1. Introduction
Weeds are a major issue in agriculture, causing 34% potential yield losses worldwide [
1]. In vegetable crops, yield decrease may be between 45 and 95% due to weed competition [
2]. In Spain, more than €370 million was spent on herbicides in 2019 out of a total sum of €1100 million spent on pesticides [
3], which makes it evident that weed control is a costly issue. Hydromulching is a technique that has been used for several decades in some countries such as the USA or Canada, mainly for erosion control, and is applied for this purpose on slopes next to highways or similar facilities [
4]. It is based on spraying a liquid blend, which solidifies on the ground, generally containing stabilized organic residues mixed with binders and water. However, due to its capacity to reduce evaporation and weed emergence, the use of this technique has been extended to greenhouses, nurseries, parks, and railway facilities [
4,
5,
6], and several different formulations have been tested.
Weed control by hydromulch is especially suitable for minor crops where herbicide use is complicated because few active ingredients are available [
7]. Other target situations are the first development phases of vegetable crops [
8] and in the first years of orchard planting, when ploughing may harm the plants. Other mulches such as polyethylene (PE), biodegradable plastic or paper mulches might be good solutions in annual vegetable crops [
9,
10,
11,
12,
13], but installation is complicated or impossible in already established perennial crops. Moreover, in perennial crops, mulching materials should assure some degree of self-incorporation, which is not guaranteed with mulch sheets that easily tear and can be wind-dispersed, polluting the environment [
14].
In the current context of reducing negative environmental impacts, weed control by hydromulch could be a useful and innovative technology [
15]. These mulches are aimed to be applied mechanically on the soil surface next to the plant stems or trunks and an application prototype is currently in its testing period. Preliminary field trials have shown that a mulch layer of around 2 cm thick can impede the emergence of annual weeds [
16].
However, there are few detailed studies published about the technique, and very little is known regarding perennial weed control. The exception is the study by [
6], who found that their hydromulch could control annual plants in a field trial, but the shoots of the perennial
Cyperus rotundus L. (purple nutsedge) pierced the layer. This species is widespread in temperate and tropical regions of the world, affecting vegetable crops, sugarcane, cotton, and other crops, and has been reported to cause 20–90% yield losses worldwide [
17].
Cynodon dactylon L. (Pers.) and
Sorghum halepense L. (Pers.) are also included in the list of the 10 worst weeds [
18] and all three, together with
Paspalum dilatatum Poiret, are common species in irrigated crops in north-eastern Spain. All of them are capable of clonal growth through rhizome production and fragmentation, particularly tubers in purple nutsedge. Populations of
C. rotundus mainly affect vegetable crops [
17], those of
C. dactylon are very persistent in fruit orchards and vineyards [
19],
S. halepense is a troublesome weed species in orchards and in maize fields [
20], and
P. dilatatum is very persistent in lawns and in other perennial crops [
21].
Preliminary characterization of hydromulches composed of recycled paper pulp, lignocellulosic crop residues, and gypsum as a binder has shown promising effects on annual weed emergence reduction in two ways: first, the percentage of seeds that were able to germinate was reduced because they died or because they went into secondary dormancy; and second, a large proportion of seedlings were unable to pass through the mulch layer and died [
22]. These formulations have been improved by adding kraft fiber to give additional strength to the mixture [
23] so that perennial weeds might be able to be retained under the mulch. However, their effects on preventing perennial weed sprouting have not been tested yet. The objective of the present work was thus to learn the potential weed control capacity of these hydromulch mixtures on four common perennial weed species in orchards and vegetables in the region with different rhizome sizes and shapes in greenhouse conditions. The results should contribute to the decision on whether or not to install subsequent field trials with these hydromulch blends on perennial weeds.
The specific aim of this work was to find if the hydromulches were able to reduce weed emergence of some of the four different perennial weed species.
2. Materials and Methods
The greenhouse trials were conducted in two locations, namely the Agròpolis facilities belonging to the Universitat Politècnica de Catalunya (UPC) in Viladecans (V) (Barcelona, Spain) and the CITA facilities in Montañana (M) (Zaragoza, Spain), for two years, 2019 and 2020 (
Table 1), so that data from four similarly designed experiments were available. Both greenhouses had glass walls and a roof, but with some differences. The one in V had overhead ventilation that opened automatically when the air temperature exceeded 25 °C. The one in M was painted with lime milk prior to the experiments, had lateral forced ventilation with a water-circulation cooling system, and the maximum temperature was set at 30 °C.
Rhizomes from the four species were collected from natural populations, washed, and kept in vermiculite at 4 °C until use. Rhizomes from
P. dilatatum and
S. halepense were collected once each year at V and the other two species in M, in all cases a few days prior to the installation of the experiment. The
C. dactylon rhizomes were cut into fragments with two nodes and some roots because roots enhance the probability of emergence [
16]. The
P. dilatatum and
S. halepense rhizomes were also cut up in such a way that the fragments employed had a minimum length of 1 cm and at least two nodes. Out of the
C. rotundus tubers collected, the biggest and the smallest were rejected and only those of medium size were used. The mean values of rhizome length and their standard errors were 27.4 ± 0.86 mm (
C. dactylon), 15.7 ± 0.16 mm (
C. rotundus), 16.6 ± 0.35 mm (
P. dilatatum), and 32.4 ± 0.78 mm (
S. halepense). Fragments and tubers without any visible sprouts were chosen.
The maximum diameter and the length of each rhizome and tuber were measured immediately before planting with a digital calliper (Mahr 16E, Esslingen, Germany, ±0.01 mm). Each rhizome was placed individually in a plastic container, hereafter called rhizome box, measuring 8 × 8 × 8 cm. Ten of these boxes were placed in a circular pot 28 cm in height and 39 cm in diameter. In previous trials, it had been observed that C. rotundus and C. dactylon plants were able to grow horizontally under the hydromulch layer for several cm until reaching the pot edge and emerging there without needing to pierce the hydromulch (data not shown). This design was thus chosen to hinder the rhizomes and tubers from sprouting without piercing the mulch, forcing them to grow vertically inside their 8 × 8 × 8 cm box. Moreover, the isolation of each rhizome/tuber in an individual rhizome box should also facilitate the detection of which rhizome/tuber was piercing and the subsequent detection of the non-sprouted rhizomes/tubers.
The first 10 cm of the pots were filled with lightweight expanded clay aggregate (8/16 mm Burés, Castelldefels, Barcelona), which was then covered with 8 cm of substrate (J2 Ecológico Burés, Castelldefels, Barcelona; a mixture of vegetable compost, Sphagnum peat and perlite; NPK 8-3-3). The ten boxes were placed on this substrate and the gaps between the boxes were filled with more substrate. Once the rhizomes were planted at a depth of 3–4 cm, they were covered with substrate and each pot received 1 L of water.
Four treatments were considered: three hydromulch compositions with promising physical characteristics [
16,
23] and a non-mulched control. All three formulations contained 16.7 L m
−2 recycled paper pulp, 1002 g m
−2 gypsum (Marfil ALGISS), and 209.3 g m
−2 kraft fiber (Capellades Paper Mill Museum, Capellades, Spain). The wheat straw hydromulch (WS) contained 833 g m
−2 of wheat straw milled with a 2.5 mm sieve, the rice husk hydromulch (RH) contained 1250 g m
−2 of rice husk, and the hydromulch based on used substrate for mushroom cultivation (UMS) contained 3100 g m
−2 of open-air dried mushroom substrate. The hydromulches were applied by pouring 2.3 L of the mixture on each pot, and the thickness of each newly applied hydromulch layer was 1.9 cm. Immediately after an application, around 20–25% of the hydromulch weight was lost by means of liquid that drained down by gravity, the amount depending on the proportion of water and fibers of the paper pulp. After the mentioned rapid initial water loss, the hydromulch slowly lost the rest of its water content over several days until it became a dry solid mulch. The pots that were not covered with hydromulch, serving as controls, received an additional 1 cm of substrate, and an additional 1.5 L of water, to compensate for the additional water received by the mulched pots.
In total, 64 pots containing 160 rhizomes of each species were used in each trial, corresponding to four replicates each year at each locality.
Pots were drip irrigated in the Viladecans trials with two emitters each feeding 0.03 L min−1 once a day for 6 min, each pot receiving 360 mL daily. In the Montañana trials, pots were watered by immersion in a basin when considered necessary after observing that the soil was dry under the mulch layer.
When no new emergences were observed for at least one week, the trials were finished. Data were collected for each individual rhizome box. The hydromulch layer was lifted carefully and the rhizomes were extracted to determine if they had sprouted or not, if any shoot had developed or not, and if the developed shoots were either trapped by the hydromulch layer or had emerged through the mulch. There were four possible types of rhizomes: (i) rhizomes without any inflated buds; (ii) rhizomes with inflated buds that did not enlarge; (iii) rhizomes with developed underground shoots that were trapped by the hydromulch; and (iv) rhizomes with developed shoots that emerged above ground by piercing the mulch layer.
The following variables, all of them expressed per pot, were computed, aggregating the data obtained in each of the 10 rhizome boxes: proportion of sprouted rhizomes (SPROUTED), proportion of rhizomes that produced emerged shoots (EMERGED), proportion of rhizomes that produced shoots that were not able to emerge but grew lengthwise, trapped under the hydromulch (TRAPPED), proportion of sprouted rhizomes whose shoot/s emerged (EMERGED/SPROUTED), and proportion of sprouted rhizomes whose shoots were all trapped under the hydromulch (TRAPPED/SPROUTED).
The variables were analyzed by means of generalized linear mixed models with a binomial distribution and logit link function. Parameters were estimated by maximum likelihood with Laplace approximation.
Three fixed effects were considered: (i) ‘hydromulch’, with four levels (non-mulched control, WS, RH and UMS), except for TRAPPED and TRAPPED/SPROUTED, which were only applicable to the three types of hydromulch; (ii) ‘species’, with four levels (C. dactylon, C. rotundus, P. dilatatum and S. halepense); and (iii) the interaction between them. Type III chi-square significance tests were performed for these three fixed effects. The random sources taken into account varied depending on the analysis, in each case seeking the best fitted model, and all of them were modeled with unstructured covariance matrices. In the first step of the analyses, the following sources were considered as possible intercept terms of the linear predictor: ‘locality’, ‘year’, ‘locality × year’, ‘block’ nested within ‘locality × year’, ‘pot’, and the covariables ‘mean rhizome length’ and ‘mean rhizome width’ per pot nested within ‘species’. After that, in a second step, some of the random sources were removed from the model if the fit statistics Akaike’s information criterion (AIC) and Pearson chi-square/degrees of freedom showed that the model fitted better without them. A third step took place when the interaction ‘hydromulch × species’ was not significant at α = 0.05: the interaction was removed as a fixed effect and the variable was reanalyzed.
The least-squares means of the levels of the main fixed effects and of the significant (
p < 0.05) interaction effect and its 95% confidence limits were computed using probability values from the chi-square distribution. Multiple comparisons were made following the Tukey–Kramer procedure. The SAS/GLIMMIX procedure [
24] was used to perform the above-mentioned generalized mixed linear models and least-squares means comparisons.
The variable EMERGED was analyzed a second time with the aim of describing the role of rhizome size in its behavior. The analysis was conducted species by species, using a generalized linear model with a binomial distribution considering the fixed effect ‘hydromulch’ with four levels, and considering the covariable ‘mean rhizome length’ one time and ‘mean rhizome width’ another time. The analyses were performed using the SAS/GENMOD procedure [
24], parameters were estimated using the logit link function and Type III analysis options; the dispersion parameter was estimated as the deviance divided by its degrees of freedom because of overdispersion, and all statistics were adjusted appropriately. Likelihood ratio statistics were used to compute the significance of the fixed effect and the sources of covariation.
3. Results and Discussion
Overall, rhizomes sprouted satisfactorily in the non-mulched control with a mean of 72%.
P. dilatatum was the species with the lowest sprouting (62%) and
C. rotundus the highest (88%). Symptoms of diseases were not observed in the non-sprouted rhizomes in any case. Base temperature for rhizome sprouting of
C. dactylon was determined at 7.7 °C [
25] and 8–10 °C for
S. halepense [
26]. An increase in sprouting was found for
C. dactylon within the range 11–33 °C [
25], 10–32 °C for
S. halepense [
27], and 10–45 °C for
C. rotundus [
28], which is coincident with the conditions in the experiments (
Table 1). With respect to moisture availability, all four species are reported to grow optimally in conditions similar to field capacity. However,
C. rotundus,
S. halepense, and
C. dactylon have a certain drought tolerance [
17,
29,
30], while flooding could be detrimental to
P. dilatatum [
31].
The results reported in
Table 2 and
Table 3 show clearly that with the tested hydromulches, perennial weed emergence could be reduced by means of three mechanisms: (i) inhibiting rhizome sprouting; (ii) preventing sprout elongation; and (iii) trapping the elongated sprouts by avoiding piercing.
RH and UMS hydromulches were effective in reducing rhizome sprouting compared to the non-mulched treatment, while WS showed non-significant differences with respect to the non-mulched treatment (
Table 2 and
Table 3). The hydromulches’ degree of efficiency varied according to the species, but no significant differences were found in the least-squares means of the sprouting among the three types of hydromulch tested. All three mulches inhibited sprouting similarly, probably because of the modified environmental conditions in the soil compared to the non-mulched treatment, for example, in light, temperature, and air-soil gas exchange. Concerning temperature, several authors have demonstrated a reduction of the maximum and an increase in the minimum temperatures in the soil when using several hydromulch formulations [
6,
32].
P. dilatatum showed the highest proportion of trapped sprouts out of the sprouted samples and
C. rotundus the lowest, this proportion being intermediate for
S. halepense and
C. dactylon (
Table 3). With regard to emergence,
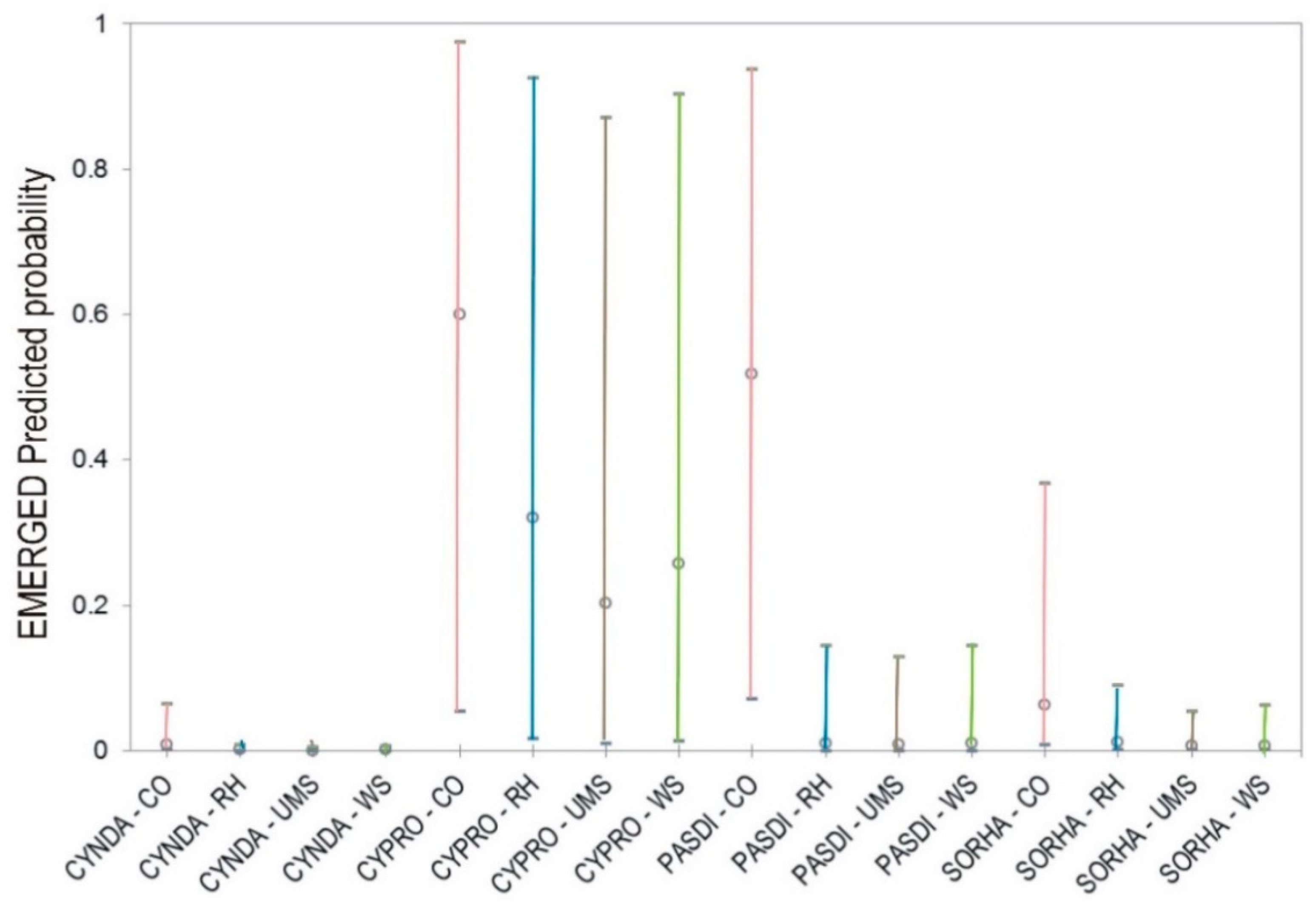
Figure 1 shows that for
P. dilatatum mean emergence was significantly higher for the non-mulched control and similar for all three mulches, while at the other extreme,
C. rotundus emerged similarly in all four treatments. These results are in accordance with our preliminary observations in field conditions (data not shown). Emergence reduction was especially high for
P. dilatatum compared to the emergence recorded in the untreated control. However, all three hydromulch compositions showed the same mean ability to suppress the emergence of the weeds, except for
C. rotundus (
Table 3,
Figure 1), the emergence of which was also not impeded by a cotton-based mulch [
6]. Although the sprouts of all four tested species specialized in penetrating the soil, the sharp leaves of
C. rotundus were shown to be the most capable of piercing the mulches. These preliminary results warrant testing the mulches in field conditions on sites with infestations of
C. dactylon,
P. dilatatum, and
S. halepense, as they performed better than for
C. rotundus. However, all mulches yielded a lower proportion of emerged shoots out of the sprouted rhizomes than the non-mulched control.
The hydromulch layers trapped, on average, the shoots of 16.3% of the sprouted
C. rotundus tubers placed in rhizome boxes (
Table 3). At the other extreme, the best control was exercised on
P. dilatatum; their rhizomes sprouted under the hydromulches, but the shoots were efficiently trapped, 87.5% on average (
Table 3). Rhizomes of
C. dactylon and
S. halepense, despite their differences in rhizome size (
Figure 2), displayed a similar and intermediate behavior: about half of the sprouted rhizomes did not pierce the hydromulch layer with any shoot (
Table 3). With the exception of
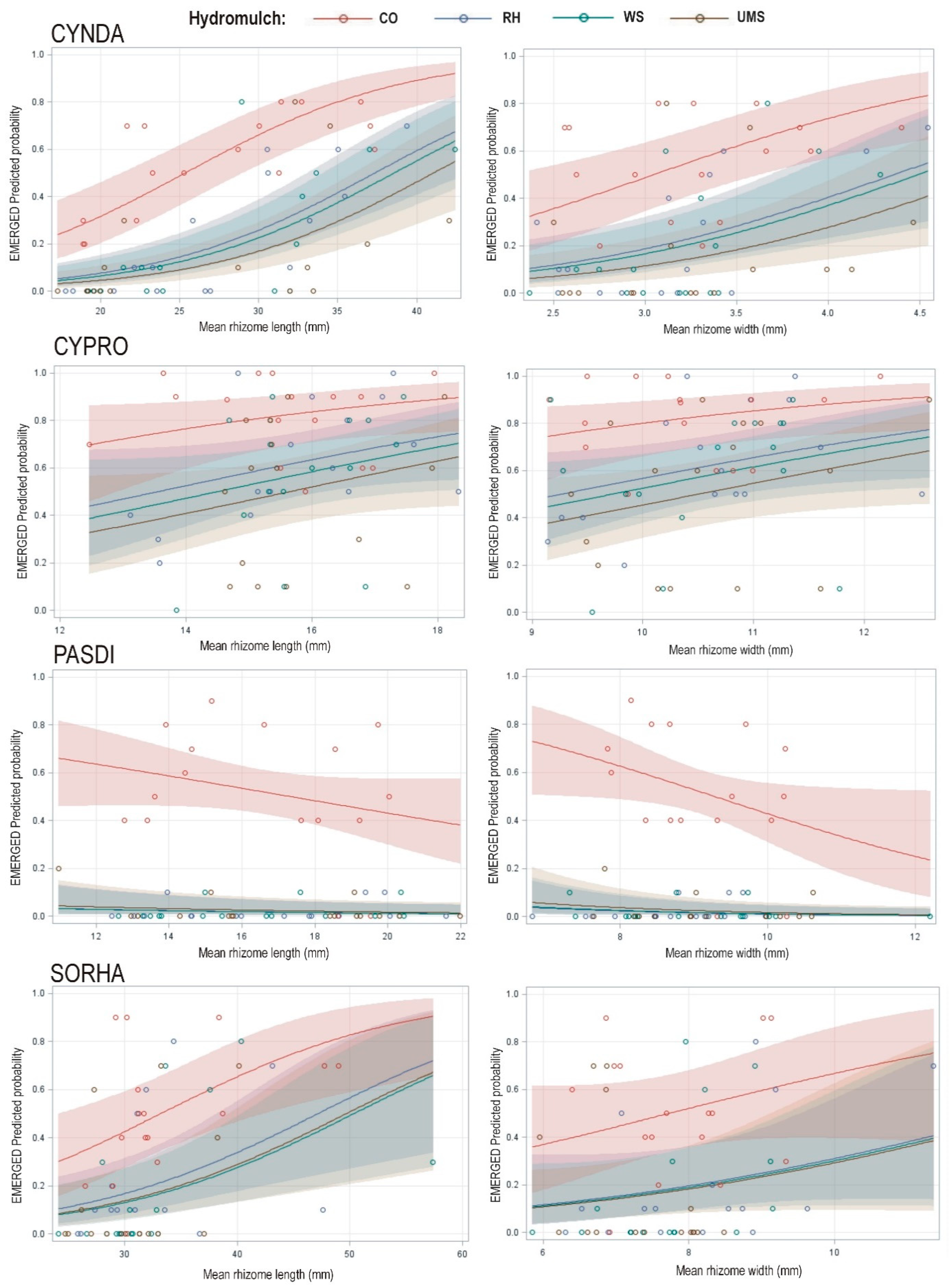
P dilatatum, the predicted emergence within species increased when the rhizome size increased (
Figure 2).
In general, the random effects gave the fitted models little variability, with the exception of ‘pot’ (
Table 2,
Figure 2). The relative prominence of the random source of variation ‘pot’ may be explained by the possibility that, despite the experimental design, aimed at achieving independence between the rhizomes of the ten boxes inside each pot, the rhizomes in the boxes might actually have affected each other. Thus, if one sprout pierced and emerged through the mulch layer, then the environment of the rhizomes located in neighboring boxes was probably changed by the lifting of the mulch, allowing some light to come through.
The ability to sprout and to produce shoots that can emerge was mediated, among other factors, by the mean rhizome length (
Table 2). In all eight generalized linear models fitted, considering for each species the fixed effect ‘hydromulch’ and one of the two covariables related to rhizome size, the hydromulch was significant at
p < 0.05. As expected, considering the least-squares mean values of this variable for the four levels of hydromulch (
Table 3), the probability of emergence was always highest in the non-mulched treatment, while there was little difference among the three types of hydromulch (
Figure 2).
Figure 2 shows that the effect of the mean rhizome length on the probability of emergence varied with the species. The results obtained for
P. dilatatum were especially forceful; the probability of emergence was independent of the mean rhizome length within the utilized range (
Figure 2). Greater mean rhizome width also implied higher probability of emergence for three species; curiously, in
P. dilatatum, increasing rhizome width reduced plant emergence in the non-mulched control treatment (
Figure 2).
C. dactylon had the thinnest rhizomes (3.2 ± 0.07 mm), followed by
S. halepense (7.8 ± 0.13 mm).
P. dilatatum, and
C. rotundus had the thickest rhizomes (9.0 ± 0.12 mm and 10.5 ± 0.11 mm respectively).






{kind=link}
{kind=link}