
Ups and Downs of the Italian Locust (Calliptamus italicus L.) Populations in the Siberian Steppes: On the Horns of Dilemmas


Abstract
1. Introduction
2. Materials and Methods
2.1. Study Territory
2.2. Observation Sites
- SE Aleksandrovskij settlement (Novosibirskaya Oblast (Novosibirsk Region), 53.67° N 78.25° E, northern steppe, studied in 2000–2008, 2015, 2018, 2019; in 2003 the local control, i.e., untreated, plot was moved about 100 m southwards, because the main part of the area was plowed) (Figure 1A).
- S Burla settlement (Altai Krai (Altaj Region), 53.23° N 78.43° E, old abandoned field, 2000–2008).
- SW Yarovoe town (Altai Krai (Altaj Region), 52.85° N 78.57° E, dry steppe (actually very old crested wheatgrass field), 1979, 1992, 1999–2008, 2015).
- E Platovka settlement (Altai Krai (Altaj Region), 52.40° N 79.05° E, dry steppe, 1999–2008, 2015).
- SE Severka settlement (Altai Krai (Altaj Region), 52.12° N 79.32° E, typical steppe, 1999–2008, 2015).
- S Pokrovka settlement (Altai Krai (Altaj Region), 52.00° N 79.37° E, typical steppe, 1999–2008, 2015).
- NW Mikhaylovskoye settlement (Altai Krai (Altaj Region), 51.82° N 79.62° E, typical steppe, 1999–2008, 2015; in 2003, we counted locusts on the adjacent steppe plot, 51.83° N 79.58° E, because we could not visit the stationary plot due to some technical problems).
- W Bor-Forpost settlement (Altai Krai (Altaj Region), 51.87° N 80.03° E, typical steppe (actually very old crested wheatgrass field), 2000–2009, 2015).
- E Ust-Volchikha settlement (Altai Krai (Altaj Region), 51.93° N 80.28° E, dry steppe, 1999–2009, 2015, 2018) (Figure 1B).
2.3. Experimental Area
2.4. Field Studies
2.5. Data Analysis
3. Results
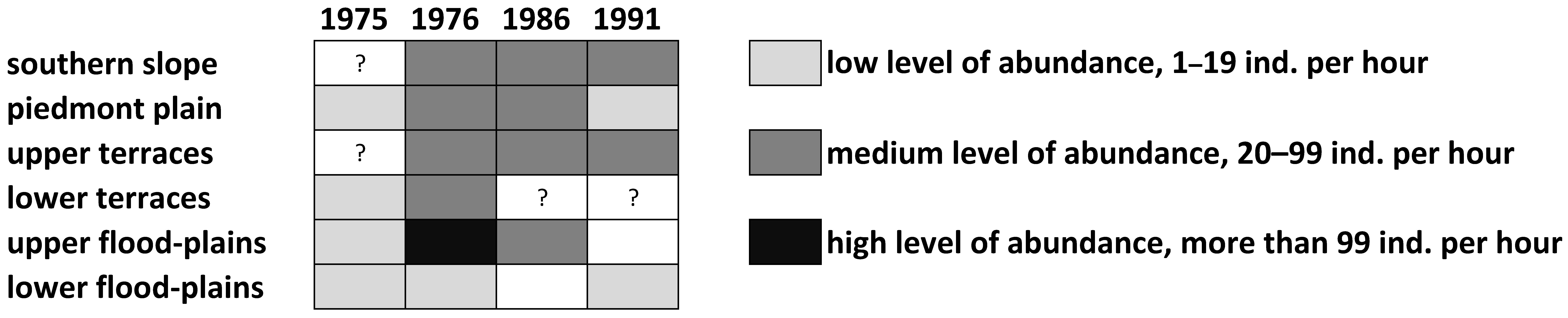
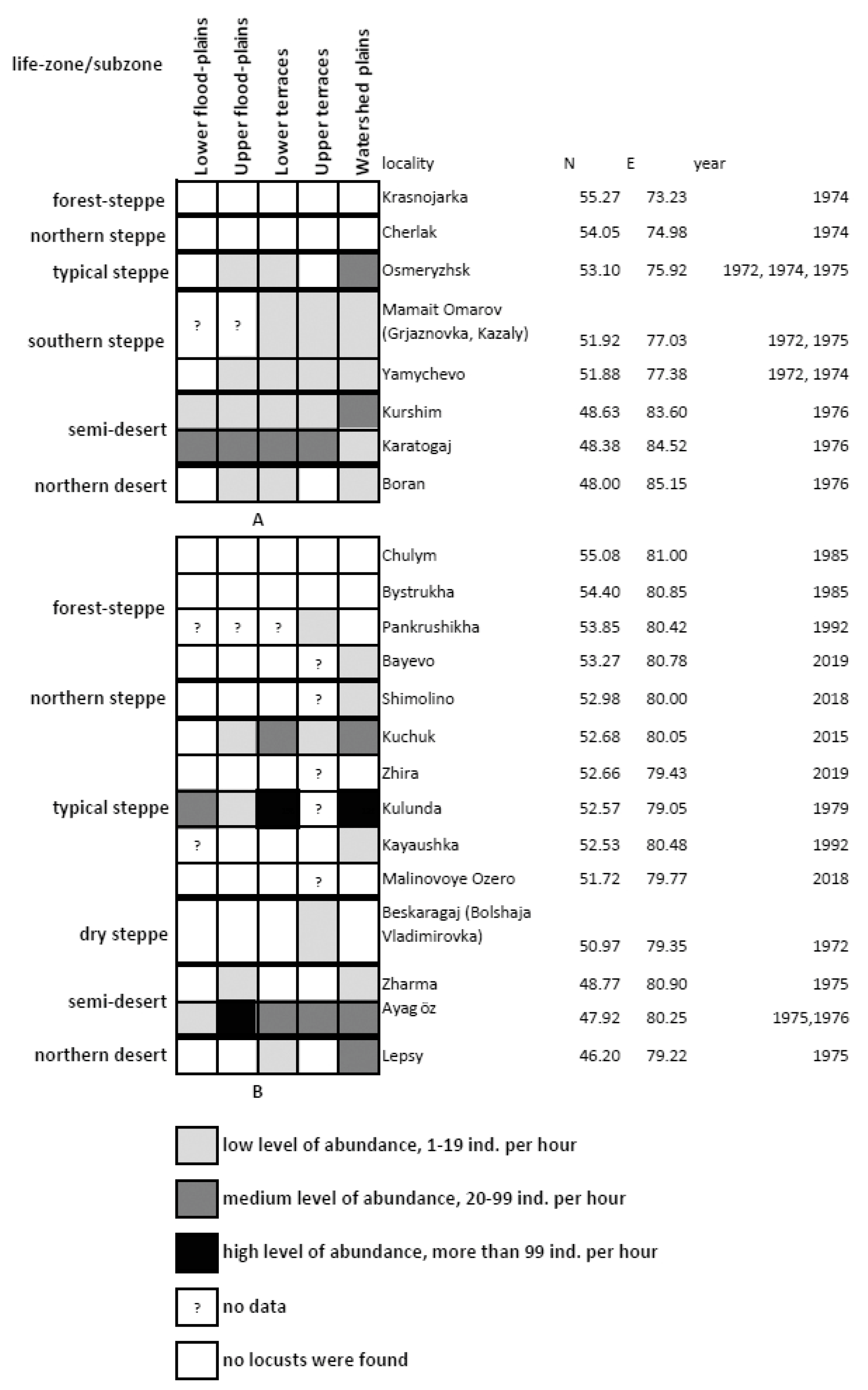
3.1. Distribution Patterns of the Italian Locust over the South-Eastern Part of West Siberian Plain and the Adjacent North-Eastern and Eastern Parts of the Kazakh Uplands
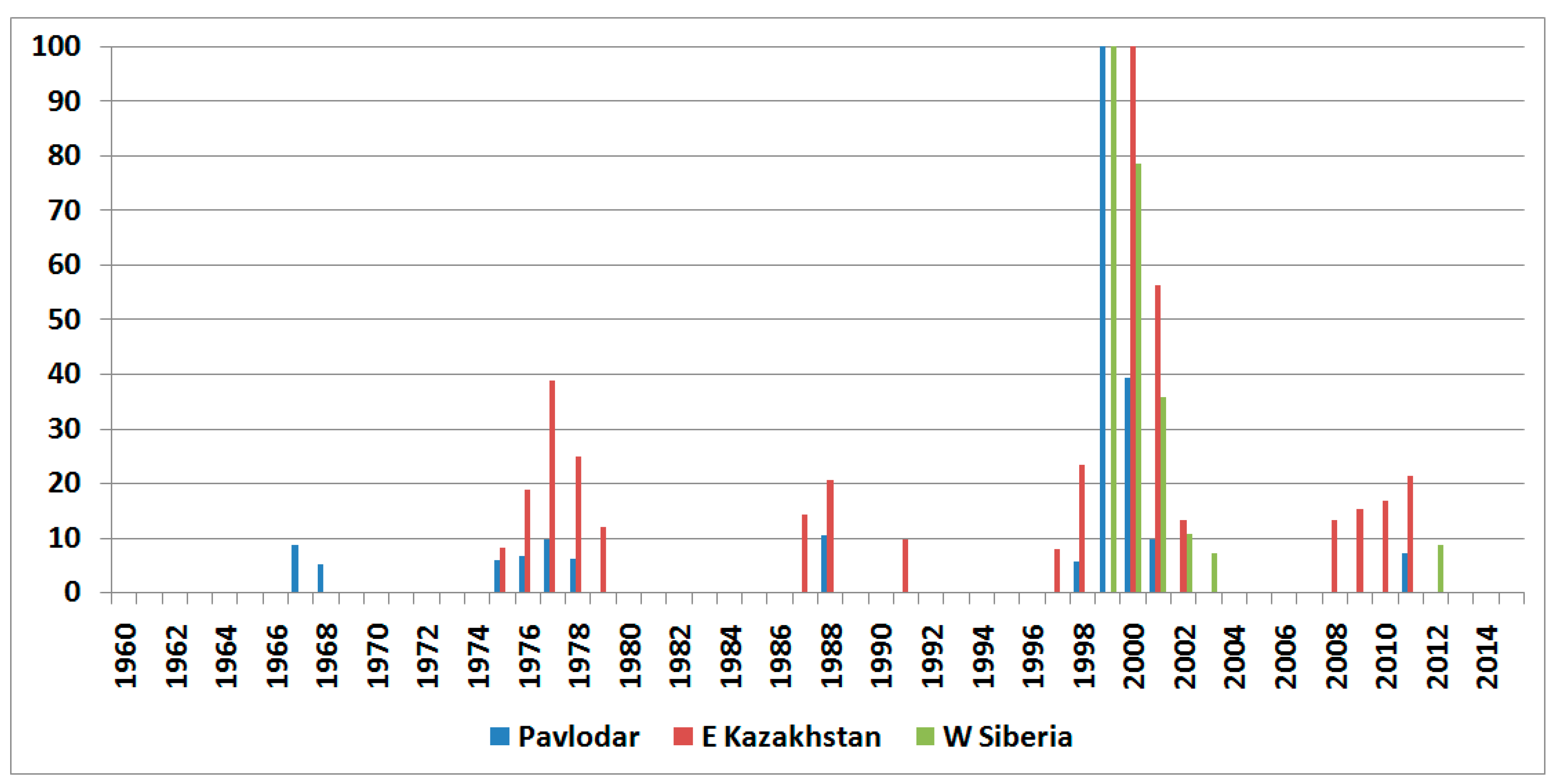
3.2. Long-Term Dynamics of the Italian Locust in the Central and Eastern Parts of Its Range: General Patterns
3.3. Peculiarities of Long-Term Dynamics of the Italian Locust in the South-Eastern Part of West Siberian Plain and in the Adjacent Parts of the Kazakh Uplands
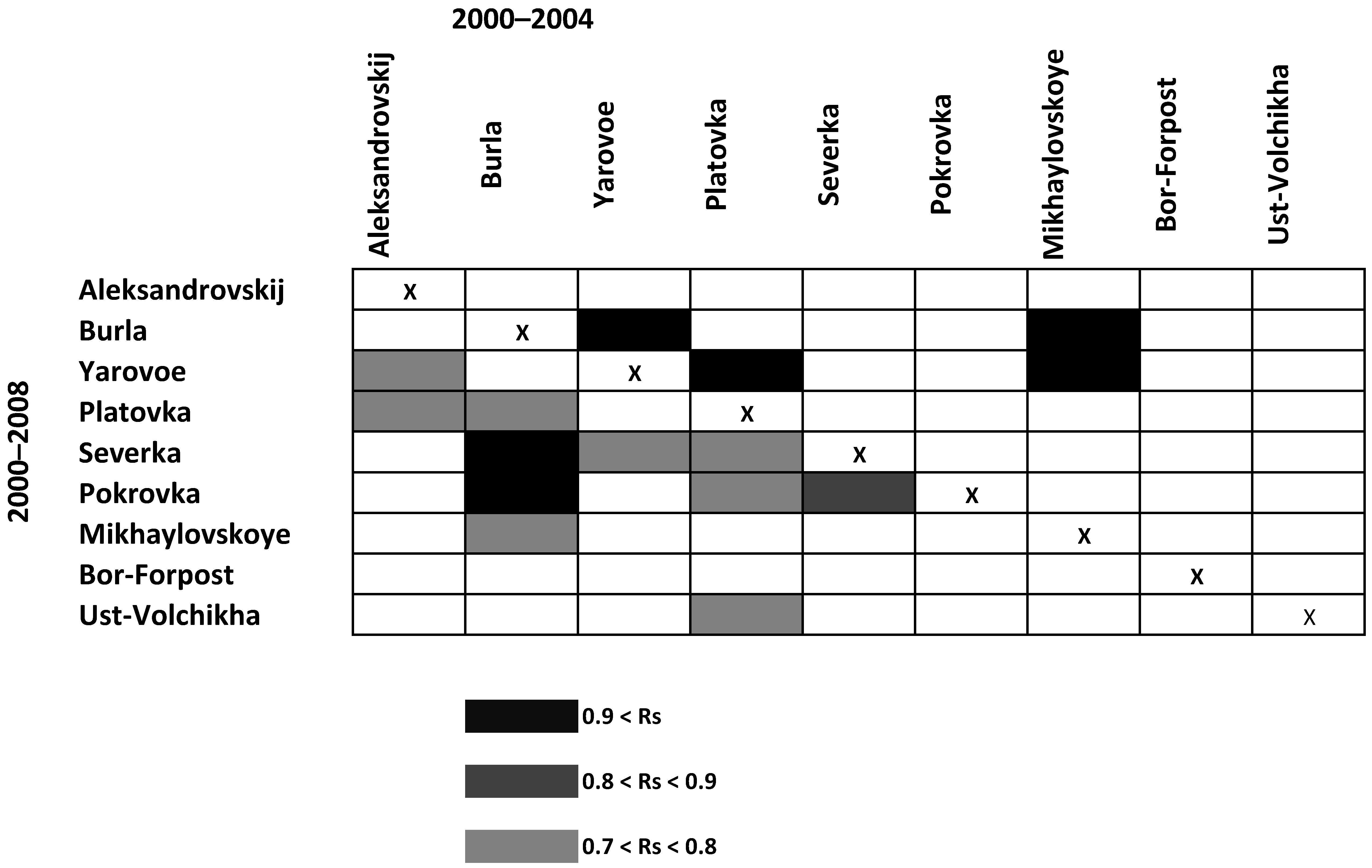
3.4. Dynamics of the Italian Locust Populations in the Kulunda Steppe in 2000–2008
- (1)
- A recession, when adults’ abundance in a local population in the middle and the end of a summer varies between relatively low and moderate (according our estimation, in the Kulunda steppe, from 0.1 up to 2.0 ind. per m2, perhaps in some cases—up to 3.2). This means that recommendations of some published manuals concerning monitoring and management of pest locusts and grasshoppers [40,41] to organize partial treatments in the next year, when the density of the Italian locust adults is more than 1 ind. per m2, are based on too conservative estimations.
- (2)
- A depression, when density is very low, commonly significantly less than 0.1 ind. per m2. In this case, a population may be stable, but scarce locusts are distributed very sporadically. For instance, in 31 July 2019, I spent about two hours on our main model site in the northern part of the Kulunda steppe (near Aleksandrovskij) finding only 10 adults of the Italian locust and estimated their average density as about 25 ind. per ha (Figure 2).
3.5. Seasonal Dynamics and Distribution of the Italian Locust Local Population in the Northern Steppe in 2000–2019
3.6. The Italian Locust and Rare Orthopteran Species
4. Discussion
4.1. May We Use for the Italian Locust Approaches and Technologies Developed for the Desert and Migratory Locusts or Not?
4.2. Monitoring, Management, and Forecasting: Is the Regional Level of Exploration Enough or Not?
4.3. Monitoring, Management, and Forecasting: When Should We Begin (or End) to Trouble or When Not?
4.4. Treatments: When Do Barriers Work and When Not?
4.5. Treatments: Can We Preserve Local Populations of Rare Orthoptera or Not?
5. Conclusions
- (1)
- Crucial parts of the range, habitats, and time lapses should be determined for the species across all species range, each region, and landscape.
- (2)
- New approaches have to be elaborated to manage populations on the landscape scale and to treat agricultural fields and rangelands.
- (3)
- Agricultural technologies per se ought to be modified significantly. For instance, in grasslands, overgrazing should be minimized, and restoration of abandoned fields that commonly harbor C. italicus should be accelerated.
- (4)
- (5)
- New approaches should be very important for field assessments of the Italian locust populations as well. For instance, real time observations by using unmanned aerial vehicles are very hopeful.
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Latchininsky, A.V.; Sergeev, M.G.; Childebaev, M.K.; Chernijakhovskij, M.E.; Lockwood, J.A.; Kambulin, V.E.; Gapparov, F.A. The Grasshoppers of Kazakhstan, Middle Asia and Adjacent Territories; Association for Applied Acridology International & University of Wyoming: Laramie, WY, USA, 2002; p. vii + 387. (In Russian) [Google Scholar]
- Zhang, L.; Lecoq, M.; Latchininsky, A.; Hunter, D. Locust and grasshopper management. Ann. Rev. Entomol. 2019, 64, 15–34. [Google Scholar] [CrossRef]
- Uvarov, B.P. Grasshoppers and Locusts; Centre for Overseas Pest Research: London, UK, 1977; Volume 2, p. 613. [Google Scholar]
- Sergeev, M.G.; Denisova, O.V.; Vanjkova, I.A. How do spatial population structures affect acridid management? In Grasshoppers and Grassland Health: Managing Grasshopper Outbreaks Without Risking Environmental Disaster; Lockwood, J.A., Latchininsky, A.V., Sergeev, M.G., Eds.; Kluwer Academic Publishers: Dordrecht, The Netherlands; Boston, MA, USA; London, UK, 2000; pp. 71–87. [Google Scholar] [CrossRef]
- Vasil’ev, K.A. The Italian locust (Calliptamus italicus L.) in Central Kazakhstan. Trudy Nauchno-Issledovatelskogo Inst. Zastshity Rastenij 1962, 7, 123–190. (In Russian) [Google Scholar]
- Bunin, L.D. The Italian Locust (Calliptamus italicus L.) in the Soil-Saving Agriculture Zone in East. Kazakhstan and Development of Treatments. Ph.D. Thesis, VIZR, Leningrad, Russia, 1979; p. 24. (In Russian). [Google Scholar]
- Azhbenov, V.K. Outbreaks and migrations of locusts in Kazakhstan. Steppe Bull. 2000, 6, 16–20. (In Russian) [Google Scholar]
- Vasil’ev, K.A. Phases of the Italian locust (Calliptamus italicus L.). Proc. USSR Acad. Sci. 1950, 74, 639–642. (In Russian) [Google Scholar]
- Sergeev, M.G.; Childebaev, M.K.; Van’kova, I.A.; Gapparov, F.A.; Kambulin, V.E.; Kokanova, E.; Latchininsky, A.V.; Pshenitsyna, L.B.; Temreshev, I.I.; Chernijakhovskij, M.E.; et al. The Italian Locust [Calliptamus italicus (Linnaeus, 1758)]: Morphology, Distribution, Ecology, Population Management, Draft Version; FAO: Rome, Italy, 2016; p. 330. Available online: http://www.fao.org/locusts-cca/bioecology/italian-locust-cit/en/ (accessed on 21 December 2020). (In Russian)
- Stolyarov, M.V. Massenvermehrungen von Calliptamus italicus L. in Südrussland im zwanzigsten Jahrhundert. Articulata 2000, 15, 99–108. [Google Scholar]
- Stolyarov, M.V. Cyclicity and some characteristics of mass reproduction of Calliptamus italicus L. in Southern Russia. Russ. J. Ecol. 2000, 31, 43–48. [Google Scholar] [CrossRef]
- Sergeev, M.G.; Van’kova, I.A. Zonal-landscape distribution of population groups of Calliptamus italicus L. (Insecta, Orthoptera, Acrididae). Sib. J. Ecol. 1996, 3, 219–225. [Google Scholar]
- Isachenko, A.G. Landscapes of the USSR; Leningrad University Publ.: Leningrad, Russia, 1985; p. 320. (In Russian) [Google Scholar]
- Smelansky, I.E.; Tishkov, A.A. The steppe biome in Russia: Ecosystem services, conservation status, and actual challenges. In Eurasian Steppes. Ecological Problems and Livelihoods in A Changing World; Werger, M.J.A., van Staalduinen, M.A., Eds.; Plant and Vegetation; Springer Science+Business Media, B.V.: Dordrecht, The Netherlands, 2012; Volume 6, pp. 45–101. [Google Scholar] [CrossRef]
- Chibilev, A.A. Steppe Eurasia: A Regional Review of Natural Diversity; Institute of Steppe and RGS: Moscow, Russia; Orenburg, Russia, 2017; p. 324. (In Russian) [Google Scholar]
- Sergeev, M.G.; Van’kova, I.A. The dynamics of a local population of the Italian locust (Calliptamus italicus L.) in an anthropogenic landscape. Contemp. Probl. Ecol. 2008, 1, 88–95. [Google Scholar] [CrossRef]
- Sergeev, M.G. Patterns of Orthoptera Distribution in North Asia; Nauka Publ.: Novosibirsk, Russia, 1986; p. 237. (In Russian) [Google Scholar]
- Gause, G.F. Studies on the ecology of the Orthoptera. Ecology 1930, 11, 307–325. [Google Scholar] [CrossRef]
- Sergeev, M.G.; Van’kova, I.A. Dynamics of the Italian locust Calliptamus italicus L. population in the southeast of the West Siberian Plain. Contemp. Probl. Ecol. 2008, 1, 204–209. [Google Scholar] [CrossRef]
- Sergeev, M.G. Distribution patterns of grasshoppers and their kin over the Eurasian Steppes. Insects 2021, 12, 77. [Google Scholar] [CrossRef] [PubMed]
- Henderson, C.F.; Tilton, E.W. Tests with acaricides against the brown wheat mite. J. Econ. Entomol. 1955, 48, 157–161. [Google Scholar] [CrossRef]
- Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. PAST: Paleontological statistics software package for education and data analysis. Palaeontol. Electron. 2001, 4, 9. [Google Scholar]
- Ili’ina, I.S.; Lapshina, E.I.; Lavrenko, N.N.; Meltser, L.I.; Romanova, E.A.; Bogojavlenskij, B.A.; Mahno, V.D. Vegetation of West Siberian Plain; Nauka Pub.: Novosibirsk, Russia, 1985; p. 251. (In Russian) [Google Scholar]
- Sergeev, M.G. Metapopulations of locusts and grasshoppers: Spatial structures, their dynamics and early warning systems. In New Strategies in Locust Control; Krall, S., Peveling, R., Ba Diallo, D., Eds.; Birkhauser Verlag: Basel, Switzerland, 1997; pp. 75–80. [Google Scholar] [CrossRef]
- Bugaev, G.S. Habitat distribution of grasshoppers in the zone of band pine forests in North-East Kazakhstan. Vestn. Selskokhoziastvennoj Nauk. Kazakhstana 1977, 6, 37–40. (In Russian) [Google Scholar]
- Bey-Bienko, G.J. The zonal and ecological distribution of Acrididae in West Siberian and Zaisan Plains. Tr. po Zastshite Rasteniy Ser. Entomol. 1930, 1, 51–90. (In Russian) [Google Scholar]
- Lindemann, K. The Italian Locust in Voronezh Province; V.I. Isaev Publ.: Voronezh, Russia, 1892; p. 52. (In Russian) [Google Scholar]
- Filip’ev, I.N. Common pests. Grasshoppers. Acridoidea. Tr. po Prikl. Entomol. 1926, 13, 57–176. (In Russian) [Google Scholar]
- Predtechensky, S.A.; Zhdanov, S.P.; Popova, A.A. Pest locusts in the USSR. Tr. po Zastshite Rastenij. Ser. Entomol. 1935, 18, 1–168. (In Russian) [Google Scholar]
- Berezhkov, R.P. The Grasshoppers of West Siberia; Tomsk University Publ.: Tomsk, Russia, 1956; p. 174. (In Russian) [Google Scholar]
- Borisenkov, E.P.; Pasetskij, V.M. Thousand-Year Chronicle of Extraordinary Natural Phenomena; Mysl Publ.: Moscow, Russia, 1988; p. 524. (In Russian) [Google Scholar]
- Govorov, D.N.; Zhivykh, A.V. (Eds.) The Report on Phytosanitary Conditions of Crops in the Russian Federation in 2018 and Pest Forecasts for 2019; Rosselkhozcenter: Moscow, Russia, 2019; p. 900. (In Russian) [Google Scholar]
- Govorov, D.N.; Zhivykh, A.V. (Eds.) The Report on Phytosanitary Conditions of Crops in the Russian Federation in 2019 and Pest Forecasts for 2020; Rosselkhozcenter: Moscow, Russia, 2020; p. 897. (In Russian) [Google Scholar]
- Kushalieva, S.A.; Avtaeva, T.A. Peculiarities of the Italian locust (Calliptamus italicus) ecology in the semi-desert life zone of the Chechen Republic. Her. Kursk State Agric. Acad. 2019, 6, 73–78. (In Russian) [Google Scholar]
- Kamynina, L.D. Locusts in the Altai Region. Steppe Bull. 2000, 6, 9–10. (In Russian) [Google Scholar]
- Chetyrkina, I.A. The Italian locust Calliptamus italicus L. in East Kazakhstan. Horae Soc. Entomol. Unionis Sov. 1958, 46, 5–67. (In Russian) [Google Scholar]
- Raevskij, V.G. Observation of the Italian locust (Calliptamus italicus L.) in the Slavgorod county in 1922. Rep. Sib. Entomol. Bur. 1924, 2, 53–58. (In Russian) [Google Scholar]
- Troitskij, D. Pest grasshoppers of the Semipalatinsk Region in 1912. Needs West. Sib. Agric. 1914, 1, 23–49. (In Russian) [Google Scholar]
- Tsyplenkov, E.P. Pest. Grasshoppers in the USSR; Kolos Publ.: Leningrad, Russia, 1970; p. 271. (In Russian) [Google Scholar]
- Naumovich, O.N.; Stolyarov, M.V.; Dolzhenko, V.I.; Nikulin, A.A.; Alekhin, V.T. Recommendation on Monitoring and Management of Pest Grasshoppers; Kolos: Moscow, Russia, 2000; p. 21. (In Russian) [Google Scholar]
- Dolzhenko, V.I.; Naumovich, O.N.; Nikulin, A.A. Pest Grasshoppers; Plant Protection and Quarantine: Moscow, Russia, 2003; p. 28. (In Russian) [Google Scholar]
- Kovalenkov, V.G.; Kuznetsova, O.V. How the Italian locust outbreak may be stopped. Plant. Prot. Quar. 2011, 9, 14–17. (In Russian) [Google Scholar]
- Sergeev, M.G.; Latchininsky, A.V.; Duranton, J.-F. Possibilities of fipronil (Adonis) application in Siberia. Plant. Prot. Quar. 2002, 3, 19–20. (In Russian) [Google Scholar]
- IUCN. The IUCN Red List of Threatened Species. Version 2020-3. 2021. Available online: https://www.iucnredlist.org (accessed on 15 January 2021).
- Popova, K.V.; Molodtsov, V.V.; Sergeev, M.G. Rare grasshoppers (Orthoptera, Acridoidea) of the Baraba and Kulunda steppes (South Siberia). Acta Biol. Sib. 2020, 6, 595–609. [Google Scholar] [CrossRef]
- Le Gall, M.; Overson, R.; Cease, A. A global review on locusts (Orthoptera: Acrididae) and their interactions with livestock grazing practices. Front. Ecol. Evol. 2019, 7, 263. [Google Scholar] [CrossRef]
- Uvarov, B.P. Locusts and Grasshoppers; Promizdat: Moscow, Russia; Leningrad, Russia, 1927; p. 306. (In Russian) [Google Scholar]
- Hemming, C.F.; Popov, G.B.; Roffey, J.; Waloff, Z. Characteristics of desert locust plague upsurges. Phil. Trans. R. Soc. Lond. B 1979, 287, 375–386. [Google Scholar] [CrossRef]
- Magor, J.I. Locust glossary. In Desert Locust Control with Existing Techniques: An Evaluation of Strategies; van Huis, A., Ed.; Wageningen Agricultural University: Wageningen, The Netherlands, 1994; pp. 119–127. [Google Scholar]
- Magor, J.I.; Lecoq, M.; Hunter, D.M. Preventive control and desert locust plagues. Crop. Prot. 2008, 27, 1527–1533. [Google Scholar] [CrossRef]
- Zimmerman, K.; Lockwood, J.A.; Latchininsky, A.V. A spatial, Markovian model of rangeland grasshopper (Orthoptera; Acrididae) population dynamics: Do long-term benefits justify suppression of infestations? Environ. Entomol. 2004, 33, 257–266. [Google Scholar] [CrossRef][Green Version]
- Kazakova, I.G.; Sergeev, M.G. The problem of delimitation of grasshopper populations. Sib. J. Ecol. 1997, 4, 315–321. (In Russian) [Google Scholar]
- Lockwood, J.A. Rangeland grasshopper ecology. In The Bionomics of Grasshoppers, Katydids and Their Kin; Gangwere, S.K., Muralirangan, M.C., Muralirangan, M., Eds.; CAB International: Oxon, UK; NewYork, NY, USA, 1997; pp. 129–146. [Google Scholar]
- Joern, A. What are the consequences of non-liner ecological interactions for grasshopper control strategies? In Grasshoppers and Grassland Health: Managing Grasshopper Outbreaks Without Risking Environmental Disaster; Lockwood, J.A., Latchininsky, A.V., Sergeev, M.G., Eds.; Kluwer Academic Publishers: Dordrecht, The Netherlands; Boston, MA, USA; London, UK, 2000; pp. 131–144. [Google Scholar] [CrossRef]
- Uvarov, B.P. Grasshoppers and Locusts; Cambridge University Press: London, UK; New York, NY, USA; Ibadan, Nigeria, 1966; Volume 1, p. xi + 481. [Google Scholar]
- Lockwood, J.A.; Lockwood, D.R. Catastrophe theory: A unified paradigm for rangeland ecosystem dynamics. J. Range Manag. 1993, 46, 282–288. [Google Scholar] [CrossRef]
- van Huis, A. Can we prevent desert locust plagues? In New Strategies in Locust Control; Krall, S., Peveling, R., Ba Diallo, D., Eds.; Birkhauser Verlag: Basel, Switzerland, 1997; pp. 453–459. [Google Scholar] [CrossRef]
- Lockwood, J.A.; Schell, S.P. Decreasing economic and environmental costs through Reduced Area and Agent Insecticide Treatments (RAATs) for the control of rangeland grasshoppers: Empirical results and their implications for pest management. J. Orthoptera Res. 1997, 6, 19–32. [Google Scholar] [CrossRef]
- Latchininsky, A.V.; Lockwood, J.A.; Sergeev, M.G. Some experience to control grasshoppers in the North American prairies. Plant. Prot. Quar. 1999, 8, 12–14. (In Russian) [Google Scholar]
- Wilps, H.; Levchenko, V.; Vernigor, A. Field trials with imidacloprid (Confidor) on the Italian locust Calliptamus italicus in Kazakhstan. J. Appl. Entomol. 2002, 126, 436–443. [Google Scholar] [CrossRef]
- Sobolev, N.N.; Sergeev, M.G. Population dynamics of grasshoppers (Orthoptera, Acridoidea) in the agricultural ecosystems of North Kazakhstan. In Human Influences on Insects’ Assemblages; Zolotarenko, G.S., Ed.; Nauka Publ.: Novosibirsk, Russia, 1985; pp. 96–104. (In Russian) [Google Scholar]
- Sergeev, M.G. La sécheresse et les schémas de distribution des criquets en Asie centrale et septentrionale. Sécheresse 1996, 7, 129–132. [Google Scholar]
- Samways, M.J.; Lockwood, J.A. Orthoptera conservation: Pests and paradoxes. J. Insect Conserv. 1998, 2, 143–149. [Google Scholar] [CrossRef]
- Sergeev, M.G. Conservation of orthopteran biological diversity relative to landscape change in temperate Eurasia. J. Insect Conserv. 1998, 2, 247–252. [Google Scholar] [CrossRef]
- Samways, M.J. Can locust control be compatible with conserving biodiversity? In Grasshoppers and Grassland Health: Managing Grasshopper Outbreaks Without Risking Environmental Disaster; Lockwood, J.A., Latchininsky, A.V., Sergeev, M.G., Eds.; Kluwer Academic Publishers: Dordrecht, The Netherlands; Boston, MA, USA; London, UK, 2000; pp. 173–179. [Google Scholar] [CrossRef]
- Latchininsky, A.V.; Sergeev, M.G. The Italian locust—Calliptamus italicus (Linnaeus) (Orthoptera): A quarter of a century after. Metaleptea 2018, 38, 47–48. [Google Scholar]
- Latchininsky, A.V. Locusts and remote sensing: A review. J. Appl. Remote Sens. 2013, 7, 075099. [Google Scholar] [CrossRef]
- Tronin, A.A.; Gornyy, V.I.; Kiselev, A.V.; Kritsuk, S.G.; Latypov, I.S. Forecasting of locust mass breeding by using satellite data. Curr. Probl. Remote Sens. Earth Space 2014, 11, 37–50. [Google Scholar]
- Klein, I.; Oppelt, N.; Kuenzer, C. Application of remote sensing data for locust research and management—A review. Insects 2021, 12, 233. [Google Scholar] [CrossRef] [PubMed]
- Stebaev, I.V.; Gukasian, A.B. Orthoptera (Tettigonoidea and Acrididae) as stimulators of microbiological processes of decomposition and mineralization of phytomass in the meadow steppes of West Siberia. Zool. Zh. 1963, 42, 216–221. (In Russian) [Google Scholar]
- Gandar, M.V. The dynamics and trophic ecology of grasshoppers (Acridoidea) in a South African savanna. Oecologia 1982, 54, 370–378. [Google Scholar] [CrossRef] [PubMed]
- Hewitt, G.B.; Onsager, J.A. A method for forecasting potential losses from grasshopper feeding on northern mixed prairie forages. J. Range Manag. 1982, 35, 53–57. [Google Scholar] [CrossRef]
- Belovsky, G.E. Do grasshoppers diminish grassland productivity? In Grasshoppers and Grassland Health: Managing Grasshopper Outbreaks without Risking Environmental Disaster; Lockwood, J.A., Latchininsky, A.V., Sergeev, M.G., Eds.; Kluwer Academic Publishers: Dordrecht, The Netherlands; Boston, MA, USA; London, UK, 2000; pp. 7–29. [Google Scholar] [CrossRef]
- Pshenitsyna, L.B.; Sergeev, M.G.; Koutsenogij, K.P.; Kovalskaja, G.A.; Chankina, O.V. Biogeochemical role of the Italian locust Calliptamusitalicus L. (Orthoptera, Acrididae). In XII Congress of Russian Entomological Society (Saint Petersburg, Russia, 19–24 August, 2002). Abstracts; Russian Entomological Society: Saint Petersburg, Russia, 2002; pp. 299–300. (In Russian) [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Year | Aleksandrovskij | Burla | Yarovoe | Platovka | Severka | Pokrovka | Mikhaylovskoye | Bor-For- post | Ust-Volchikha |
|---|---|---|---|---|---|---|---|---|---|
| 1999 | ? | ? | 0.48 ± 0.27 | 0.80 ± 0.33 | 0.32 ± 0.22 | ~1 | 3.2 ± 0.61 | ? | 0.32 ± 0.22 |
| 2000 | 24.32 ± 2.34 | 9.60 ± 0.85 | 6.40 ± 0.65 | 4.48 ± 0.48 | 2.88 ± 0.67 | 5.28 ± 0.86 | 6.56 ± 0.80 | 0.96 ± 0.35 | 0.48 ± 0.27 |
| 2001 | 2.77 ± 0.70 | 5.60 ± 0.73 | 4.16 ± 0.63 | 3.04 ± 0.53 | 7.36 ± 0.82 | 4.00 ± 0.65 | 0.96 ± 0.35 | 0.08 ± 0.08 | 1.60 ± 0.46 |
| 2002 | 0.16 ± 0.16 | 3.20 ± 1.31 | 0.16 ± 0.16 | 1.28 ± 0.89 | 1.92 ± 1.06 | 6.40 ± 1.85 | 0.32 ± 0.32 | 1.92 ± 0.74 | 1.92 ± 1.06 |
| 2003 | + | 0.16 ± 0.16 | + | 0.16 ± 0.16 | 0.32 ± 0.23 | 0.13 ± 0.13 | 0.16 ± 0.16 | 0.32 ± 0.23 | 0 |
| 2004 | 0.32 ± 0.32 | + | + | 0.96 ± 0.54 | + | 0 | 0 | 0.40± 0.27 | 0.32± 0.32 |
| 2005 | 0.32 ± 0.23 | 0 | 0.38 ± 0.22 | 0.53 ± 0.37 | 0.05 ± 0.05 | 0 | 0 | 0.85 ± 0.29 | + |
| 2006 | + | 0.64 ± 0.45 | 0.21 ± 0.15 | 0.43 ± 0.30 | 1.28 ± 0.62 | 0.21 ± 0.21 | 0 | 0.75 ± 0.028 | 0.16 ± 0.16 |
| 2007 | 0.32 ± 0.23 | 0.96 ± 0.54 | 0.64 ± 0.32 | 1.16 ± 0.69 | 1.92 ± 0.74 | 1.07 ± 0.46 | 0 | 1.15 ± 0.37 | + |
| 2008 | 0.48 ± 0.27 | 0.16 ± 0.16 | 0.51 ± 0.25 | 0.96 ± 0.54 | 0.64 ± 0.45 | 0.32 ± 0.32 | 0 | 1.54 ± 0.46 | 0 |
| Year | Aleksandrovskij | Burla | Yarovoe | Platovka | Severka | Pokrovka | Mikhaylovskoye | Bor-Forpost | Ust-Volchikha |
|---|---|---|---|---|---|---|---|---|---|
| 1999–2000 | ? | ? | 41.5 0.0000 | 85 0.00001 | 202.5 0.0328 | 125 0.0003 | 167.5 0.000 | ? | n.s. |
| 2000–2001 | 469 0.0000 | 195.5 0.0024 | 196.5 0.0250 | n.s. | 126.5 0.0003 | n.s. | 166 0.0045 | n.s. | n.s. |
| 2001–2002 | n.s. | 165.5 0.0042 | 84.5 0.00001 | 153.5 0.0020 | 94 0.00002 | n.s. | n.s. | n.s. | n.s. |
| 2002–2003 | n.s. | n.s. | n.s. | n.s. | n.s. | 1012 0.0055 | n.s. | n.s. | n.s. |
| Year | Aleksandrovskij | Ust-Volchikha | ||||||
|---|---|---|---|---|---|---|---|---|
| Density | Species Behavior | E/F Females | E/F Males | Density | Species Behavior | E/F Females | E/F Males | |
| 1999 | ? | ? | ? | ? | 0.32 ± 0.22 | No bands and swarms | 1.576 ± 0.004 | 1.609 ± 0.006 |
| 2000 | 24.32 ± 2.34 | Numerous bands, small swarms, active local flights | 1.514 ± 0.016 | 1.539 ± 0.013 | 0.48 ± 0.27 | Very small and scarce swarms | 1.368 ± 0.010 | 1.392 ± 0.012 |
| 2001 | 2.77 ± 0.70 | No bands and swarms | 1.395 ± 0.013 | 1.406 ± 0.018 | 1.60 ± 0.46 | Very small and scarce swarms | 1.473 ± 0.011 | 1.479 ± 0.009 |
| 2008 | 0.48 ± 0.27 | No bands and swarms | 1.484 ± 0.038 | 1.384 ± 0.024 | 0 | No bands and swarms | ? | ? |
| Plot | 7 June | 12 June | 13 June | 15 June | 16 June | 23 June | 26 June | 5 July | 15 July | |
|---|---|---|---|---|---|---|---|---|---|---|
| Type | Treated/Untreated | D1 | D6 | D7 | D9 | D10 | D17 | D20 | D29 | D39 |
| Blanket | 31.4 | 70.0 | 100 | 93.7 | 100 | 100 | 76.0 | 100 | 86.1 | |
| 1:1 | Treated | 57.2 | 81.3 | 100 | 100 | 93.2 | 100 | 100 | 91.7 | 100 |
| Untreated | 35.7 | 43.8 | 77.5 | 80.3 | 79.5 | 70.8 | 55.0 | 50.0 | 65.4 | |
| 1:2 | Treated | 46.4 | 81.3 | 100 | 96.1 | 100 | 100 | 100 | 100 | 82.7 |
| Untreated | 57.2 | 62.5 | 70.0 | 56.6 | 86.4 | 66.7 | - | 8.3 | 56.7 | |
| 1:3 | Treated | 14.3 | 70.0 | 76.0 | 81.1 | 90.1 | 86.8 | 100 | 33.3 | 86.1 |
| Untreated | 14.3 | 10.0 | 52.0 | 36.9 | 78.2 | 33.9 | 4.0 | - | 58.4 | |
| Year | Aleksandrovskij | Burla | Yarovoe | Platovka | Severka | Pokrovka | Mikhaylovskoye | Bor-Forpost | Ust-Volchikha |
|---|---|---|---|---|---|---|---|---|---|
| 1992 | ? | ? | AB, NA | ? | ? | ||||
| 1999 | ? | ? | MP | ? | |||||
| 2000 | GG, MP b | AB, NA | |||||||
| 2001 | AB, GG c, MP | GG | NA | GG, SP a | |||||
| 2002 | AB, MP d | AB | |||||||
| 2003 | |||||||||
| 2004 | GG, MP e | ||||||||
| 2005 | GG | GG | AB | GG, SP | |||||
| 2006 | GG | AB, MP | MP a | ||||||
| 2007 | MP | GG a | |||||||
| 2008 | GG e, MP e | GG | GG | GG | |||||
| 2015 | GG | MP | MP |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sergeev, M.G. Ups and Downs of the Italian Locust (Calliptamus italicus L.) Populations in the Siberian Steppes: On the Horns of Dilemmas. Agronomy 2021, 11, 746. https://doi.org/10.3390/agronomy11040746
Sergeev MG. Ups and Downs of the Italian Locust (Calliptamus italicus L.) Populations in the Siberian Steppes: On the Horns of Dilemmas. Agronomy. 2021; 11(4):746. https://doi.org/10.3390/agronomy11040746
Chicago/Turabian StyleSergeev, Michael G. 2021. "Ups and Downs of the Italian Locust (Calliptamus italicus L.) Populations in the Siberian Steppes: On the Horns of Dilemmas" Agronomy 11, no. 4: 746. https://doi.org/10.3390/agronomy11040746
APA StyleSergeev, M. G. (2021). Ups and Downs of the Italian Locust (Calliptamus italicus L.) Populations in the Siberian Steppes: On the Horns of Dilemmas. Agronomy, 11(4), 746. https://doi.org/10.3390/agronomy11040746