected to estimation of their genetic diversity parameters, such as the number of total, effective and private alleles, the Shannon’s Information (I) and the diversity (h) indices. Analysis of Molecular Variance (AMOVA) was performed in order to explore the proportion of polymorphism distributed among and within the cowpea metapopulations, using GenAlEx 6.5 [40].
Genetic distances between the 8 metapopulations were calculated according to Nei’s genetic distance [41] and a dissimilarity matrix was generated, which was subjected to tree construction, under Unweighted Pair Group Method Analysis (UPGMA), using R software version 4.2 [34]. The Polymorphic Information Content of the SSR markers was calculated according to Smith et al. (1997) [42].
STRUCTURE v. 2.3.4 software was used to establish the metapopulation structure formed by 87 cowpea landraces, under the Bayesian clustering method [43]. The lengths of the burn-in and Markov Chain Monte Carlo (MCMC) were both set at 10,000 iterations [44]. Thirty runs for each K-value were performed with K ranging from 1 to 10, in order to obtain an accurate estimation of the number of populations. STRUCTURE Harvester was used to calculate Delta-K and the appropriate K value was chosen [45].
3. Results
3.1. Seed Morphological Diversity
The seed morphological measurements were statistically analyzed and summary statistics are presented in Table 1. Details on the seed coat color, eye color and testa texture are provided in Supplementary Table S1. According to the MANOVA, statistically significant differences were detected among the 87 accessions (Wilk’s lambda = 0.2, p < 0.01). More specifically, a wide variation was observed within VuPop1, VuPop2, VuPop3, VuPop4 and VuPop8 regarding the HSW and the seed area, contrary to VuPop5, VuPop6 and VuPop7, where most of the accessions approximate the mean value (Figure 2a).

Table 1.
Summary statistics of the seed morphological characteristics.
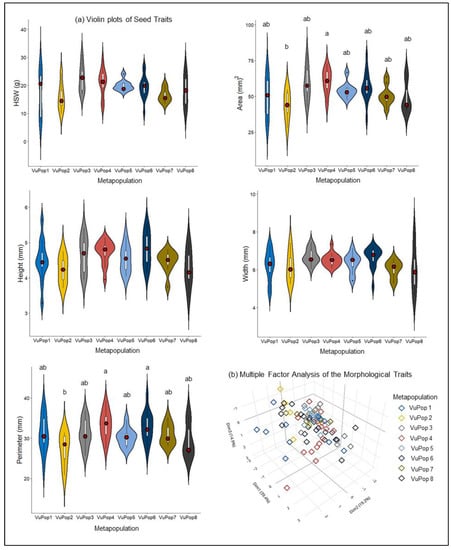
Figure 2.
(a) Violin plots of the HSW and seed geometrical features for the 8 cowpea metapopulations. Metapopulations that share no common letter are significantly different according to Tukey’s HSD (p < 0.05); (b) Multiple Factor Analysis plot of the cowpea metapopulations’ seed features, where the first three axes explain 73.5% of the observed variability.
Partitioning of the variance and grouping of the assumed populations with respect to their quantitative (hundred-seed weight, seed area, perimeter, circularity, width, Gray value, integrated density) and qualitative (seed color, seed shape, eye color, testa texture) seed morphological attributes were estimated by Factorial Mixed Data Analysis (FMDA). The quantitative traits include the hundred-seed weight (HSW), seed area (A), perimeter (P), circularity (C), height (H), width (W), Gray value (Gv) and integrated density (ID), whereas the qualitative traits include the seed color (SC), shape (SS), eye color (EC) and the testa texture (TT). All populations were grouped in a single cluster in the 3D MFA plot, where the first three dimensions represent 73.5% of the observed variability (Figure 2b).
A Pearson’s correlation test revealed a significantly very strong positive correlation between HSW and seed area (0.83), between area and perimeter (0.84) and between Gray value (mean and median) and cream seed color (0.85), suggesting that measurement of a single feature is sufficient to represent other highly correlated characteristics. Medium positive correlations were detected between seed height and area (0.6), perimeter (0.59) and weight (0.47), between seed width and HSW (0.5), area (0.63) and perimeter (0.56), as well as between Gray value and B eye color (0.53) and rough testa texture (0.6). Noteworthily, medium positive correlations were demonstrated between cream seed color and HSW (0.58), area (0.5), B eye color (0.64) and rough testa texture (0.64). Contrarily, moderate negative correlations were detected among brown seed color and HSW (−0.54), area (−0.49) and width (−0.4). Those data indicate the preference of local cowpea germplasm cultivation and breeding in Greece, which is directed towards cream seeded landraces (Figure 3).
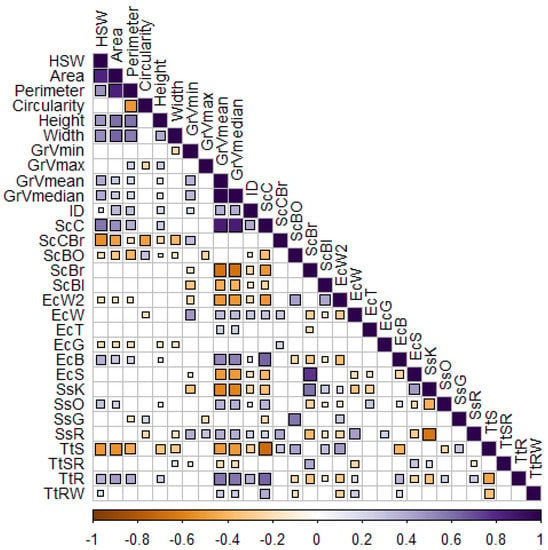
Figure 3.
Pearson’s corrplot of the seed morphological traits. Non-significant correlations were omitted from the plot (p > 0.05). Seed trait abbreviations are explained in the Abbreviations.
3.2. Molecular Genetic Analysis
Genotyping of the eight cowpea metapopulations with 14 polymorphic microsatellites generated, in total, 78 different alleles. The number of alleles per SSR primer set ranged from 2 (VM22, VM36, VM39, SSR1, SSR6421, SSR6243) to 16 (SSR6239), with an average of 5.5. The polymorphic information content (PIC), a measure indicative of the allelic diversity for a specific locus, varied from 0.1 (SSR6819) to 0.8 (SSR6807), with an average of 0.35 (Supplementary Table S4). The microsatellite loci that exhibited the highest PIC values were SSR6807, SSR6239 and SSR6866, indicating their usefulness in discriminating different genotypes. A summary of the molecular diversity components for the eight cowpea metapopulations is provided in Table 2.
Table 2.
Summary of the genetic diversity statistics over 14 SSR loci.
The Analysis of Molecular Variance (AMOVA) revealed that 93% of the observed variability was assigned within each cowpea metapopulation (Supplementary Table S5) and the UPGMA arranged the metapopulations into two distinct clusters (Figure 4a,b). Cluster I is comprised of the landraces that originate in Dodecanese islands, whereas cluster II is formed by the rest of the metapopulations.
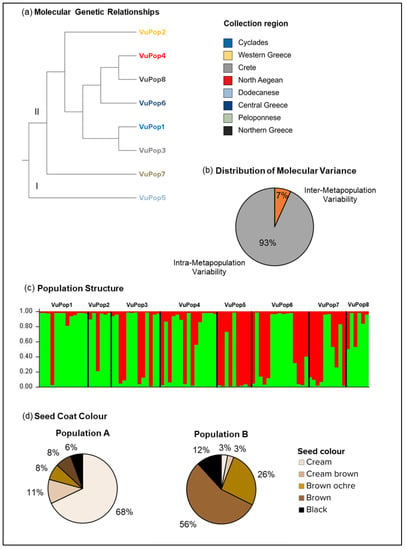
Figure 4.
(a) SSR-generated dendrogram of the 8 cowpea metapopulations, distributing the landraces in two main clusters; (b) distribution of variance according to AMOVA; (c) population structure graph of the estimated membership of the 87 cowpea landraces, based on 14 SSR markers (K = 2), indicating the emergence of two subpopulations; and (d) seed coat color distribution among the two populations.
According to the ΔΚ population structure analysis, the 87 accessions were grouped into two assumed populations (A, B) (Figure 4c). Population A comprises the majority of the accessions (61%), while population B gathered 39% of the accessions (likelihood threshold 0.55). Both populations were mixed in composition, in that they included accessions from all regional territories. The majority of the accessions from Cyclades (93.2%), Crete (60%), Western Greece (83,3%), North Aegean (69.5%) and Northern Greece (71.4%) are arranged in population A, whereas the majority of the accessions from the Dodecanese (77.7%) and Peloponnese (66.6%) are grouped in population B. The accessions from Central Greece are distributed equally in both populations. Regarding the seed coat color, population A comprises mostly cream/cream brown landraces (79%), while a smaller part concerns brown/brown ochre (16%) and black (6%) seeded landraces. Contrarily, population B includes almost exclusively dark seeded landraces (brown ochre to brown seed coat at 82%, black seed coat at 12%), and only 6% of the landraces are a cream-brown color (Figure 4d). We estimated the Fst values for both populations (FstA = 0.23, FstB = 0.16), deducing moderate differentiation levels of one another (Fstmean = 0.2), and calculated the average distances among the individuals in each population (HeA = 0.23, HeB = 0.26).
Molecular marker KAD-61 was successfully detected in all accessions; however, the amplicon size of 414 bp in all accessions was larger than suggested in the literature (up to 200 bp). Sequence analysis of the amplicon from all accessions revealed 95% coverage and 99.5% identity to the corresponding nucleotide sequence coding for the LRR receptor-like serine/threonine-protein kinase FLS2, which signals for plant innate immunity responses. However, due to the observed ambiguity in the amplicon size, the use of this marker requires further investigation to confirm, in vivo, the assessed genetic resistance in the given germplasm collection.
Further molecular screening of the germplasm for the presence of resistance-conferring alleles to Aphis craccivora (Koch) with the microsatellite CP-171-172 revealed that 12 accessions were associated with the resistant genotype (VuGR3, VuGR33, VuGR35, VuGR41, VuG47, VuGR49, VuGR56, VuGR76, VuGR77, VuGR78, VuGR85, VuGR86). However, this marker was detected only in 35 out of the 87 accessions.
4. Discussion
Orphan crops such as cowpea are vital in reassuring food security in vulnerable regions worldwide, underlining their continuous cultivation and untapped diversity. Despite their importance to agriculture, only recently has the species gained breeders’ attention, in order to increase plant-protein production to meet food crisis challenges due to climate change [46]. Cultivated cowpea’s germplasm genetic base is narrow because of its highly self-pollinating nature and to precedent breeding programs, as improved elite lines were constantly used as parents in crosses to generate segregating populations, exhibiting limited gene flow. On the other hand, the genetic diversity in the local germplasm remains untapped, constraining the effective incorporation of locally adapted landraces and alleles in breeding strategies.
Landraces are defined as “geographically or ecologically distinctive populations, which have evolved under cultivation and become locally adapted to various environments” [47]. Contrary to modern varieties, landraces have not been exposed to systematic plant-breeding procedures and are diverse in their genetic composition, both within and between populations. They provide a broad representation of the natural variation that is present in a species and constitute the primary gene pool available to breeders for improvements in crops. Hence, landraces constitute a dynamic component in the cultivated species genetic diversity. Screening of eight Greek metapopulations, composed of 87 landraces with 14 microsatellites, revealed a range of 2 to 16 alleles per locus. Among others, some of those molecular markers were previously employed for genotyping a plethora of cowpea cultivars, breeding lines, landraces and wild relatives, and reported similar [21,48,49,50] and more limited allele ranges [16,19,51]. The number of alleles per locus is highly associated with the number of accessions subjected to SSR genotyping, the selected markers and the type of the germplasm, and is indicative of the polymorphic content of the germplasm under evaluation. Allele ranges similar to our results were previously detected in up to 7-fold more multitudinous study populations of African origin [49]. It is, therefore, deducible that the accessions that were evaluated in the present study hold a significant amount of untapped diversity, which should be taken into consideration for incorporation in breeding strategies seeking the expansion of the species germplasm genetic base.
PIC values and gene diversity (also called expected heterozygosity) are both estimates of the genetic polymorphism present in populations and are used for the assessment of the quality of a marker [52]. The PIC values represent the informativeness of molecular markers, which is categorized as low (PIC < 0.25), moderate (0.25 < PIC < 0.5) and high (PIC > 0.5), respectively. In this study, we used 14 SSR markers to evaluate the genetic diversity of 87 cowpea accessions. Most SSRs exhibited moderate to high PIC values, ranging from 0.1 to 0.8, with an average of 0.35 (Supplementary Table S4). The low PIC values observed in some of the SSRs are explained by the frequencies of the alleles’ presence in those loci. Disproportionate frequency distribution of a locus’ alleles results in the generation of low PIC values (e.g., VM36, SSR6819, SSR6215, SSR6243) (Supplementary Table S3). However, most of the microsatellites used in our study detected a great number of alleles in the different metapopulations, indicating the existence of a highly polymorphic germplasm, and also revealing highly variable loci (SSR6807, SSR6866 and SSR6239 detected 11, 12 and 16 alleles, respectively). Additionally, the comparatively low levels of gene diversity observed in our study (0.2) could be explained by the predominant self-pollinating nature of cowpea. The cowpea’s genetic base is considered narrow and the inherent self-pollination mechanism further contributes to its maintenance. However, landraces from the Mediterranean basin have demonstrated significant diversity [6,19,28,53,54]. Noteworthily, the metapopulations collected in the Aegean islands (Cyclades, Dodecanese and North Aegean) hold the highest amounts of polymorphism among the germplasm under evaluation (73–82%). This is most likely attributed to the geographical position of the Aegean islands. Closely neighboring both Europe and Asia, these insular complexes have served as intermediate docking stations for maritime traders for millennia [55]. As previously proposed, cowpea was probably transferred through Asia minor, constantly enriching the islands’ gene pool with new alleles, forming a crucible cowpea germplasm [3]. The AMOVA proposed that the detected variability is distributed within each metapopulation by 93%. This probably highlights the allocation of several accessions with preferable traits in various parts of Greece, through trading networks, and hybridization with local germplasm, subsequently giving formation to variable cowpea germplasms throughout the different regions [56]. In support of this claim, Xiong et al. (2016), having performed a robust SNP genotyping analysis on a world cowpea collection from 56 countries, reported that the majority of the diversity exists within given geographic regions, rather than among the different regions [57]. Moreover, agriculture in the Aegean islands is confined in small terrace fields that are scattered amid steep slopes [58]. Considering the prevalent self-pollinating nature of cowpea, the emergence of isolated genetic pools is likely.
To further investigate the heterogeneity among different metapopulations, analysis of population structure presumed the emergence of two groups, where most of the accessions from Cyclades, Crete, Western Greece, North Aegean and Northern Greece are arranged in one homogeneous population and the majority of the accessions from the Dodecanese and Peloponnese in another, making evident the germplasm expansion throughout Greece, the potential homogeneity of landraces of distant territories and the heterogeneity in germplasms cultivated in neighboring regions (Cyclades–Dodecanese). Interestingly, the accessions from the Dodecanese islands seem more distinct from the rest of the accessions (Figure 4a). Central Greece is a nodal champaign in the middle of Greece concentrated with intensive agricultural production, characterized by a variety of microclimatic conditions between the northern–southern and the eastern–western parts of its territory. Based on the structure analysis results, accessions that originate from Central Greece were equally divided in both the aforementioned populations, suggesting a diverse cowpea germplasm hotspot.
Legumes, as a milestone staple in the Mediterranean diet, are strongly connected to the socio-economic profile of a region [59]. Different cowpea productive regions cultivate certain varieties more than others, in terms of consumer-desired seed traits, such as the coat color and pattern, features determinant of quality and use. Usually, light-colored seeds are destined for flour, while darker seeds are used as whole beans. Every year, Greek islands are visited by a vast number of tourists, of varicolored gastronomical backgrounds, hence, determining the culinary profile of these regions and, consequently, defining the regional agriculture [60]. In the Dodecanese islands, Crete and Peloponnese, most of the accessions are dark-seeded (Figure 5). Dark-seeded varieties were found to be more resilient under water deficit and extreme heat conditions, with little chlorophyl reduction and high levels of proline, an osmolyte compound, contributing to the cells’ osmotic adjustment and stabilization of subcellular compartments [61]. Therefore, potential selection of adaptive accessions with dark seed coats would justify the cultivation of mostly brown-seeded landraces, as in these areas, irrigated lands are limited and are further decreased as the regional climate is characterized by increased temperatures, which will further rise due to climate change [62].
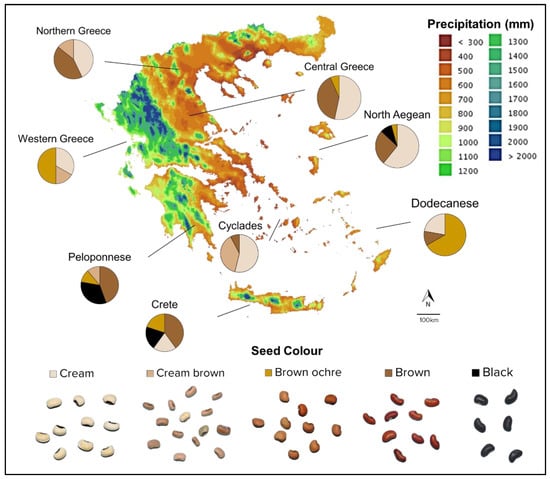
Figure 5.
Map of Greece showing average annual precipitation (mm) and the composition of the different cowpea populations regarding their seed color (http://climatlas.hnms.gr/, accessed on 30 September 2022).
On the other hand, every year from mid-May to mid-September, the North Aegean and Cyclades receive strong Etesian northern winds, also known as “meltemia”, which contribute to the establishment of milder summer temperatures, benefiting completion of seed filling and protecting from abrupt temperature increases, compared to the mainland and the Dodecanese [63]. Taking into account that cowpea is a warm season crop and harvesting occurs in early summer, this phenomenon buttresses the cultivation of white-seeded cowpea accessions in those regions.
Dark seed color is also strongly associated with overaccumulation of several secondary metabolites, such as flavanols (catechin, catechin glucoside and epicatechin) and anthocyanins (delphinidin), compounds of pronounced nutraceutical, pharmaceutical and medicinal interest [64,65,66]. Apart from that, the seeds’ composition on functional proteins justifies the need to transform cowpea from cash crop to industrial crop [67]. Protein content in cowpea is reported to reach up to 40% of the seed dry weight [68] and is found to be most abundant in cream-colored varieties [69]. The seed color and pattern along with seed size are determinants of the cowpea’s market price and have been considered as major breeding targets [70]. Cowpea is characterized by high intra-plant variability regarding the seed weight and protein content, which is attributed to indeterminate flowering and pod position. Development of high-yielding, uniformly flowering cultivars, with constant numbers of seeds per pod would increase the seed size and protein content homogeneity and would promote the industrialization of the crop [71]. Many Greek accessions examined in this study are considered highly productive, as they present HSW values up to 31 g (VuGR23), higher than in previously described commercial and local germplasms from Africa and India, indicating their potential for further breeding into elite cultivars [72,73,74,75,76]. Similar results were reported in Southern European landraces from Italy and the Iberian peninsula that further support the claim of germplasm migration inside the Mediterranean basin through trade from Asia minor [29]. The landraces collected in the Northern Greek territories demonstrate slightly higher values regarding the overall seed size and weight. Both features are considered significantly heritable and this discrimination may be attributed to the more favorable average climatic conditions in northern regions, where the crop can receive adequate precipitation and irrigation, promoting sufficient seed filling prior harvest, in contrast to the southern territories, which are more drought-prone. Moreover, Central and Northern Greece are run by several streams, rivers and aquifers, and intensive irrigated agriculture allows for safeguarding production that would otherwise be lost in non-irrigated cultivation schemes, under drought waves.
Cowpea, being mostly cultivated in semiarid areas, is well adapted to abiotic and biotic stresses. However, infestation with Aphis craccivora is challenging for the crop and may result in major yield losses. Cowpea aphid is phytotoxic and damages the crop, regardless of pest densities. Along with the direct damage caused by the sap imbibition, A. craccivora is responsible for the transmission of more than 14 viruses to cowpea and other pulses, rendering the need for successful aphid population management in cowpea cultivation systems imperative [77]. Despite the use of insecticides, cultural and integrated pest management for controlling aphid-induced damage to crops, methods have not been very effective, as smallholder farmers growing cowpea in marginal areas cannot afford their cost, in addition questioning their environmental footprint. Thus, development of cultivars with enhanced endogenous resistance is the most efficient and sustainable alternative to chemical pest control, sustaining crop production for longer periods. Conducting resistance bioassays in large breeding populations is a time-consuming and wearisome procedure. Development of molecular markers perfectly linked to the resistant phenotype would promote rapid selection of the preferred genotypes. Thus far, efforts have been made in detecting resistant genotypes and elucidating the molecular mechanism underlying aphid resistance [77]. Antibiosis is proposed as the principal defense response of the African-resistant germplasm, in blocking sap ingestion by cowpea aphids, secondarily supported by antixenosis mechanisms [78,79,80]. Investigation of the genetic mechanism regulating aphid resistance in TVu-2876 indicated the action of a single dominant gene [36]. Molecular screening of the Greek germplasm revealed the presence of the resistant TVu-2876 genotype in 12 landraces, highlighting a promising source of aphid genetic resistance that needs to be further examined. The majority of them originated in Peloponnese, Central Greece and North Aegean regions. Out of them, seven are dark-seeded, while five of them are cream black-eyed seeds. These landraces could be incorporated in gene pyramiding assays in cowpea breeding strategies. Dark seed coats are rich in secondary metabolites that contribute to defense responses against biotic stresses [81]. Varieties with a high abundance of seed secondary metabolites are predisposed to biosynthesize high amounts of such compounds when needed. Previously, positive correlation has been detected between the plant flavonoid content and abundance and the resistance to aphids. More specifically, kaempferol, quercetin and isorhamnetin cowpea chemotypes were associated with resistance responses, with quercetin and isorhamnetin demonstrating a direct involvement in the resistance mechanism [82,83]. Sequencing and alignment of the KAD-61 amplicon to the V. unguiculata genome detected absolute homology to the leucine-rich repeat (LRR) receptor-like serine/threonine-protein kinase (RLK) FLS2 gene of V. unguiculata ssp. sesquipedialis cultivar Xiabao 2 genome [84]. The transmembrane receptor kinase FLAGELLIN SENSITIVE2 (FLS2) is essential for flagellin perception and is integral in plant defense responses against pathogens [85]. With respect to the germplasm under evaluation, no nucleotide sequence variations were detected among the cowpea landraces in the aforementioned locus. Moreover, the amplicon size of KAD-61 was the same among all landraces, divergent to that proposed in the literature. Hence, KAD-61 is considered unreliable for discriminating among resistance and susceptibility conferring alleles in the given accessions. More profound genomic analyses are needed for the detection of resistant alleles in the present germplasm and the development of linked molecular markers that would accelerate selection for aphid resistance. Further research is also suggested regarding the in vivo demonstration of aphid resistance in VuGR3, VuGR33, VuGR35, VuGR41, VuG47, VuGR49, VuGR56, VuGR76, VuGR77, VuGR78, VuGR85 and VuGR86, as depicted, presaged by marker CP-171-172 to harbor the resistant TVu-2876 genotype. Contemporary biotechnological approaches providing insect and pathogen resistance through molecular network engineering have already been successfully employed in cowpea and would assist cowpea breeding for increased plasticity, subsequently promoting cowpea productivity [86].
5. Conclusions
Cowpea is considered a staple, resilient and underutilized legume, cultivated in low-input agricultural systems and a promising industrial protein-rich crop. Cowpea experienced two bottleneck effects during domestication and the breeders’ tendency of using improved elite lines as parental material for the generation of segregating populations in breeding programs resulted in limiting the crop’s genetic base. Therefore, scrutinization of unexploited local germplasms is substantial for species gene pool enrichment. Significant polymorphism was detected among 87 Greek landraces, regarding seed morphological characteristics, seed color and molecular genetic diversity. Many of them are considered high-producing, when compared to other cultivated accessions, and 12 of them possess the A. craccivora resistance-conferring CP-171-172 allele, which would be highly favorable for incorporation in cowpea breeding programs, towards pyramiding aphid resistance to elite germplasms. Furthermore, future genome-wide association studies are proposed for the germplasm evaluated in this study, for the detection of desirable alleles regarding biotic and abiotic stress resilience and their potential introgression to elite germplasms, as climate change imposes the development of adaptive cultivars for safeguarding plant–protein production in vulnerable Mediterranean agro-ecosystems.
Supplementary Materials
The following are available online at https://www.mdpi.com/article/10.3390/agronomy13010274/s1, Table S1. Details on the collection sites, seed color and eye color of the V. unguiculata landraces. Table S2. SSR primer sequences used in the study. Table S3. SSR scoring binary matrix, where “1” indicates the presence and “0” indicates the absence of a specific allele. Table S4. Polymorphic Information Content (PIC) value for the SSR markers used for genotyping 87 cowpea landraces. Table S5. AMOVA results based on genotypic analysis with 14 SSR molecular markers.
Author Contributions
Conceptualization, P.V.M.; methodology, I.Z., M.S.; formal analysis, I.Z. and M.S.; investigation, I.Z., M.S. and P.V.M.; resources, P.V.M.; writing—original draft preparation, I.Z. writing—review and editing, I.Z. and P.V.M.; project administration, P.V.M.; funding acquisition, P.V.M. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
Abbreviations
| ScC | Seed color Cream |
| ScCBr | Seed color Cream-Brown |
| ScBO | Seed color Brown-Ochre |
| ScBr | Seed color Brown |
| ScBl | Seed color Black |
| EcW2 | Eye color W2 |
| EcW | Eye color W |
| EcT | Eye color T |
| EcG | Eye color G |
| EcB | Eye color B |
| EcS | Eye color S |
| SsK | Seed shape Kidney |
| SsO | Seed shape Ovoid |
| SsG | Seed shape Globose |
| SsR | Seed shape Rhomboid |
| TtS | Testa texture Smooth |
| TtSR | Testa texture Smooth to Rough |
| TtR | Testa texture Rough |
| TtRW | Testa texture Rough to Wrinkled |
References
- Bundervoet, T.; Dávalos, M.E.; Garcia, N. The Short-Term Impacts of COVID-19 on Households in Developing Countries; Policy Research Working Paper; No. 9582; World Bank: Washington, DC, USA, 2021; Available online: https://hdl.handle.net/10986/35290 (accessed on 25 September 2022).
- Jimenez-Lopez, J.C.; Singh, K.B.; Clemente, A.; Nelson, M.N.; Ochatt, S.; Smith, P.M.C.C. Editorial: Legumes for Global Food Security. Frontiers 2020, 11, 926. [Google Scholar] [CrossRef]
- Herniter, I.A.; Muñoz-Amatriaín, M.; Close, T.J.; Muñoz-Amatriaín, M.; Close, T.J. Genetic, textual, and archeological evidence of the historical global spread of cowpea (Vigna unguiculata [L.] Walp.). Legum. Sci. 2020, 2, e57. [Google Scholar] [CrossRef]
- Carvalho, M.; Lino-Neto, T.; Rosa, E.; Carnide, V. Cowpea: A legume crop for a challenging environment. J. Sci. Food Agric. 2017, 97, 4273–4284. [Google Scholar] [CrossRef] [PubMed]
- De Luca, D.; Cennamo, P.; Del Guacchio, E.; Di Novella, R.; Caputo, P. Conservation and genetic characterisation of common bean landraces from Cilento region (Southern Italy): High differentiation in spite of low genetic diversity. Genetica 2018, 146, 29–44. [Google Scholar] [CrossRef] [PubMed]
- Lazaridi, E.; Ntatsi, G.; Fernández, J.A.; Karapanos, I.; Carnide, V.; Savvas, D.; Bebeli, P.J. Phenotypic diversity and evaluation of fresh pods of cowpea landraces from Southern Europe. J. Sci. Food Agric. 2017, 97, 4326–4333. [Google Scholar] [CrossRef] [PubMed]
- Anago, F.N.; Agbangba, E.C.; Oussou, B.T.C.; Dagbenonbakin, G.D.; Amadji, L.G. Cultivation of cowpea challenges in west africa for food security: Analysis of factors driving yield gap in Benin. Agronomy 2021, 11, 1139. [Google Scholar] [CrossRef]
- Karuwal, R.L.; Suharsono, S.; Tjahjoleksono, A.; Hanif, N. Short Communication: Characterization and nutrient analysis of seed of local cowpea (Vigna unguiculata) varieties from Southwest Maluku, Indonesia. Biodiversitas 2021, 22, 85–91. [Google Scholar] [CrossRef]
- Jayathilake, C.; Visvanathan, R.; Deen, A.; Bangamuwage, R.; Jayawardana, B.C.; Nammi, S.; Liyanage, R. Cowpea: An overview on its nutritional facts and health benefits. J. Sci. Food Agric. 2018, 98, 4793–4806. [Google Scholar] [CrossRef]
- Baoua, I.; Rabé, M.M.; Murdock, L.L.; Baributsa, D. Cowpea production constraints on smallholders’ farms in Maradi and Zinder regions, Niger. Crop Prot. 2021, 142, 105533. [Google Scholar] [CrossRef]
- Kamara, A.Y.; Omoigui, L.O.; Kamai, N.; Ewansiha, S.U.; Ajeigbe, H.A. Improving cultivation of cowpea in West Africa. In Achieving Sustainable Cultivation of Grain Legumes; Burleigh Dodds Science Publishing: Cambridge, UK, 2018; Volume 2, pp. 235–252. ISBN 9781786761408. [Google Scholar]
- FAOSTAT. Available online: http://www.fao.org/faostat/en/#data/QCL/visualize (accessed on 25 September 2022).
- Ibrahim, M.; David, A.A.; Mshelia, J.S.; John, A.B.; Sunday, B.A.; Stephanie, S.B.-U. Effect of climate variation on the yield of cowpea (Vigna unguiculata). Afr. J. Agric. Res. 2021, 17, 456–462. [Google Scholar] [CrossRef]
- Busconi, M.; Gumede, M.T.; Shegro Gerrano, A.; Amelework, A.B.; Modi, A.T. Analysis of Genetic Diversity and Population Structure of Cowpea (Vigna unguiculata (L.) Walp) Genotypes Using Single Nucleotide Polymorphism Markers. Plants 2022, 11, 3480. [Google Scholar] [CrossRef]
- Dagnon, Y.D.; Palanga, K.K.; Bammite, D.; Bodian, A.; Akabassi, G.C.; Foncéka, D.; Tozo, K. Genetic diversity and population structure of cowpea [Vigna unguiculata (L.) Walp.] accessions from Togo using SSR markers. PLoS ONE 2022, 17, e0252362. [Google Scholar] [CrossRef] [PubMed]
- Diouf, D.; Hilu, K.W. Microsatellites and RAPD markers to study genetic relationships among cowpea breeding lines and local varieties in Senegal. Genet. Resour. Crop Evol. 2005, 52, 1057–1067. [Google Scholar] [CrossRef]
- Fang, J.; Chao, C.C.T.; Roberts, P.A.; Ehlers, J.D. Genetic diversity of cowpea [Vigna unguiculata (L.) Walp.] in four West African and USA breeding programs as determined by AFLP analysis. Genet. Resour. Crop Evol. 2007, 54, 1197–1209. [Google Scholar] [CrossRef]
- Ghalmi, N.; Malice, M.; Jacquemin, J.M.; Ounane, S.M.; Mekliche, L.; Baudoin, J.P. Morphological and molecular diversity within Algerian cowpea (Vigna unguiculata (L.) Walp.) landraces. Genet. Resour. Crop Evol. 2010, 57, 371–386. [Google Scholar] [CrossRef]
- Asare, A.T.; Gowda, B.S.; Galyuon, I.K.A.; Aboagye, L.L.; Takrama, J.F.; Timko, M.P. Assessment of the genetic diversity in cowpea (Vigna unguiculata L. Walp.) germplasm from Ghana using simple sequence repeat markers. Plant Genet. Resour. 2010, 8, 142–150. [Google Scholar] [CrossRef]
- Li, C.D.; Fatokun, C.A.; Ubi, B.; Singh, B.B.; Scoles, G.J. Determining genetic similarities and relationships among cowpea breeding lines and cultivars by microsatellite markers. Crop Sci. 2001, 41, 189–197. [Google Scholar] [CrossRef]
- Badiane, F.A.; Gowda, B.S.; Cissé, N.; Diouf, D.; Sadio, O.; Timko, M.P. Genetic relationship of cowpea (Vigna unguiculata) varieties from Senegal based on SSR markers. Genet. Mol. Res. 2012, 11, 292–304. [Google Scholar] [CrossRef]
- Skendžić, S.; Zovko, M.; Živković, I.P.; Lešić, V.; Lemić, D. The impact of climate change on agricultural insect pests. Insects 2021, 12, 440. [Google Scholar] [CrossRef]
- Mahesha, H.S.; Keerthi, M.C.; Shivakumar, K.V.; Bhargavi, H.A.; Saini, R.P.; Manjunatha, L.; Hickok, D.; Blair, M.W. Development of Biotic Stress Resistant Cowpea. In Genomic Designing for Biotic Stress Resistant Pulse Crops; Springer: Cham, Switzerland, 2022; pp. 213–251. [Google Scholar]
- Bhusal, S.J.; Koch, R.L.; Lorenz, A.J. Variation in Soybean Aphid (Hemiptera: Aphididae) Biotypes within Fields. J. Econ. Entomol. 2021, 114, 1336–1344. [Google Scholar] [CrossRef]
- Singh, K.S.; Cordeiro, E.M.G.; Troczka, B.J.; Pym, A.; Mackisack, J.; Mathers, T.C.; Duarte, A.; Legeai, F.; Robin, S.; Bielza, P.; et al. Global patterns in genomic diversity underpinning the evolution of insecticide resistance in the aphid crop pest Myzus persicae. Commun. Biol. 2021, 4, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Bellucci, E.; Bitocchi, E.; Rau, D.; Nanni, L.; Ferradini, N.; Giardini, A.; Rodriguez, M.; Attene, G.; Papa, R. Population structure of barley landrace populations and gene-flow with modern varieties. PLoS ONE 2013, 8, e83891. [Google Scholar] [CrossRef] [PubMed]
- Negri, V. Agro-biodiversity conservation in Europe: Ethical issues. J. Agric. Environ. Ethics 2005, 18, 3–25. [Google Scholar] [CrossRef]
- Lazaridi, E.; Ntatsi, G.; Savvas, D.; Bebeli, P.J. Diversity in cowpea (Vigna unguiculata (L.) Walp.) local populations from Greece. Genet. Resour. Crop Evol. 2017, 64, 1529–1551. [Google Scholar] [CrossRef]
- Monteiro, E.; Castro, I.; Carvalho, M.; Martín, J.P.; Rosa, E.; Carnide, V. Iberian Peninsula cowpea diversity: Chloroplast, microsatellite and morpho-agronomic variability. Syst. Biodivers. 2020, 19, 121–134. [Google Scholar] [CrossRef]
- Muñoz-Amatriaín, M.; Lo, S.; Herniter, I.A.; Boukar, O.; Fatokun, C.; Carvalho, M.; Castro, I.; Guo, Y.N.; Huynh, B.L.; Roberts, P.A.; et al. The UCR Minicore: A resource for cowpea research and breeding. Legum. Sci. 2021, 3, e95. [Google Scholar] [CrossRef]
- Carvalho, M.; Muñoz-Amatriaín, M.; Castro, I.; Lino-Neto, T.; Matos, M.; Egea-Cortines, M.; Rosa, E.; Close, T.; Carnide, V. Genetic diversity and structure of Iberian Peninsula cowpeas compared to world-wide cowpea accessions using high density SNP markers. BMC Genom. 2017, 18, 121–134. [Google Scholar] [CrossRef]
- International Board for Plant Genetic Resources (IBPGR) Descriptors for Cowpea. Available online: https://www.bioversityinternational.org/e-library/publications/detail/descriptors-for-cowpea/ (accessed on 11 August 2022).
- Tsanakas, G.F.; Mylona, P.V.; Koura, K.; Gleridou, A.; Polidoros, A.N. Genetic diversity analysis of the Greek lentil (Lens culinaris) landrace “Eglouvis” using morphological and molecular markers. Plant Genet. Resour. Characterisation Util. 2018, 16, 469–477. [Google Scholar] [CrossRef]
- R Core Team. R Core Team 2021 R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021; Available online: https://www.R-project.org/ (accessed on 28 August 2022).
- Andargie, M.; Pasquet, R.S.; Gowda, B.S.; Muluvi, G.M.; Timko, M.P. Construction of a SSR-based genetic map and identification of QTL for domestication traits using recombinant inbred lines from a cross between wild and cultivated cowpea (V. unguiculata (L.) Walp.). Mol. Breed. 2011, 28, 413–420. [Google Scholar] [CrossRef]
- Omoigui, L.O.; Ekeuro, G.C.; Kamara, A.Y.; Bello, L.L.; Timko, M.P.; Ogunwolu, G.O. New sources of aphids [Aphis craccivora (Koch)] resistance in cowpea germplasm using phenotypic and molecular marker approaches. Euphytica 2017, 213, 178. [Google Scholar] [CrossRef]
- Goor, R.M.; Hoffman, D.; Riley, G.R. Novel Method for Accurately Assessing Pull-up Artifacts in STR Analysis. Forensic Sci. Int. Genet. 2021, 51, 102410. [Google Scholar] [CrossRef] [PubMed]
- Casañas, F.; Simó, J.; Casals, J.; Prohens, J. Toward an evolved concept of landrace. Front. Plant Sci. 2017, 8, 145. [Google Scholar] [CrossRef] [PubMed]
- Kyratzis, A.C.; Nikoloudakis, N.; Katsiotis, A. Genetic variability in landraces populations and the risk to lose genetic variation. The example of landrace “Kyperounda” and its implications for ex situ conservation. PLoS ONE 2019, 14, e0224255. [Google Scholar] [CrossRef] [PubMed]
- Peakall, R.; Smouse, P.E. GENALEX 6: Genetic analysis in Excel. Population genetic software for teaching and research. Mol. Ecol. Notes 2006, 6, 288–295. [Google Scholar] [CrossRef]
- Nei, M.; Tajima, F.; Tateno, Y. Accuracy of estimated phylogenetic trees from molecular data—II. Gene frequency data. J. Mol. Evol. 1983, 19, 153–170. [Google Scholar] [CrossRef]
- Smith, J.S.C.; Chin, E.C.L.; Shu, H.; Smith, O.S.; Wall, S.J.; Senior, M.L.; Mitchell, S.E.; Kresovich, S.; Ziegle, J. An evaluation of the utility of SSR loci as molecular markers in maize (Zea mays L.): Comparisons with data from RFLPS and pedigree. Theor. Appl. Genet. 1997, 95, 163–173. [Google Scholar] [CrossRef]
- Pritchard, J.K.; Stephens, M.; Donnelly, P. Inference of Population Structure Using Multilocus Genotype Data. Genetics 2000, 155, 945–959. [Google Scholar] [CrossRef]
- Evanno, G.; Regnaut, S.; Goudet, J. Detecting the number of clusters of individuals using the software structure: A simulation study. Mol. Ecol. 2005, 14, 2611–2620. [Google Scholar] [CrossRef]
- Earl, D.A.; vonHoldt, B.M. STRUCTURE HARVESTER: A website and program for visualizing STRUCTURE output and implementing the Evanno method. Conserv. Genet. Resour. 2011, 4, 359–361. [Google Scholar] [CrossRef]
- Tadele, Z. Orphan crops: Their importance and the urgency of improvement. Planta 2019, 250, 677–694. [Google Scholar] [CrossRef]
- Dwivedi, S.L.; Ceccarelli, S.; Blair, M.W.; Upadhyaya, H.D.; Are, A.K.; Ortiz, R. Landrace Germplasm for Improving Yield and Abiotic Stress Adaptation. Trends Plant Sci. 2016, 21, 31–42. [Google Scholar] [CrossRef] [PubMed]
- Feng, Y.; Ryan, U.M.; Xiao, L. Genetic Diversity and Population Structure of Cryptosporidium. Trends Parasitol. 2018, 34, 997–1011. [Google Scholar] [CrossRef] [PubMed]
- Sarr, A.; Bodian, A.; Gbedevi, K.M.; Ndir, K.N.; Ajewole, O.O.; Gueye, B.; Foncéka, D.; Diop, E.A.M.C.; Diop, B.M.; Cissé, N.; et al. Genetic Diversity and Population Structure Analyses of Wild Relatives and Cultivated Cowpea (Vigna unguiculata (L.) Walp.) from Senegal Using Simple Sequence Repeat Markers. Plant Mol. Biol. Rep. 2021, 39, 112–124. [Google Scholar] [CrossRef]
- Ali, Z.B.; Yao, K.N.; Odeny, D.A.; Kyalo, M.; Skilton, R.; Eltahir, I.M. Assessing the genetic diversity of cowpea [Vigna unguiculata (L.) Walp.] accessions from Sudan using simple sequence repeat (SSR) markers. Afr. J. Plant Sci. 2015, 9, 293–304. [Google Scholar] [CrossRef]
- Ouedraogo, J.; Sawadago, M.; Tignegre, J.-B.; Drabo, I.; Balma, D. Caractérisation agro-morphologique et moléculaire de cultivars locaux de niébé (Vigna unguiculata) du Burkina Faso. Cameroon J. Exp. Biol. 2010, 6, 31–40. [Google Scholar] [CrossRef]
- Serrote, C.M.L.; Reiniger, L.R.S.; Silva, K.B.; Dos Santos Rabaiolli, S.M.; Stefanel, C.M. Determining the Polymorphism Information Content of a molecular marker. Gene 2020, 726, 144175. [Google Scholar] [CrossRef]
- Bozokalfa, M.K.; Aşçioğul, T.K.; Eşiyok, D. Genetic diversity of farmer-preferred cowpea (Vigna unguiculata L. Walp) landraces in Turkey and evaluation of their relationships based on agromorphological traits. Genetika 2017, 49, 935–957. [Google Scholar] [CrossRef]
- Lioi, L.; Morgese, A.; Cifarelli, S.; Sonnante, G. Germplasm collection, genetic diversity and on-farm conservation of cowpea [Vigna unguiculata (L.) Walp.] landraces from Apulia region (Southern Italy). Genet. Resour. Crop Evol. 2019, 66, 165–175. [Google Scholar] [CrossRef]
- Horejs, B.; Milić, B.; Ostmann, F.; Thanheiser, U.; Weninger, B.; Galik, A. The Aegean in the Early 7th Millennium BC: Maritime Networks and Colonization. J. World Prehistory 2015, 28, 289–330. [Google Scholar] [CrossRef]
- Zeven, A.C. Landraces: A review of definitions and classifications. Euphytica 1998, 104, 127–139. [Google Scholar] [CrossRef]
- Xiong, H.; Shi, A.; Mou, B.; Qin, J.; Motes, D.; Lu, W.; Ma, J.; Weng, Y.; Yang, W.; Wu, D. Genetic Diversity and Population Structure of Cowpea (Vigna unguiculata L. Walp). PLoS ONE 2016, 11, e0160941. [Google Scholar] [CrossRef] [PubMed]
- Sakellariou, M.; Psiloglou, B.E.; Giannakopoulos, C.; Mylona, P.V. Integration of Abandoned Lands in Sustainable Agriculture: The Case of Terraced Landscape Re-Cultivation in Mediterranean Island Conditions. Land 2021, 10, 457. [Google Scholar] [CrossRef]
- Piergiovanni, A.R. Legumes: Staple foods used in rituals and festive events of Apulia region (Southern Italy). Food Cult. Soc. 2021, 24, 543–561. [Google Scholar] [CrossRef]
- Laeis, G.C.M.; Scheyvens, R.A.; Morris, C. Cuisine: A new concept for analysing tourism-agriculture linkages? J. Tour. Cult. Chang. 2020, 18, 643–658. [Google Scholar] [CrossRef]
- Bandurska, H.; Niedziela, J.; Pietrowska-Borek, M.; Nuc, K.; Chadzinikolau, T.; Radzikowska, D. Regulation of proline biosynthesis and resistance to drought stress in two barley (Hordeum vulgare L.) genotypes of different origin. Plant Physiol. Biochem. 2017, 118, 427–437. [Google Scholar] [CrossRef]
- Myronidis, D.; Nikolaos, T. Changes in climatic patterns and tourism and their concomitant effect on drinking water transfers into the region of South Aegean, Greece. Stoch. Environ. Res. Risk Assess. 2021, 35, 1725–1739. [Google Scholar] [CrossRef]
- Papadopoulos, A.; Varlas, G. Weather Systems Affecting the Meteorological Conditions over the Aegean Sea; Springer: Berlin/Heidelberg, Germany, 2020; pp. 1–25. [Google Scholar]
- Tsamo, A.T.; Mohammed, H.; Mohammed, M.; Papoh Ndibewu, P.; Dapare Dakora, F. Seed coat metabolite profiling of cowpea (Vigna unguiculata L. Walp.) accessions from Ghana using UPLC-PDA-QTOF-MS and chemometrics. Nat. Prod. Res. 2020, 34, 1158–1162. [Google Scholar] [CrossRef]
- Chen, Z.; Zhang, R.; Shi, W.; Li, L.; Liu, H.; Liu, Z.; Wu, L. The Multifunctional Benefits of Naturally Occurring Delphinidin and Its Glycosides. J. Agric. Food Chem. 2019, 67, 11288–11306. [Google Scholar] [CrossRef]
- Bae, J.; Kim, N.; Shin, Y.; Kim, S.-Y.; Kim, Y.-J. Activity of catechins and their applications. Biomed. Dermatol. 2020, 4, 1–10. [Google Scholar] [CrossRef]
- Carneiro da Silva, A.; de Freitas Barbosa, M.; Bento da Silva, P.; Peres de Oliveira, J.; Loureiro da Silva, T.; Lopes Teixeira Junior, D.; de Moura Rocha, M. Health Benefits and Industrial Applications of Functional Cowpea Seed Proteins. In Grain and Seed Proteins Functionality; IntechOpen: London, UK, 2021. [Google Scholar] [CrossRef]
- Dakora, F.D.; Belane, A.K. Evaluation of Protein and Micronutrient Levels in Edible Cowpea (Vigna unguiculata L. Walp.) Leaves and Seeds. Front. Sustain. Food Syst. 2019, 3, 70. [Google Scholar] [CrossRef]
- Weng, Y.; Qin, J.; Eaton, S.; Yang, Y.; Ravelombola, W.S.; Shi, A. Evaluation of Seed Protein Content in USDA Cowpea Germplasm. HortScience 2019, 54, 814–817. [Google Scholar] [CrossRef]
- Lo, S.; Muñoz-Amatriaín, M.; Boukar, O.; Herniter, I.; Cisse, N.; Guo, Y.N.; Roberts, P.A.; Xu, S.; Fatokun, C.; Close, T.J. Identification of QTL controlling domestication-related traits in cowpea (Vigna unguiculata L. Walp). Sci. Rep. 2018, 8, 6261. [Google Scholar] [CrossRef] [PubMed]
- Iseki, K.; Olaleye, O.; Ishikawa, H. Intra-plant variation in seed weight and seed protein content of cowpea. Plant Prod. Sci. 2020, 23, 103–113. [Google Scholar] [CrossRef]
- Nalawade, A.D.; Patil, S.M.; Rajwade, P.R.; Kauthale, V.K. Evaluation of Cowpea Germplasm by using Agro-Morphological Characters. Indian J. Agric. Res. 2021, 55, 364–368. [Google Scholar] [CrossRef]
- Gondwe, T.M.; Alamu, E.O.; Mdziniso, P.; Maziya-Dixon, B. Cowpea (Vigna unguiculata (L.) Walp) for food security: An evaluation of end-user traits of improved varieties in Swaziland. Sci. Rep. 2019, 9, 15991. [Google Scholar] [CrossRef]
- Mofokeng, M.A.; Mashilo, J.; Rantso, P.; Shimelis, H. Genetic variation and genetic advance in cowpea based on yield and yield-related traits. Acta Agric. Scand. 2020, 70, 381–391. [Google Scholar] [CrossRef]
- Gerrano, A.S.; Jansen van Rensburg, W.S.; Kutu, F.R. Agronomic evaluation and identification of potential cowpea (Vigna unguiculata L. Walp) genotypes in South Africa. Acta Agric. Scand. 2019, 69, 295–303. [Google Scholar] [CrossRef]
- Iseki, K.; Ikazaki, K.; Batieno, J.B. Cowpea yield variation in three dominant soil types in the Sudan Savanna of West Africa. Field Crops Res. 2021, 261, 108012. [Google Scholar] [CrossRef]
- Mofokeng, M.A.; Gerrano, A.S. Efforts in Breeding Cowpea for Aphid Resistance: A Review; Taylor & Francis: Abingdon, UK, 2021; Volume 71, pp. 489–497. [Google Scholar]
- MacWilliams, J.R.; Chesnais, Q.; Nabity, P.; Mauck, K.; Kaloshian, I. Cowpea aphid resistance in cowpea line CB77 functions primarily through antibiosis and eliminates phytotoxic symptoms of aphid feeding. J. Pest Sci. 2022, 1–15. [Google Scholar] [CrossRef]
- Kamphuis, L.G.; Gao, L.; Singh, K.B. Identification and characterization of resistance to cowpea aphid (Aphis craccivora Koch) in Medicago truncatula. BMC Plant Biol. 2012, 12, 101. [Google Scholar] [CrossRef]
- Huynh, B.L.; Ehlers, J.D.; Ndeve, A.; Wanamaker, S.; Lucas, M.R.; Close, T.J.; Roberts, P.A. Genetic mapping and legume synteny of aphid resistance in African cowpea (Vigna unguiculata L. Walp.) grown in California. Mol. Breed. 2015, 35, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Kpoviessi, A.D.; Agbahoungba, S.; Agoyi, E.E.; Nuwamanya, E.; Assogbadjo, A.E.; Chougourou, D.C.; Adoukonou-Sagbadja, H. Primary and secondary metabolite compounds in cowpea seeds resistant to the cowpea bruchid [Callosobruchus maculatus (F.)] in postharvest storage. J. Stored Prod. Res. 2021, 93, 101858. [Google Scholar] [CrossRef]
- Lattanzio, V.; Arpaia, S.; Cardinali, A.; Di Venere, D.; Linsalata, V. Endogenous flavonoids in resistance mechanism of Vigna role of to aphids. J. Agric. Food Chem. 2000, 48, 5316–5320. [Google Scholar] [CrossRef] [PubMed]
- Togola, A.; Boukar, O.; Servent, A.; Chamarthi, S.; Tamò, M.; Fatokun, C. Identification of sources of resistance in cowpea mini core accessions to Aphis craccivora Koch (Homoptera: Aphididae) and their biochemical characterization. Euphytica 2020, 216, 88. [Google Scholar] [CrossRef]
- Xia, Q.; Pan, L.; Zhang, R.; Ni, X.; Wang, Y.; Dong, X.; Gao, Y.; Zhang, Z.; Kui, L.; Li, Y.; et al. The genome assembly of asparagus bean, Vigna unguiculata ssp. sesquipedialis. Sci. Data 2019, 6, 124. [Google Scholar] [CrossRef] [PubMed]
- Liang, X.; Zhang, J. Regulation of plant responses to biotic and abiotic stress by receptor-like cytoplasmic kinases. Stress Biol. 2022, 2, 25. [Google Scholar] [CrossRef]
- Muthuvel, J.; Saharia, M.; Kumar, S.; Abiala, M.A.; Rao, G.J.N.; Sahoo, L. Progress in Genetic Engineering of Cowpea for Insect Pest and Virus Resistance. In Genetically Modified Crops; Springer: Singapore, 2021; pp. 115–137. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).