
Analysis of RAZORMIN® as a Biostimulant and Its Effect on the Phytotoxicity Mitigation Caused by Fungicide Azoxystrobin in Pepper

 , , ,
, , ,  , , and
, , and
Abstract
:1. Introduction
- i.
- inducer/promoter of metabolic, photosynthetic, and nutritional efficiency in plants (biostimulant effect);
- ii.
- inducer/promoter of plant defense responses against fungicide application (abiotic stress tolerance).
2. Materials and Methods
2.1. Plant Material and Growth Conditions
2.2. RAZORMIN®
2.3. Experiments and Treatments
2.3.1. Experiment 1: Efficacy of RAZORMIN® as a Biostimulant
2.3.2. Experiment 2: Efficacy of RAZORMIN® in Reducing the Phytotoxicity of the Fungicide Azoxystrobin
2.4. Plant Sampling
2.5. Relative Growth Rate (RGR)
2.6. Analysis of Photosynthetic Efficiency
2.7. Determination of Hormone Profile
2.8. Determination of Amino Acids and Soluble Proteins
2.9. Nutrient Content and Efficiency of Nutrient Utilization
2.10. Fluorescence Analysis of Chlorophyll a
2.11. Photosynthetic Pigments Concentration
2.12. Determination of the Concentration of Oxidative Stress Indicators (MDA, H2O2 and O2−)
2.13. Determination of the Concentration of Total Phenols, Flavonoids and Anthocyanins
2.14. Statistical Analysis
3. Results and Discussion
3.1. Experiment 1: Efficacy of RAZORMIN® as a Biostimulant
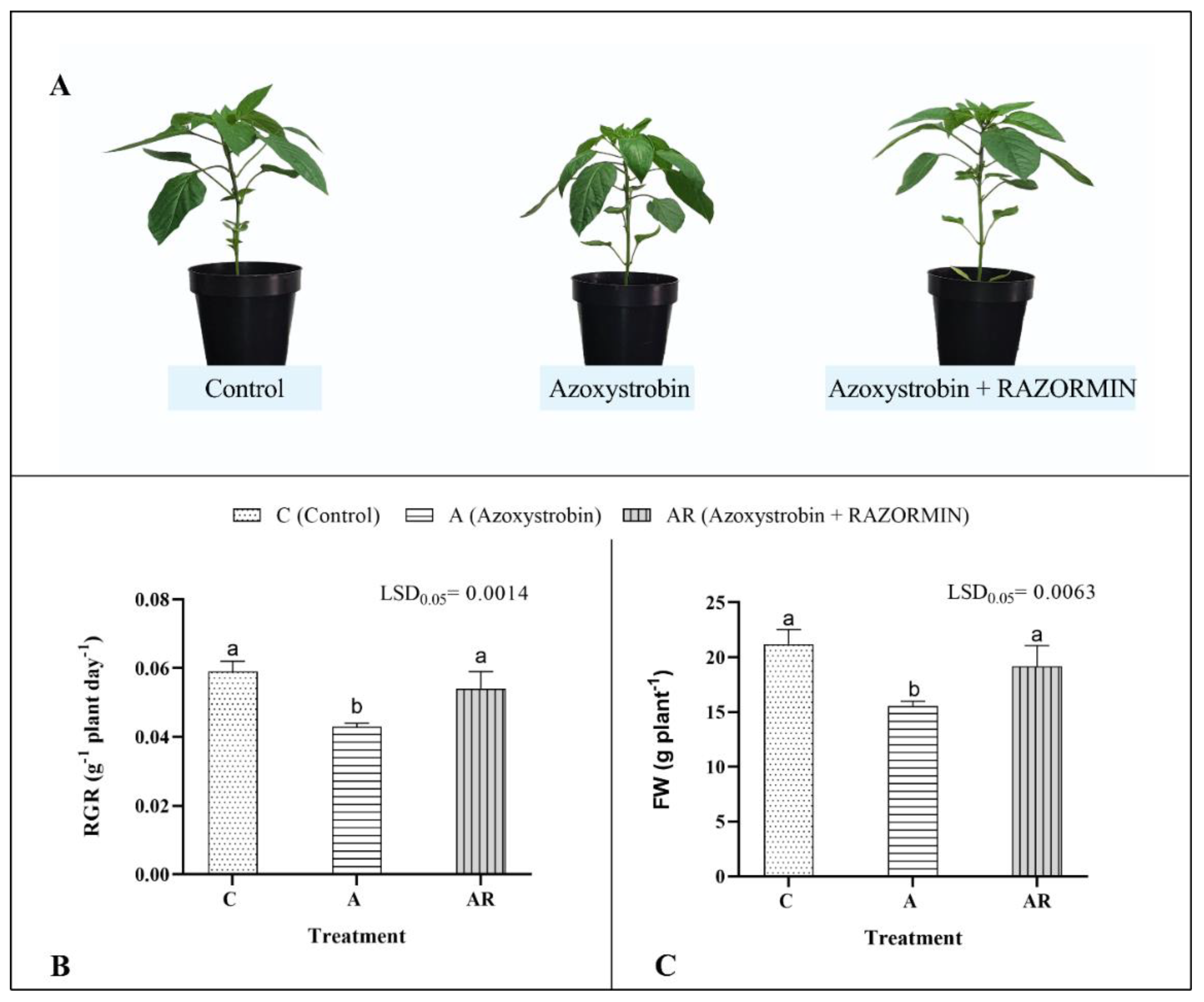
3.2. Experiment 2: Efficacy of RAZORMIN® as a Biostimulant Reducing the Phytotoxicity of the Fungicide Azoxystrobin
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Brown, P.; Saa, S. Biostimulants in agriculture. Front. Plant Sci. 2015, 6, 671. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yakhin, O.I.; Lubyanov, A.A.; Yakhin, I.A.; Brown, P.H. Biostimulants in Plant Science: A Global Perspective. Front. Plant Sci. 2017, 7, 2049. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zulfiqar, F.; Casadesús, A.; Brockman, H.; Munné-Bosch, S. An overview of plant-based natural biostimulants for sustainable horticulture with a particular focus on moringa leaf extracts. Plant Sci. 2020, 295, 110194. [Google Scholar] [CrossRef] [PubMed]
- Ricci, M.; Tilbury, L.; Daridon, B.; Sukalac, K. General Principles to Justify Plant Biostimulant Claims. Front. Plant Sci. 2019, 10. [Google Scholar] [CrossRef]
- Regulation (EU). Fertilising Products Regulation (FPR) (EU) 2019/1009. Available online: https://eur-lex.europa.eu/eli/reg/2019/1009/oj (accessed on 2 June 2022).
- Briglia, N.; Petrozza, A.; Hoeberichts, F.A.; Verhoef, N.; Povero, G. Investigating the Impact of Biostimulants on the Row Crops Corn and Soybean Using High-Efficiency Phenotyping and Next Generation Sequencing. Agronomy 2019, 9, 761. [Google Scholar] [CrossRef] [Green Version]
- EBIC. European Biostimulant Industry Council. Available online: https://biostimulants.eu/wp-content/uploads/2019/10/EBIC-Brochure-English.pdf (accessed on 2 June 2022).
- Zarzecka, K.; Gugała, M.; Mystkowska, I.; Sikorska, A. Total and True Protein Content in Potato Tubers Depending on Herbicides and Biostimulants. Agronomy 2020, 10, 1106. [Google Scholar] [CrossRef]
- De Saeger, J.; Van Praet, S.; Vereecke, D.; Park, J.; Jacques, S.; Han, T.; Depuydt, S. Toward the molecular understanding of the action mechanism of Ascophyllum nodosum extracts on plants. J. Appl. Phycol. 2020, 32, 573–597. [Google Scholar] [CrossRef] [Green Version]
- Caradonia, F.; Ronga, D.; Flore, A.; Barbieri, R.; Moulin, L.; Terzi, V.; Francia, E. Biostimulants and cherry rootstock in-creased tomato fruit yield and quality in sustainable farming systems. Ital. J. Agron. 2020, 15, 121–131. [Google Scholar]
- Pobereżny, J.; Szczepanek, M.; Wszelaczyńska, E.; Prus, P. The Quality of Carrot after Field Biostimulant Application and after Storage. Sustainability 2020, 12, 1386. [Google Scholar] [CrossRef] [Green Version]
- Kocira, S.; Szparaga, A.; Findura, P.; Treder, K. Modification of yield and fiber fractions biosynthesis in Phaseolus vulgaris L. by treatment with biostimulants containing amino acids and seaweed extract. Agronomy 2020, 10, 1338. [Google Scholar] [CrossRef]
- Rouphael, Y.; Colla, G. Biostimulants in agriculture. Front. Plant Sci. 2020, 11, 40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Drobek, M.; Frąc, M.; Cybulska, J. Plant Biostimulants: Importance of the Quality and Yield of Horticultural Crops and the Improvement of Plant Tolerance to Abiotic Stress—A Review. Agronomy 2019, 9, 335. [Google Scholar] [CrossRef] [Green Version]
- Hasanuzzaman, M.; Parvin, K.; Bardhan, K.; Nahar, K.; Anee, T.I.; Masud, A.A.C.; Fotopoulos, V. Biostimulants for the Regulation of Reactive Oxygen Species Metabolism in Plants under Abiotic Stress. Cells 2021, 10, 2537. [Google Scholar] [CrossRef] [PubMed]
- Francesca, S.; Arena, C.; Hay Mele, B.; Schettini, C.; Ambrosino, P.; Barone, A.; Rigano, M.M. The Use of a Plant-Based Biostimulant Improves Plant Performances and Fruit Quality in Tomato Plants Grown at Elevated Temperatures. Agronomy 2020, 10, 363. [Google Scholar] [CrossRef] [Green Version]
- Giordano, M.; El-Nakhel, C.; Caruso, G.; Cozzolino, E.; De Pascale, S.; Kyriacou, M.C.; Colla, G.; Rouphael, Y. Stand-Alone and Combinatorial Effects of Plant-based Biostimulants on the Production and Leaf Quality of Perennial Wall Rocket. Plants 2020, 9, 922. [Google Scholar] [CrossRef]
- Van Oosten, M.J.; Pepe, O.; De Pascale, S.; Silletti, S.; Maggio, A. The role of biostimulants and bioeffectors as alleviators of abiotic stress in crop plants. Chem. Biol. Technol. Agric. 2017, 4, 5. [Google Scholar] [CrossRef] [Green Version]
- Lucini, L.; Miras-Moreno, B.; Rouphael, Y.; Cardarelli, M.; Colla, G. Combining Molecular Weight Fractionation and Metabolomics to Elucidate the Bioactivity of Vegetal Protein Hydrolysates in Tomato Plants. Front. Plant Sci. 2020, 11, 976. [Google Scholar] [CrossRef]
- Petit, A.-N.; Fontaine, F.; Vatsa, P.; Clément, C.; Vaillant-Gaveau, N. Fungicide impacts on photosynthesis in crop plants. Photosynth. Res. 2012, 111, 315–326. [Google Scholar] [CrossRef]
- Junqueira, V.B.; Müller, C.; Rodrigues, A.A.; Amaral, T.S.; Batista, P.F.; Silva, A.A.; Costa, A.C. Do fungicides affect the physiology, reproductive development and productivity of healthy soybean plants? Pestic. Biochem. Physiol. 2021, 172, 104754. [Google Scholar] [CrossRef]
- Amaro, A.C.E.; Baron, D.; Ono, E.O.; Rodrigues, J.D. Physiological effects of strobilurin and carboxamides on plants: An overview. Acta Physiol. Plant. 2020, 42, 4. [Google Scholar] [CrossRef]
- Basha, C.J.; Soniya, M.; Ganiger, P.C. Field evaluation of fungicides against powdery mildew of chilli (Capsicum annuum L.). Int. J. Plant Prot. 2017, 10, 329–332. [Google Scholar] [CrossRef]
- Karkanis, A.; Bilalis, D.; Efthimiadou, A.; Katsenios, N. Effects of field bindweed (Convolvulus arvensis L.) and powdery mildew [Leveillula taurica (Lev.) Arn.] on pepper growth and yield–Short communication. Hortic. Sci. 2012, 39, 135–138. [Google Scholar] [CrossRef] [Green Version]
- Madhavan, S.; Adhipathi, P.; Velazhahan, R.; Paranidharan, V.; Karthikeyan, M. Management of chilli (Capsicum annuum) anthracnose using fungicides and biocontrol agents. Indian Phytopathol. 2017, 70, 86–90. [Google Scholar] [CrossRef] [Green Version]
- Kharkwal, M.; Mishra, J.; Prasad, R. In vitro evaluation of different fungicides in inhibiting the mycelia growth of Colle-totrichum capsici causing Anthracnose of chili. J. Pharmacogn. Phytochem. 2020, 9, 919–921. [Google Scholar]
- Ahiladevi, P.; Prakasm, V. Effect of Azoxystrobin 25 SC on the biochemical parameters of chilli (Capsicum annuum L.). Karnataka J. Agric. Sci. 2014, 27, 355–356. [Google Scholar]
- Gullino, M.L.; Minuto, A.; Gilardi, G.; Garibaldi, A. Efficacy of azoxystrobin and other strobilurins against Fusarium wilts of carnation, cyclamen and Paris daisy. Crop Prot. 2002, 21, 57–61. [Google Scholar] [CrossRef]
- Mahajan, N.S.; Mishra, M.; Tamhane, V.A.; Gupta, V.S.; Giri, A.P. Stress inducible proteomic changes in Capsicum annuum leaves. Plant Physiol. Biochem. 2014, 74, 212–217. [Google Scholar] [CrossRef]
- Kong, X.-M.; Zhou, Q.; Luo, F.; Wei, B.-D.; Wang, Y.-J.; Sun, H.-J.; Zhao, Y.-B.; Ji, S.-J. Transcriptome analysis of harvested bell peppers (Capsicum annuum L.) in response to cold stress. Plant Physiol. Biochem. 2019, 139, 314–324. [Google Scholar] [CrossRef]
- Wang, H.; Liu, Z.; Luo, S.; Li, J.; Zhang, J.; Li, L.; Xie, J. 5-Aminolevulinic acid and hydrogen sulphide alleviate chilling stress in pepper (Capsicum annuum L.) seedlings by enhancing chlorophyll synthesis pathway. Plant Physiol. Biochem. 2021, 167, 567–576. [Google Scholar] [CrossRef]
- Agliassa, C.; Mannino, G.; Molino, D.; Cavalletto, S.; Contartese, V.; Bertea, C.M.; Secchi, F. A new protein hydroly-sate-based biostimulant applied by fertigation promotes relief from drought stress in Capsicum annuum L. Plant Physiol. Biochem. 2021, 166, 1076–1086. [Google Scholar] [CrossRef]
- Tort, N.; Turkyilmaz, B. Physiological Effects of Captan Fungicide on Pepper (Capsicum annuum L.) Plant. Pak. J. Biol. Sci. 2003, 6, 2026–2029. [Google Scholar] [CrossRef] [Green Version]
- Barrajón-Catalán, E.; Álvarez-Martínez, F.J.; Borrás, F.; Pérez, D.; Herrero, N.; Ruiz, J.J.; Micol, V. Metabolomic analysis of the effects of a commercial complex biostimulant on pepper crops. Food Chem. 2020, 310, 125818. [Google Scholar] [CrossRef]
- Parađiković, N.; Teklić, T.; Zeljković, S.; Lisjak, M.; Špoljarević, M. Biostimulants research in some horticultural plant species—A review. Food Energy Secur. 2019, 8, e00162. [Google Scholar] [CrossRef]
- Bellaloui, N.; Brown, P.H. Cultivar differences in boron uptake and distribution in celery (Apium graveolens), tomato (Lycopersicon esculentum) and wheat (Triticum aestivum). Plant Soil 1998, 198, 153–158. [Google Scholar] [CrossRef]
- Strasser, R.J.; Srivastava, A.; Tsimilli-Michael, M. The fluorescence transient as a tool to characterize and screen photo-synthetic samples. In Probing Photosynthesis: Mechanisms, Regulation and Adaptation; CRC Press: Boca Raton, FL, USA, 2000; pp. 445–483. [Google Scholar]
- Ghanem, M.E.; Albacete, A.; Martínez-Andújar, C.; Acosta, M.; Romero-Aranda, R.; Dodd, I.C.; Lutts, S.; Pérez-Alfocea, F. Hormonal changes during salinity-induced leaf senescence in tomato (Solanum lycopersicum L.). J. Exp. Bot. 2008, 59, 3039–3050. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Yemm, E.; Cocking, E.; Ricketts, R. The determination of amino-acids with ninhydrin. Analyst 1955, 80, 209–214. [Google Scholar] [CrossRef]
- Wolf, B. A comprehensive system of leaf analyses and its use for diagnosing crop nutrient status. Commun. Soil Sci. Plant Anal. 1982, 13, 1035–1059. [Google Scholar] [CrossRef]
- Krom, M.D. Spectrophotometric determination of ammonia: A study of a modified Berthelot reaction using salicylate and dichloroisocyanurate. Analyst 1980, 105, 305–316. [Google Scholar] [CrossRef]
- Xu, G.; Fan, X.; Miller, A.J. Plant Nitrogen Assimilation and Use Efficiency. Annu. Rev. Plant Biol. 2012, 63, 153–182. [Google Scholar] [CrossRef] [Green Version]
- Wellburn, A.R. The Spectral Determination of Chlorophylls a and b, as well as Total Carotenoids, Using Various Solvents with Spectrophotometers of Different Resolution. J. Plant Physiol. 1994, 144, 307–313. [Google Scholar] [CrossRef]
- Fu, J.; Huang, B. Involvement of antioxidants and lipid peroxidation in the adaptation of two cool-season grasses to lo-calized drought stress. Environ. Exp. Bot. 2001, 45, 105–114. [Google Scholar] [CrossRef]
- Heath, R.L.; Packer, L. Photoperoxidation in isolated chloroplasts: I. Kinetics and stoichiometry of fatty acid peroxidation. Arch. Biochem. Biophys. 1968, 125, 189–198. [Google Scholar] [CrossRef]
- Mukherjee, S.P.; Choudhuri, M.A. Implications of water stress-induced changes in the levels of endogenous ascorbic acid and hydrogen peroxide in Vigna seedlings. Physiol. Plant. 1983, 58, 166–170. [Google Scholar] [CrossRef]
- Zhongguang, L.; Ming, G. Improvement of measurement method for superoxide anion radical in plant. Acta Bot. Yunnanica 2005, 27, 211–216. [Google Scholar]
- Rivero, R.M.; Ruiz, J.M.; García, P.C.; López-Lefebre, L.R.; Sánchez, E.; Romero, L. Resistance to cold and heat stress: Accumulation of phenolic compounds in tomato and watermelon plants. Plant Sci. 2001, 160, 315–321. [Google Scholar] [CrossRef]
- Kim, D.-O.; Jeong, S.W.; Lee, C.Y. Antioxidant capacity of phenolic phytochemicals from various cultivars of plums. Food Chem. 2003, 81, 321–326. [Google Scholar] [CrossRef]
- Giusti, M.M.; Wrolstad, R.E. Characterization and Measurement of Anthocyanins by UV-Visible Spectroscopy. Curr. Protoc. Food Anal. Chem. 2001, F1.2.1–F1.2.13. [Google Scholar] [CrossRef]
- Ashour, M.; Hassan, S.M.; Elshobary, M.E.; Ammar, G.A.; Gaber, A.; Alsanie, W.F.; Mansour, A.T.; El-Shenody, R. Impact of commercial seaweed liquid extract (TAM®) biostimulant and its bioactive molecules on Growth and Antioxidant Activities of Hot Pepper (Capsicum annuum). Plants 2021, 10, 1045. [Google Scholar] [CrossRef]
- Di Mola, I.; Ottaiano, L.; Cozzolino, E.; Senatore, M.; Giordano, M.; El-Nakhel, C.; Sacco, A.; Rouphael, Y.; Colla, G.; Mori, M. Plant-Based Biostimulants Influence the Agronomical, Physiological, and Qualitative Responses of Baby Rocket Leaves under Diverse Nitrogen Conditions. Plants 2019, 8, 522. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cho, M.-H.; Park, H.L.; Hahn, T.-R. Engineering leaf carbon metabolism to improve plant productivity. Plant Biotechnol. Rep. 2015, 9, 1–10. [Google Scholar] [CrossRef]
- Bulgari, R.; Morgutti, S.; Cocetta, G.; Negrini, N.; Farris, S.; Calcante, A.; Spinardi, A.; Ferrari, E.; Mignani, I.; Oberti, R.; et al. Evaluation of Borage Extracts as Potential Biostimulant Using a Phenomic, Agronomic, Physiological, and Biochemical Approach. Front. Plant Sci. 2017, 8, 935. [Google Scholar] [CrossRef] [PubMed]
- Choi, S.; Colla, G.; Cardarelli, M.; Kim, H.-J. Effects of Plant-Derived Protein Hydrolysates on Yield, Quality, and Nitrogen Use Efficiency of Greenhouse Grown Lettuce and Tomato. Agronomy 2022, 12, 1018. [Google Scholar] [CrossRef]
- Khan, N.; Bano, A.; Ali, S.; Babar, M.A. Crosstalk amongst phytohormones from planta and PGPR under biotic and abiotic stresses. Plant Growth Regul. 2020, 90, 189–203. [Google Scholar] [CrossRef]
- Boudet, A.-M. Evolution and current status of research in phenolic compounds. Phytochemistry 2007, 68, 2722–2735. [Google Scholar] [CrossRef] [PubMed]
- De Bang, T.C.; Husted, S.; Laursen, K.H.; Persson, D.P.; Schjoerring, J.K. The molecular–physiological functions of mineral macronutrients and their consequences for deficiency symptoms in plants. New Phytol. 2021, 229, 2446–2469. [Google Scholar] [CrossRef] [PubMed]
- Iqbal, A.; Qiang, D.; Zhun, W.; Xiangru, W.; Huiping, G.; Hengheng, Z.; Nianchang, P.; Xiling, Z.; Meizhen, S. Growth and nitrogen metabolism are associated with nitrogen-use efficiency in cotton genotypes. Plant Physiol. Biochem. 2020, 149, 61–74. [Google Scholar] [CrossRef]
- Congreves, K.A.; Otchere, O.; Ferland, D.; Farzadfar, S.; Williams, S.; Arcand, M.M. Nitrogen Use Efficiency Definitions of Today and Tomorrow. Front. Plant Sci. 2021, 12, 637108. [Google Scholar] [CrossRef]
- Colla, G.; Rouphael, Y.; Canaguier, R.; Svecova, E.; Cardarelli, M. Biostimulant action of a plant-derived protein hydrol-ysate produced through enzymatic hydrolysis. Front. Plant Sci. 2014, 5, 448. [Google Scholar] [CrossRef] [Green Version]
- Epstein, E.; Bloom, A. Mineral Nutrition of Plants: Principles and Perspectives, 2nd ed.; Sinauer Associates Inc.: Sunderland, UK, 2005; Volume 2005. [Google Scholar]
- Maathuis, F.J.M. Physiological functions of mineral macronutrients. Curr. Opin. Plant Biol. 2009, 12, 250–258. [Google Scholar] [CrossRef]
- Rouphael, Y.; Giordano, M.; Cardarelli, M.; Cozzolino, E.; Mori, M.; Kyriacou, M.C.; Bonini, P.; Colla, G. Plant-and sea-weed-based extracts increase yield but differentially modulate nutritional quality of greenhouse spinach through biostimulant action. Agronomy 2018, 8, 126. [Google Scholar] [CrossRef] [Green Version]
- Hänsch, R.; Mendel, R.R. Physiological functions of mineral micronutrients (Cu, Zn, Mn, Fe, Ni, Mo, B, Cl). Curr. Opin. Plant Biol. 2009, 12, 259–266. [Google Scholar] [CrossRef] [PubMed]
- Ali, E.; Hassan, F.; Elgimabi, M. Improving the growth, yield and volatile oil content of Pelargonium graveolens L. Herit by foliar application with moringa leaf extract through motivating physiological and biochemical parameters. S. Afr. J. Bot. 2018, 119, 383–389. [Google Scholar] [CrossRef]
- Abdalla, M.M. The potential of Moringa oleifera extract as a biostimulant in enhancing the growth, biochemical and hormonal contents in rocket (Eruca vesicaria subsp. sativa) plants. Int. J. Plant Physiol. Biochem. 2013, 5, 42–49. [Google Scholar] [CrossRef]
- Debona, D.; Nascimento, K.; Gomes, J.; Aucique-Perez, C.; Rodrigues, F. Physiological changes promoted by a strobilurin fungicide in the rice-Bipolaris oryzae interaction. Pestic. Biochem. Physiol. 2016, 130, 8–16. [Google Scholar] [CrossRef] [PubMed]
- Nason, M.A.; Farrar, J.; Bartlett, D. Strobilurin fungicides induce changes in photosynthetic gas exchange that do not improve water use efficiency of plants grown under conditions of water stress. Pest Manag. Sci. 2007, 63, 1191–1200. [Google Scholar] [CrossRef] [PubMed]
- Yadav, S.; Atri, N. Reactive Oxygen Species and Response of the Calvin–Benson Cycle: An Overview. In Reactive Oxygen Species in Plants: Boon or Bane-Revisiting the Role of ROS; John Wiley & Sons Ltd.: Oxford, UK, 2017; p. 157. [Google Scholar]
- Jalakas, P.; Tulva, I.; Kangor, T.; Sooväli, P.; Rasulov, B.; Tamm, Ü.; Koppel, M.; Kollist, H.; Merilo, E. Gas exchange-yield relationships of malting barley genotypes treated with fungicides and biostimulants. Eur. J. Agron. 2018, 99, 129–137. [Google Scholar] [CrossRef]
- Shahid, M.; Ahmed, B.; Zaidi, A.; Khan, M.S. Toxicity of fungicides to Pisum sativum: A study of oxidative damage, growth suppression, cellular death and morpho-anatomical changes. RSC Adv. 2018, 8, 38483–38498. [Google Scholar] [CrossRef] [Green Version]
- Mayer, A.; Harel, E. Phenoloxidases and their significance in fruit and vegetables. Food Enzymol. 1991, 1, 373–398. [Google Scholar]
- Havaux, M. Carotenoids as membrane stabilizers in chloroplasts. Trends Plant Sci. 1998, 3, 147–151. [Google Scholar] [CrossRef]
- Parađiković, N.; Vinković, T.; Vinković Vrček, I.; Žuntar, I.; Bojić, M.; Medić-Šarić, M. Effect of natural biostimulants on yield and nutritional quality: An example of sweet yellow pepper (Capsicum annuum L.) plants. J. Sci. Food Agric. 2011, 91, 2146–2152. [Google Scholar] [CrossRef] [PubMed]
- Klokić, I.; Koleška, I.; Hasanagić, D.; Murtić, S.; Bosančić, B.; Todorović, V. Biostimulants’ influence on tomato fruit char-acteristics at conventional and low-input NPK regime. Acta Agric. Scand. Sect. B Soil Plant Sci. 2020, 70, 233–240. [Google Scholar]


{kind=link}
{kind=link}
| Product | Percentage |
|---|---|
| Free amino acids | 7% w/w |
| Polysaccharides | 3% w/w |
| Total nitrogen (N) | 4% w/w |
| Organic nitrogen (N) | 2.1% w/w |
| Nitric nitrogen (N) | 0.9% w/w |
| Ammoniacal nitrogen (N) | 1% w/w |
| Phosphorous pentoxide (P2O5) water soluble | 4% w/w |
| Potassium oxide (K2O) water soluble | 3% w/w |
| Iron (Fe) water soluble | 0.4% w/w |
| Manganese (Mn) water-soluble | 0.1% w/w |
| Boron (B) water-soluble | 0.1% w/w |
| Zinc (Zn) soluble in water | 0.085% w/w |
| Copper (Cu) water-soluble | 0.02% w/w |
| Molybdenum (Mo) water-soluble | 0.01% w/w |
| Treatments | A (µmol m⁻2 s⁻1) | E (mmol m⁻2 s⁻1) | gsw (mmol m⁻2 s⁻1) | WUE | Soluble Sugars (µg g⁻1 FW) |
|---|---|---|---|---|---|
| Control | 3.10 ± 1.08 | 0.38 ± 0.08 | 25.72 ± 5.54 | 7.90 ± 1.23 | 55.19 ± 3.50 |
| RAZORMIN® | 4.89 ± 0.75 | 0.55 ± 0.05 | 38.94 ± 3.23 | 8.81 ± 0.78 | 83.22 ± 6.88 |
| p-value | * | * | ** | NS | ** |
| LSD0.05 | 1.60 | 0.11 | 7.84 | 1.78 | 12.37 |
| Hormones | Control | RAZORMIN® | p-Value |
|---|---|---|---|
| IAA | 0.96 ± 0.09 | 1.11 ± 0.08 | * |
| tZ | 357.2 ± 8.95 | 399.2 ± 8.44 | *** |
| iP | 0.42 ± 0.06 | 0.47 ± 0.09 | NS |
| GA1 | 0.15 ± 0.01 | 0.36 ± 0.01 | *** |
| GA3 | 0.06 ± 0.01 | 0.14 ± 0.01 | *** |
| GA4 | 0.17 ± 0.01 | 0.25 ± 0.02 | *** |
| ABA | 73.95 ± 4.71 | 77.50 ± 2.25 | NS |
| ACC | 19.76 ± 0.71 | 20.11 ± 0.96 | NS |
| JA | 542.9 ± 8.58 | 539.6 ± 8.53 | NS |
| SA | 4033 ± 63.76 | 4373 ± 64.39 | *** |
| Treatments | Soluble Amino Acids (mg g⁻1 FW) | Soluble Proteins (mg g⁻1 FW) | N (mg g−1 DW) | NUtE (g2 DW mg−1 N) |
|---|---|---|---|---|
| Control | 2.04 ± 0.10 | 0.68 ± 0.02 | 61.36 ± 4.72 | 0.16 ± 0.01 |
| RAZORMIN® | 3.25 ± 0.14 | 0.69 ± 0.02 | 67.00 ± 4.80 | 0.21 ± 0.02 |
| p-Value | ** | NS | ** | *** |
| LSD0.05 | 0.17 | 0.03 | 4.75 | 0.01 |
| Treatments | P (mg g−1 DW) | K (mg g−1 DW) | Ca (mg g−1 DW) | Mg (mg g−1 DW) | S (mg g−1 DW) |
|---|---|---|---|---|---|
| Control | 5.59 ± 0.79 | 31.88 ± 2.05 | 19.62 ± 1.47 | 6.82 ± 0.36 | 4.52 ± 0.19 |
| RAZORMIN® | 6.58 ± 0.18 | 35.28 ± 0.53 | 21.95 ± 0.39 | 7.87 ± 0.43 | 5.20 ± 0.21 |
| p-value | * | * | * | ** | * |
| LSD0.05 | 1.30 | 3.39 | 2.43 | 0.90 | 0.45 |
| Treatments | Fe (µg g−1 DW) | Cu (µg g−1 DW) | Mn (µg g−1 DW) | Zn (µg g−1 DW) | Mo (µg g−1 DW) | B (µg g−1 DW) |
|---|---|---|---|---|---|---|
| Control | 180.35 ± 18.51 | 16.66 ± 0.89 | 46.39 ± 3.45 | 49.75 ± 4.12 | 1.88 ± 0.19 | 29.99 ± 2.90 |
| RAZORMIN® | 234.62 ± 24.38 | 17.00 ± 1.48 | 42.49 ± 2.27 | 57.35 ± 3.44 | 2.39 ± 0.09 | 42.06 ± 1.68 |
| p-value | ** | NS | NS | ** | ** | *** |
| LSD0.05 | 49.07 | 2.76 | 6.62 | 8.60 | 0.32 | 5.37 |
| Treatments | Fv/Fm | RC/ABS | PIABS | 1 − Vj |
|---|---|---|---|---|
| Control | 0.815 ± 0.001 a | 0.81 ± 0.09 a | 7.48 ± 1.84 a | 0.67 ± 0.03 a |
| Azoxystrobin | 0.814 ± 0.004 a | 0.83 ± 0.07 a | 7.44 ± 1.35 a | 0.67 ± 0.02 a |
| Azoxystrobin + RAZORMIN® | 0.814 ± 0.008 a | 0.80 ± 0.05 a | 7.52 ± 1.06 a | 0.68 ± 0.02 a |
| p-value | NS | NS | NS | NS |
| LSD0.05 | 0.006 | 0.08 | 1.78 | 0.02 |
| Treatments | Chl a (mg g−1 FW) | Chl b (mg g−1 FW) | Carotenoids (mg g−1 FW) |
|---|---|---|---|
| Control | 0.56 ± 0.01 a | 0.31 ± 0.01 a | 0.37 ± 0.01 a |
| Azoxystrobin | 0.50 ± 0.02 b | 0.29 ± 0.01 a | 0.35 ± 0.02 a |
| Azoxystrobin + RAZORMIN® | 0.56 ± 0.01 a | 0.30 ± 0.01 a | 0.37± 0.01 a |
| p-Value | * | NS | NS |
| LSD0.05 | 0.05 | 0.03 | 0.04 |
| Treatments | A (µmol m⁻2 s⁻1) | E (mmol m⁻2 s⁻1) | gsw (mmol m⁻2 s⁻1) | WUE |
|---|---|---|---|---|
| Control | 8.61 ± 1.45 a | 1.09 ± 0.20 a | 81.43 ± 11.57 a | 7.94 ± 1.11 b |
| Azoxystrobin | 7.81 ± 0.88 a | 0.61 ± 0.14 b | 42.01 ± 9.89 b | 13.04 ± 1.42 a |
| Azoxystrobin + RAZORMIN® | 7.13 ± 0.95 a | 0.48 ± 0.09 b | 34.15 ± 6.61 b | 14.91 ± 0.99 a |
| p-Value | NS | ** | ** | *** |
| LSD0.05 | 2.23 | 0.30 | 19.14 | 2.37 |
| Treatments | MDA (µM g−1 FW) | H2O2 (µg g−1 FW) | O2− (µg g−1 FW) |
|---|---|---|---|
| Control | 1.27 ± 0.21 b | 54.44 ± 2.17 b | 4.50 ± 0.06 b |
| Azoxystrobin | 2.87 ± 0.28 a | 88.06 ± 1.38 a | 6.10 ± 0.04 a |
| Azoxystrobin + RAZORMIN® | 1.35 ± 0.14 b | 60.36 ± 2.56 b | 5.09 ± 0.13 b |
| p-value | *** | ** | *** |
| LSD0.05 | 0.42 | 4.18 | 0.49 |
| Treatments | Total Phenols (mg g−1 FW) | Flavonoids (mg g−1 FW) | Anthocyanins (mg g−1 FW) |
|---|---|---|---|
| Control | 23.60 ± 0.92 c | 19.67 ± 0.29 b | 2.98 ± 0.22 b |
| Azoxystrobin | 26.99 ± 0.99 b | 21.01 ± 0.86 b | 1.47 ± 0.16 c |
| Azoxystrobin + RAZORMIN® | 33.00 ± 0.56 a | 25.73 ± 0.89 a | 6.95 ± 0.52 a |
| p-Value | *** | *** | *** |
| LSD0.05 | 1.68 | 1.38 | 0.68 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ahmad, A.; Navarro-León, E.; Izquierdo-Ramos, M.J.; Rios, J.J.; Blasco, B.; Navarro-Morillo, I.; Ruiz, J.M. Analysis of RAZORMIN® as a Biostimulant and Its Effect on the Phytotoxicity Mitigation Caused by Fungicide Azoxystrobin in Pepper. Agronomy 2022, 12, 1418. https://doi.org/10.3390/agronomy12061418
Ahmad A, Navarro-León E, Izquierdo-Ramos MJ, Rios JJ, Blasco B, Navarro-Morillo I, Ruiz JM. Analysis of RAZORMIN® as a Biostimulant and Its Effect on the Phytotoxicity Mitigation Caused by Fungicide Azoxystrobin in Pepper. Agronomy. 2022; 12(6):1418. https://doi.org/10.3390/agronomy12061418
Chicago/Turabian StyleAhmad, Ali, Eloy Navarro-León, María José Izquierdo-Ramos, Juan José Rios, Begoña Blasco, Iván Navarro-Morillo, and Juan Manuel Ruiz. 2022. "Analysis of RAZORMIN® as a Biostimulant and Its Effect on the Phytotoxicity Mitigation Caused by Fungicide Azoxystrobin in Pepper" Agronomy 12, no. 6: 1418. https://doi.org/10.3390/agronomy12061418