
Transcriptome Analysis of ‘Kyoho’ Grapevine Leaves Identifies Heat Response Genes Involved in the Transcriptional Regulation of Photosynthesis and Abscisic Acid
 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials and Treatments
2.2. RNA-Seq Analysis
2.3. Gene Function Annotations and Classifications
2.4. Identification of Differentially Expressed Genes (DEGs)
2.5. Validation of RNA-Seq by Quantitative Real-Time RT-PCR
2.6. Analysis of Photosynthetic Gas Exchange
2.7. Measurement of Chlorophyll Fluorescence
2.8. Measurement of Endogenous ABA Content
2.9. Statistical Analysis
3. Results
3.1. Illumina Sequencing
3.2. DEGs among the HS Treatment and Control in Grape
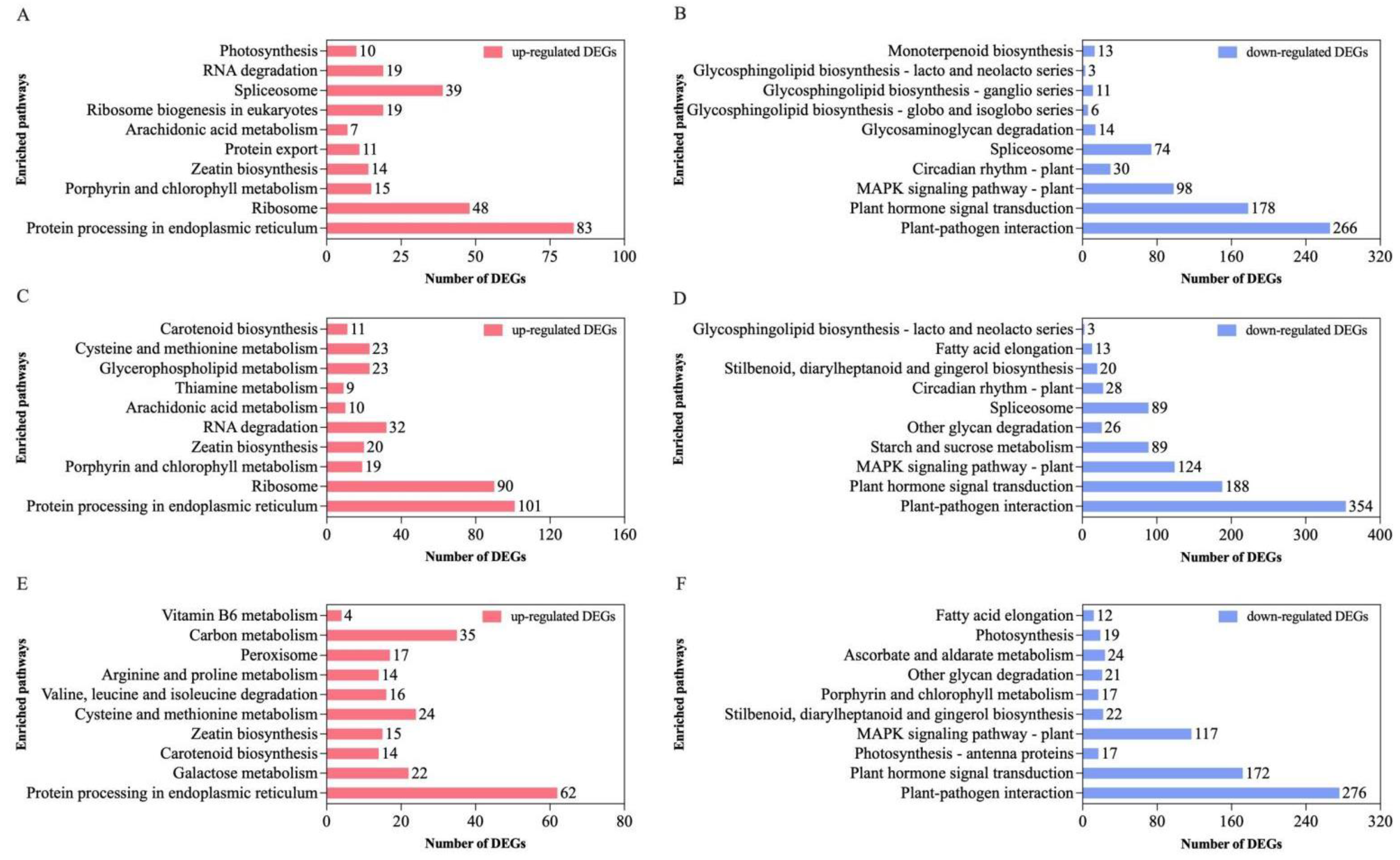
3.3. KEGG Pathway Enrichment Analysis of DEGs
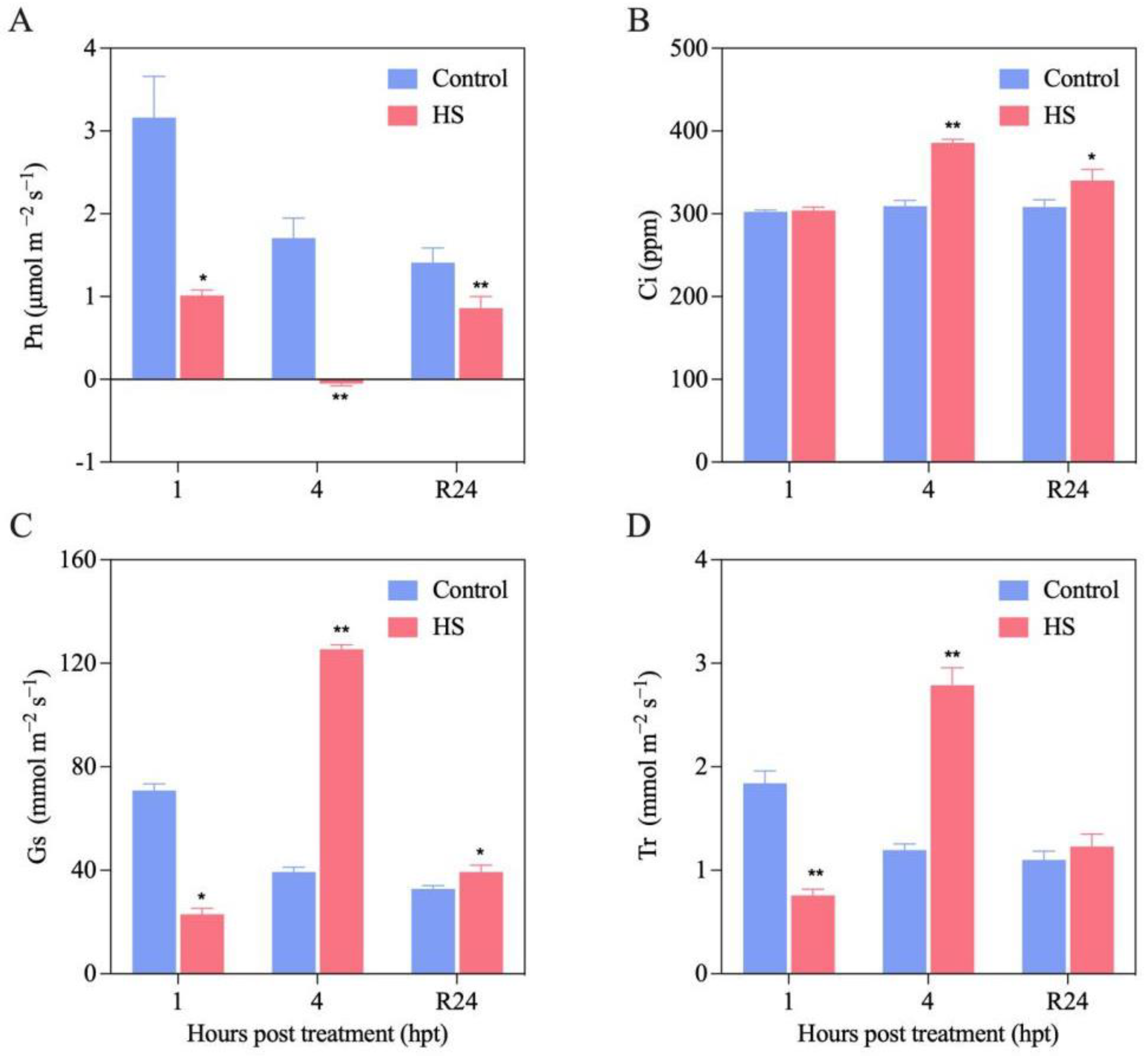
3.4. Effect of HS Treatment on Gas Exchange
3.5. Effect of HS Treatment on Donor Side, Reaction Center and Acceptor Side of PSⅡ
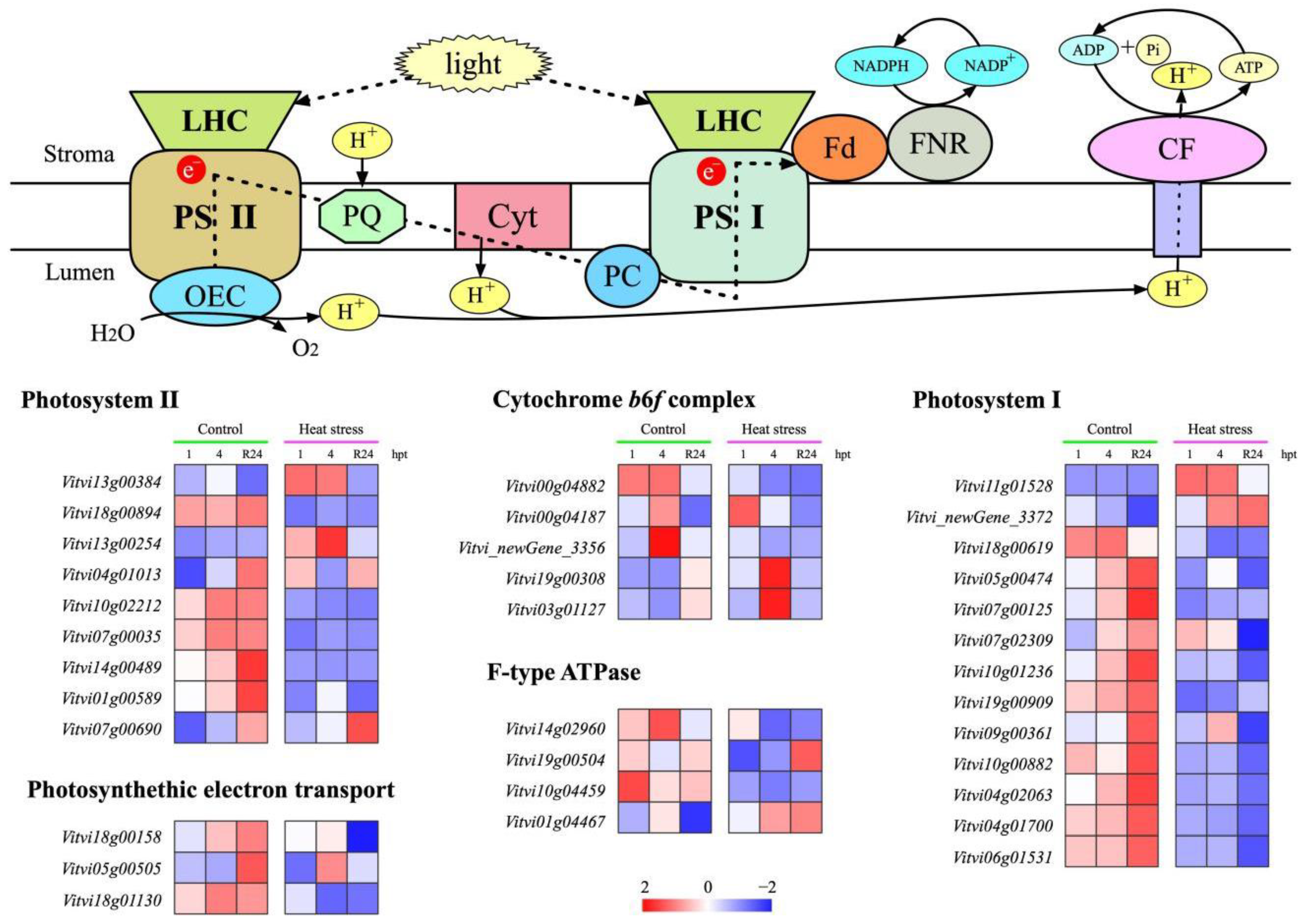
3.6. Effect of HS Treatment on Expression of Photosynthesis-Related Genes
3.7. Effect of HS Treatment on Expression of ABA Biosynthesis, Catabolism and Signal Transduction-Related Genes
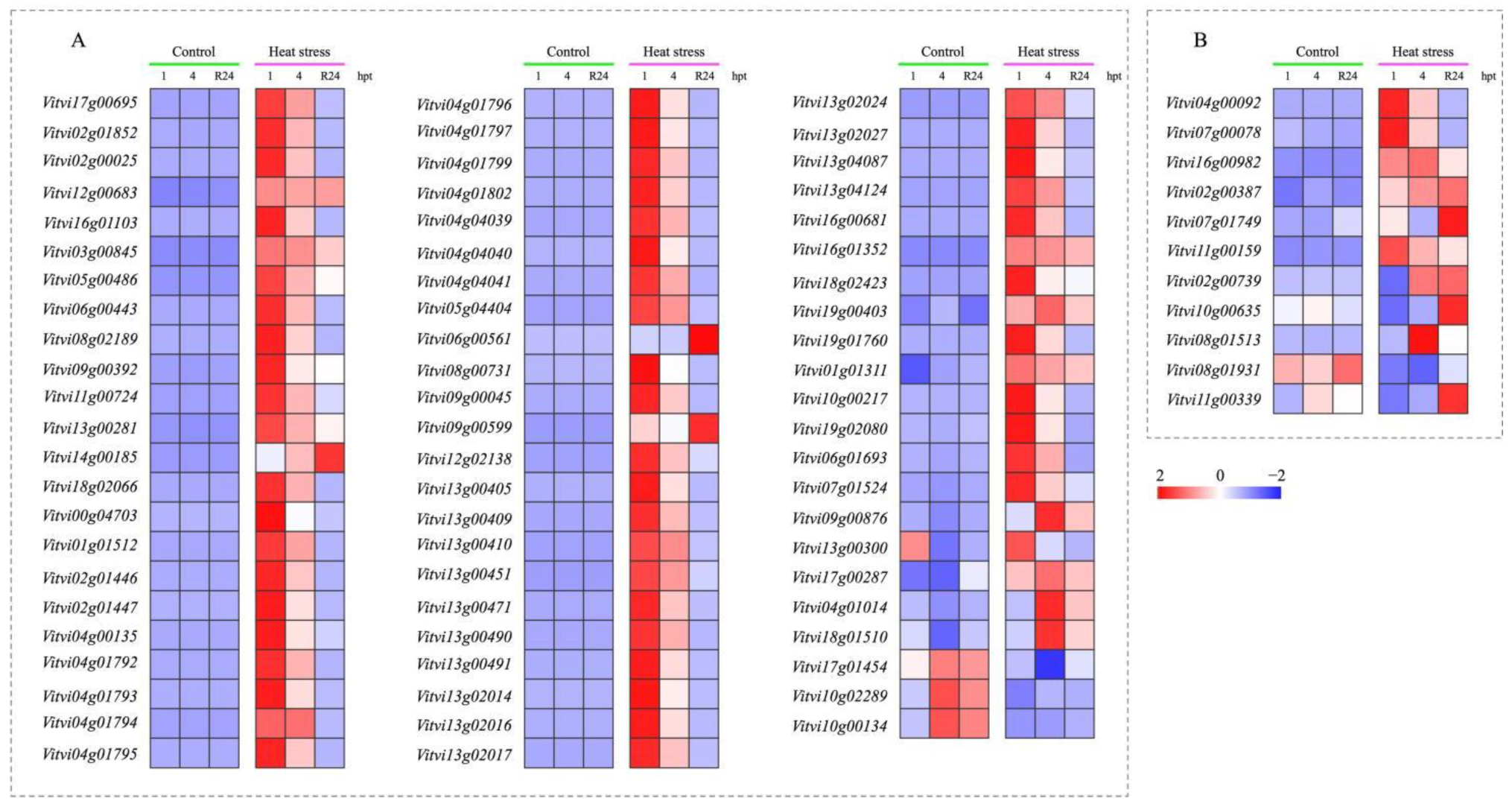
3.8. Effect of HS Treatment on Expression of HSPs and HSFs
4. Discussion
4.1. Photosynthesis Was Inhibited by HS Treatment
4.2. ABA Biosynthesis, Catabolism and Signal Transduction Was Affected by HS Treatment
4.3. Heat Shock Proteins and Heat Shock Factors Were Mostly Up-Regulated by HS Treatment
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| HS | heat stress |
| ABA | abscisic acid |
| HSFs | heat shock transcription factors |
| HSPs | heat shock proteins |
| iiTRAQ | isobaric tags for relative and absolute quantitation |
| qRT-PCR | quantitative real-time RT-PCR |
| FPKM | fragments per kilobase of transcript per million mapped reads |
| NR | NCBI non-redundant protein sequence database |
| COG | Cluster of Orthologous Groups |
| Pfam | The database of Homologous protein family |
| KEGG | The database of Kyoto Encyclopedia of Genes and Genomes |
| GO | Gene Ontology |
| DEGs | differentially expressed genes |
| Pn | Net photosynthetic rate |
| Gs | stomatal conductance |
| Ci | substomatal CO2 concentration |
| Tr | transpiration rate |
| PPFD | photosynthetic photon flux density |
| PSⅡ | photosystem Ⅱ |
| PSⅠ | photosystem Ⅰ |
| Fv/Fm | maximum quantum efficiency of PSⅡ photochemistry |
| CSm | excited leaf cross-section |
| ABS/CSm | absorption flux per CSm approximated |
| TRo/CSm | trapped energy flux per CSm |
| RC/CSm | percentage of active/inactive reaction centers per CSm |
| ETo/CSm | electron transport flux per CSm |
| DIo/CSm | dissipated energy flux per CSm |
| Hpt | hour post treatment |
| R24 hpt | 24 h after recovery from treatment |
| PSY | phytoene synthase |
| CrtZ | beta-carotene hydroxylase |
| ZEP | zeaxanthin epoxidase |
| NCED | 9-cis-epoxycarotenoid dioxygenase |
| ABAO | abscisic-aldehyde oxidase |
| ABA8ox | ABA 8′-hydroxylase |
| ABA-GT | ABA glucosyltransferase |
| PA | phaseic acid |
| ABA-GE | ABA glucosyl-ester |
| PP2C | type 2C serine threonine protein phosphatase |
| SnRK2 | SNF1-related type 2 protein kinases |
| ABF | ABA-responsive element binding factor |
| ABI | ABA-insensitive |
| OEC | oxygen-evolving complex |
| LHCⅡ | PSⅡ-light-harvesting complex Ⅱ |
References
- Vivier, M.A.; Pretorius, I.S. Genetically tailored grapevines for the wine industry. Trends Biotechnol. 2002, 20, 472–478. [Google Scholar] [CrossRef]
- Cramer, G.; Urano, K.; Delrot, S.; Pezzotti, M.; Shinozaki, K. Effects of abiotic stress on plants: A systems biology perspective. BMC Plant Biol. 2011, 11, 163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, H.G.; Liu, G.J.; Liu, G.T.; Yan, B.F.; Duan, W.; Wang, L.J.; Li, S.H. Comparison of investigation methods of heat injury in grapevine (Vitis) and assessment to heat tolerance in different cultivars and species. BMC Plant Biol. 2014, 14, 156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Venios, X.; Korkas, E.; Nisiotou, A.; Banilas, G. Grapevine responses to heat stress and global warming. Plants 2020, 9, 1754. [Google Scholar] [CrossRef] [PubMed]
- Liu, G.T.; Wang, J.F.; Cramer, G.; Dai, Z.W.; Duan, W.; Xu, H.G.; Wu, B.H.; Fan, P.G.; Wang, L.J.; Li, S.H. Transcriptomic analysis of grape (Vitis vinifera L.) leaves during and after recovery from heat stress. BMC Plant Biol. 2012, 12, 174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cohen, S.D.; Tarara, J.M.; Gambetta, G.A.; Matthews, M.A.; Kennedy, J.A. Impact of diurnal temperature variation on grape berry development, proanthocyanidin accumulation, and the expression of flavonoid pathway genes. J. Exp. Bot. 2012, 63, 2655–2665. [Google Scholar] [CrossRef] [Green Version]
- Greer, D.H.; Weedon, M.M. The impact of high temperatures on Vitis vinifera cv. Semilion grapevine performance and berry ripening. Front. Plant Sci. 2013, 4, 491. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.H.; Huang, W.D.; Liu, Y.P.; Pan, Q.H. Effects of temperature acclimation pretreatment on the ultrastructure of mesophyll cells in young grape plants (Vitis vinifera L. cv. Jingxiu) under cross-temperature stresses. J. Integr. Plant Biol. 2005, 47, 959–970. [Google Scholar] [CrossRef]
- Kriedemann, P.E. Photosynthesis in vine leaves as a function of light intensity, temperature and leaf age. Vitis 1968, 7, 213–220. [Google Scholar]
- Ferrini, F.; Mattii, G.B.; Nicese, F.P. Effect of temperature on key physiological responses of grapevine leaf. Am. J. Enol. Viticult. 1995, 46, 375–379. [Google Scholar]
- Schultz, H.R. Extension of a Farquhar model for limitations of leaf photosynthesis induced by light environment, phenology and leaf age in grapevines (Vitis vinifera L. cv. White Riesling and Zinfandel). Funct. Plant Biol. 2003, 30, 673–687. [Google Scholar] [CrossRef]
- Yu, D.J.; Kim, S.J.; Lee, H.J. Stomatal and non-stomatal limitations to photosynthesis in field-grown grapevine cultivars. Biol. Plant. 2009, 53, 133–137. [Google Scholar] [CrossRef]
- Zsofi, Z.; Varadi, G.; Balo, B.; Marschall, M.; Nagy, Z. Heat acclimation of grapevine leaf photosynthesis: Mezzo- and macroclimate aspects. Funct. Plant Biol. 2009, 36, 310–322. [Google Scholar] [CrossRef]
- Luo, H.B.; Ma, L.; Xi, H.F.; Duan, W.; Li, S.H.; Loescher, W.; Wang, J.F.; Wang, L.J. Photosynthetic responses to heat treatments at different temperatures and following recovery in grapevine (Vitis amurensis L.) leaves. PLoS ONE 2011, 6, e23033. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.J.; Fan, L.; Loescher, W.; Duan, W.; Liu, G.J.; Cheng, J.S.; Luo, H.B.; Li, S.H. Salicylic acid alleviates decreases in photosynthesis under heat stress and accelerates recovery in grapevine leaves. BMC Plant Biol. 2010, 10, 34. [Google Scholar] [CrossRef] [Green Version]
- Greer, D.H.; Weedon, M.M. Modelling photosynthetic responses to temperature of grapevine (Vitis vinifera cv. Semillon) leaves on vines grown in a hot climate. Plant Cell Environ. 2012, 35, 1050–1064. [Google Scholar] [CrossRef]
- Greer, D.H.; Weston, C. Heat stress affects flowering, berry growth, sugar accumulation and photosynthesis of Vitis vinifera cv. Semillon grapevines grown in a controlled environment. Func. Plant Biol. 2010, 37, 206–214. [Google Scholar] [CrossRef]
- Sepulveda, G.; Kliewer, W.M. Stomatal response of three grapevine cultivars (Vitis vinifera L.) to high temperature. Am. J. Enol. Vitic. 1986, 37, 44–52. [Google Scholar]
- Baron, K.N.; Schroeder, D.F.; Stasolla, C. Transcriptional response of abscisic acid (ABA) metabolism and transport to cold and heat stress applied at the reproductive stage of development in Arabidopsis thaliana. Plant Sci. 2012, 188–189, 48–59. [Google Scholar] [CrossRef]
- Sharma, E.; Borah, P.; Kaur, A.; Bhatnagar, A.; Mohapatra, T.; Kapoor, S.; Khurana, J.P. A comprehensive transcriptome analysis of contrasting rice cultivars highlights the role of auxin and ABA responsive genes in heat stress response. Genomics 2021, 113, 1247–1261. [Google Scholar] [CrossRef]
- Nambara, E.; Marion-Poll, A. Abscisic acid biosynthesis and catabolism. Annu. Rev. Plant Biol. 2005, 56, 165–185. [Google Scholar] [CrossRef] [Green Version]
- Larkindale, J.; Hall, J.D.; Knight, M.R.; Vierling, E. Heat stress phenotypes of Arabidopsis mutants implicate multiple signaling pathways in the acquisition of thermotolerance. Plant Physiol. 2005, 138, 882–897. [Google Scholar] [CrossRef] [Green Version]
- Larkindale, J.; Huang, B. Thermotolerance and antioxidant systems in Agrostis stolonifera: Involvement of salicylic acid, abscisic acid, calcium, hydrogen peroxide, and ethylene. J. Plant Physiol. 2004, 161, 405–413. [Google Scholar] [CrossRef]
- Gong, M.; Li, Y.J.; Chen, S.Z. Abscisic acid induced thermotolerance in maize seedlings is mediated by Ca2+ and associated with antioxidant systems. J. Plant Physiol. 1998, 153, 615–621. [Google Scholar] [CrossRef]
- Robertson, A.J.; Ishikawa, M.; Gusta, L.V.; MacKenzie, S.L. Abscisic acid-Induced heat tolerance in bromus inermis leyss cell-suspension cultures (heat-stable, abscisic acid-responsive polypeptides in combination with sucrose confer enhanced thermostability). Plant Physiol. 1994, 105, 181–190. [Google Scholar] [CrossRef] [Green Version]
- Zarrouk, O.; Brunetti, C.; Egipto, R.; Pinheiro, C.; Genebra, T.; Gori, A.; Lopes, C.M.; Tattini, M.; Chaves, M.M. Grape ripening is regulated by deficit irrigation/elevated temperatures according to cluster position in the canopy. Front. Plant Sci. 2016, 7, 1640. [Google Scholar] [CrossRef] [Green Version]
- Pillet, J.; Egert, A.; Pieri, P.; Lecourieux, F.; Kappel, C.; Charon, J.; Gomes, E.; Keller, F.; Delrot, S.; Lecourieux, D. VvGOLS1 and VvHsfA2 are involved in the heat stress responses in grapevine berries. Plant Cell Physiol. 2012, 53, 1776–1792. [Google Scholar] [CrossRef]
- Peng, S.B.; Zhu, Z.G.; Zhao, K.; Shi, J.L.; Yang, Y.Z.; He, M.Y.; Wang, Y.J. A novel heat shock transcription factor, VpHsf1, from Chinese Wild Vitis pseudoreticulata is involved in biotic and abiotic stresses. Plant Mol. Biol. Rep. 2013, 31, 240–247. [Google Scholar] [CrossRef]
- Liu, G.T.; Chai, F.M.; Wang, Y.; Jiang, J.Z.; Duan, W.; Wang, Y.T.; Wang, F.F.; Li, S.H.; Wang, L.J. Genome-wide identification and classification of HSF family in grape, and their transcriptional analysis under heat acclimation and heat stress. Hortic. Plant J. 2018, 4, 133–143. [Google Scholar] [CrossRef]
- Kobayashi, M.; Katoh, H.; Takayanagi, T.; Suzuki, S. Characterization of thermotolerance-related genes in grapevine (Vitis vinifera). J. Plant Physiol. 2010, 167, 812–819. [Google Scholar] [CrossRef]
- Kobayashi, M.; Takato, H.; Fujita, K.; Suzuki, S. HSG1, a grape Bcl-2-associated athanogene, promotes floral transition by activating CONSTANS expression in transgenic Arabidopsis plant. Mol. Biol. Rep. 2012, 39, 4367–4374. [Google Scholar] [CrossRef] [PubMed]
- Liu, G.T.; Ma, L.; Duan, W.; Wang, B.C.; Li, J.H.; Xu, H.G.; Yan, X.Q.; Yan, B.F.; Li, S.H.; Wang, L.J. Differential proteomic analysis of grapevine leaves by iTRAQ reveals responses to heat stress and subsequent recovery. BMC Plant Biol. 2014, 14, 110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, J.F.; Liu, X.N.; Liu, C.H.; Liu, G.T.; Li, S.H.; Wang, L.J. Integrating omics and alternative splicing reveals insights into grape response to high temperature. Plant Physiol. 2017, 173, 1502–1518. [Google Scholar] [CrossRef] [PubMed]
- Rienth, M.; Torregrosa, L.; Sarah, G.; Ardisson, M.; Brillouet, J.-M.; Romieu, C. Temperature desynchronizes sugar and organic acid metabolism in ripening grapevine fruits and remodels their transcriptome. BMC Plant Biol. 2016, 16, 164. [Google Scholar] [CrossRef]
- Pucker, B.; Schwandner, A.; Becker, S.; Hausmann, L.; Viehöver, P.; Töpfer, R.; Weisshaar, B.; Holtgräwe, D. RNA-Seq time series of Vitis vinifera bud development reveals correlation of expression patterns with the local temperature profile. Plants 2020, 9, 1548. [Google Scholar] [CrossRef]
- Toups, H.S.; Cochetel, N.; Gray, D.; Cramer, G.R. VviERF6Ls: An expanded clade in Vitis responds transcriptionally to abiotic and biotic stresses and berry development. BMC Genom. 2020, 21, 472. [Google Scholar] [CrossRef]
- Jallion, O.; Aury, J.M.; Noel, B.; Policriti, A.; Clepet, C.; Casagrande, A.; Choisne, N.; Aubourg, S.; Vitulo, N.; Jubin, C.; et al. The grapevine genome sequence suggests ancestral hexaploidization in major angiosperm phyla. Nature 2007, 449, 463–467. [Google Scholar]
- Kim, D.; Langmead, B.; Salzberg, S.L. HISAT: A fast spliced aligner with low memory requirements. Nat. Methods 2015, 12, 357–360. [Google Scholar] [CrossRef] [Green Version]
- Trapnell, C.; Williams, B.A.; Pertea, G.; Mortazavi, A.; Kwan, G.; van Baren, M.J.; Salzberg, S.L.; Wold, B.J.; Pachter, L. Transcript assembly and quantification by RNA-Seq reveals unannotated transcripts and isoform switching during cell differentiation. Nat. Biotechnol. 2010, 28, 511–515. [Google Scholar] [CrossRef] [Green Version]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [Green Version]
- Young, M.D.; Wakefield, M.J.; Smyth, G.K.; Oshlack, A. Gene ontology analysis for RNA-seq: Accounting for selection bias. Genome Biol. 2010, 11, R14. [Google Scholar] [CrossRef] [Green Version]
- Mao, X.; Cai, T.; Olyarchuk, J.G.; Wei, L. Automated genome annotation and pathway identification using the KEGG Orthology (KO) as a controlled vocabulary. Bioinformatics 2005, 21, 3787–3793. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Sitko, K.; Rusinowski, S.; Pogrzeba, M.; Daszkowska-Golec, A.; Gieroń, Ż.; Kalaji, H.; Małkowski, E. Development and aging of photosynthetic apparatus of Vitis vinifera L. during growing season. Photosynthetica 2020, 58, 186–193. [Google Scholar] [CrossRef]
- Yang, J.C.; Zhang, J.H.; Wang, Z.Q.; Zhu, Q.S.; Wang, W. Hormonal changes in the grains of rice subjected to water stress during grain filling. Plant J. 2001, 127, 315–323. [Google Scholar] [CrossRef] [Green Version]
- Chen, W.K.; Yu, K.J.; Liu, B.; Lan, Y.B.; Sun, R.Z.; Li, Q.; He, F.; Pan, Q.H.; Duan, C.Q.; Wang, J. Comparison of transcriptional expression patterns of carotenoid metabolism in ‘Cabernet Sauvignon’ grapes from two regions with distinct climate. J. Plant Physiol. 2017, 213, 75–86. [Google Scholar] [CrossRef]
- Ohama, N.; Sato, H.; Shinozaki, K.; Yamaguchi-Shinozaki, K. Transcriptional regulatory network of plant heat stress response. Trends Plant Sci. 2017, 22, 53–65. [Google Scholar] [CrossRef]
- Ju, Y.; Min, Z.; Zhang, Y.; Zhang, K.-k.; Liu, M.; Fang, Y. Transcriptome profiling provide new insights into the molecular mechanism of grapevine response to heat, drought, and combined stress. Sci. Hortic. 2021, 286, 110076. [Google Scholar] [CrossRef]
- Liu, M.; Ju, Y.; Min, Z.; Fang, Y.; Meng, J. Transcriptome analysis of grape leaves reveals insights into response to heat acclimation. Sci. Hortic. 2020, 272, 109554. [Google Scholar] [CrossRef]
- Coate, J.E.; Doyle, J.J. Quantifying whole transcriptome size, a prerequisite for understanding transcriptome evolution across species: An example from a plant allopolyploid. Genome Biol. Evol. 2010, 2, 534–546. [Google Scholar] [CrossRef] [Green Version]
- Pirrello, J.; Deluche, C.; Frangne, N.; Gévaudant, F.; Maza, E.; Djari, A.; Bourge, M.; Renaudin, J.-P.; Brown, S.; Bowler, C.; et al. Transcriptome profiling of sorted endoreduplicated nuclei from tomato fruits: How the global shift in expression ascribed to DNA ploidy influences RNA-Seq data normalization and interpretation. Plant J. 2018, 93, 387–398. [Google Scholar] [CrossRef] [Green Version]
- Sharma, A.; Kumar, V.; Shahzad, B.; Ramakrishnan, M.; Singh Sidhu, G.P.; Bali, A.S.; Handa, N.; Kapoor, D.; Yadav, P.; Khanna, K.; et al. Photosynthetic response of plants under different abiotic stresses: A Review. J. Plant Growth Regul. 2020, 39, 509–531. [Google Scholar] [CrossRef]
- Li, Y.T.; Xu, W.W.; Ren, B.Z.; Zhao, B.; Zhang, J.W.; Liu, P.; Zhang, Z.S. High temperature reduces photosynthesis in maize leaves by damaging chloroplast ultrastructure and photosystem II. J. Agron. Crop Sci. 2020, 00, 1–17. [Google Scholar] [CrossRef]
- Allario, T.; Brumos, J.; Colmenero-Flores, J.M.; Iglesias, D.J.; Pina, J.A.; Navarro, L.; Talon, M.; Ollitrault, P.; Morillon, R. Tetraploid Rangpur lime rootstock increases drought tolerance via enhanced constitutive root abscisic acid production. Plant Cell Environ. 2013, 36, 856–868. [Google Scholar] [CrossRef]
- Matsui, A.; Ryugo, K.; Kliewer, W.M. Lowered berry quality due to heat stress at the early ripening stage of berry growth in a seeded grapevine, Vitis vinifera L. Res. Bull. Fac. Agric. Gifa Univ. 1991, 56, 139–145. [Google Scholar]
- Godde, D.; Bormnan, J.F. Regulation of photosynthesis in higher plants. In Molecular to Global Photosynthesis; Archer, M.D., Barber, J., Eds.; Imperial College Press: London, UK, 2004; pp. 221–286. [Google Scholar]
- Salvucci, M.E.; Crafts-Brandner, S.J. Relationship between the heat tolerance of photosynthesis and the thermal stability of rubisco activase in plants from contrasting thermal environments. Plant Physiol. 2004, 134, 1460–1470. [Google Scholar] [CrossRef] [Green Version]
- Way, D.A.; Oren, R. Differential responses to changes in growth temperature between trees from different functional groups and biomes: A review and synthesis of data. Physiology 2010, 30, 669–688. [Google Scholar] [CrossRef] [Green Version]
- Feng, B.; Liu, P.; Li, G.; Dong, S.T.; Wang, F.H.; Kong, L.A.; Zhang, J.W. Effect of Heat Stress on the Photosynthetic characteristics in flag leaves at the grain-filling stage of different heat-resistant winter wheat varieties. J. Agron. Crop Sci. 2014, 200, 143–155. [Google Scholar] [CrossRef]
- Bernardo, S.; Dinis, L.-T.; Machado, N.; Moutinho-Pereira, J. Grapevine abiotic stress assessment and search for sustainable adaptation strategies in Mediterranean-like climates. A review. Agron. Sustain. Dev. 2018, 38, 66. [Google Scholar] [CrossRef] [Green Version]
- Brestic, M.; Zivcak, M.; Olsovska, K.; Kalaji, H.M.; Shao, H.B.; Hakeem, K.R. Heat signaling and stress responses in photosynthesis. In Plant Signaling: Understanding the Molecular Crosstalk; Hakeem, K.R., Rehman, R.U., Tahir, I., Eds.; Springer: New Delhi, India, 2014; pp. 241–256. [Google Scholar]
- Kun, Z.; Bai-hong, C.; Yan, H.; Rui, Y.; Yu-an, W. Effects of short-term heat stress on PSII and subsequent recovery for senescent leaves of Vitis vinifera L. cv. Red Globe. J. Integr. Agr. 2018, 17, 2683–2693. [Google Scholar]
- Ifuku, K.; Ido, K.; Sato, F. Molecular functions of PsbP and PsbQ proteins in the photosystem II supercomplex. J. Photochem. Photobiol. B Biol. 2011, 104, 158–164. [Google Scholar] [CrossRef] [PubMed]
- Sakata, S.; Mizusawa, N.; Kubota-Kawai, H.; Sakurai, I.; Wada, H. Psb28 is involved in recovery of photosystem II at high temperature in Synechocystis sp. PCC 6803. B.B.A. Bioenerg. 2013, 1827, 50–59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, L.J.; Loescher, W.; Duan, W.; Li, W.D.; Yang, S.H.; Li, S.H. Heat acclimation induced acquired heat tolerance and cross adaptation in different grape cultivars: Relationships to photosynthetic energy partitioning. Funct. Plant Biol. 2009, 36, 516–526. [Google Scholar] [CrossRef] [PubMed]
- Roháček, K. Chlorophyll fluorescence parameters: The definitions, photosynthetic meaning, and mutual relationships. Photosynthetica 2002, 40, 13–29. [Google Scholar] [CrossRef]
- Zhang, Q.C.; Li, Y.; Xu, W.G.; Zhang, Y.; Qi, X.L.; Fang, Y.H.; Peng, C.J. Joint expression of Zmpepc, Zmppdk, and Zmnadp-me is more efficient than expression of one or two of those genes in improving the photosynthesis of Arabidopsis. Plant Physiol. Biochem. 2021, 158, 410–419. [Google Scholar] [CrossRef]
- Cazzonelli, C.I.; Pogson, B.J. Source to sink: Regulation of carotenoid biosynthesis in plants. Trends Plant Sci. 2010, 5, 266–274. [Google Scholar] [CrossRef]
- Guzman, I.; Hamby, S.; Romero, J.; Bosland, P.W.; O’Connell, M.A. Variability of carotenoid biosynthesis in orange colored Capsicum spp. Plant Sci. 2010, 179, 49–59. [Google Scholar] [CrossRef] [Green Version]
- Sugiyama, A.; Ikoma, Y.; Fujii, H.; Shimada, T.; Endo, T.; Shimizu, T.; Omura, M. Structure and expression levels of alleles of Citrus zeaxanthin epoxidase genes. J. Jpn. Soc. Hort. Sci. 2010, 79, 263–274. [Google Scholar] [CrossRef] [Green Version]
- Zhang, M.; Leng, P.; Zhang, G.L.; Li, X.X. Cloning and functional analysis of 9-cis-epoxycarotenoid dioxygenase (NCED) genes encoding a key enzyme during abscisic acid biosynthesis from peach and grape fruits. J. Plant Physiol. 2009, 166, 1241–1252. [Google Scholar] [CrossRef]
- Seo, M.; Koshiba, T. Complex regulation of ABA biosynthesis in plants. Trends Plant Sci. 2002, 7, 41–48. [Google Scholar] [CrossRef]
- Boneh, U.; Biton, I.; Schwartz, A.; Ben-Ari, G. Characterization of the ABA signal transduction pathway in Vitis vinifera. Plant Sci. 2012, 187, 89–96. [Google Scholar] [CrossRef]
- Lee, S.C.; Luan, S. ABA signal transduction at the crossroad of biotic and abiotic stress responses. Plant Cell Environ. 2012, 35, 53–60. [Google Scholar] [CrossRef]
- Klingler, J.P.; Batelli, G.; Zhu, J.K. ABA receptors: The START of a new paradigm in phytohormone signalling. J. Exp. Bot. 2010, 61, 3199–3210. [Google Scholar] [CrossRef] [Green Version]
- Schroeder, J.I.; Allen, G.J.; Hugouvieux, V.; Kwak, J.M.; Waner, D. Guard cell signal transduction. Annu. Rev. Plant Physiol. Plant Biol. 2001, 52, 627–658. [Google Scholar] [CrossRef] [Green Version]
- Sun, J.; Ren, L.P.; Cheng, Y.; Gao, J.J.; Dong, B.; Chen, S.M.; Chen, F.D.; Jiang, J.F. Identification of differentially expressed genes in Chrysanthemum nankingense (Asteraceae) under heat stress by RNA Seq. Gene 2014, 552, 59–66. [Google Scholar] [CrossRef]
- Bokszczanin, K.L.; Fragkostefanakis, S. Perspectives on deciphering mechanisms underlying plant heat stress response and thermotolerance. Front. Plant Sci. 2013, 4, 315. [Google Scholar] [CrossRef]
- Kotak, S.; Larkindale, J.; Lee, U.; Koskull-Doring, P.V.; Vierling, E.; Scharf, K.D. Complexity of the heat stress response in plants. Curr. Opin. Plant Biol. 2007, 10, 310–316. [Google Scholar] [CrossRef]
- Saidi, Y.; Finka, A.; Goloubinoff, P. Heat perception and signalling in plants: A tortuous path to thermotolerance. New Phytol. 2011, 190, 556–565. [Google Scholar] [CrossRef]
- Huang, B.R.; Xu, C.P. Identification and characterization of proteins associated with plant tolerance to heat stress. J. Integr. Plant Biol. 2008, 50, 1230–1237. [Google Scholar] [CrossRef]
- Wahid, A.; Gelani, S.; Ashraf, M.; Foolad, M.R. Heat tolerance in plants: An overview. Environ. Exp. Bot. 2007, 61, 199–223. [Google Scholar] [CrossRef]
- Heckathorn, S.A.; Downs, C.A.; Sharker, T.D.; Coleman, J.S. The small, methionine-rich chloroplast heat-shock protein protects photosystem II electron transport during heat stress. Plant Physiol. 1998, 116, 439–444. [Google Scholar] [CrossRef] [Green Version]
- Sanmiya, K.; Suzuki, K.; Egawa, Y.; Shono, M. Mitochondrial small heat-shock protein enhances thermotolerance in tobacco plants. FEBS Lett. 2004, 557, 265–268. [Google Scholar] [CrossRef] [Green Version]
- Chuang, H.; Khurana, N.; Nijhavan, A.; Khurana, J.P.; Khurana, P. The wheat chloroplastic small heat shock protein (sHSP26) is involved in seed maturation and germination and imparts tolerance to heat stress. Plant Cell Environ. 2012, 35, 1912–1931. [Google Scholar]
- Fang, L.C.; Tong, J.; Dong, Y.F.; Xu, D.Y.; Mao, J.; Zhou, Y. De novo RNA sequencing transcriptome of Rhododendron obtusum identified the early heat response genes involved in the transcriptional regulation of photosynthesis. PLoS ONE 2017, 12, e0186376. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.M.; Chen, S.Y.; Shi, X.; Liu, D.N.; Zhao, P.; Lu, Y.Z.; Cheng, Y.B.; Liu, Z.S.; Nie, X.J.; Song, W.N.; et al. Hybrid sequencing reveals insight into heat sensing and signaling of bread wheat. Plant J. 2019, 98, 1015–1032. [Google Scholar] [CrossRef]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guo, R.; Lin, L.; Huang, G.; Shi, X.; Wei, R.; Han, J.; Zhou, S.; Zhang, Y.; Xie, T.; Bai, X.; et al. Transcriptome Analysis of ‘Kyoho’ Grapevine Leaves Identifies Heat Response Genes Involved in the Transcriptional Regulation of Photosynthesis and Abscisic Acid. Agronomy 2022, 12, 2591. https://doi.org/10.3390/agronomy12102591
Guo R, Lin L, Huang G, Shi X, Wei R, Han J, Zhou S, Zhang Y, Xie T, Bai X, et al. Transcriptome Analysis of ‘Kyoho’ Grapevine Leaves Identifies Heat Response Genes Involved in the Transcriptional Regulation of Photosynthesis and Abscisic Acid. Agronomy. 2022; 12(10):2591. https://doi.org/10.3390/agronomy12102591
Chicago/Turabian StyleGuo, Rongrong, Ling Lin, Guiyuan Huang, Xiaofang Shi, Rongfu Wei, Jiayu Han, Sihong Zhou, Ying Zhang, Taili Xie, Xianjin Bai, and et al. 2022. "Transcriptome Analysis of ‘Kyoho’ Grapevine Leaves Identifies Heat Response Genes Involved in the Transcriptional Regulation of Photosynthesis and Abscisic Acid" Agronomy 12, no. 10: 2591. https://doi.org/10.3390/agronomy12102591