Evidence for a Causal Relationship between the Solar Cycle and Locust Abundance

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
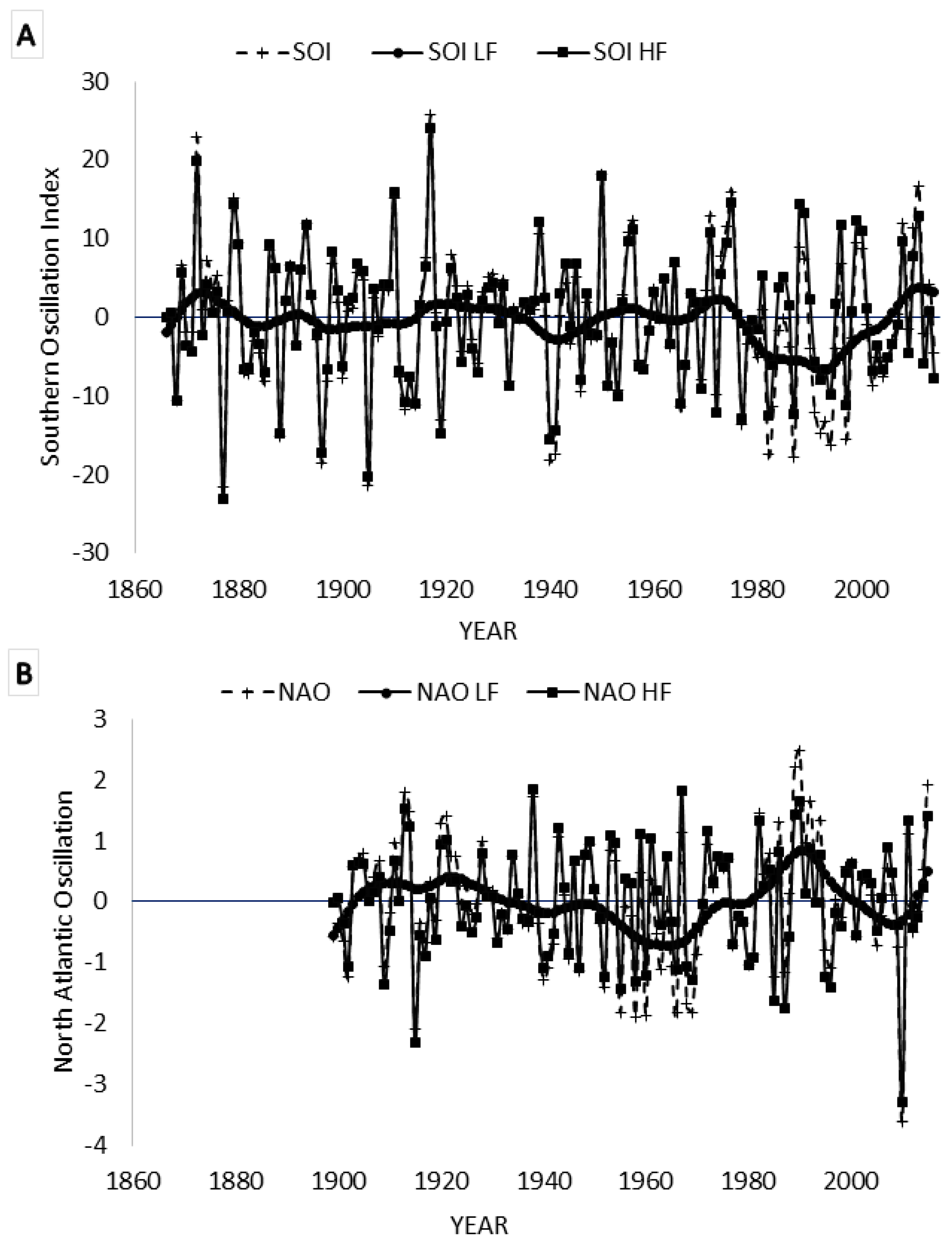
2.1. Data Sets
2.2. Statistical Analyses
3. Results
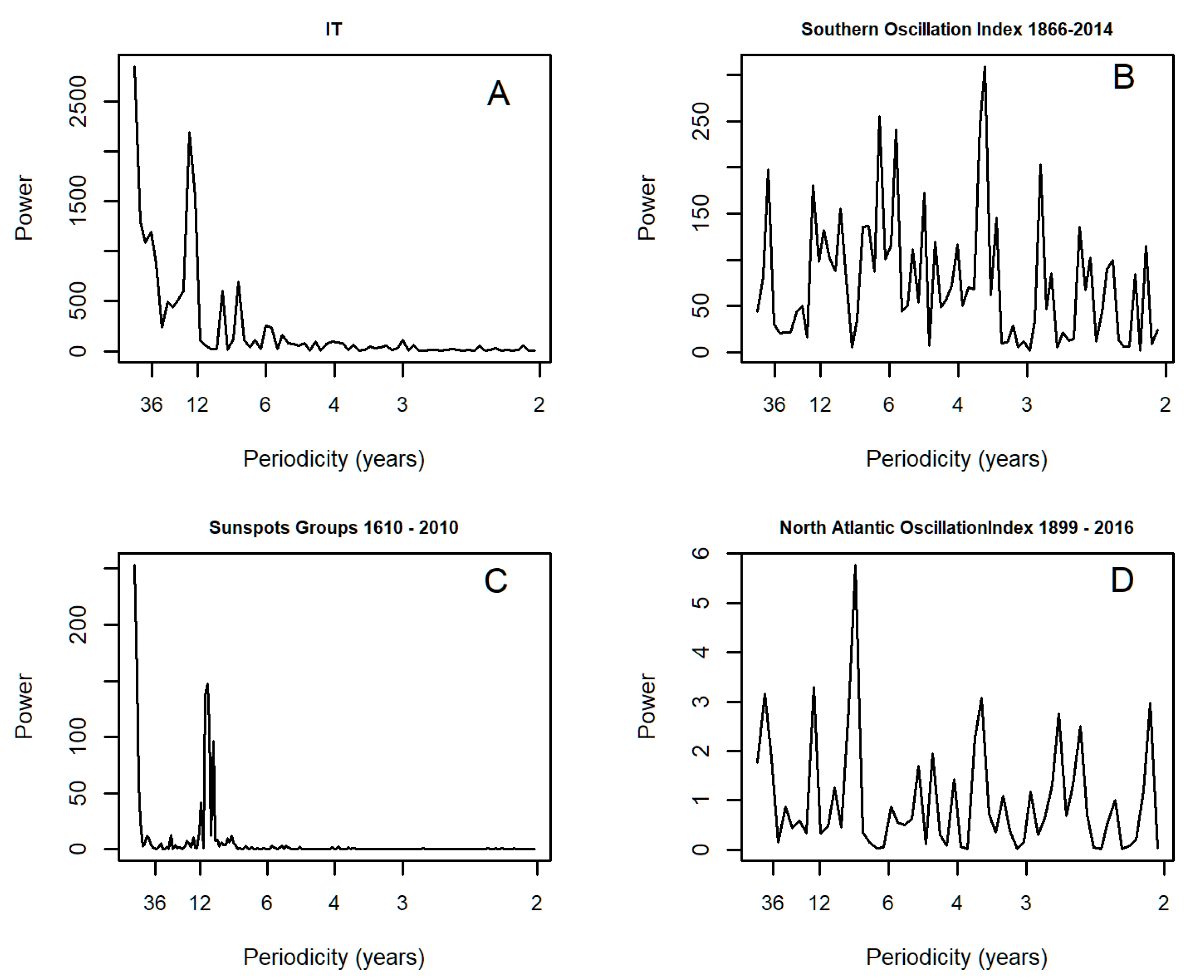
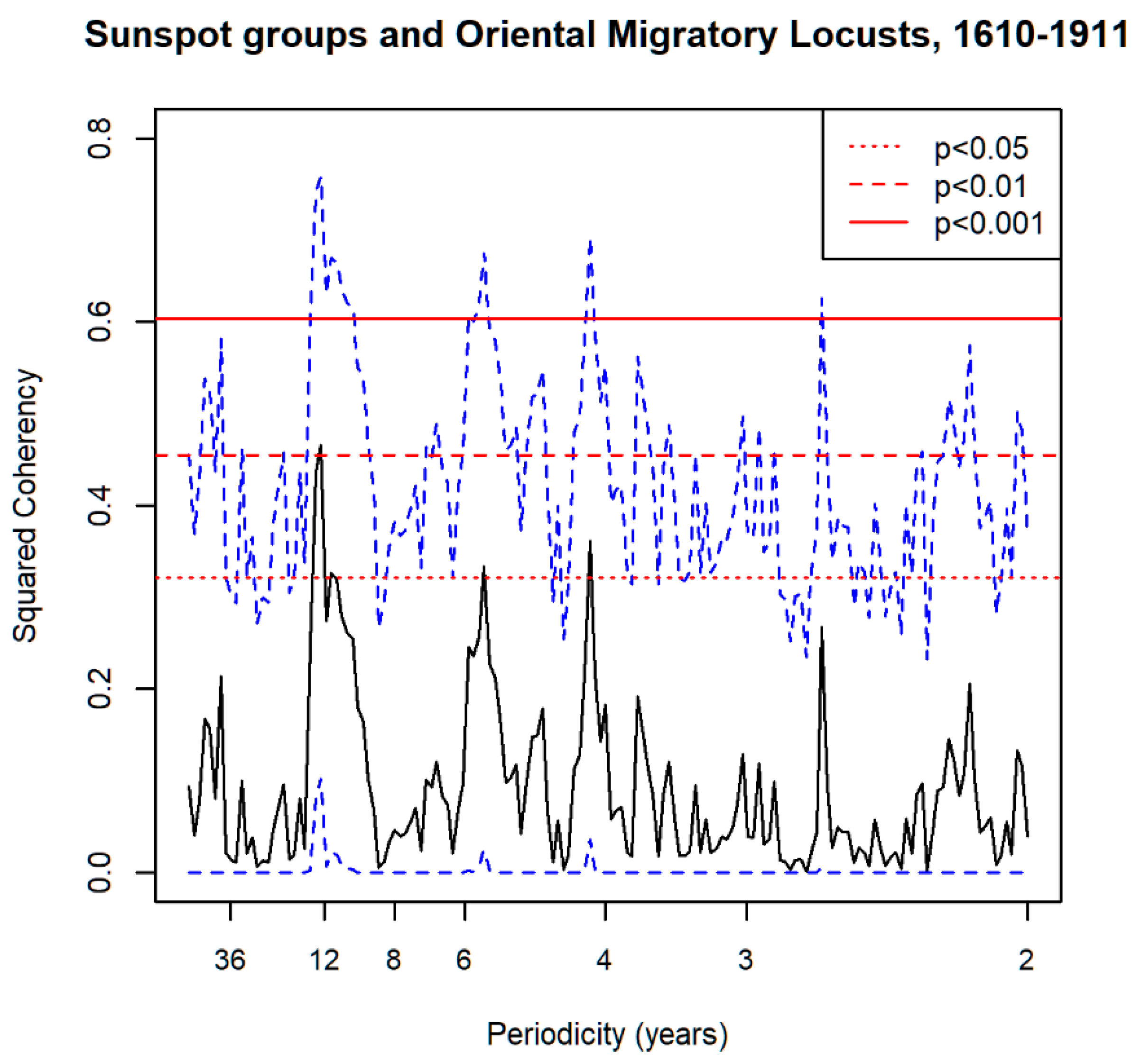
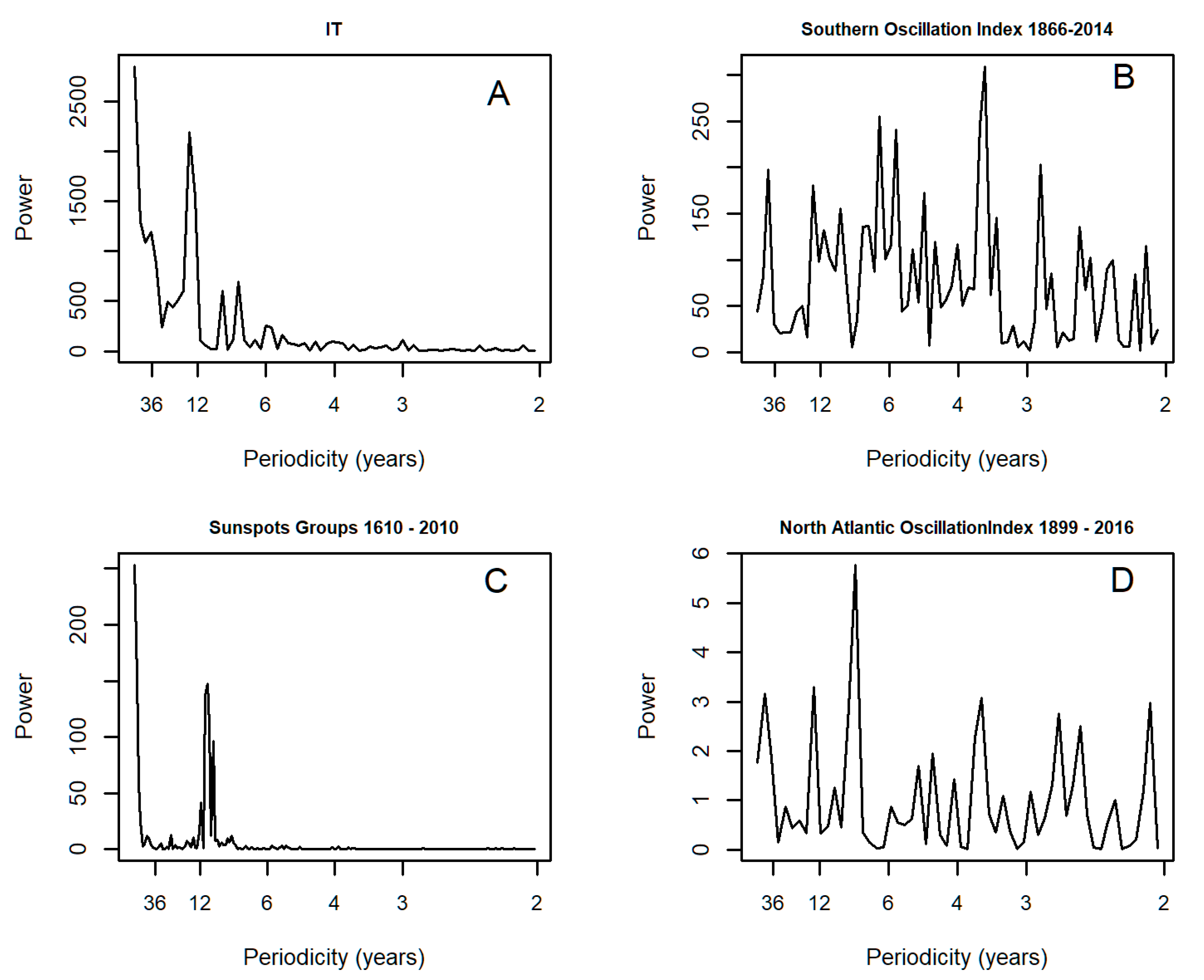
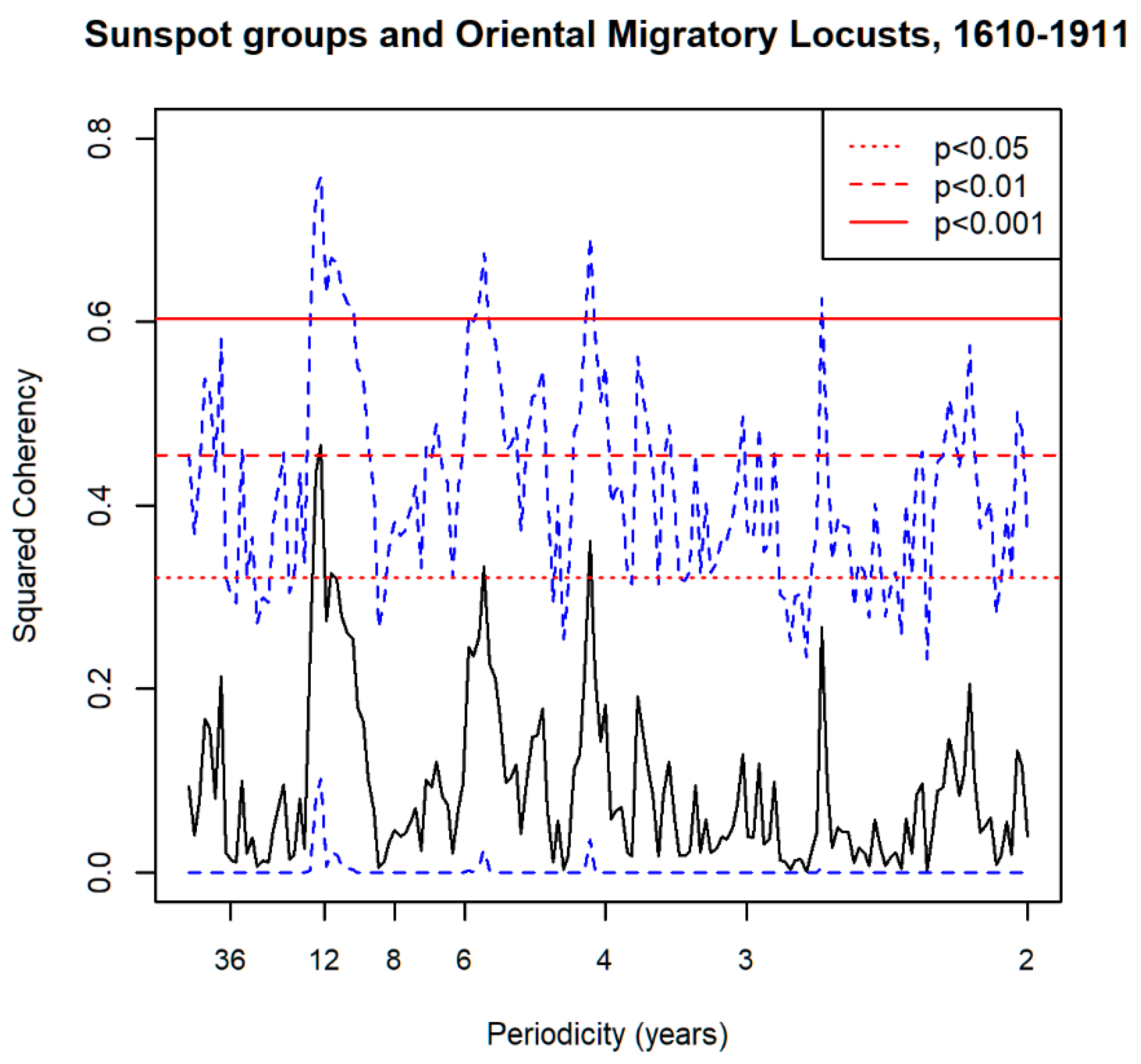
3.1. Spectral Analysis: Oriental Migratory Locusts
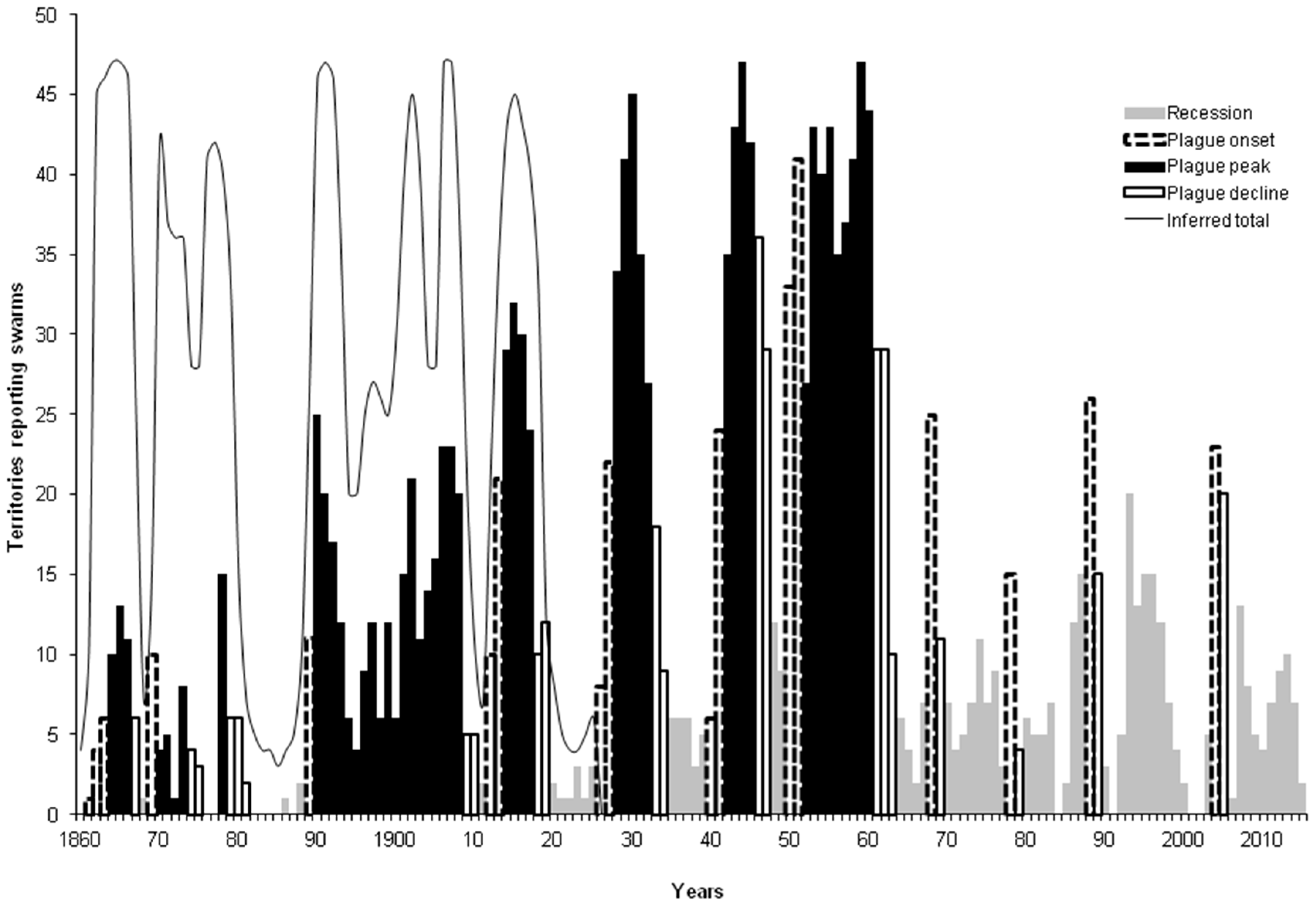
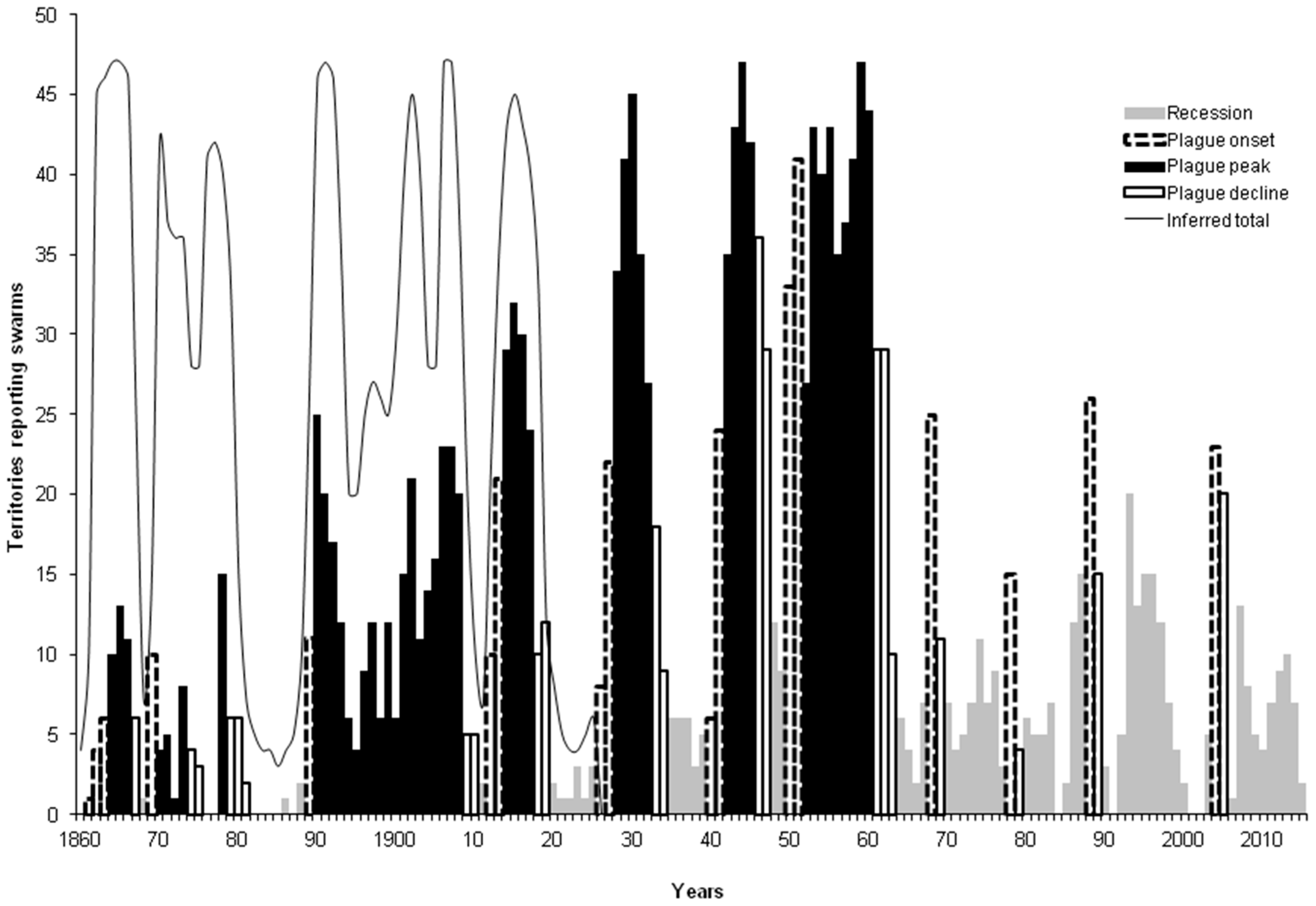
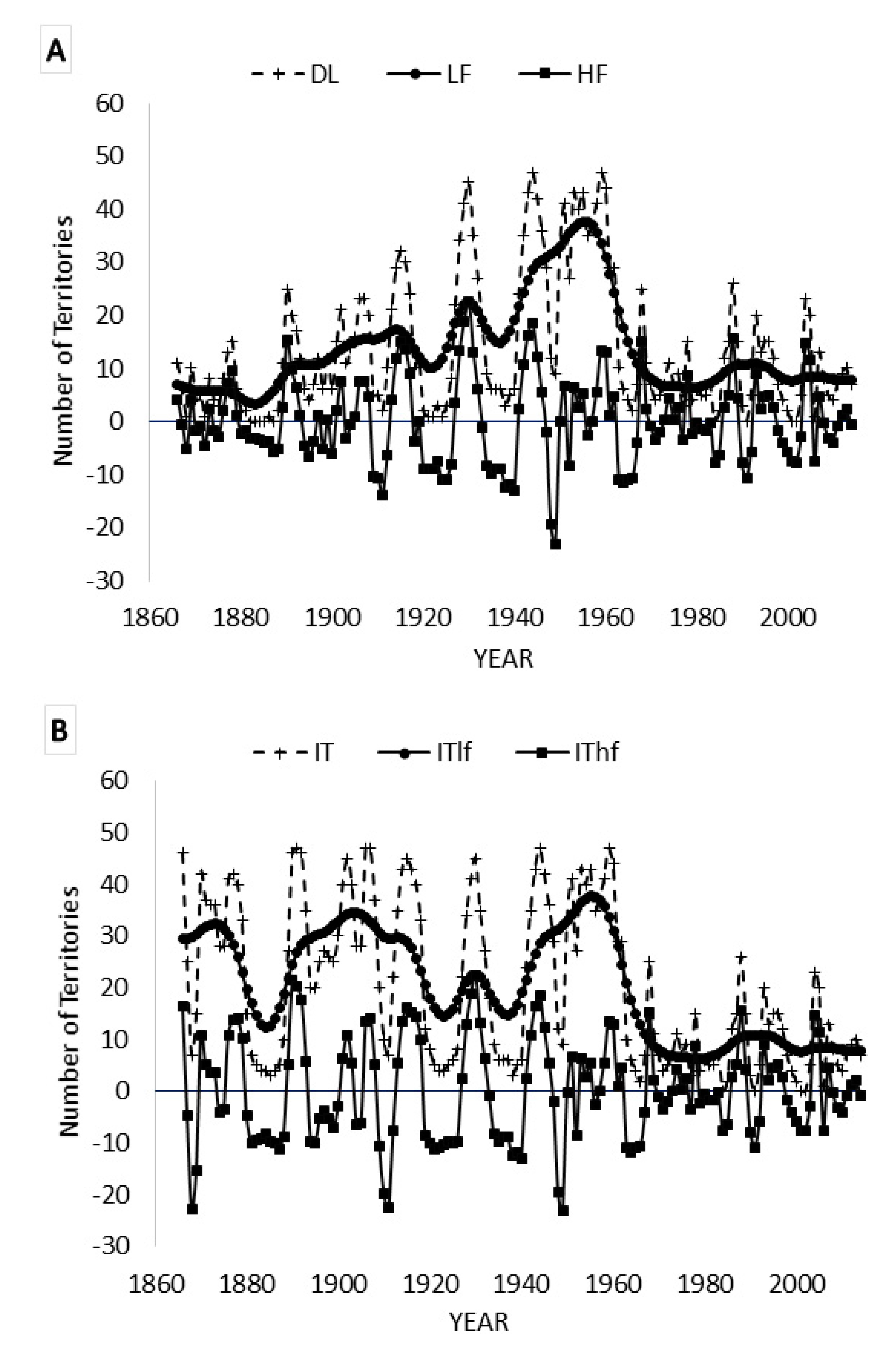
3.2. Spectral Analysis: Numbers of Territories Infested with Desert Locust Swarms, 1866–2010
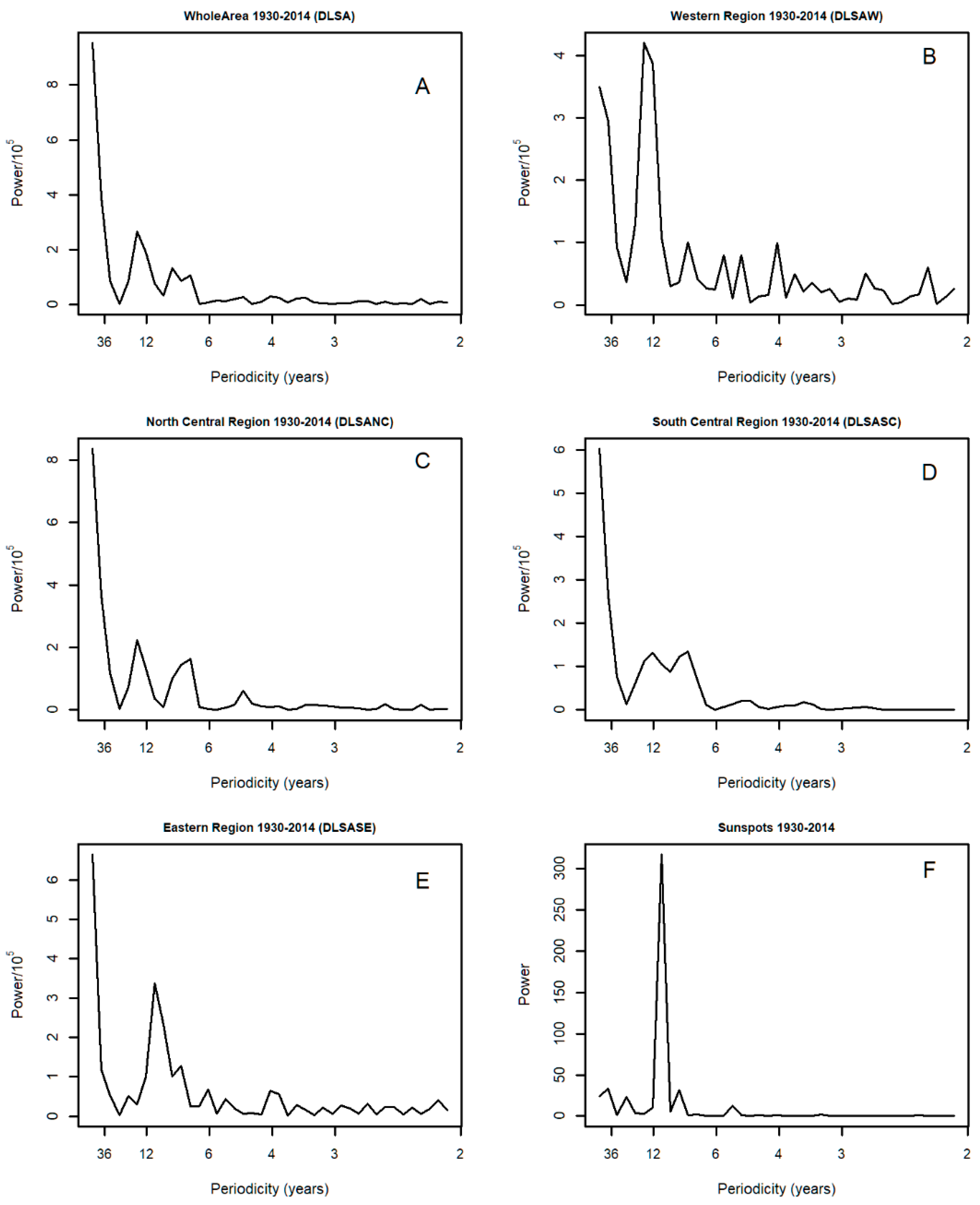
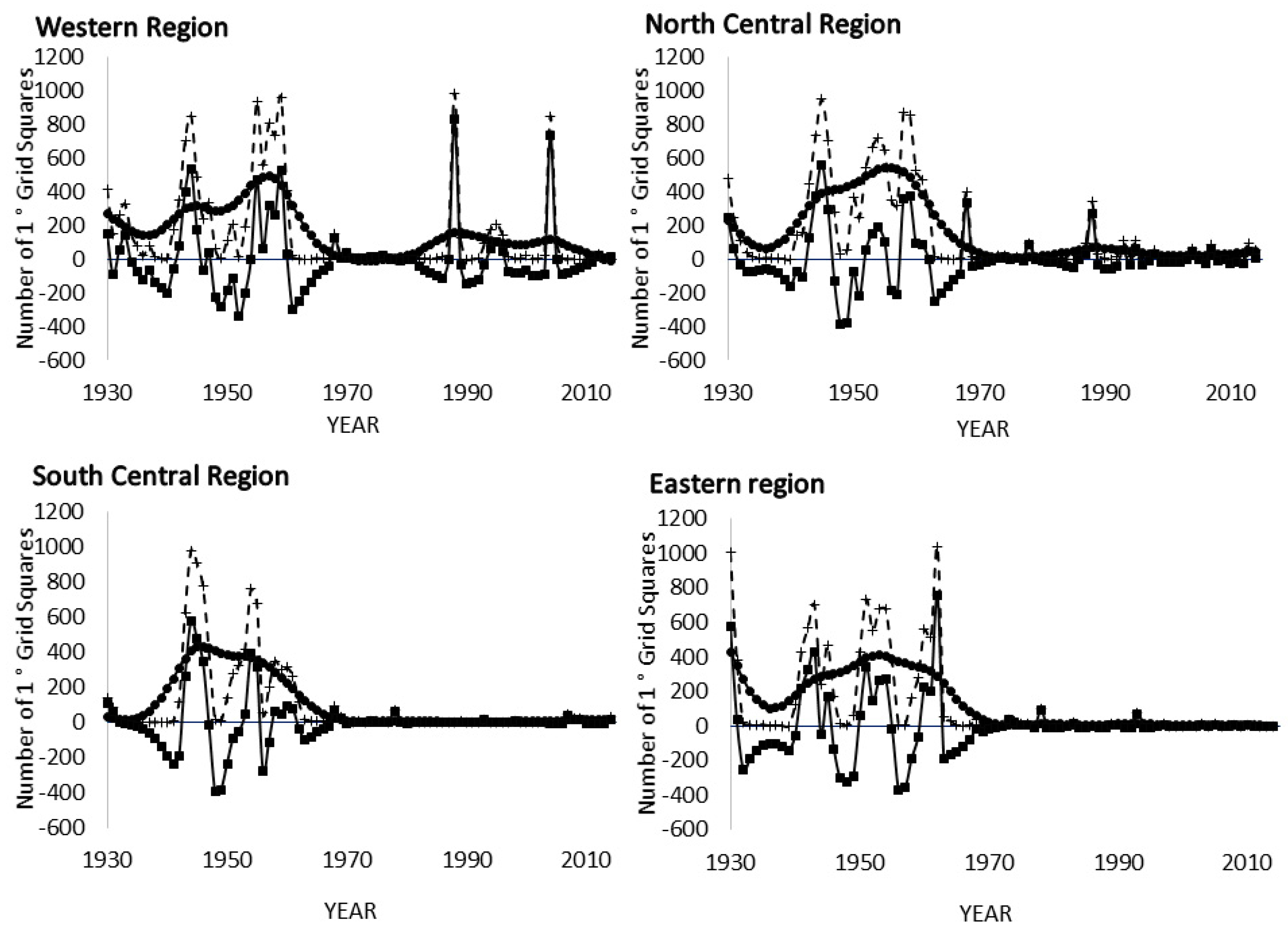
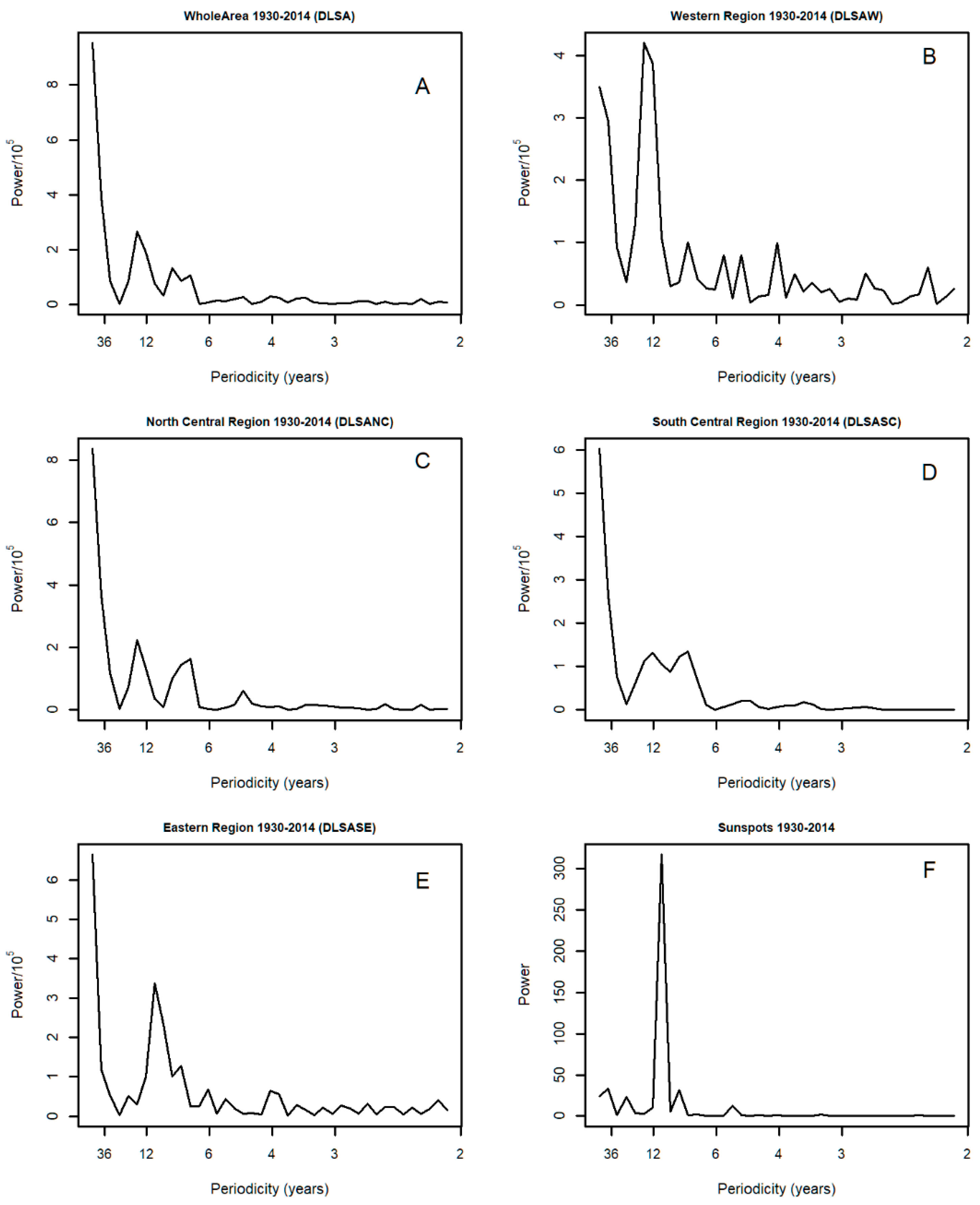
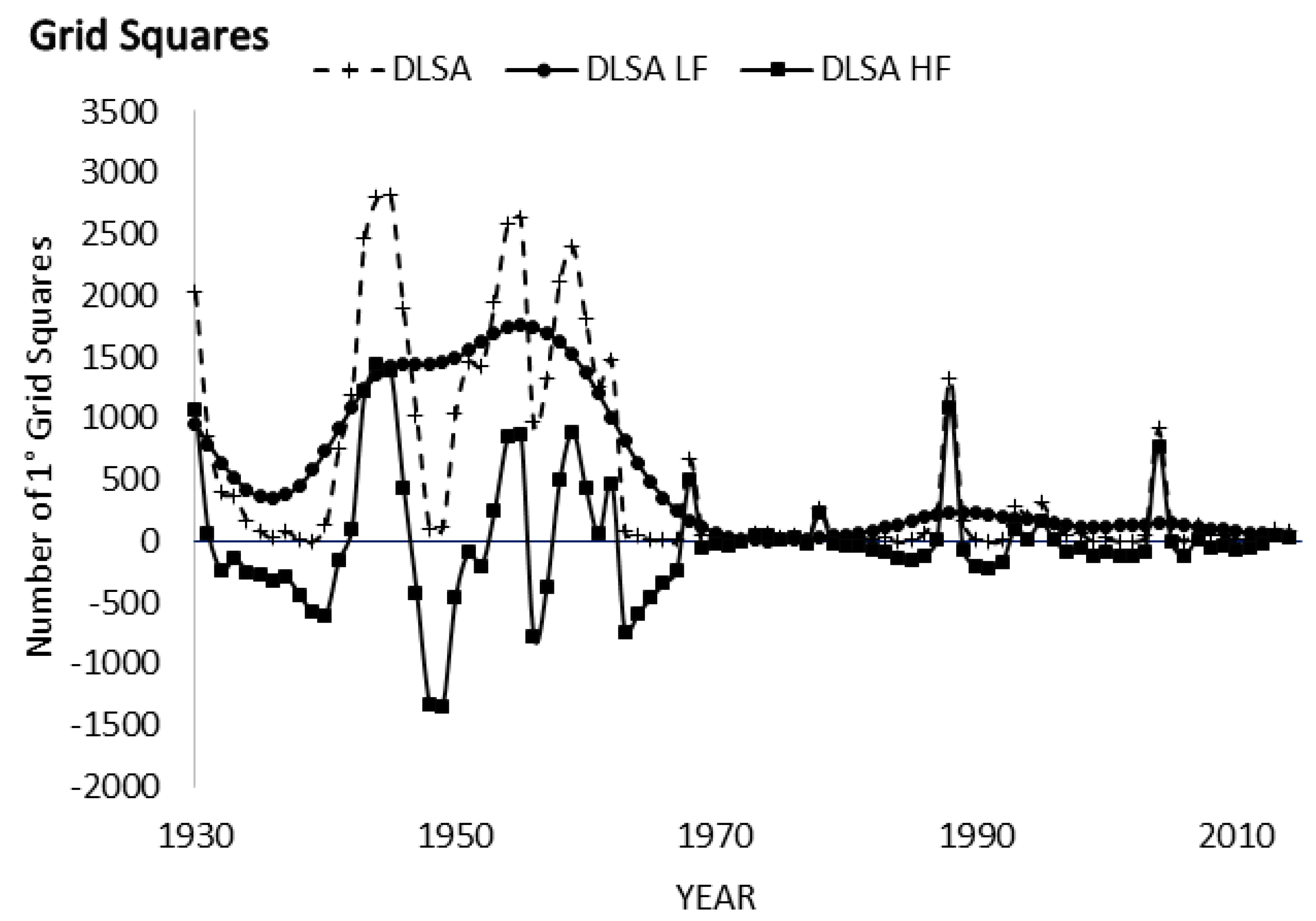
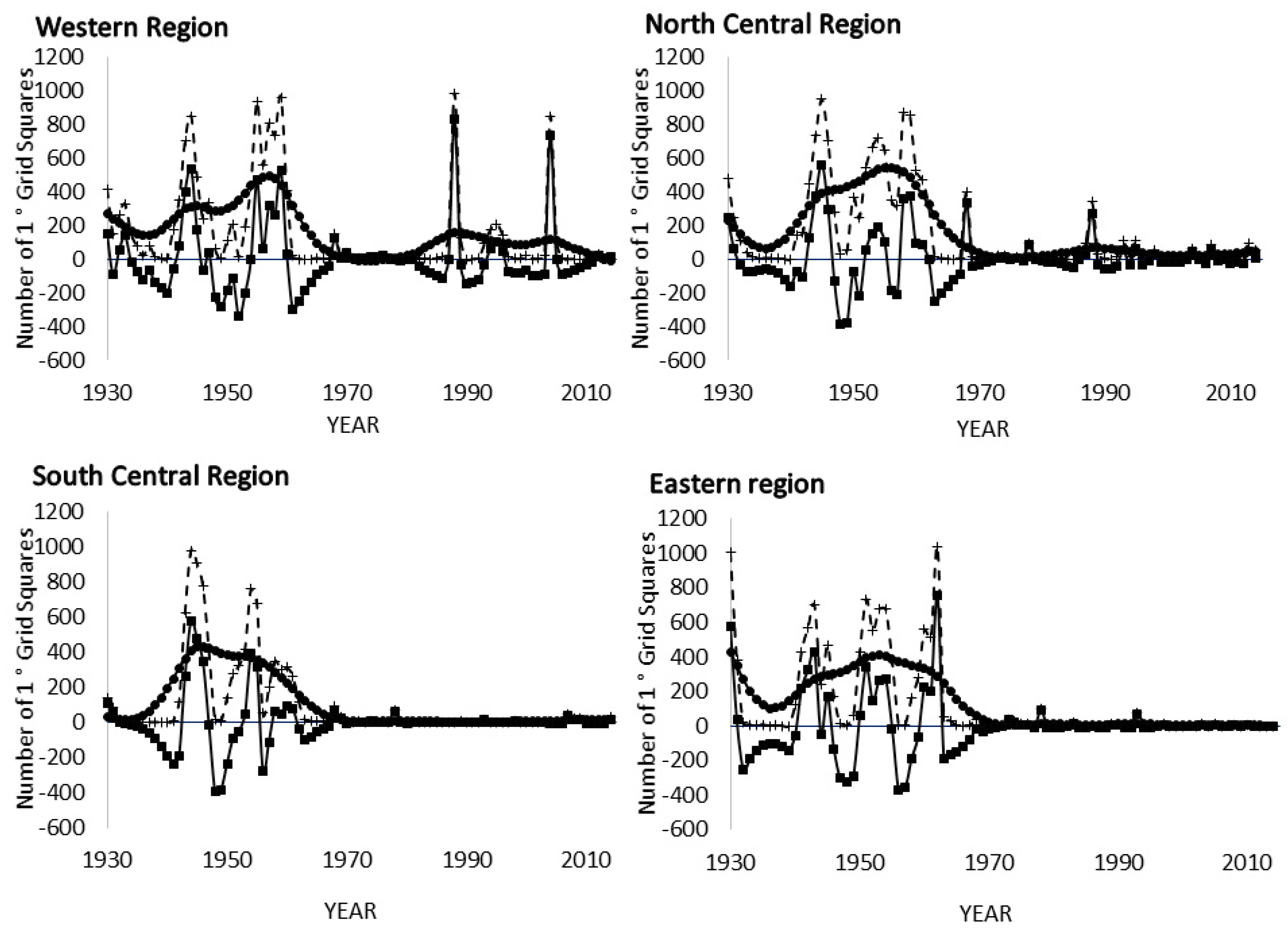
3.3. Spectral Analysis: Numbers of 1° Grid Squares Infested with Desert Locust Swarms, 1930–2014
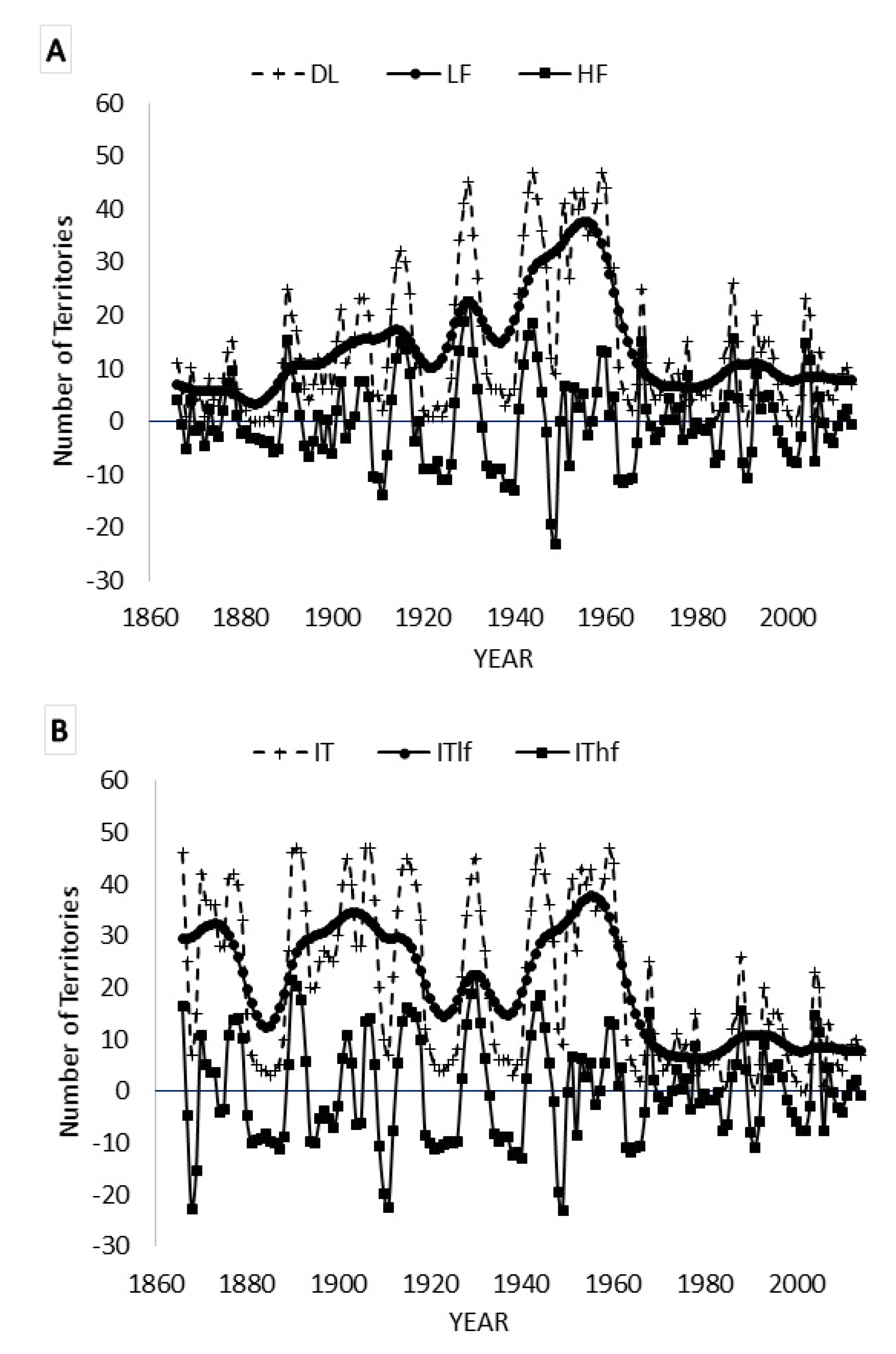
3.4. Kalman Filtering
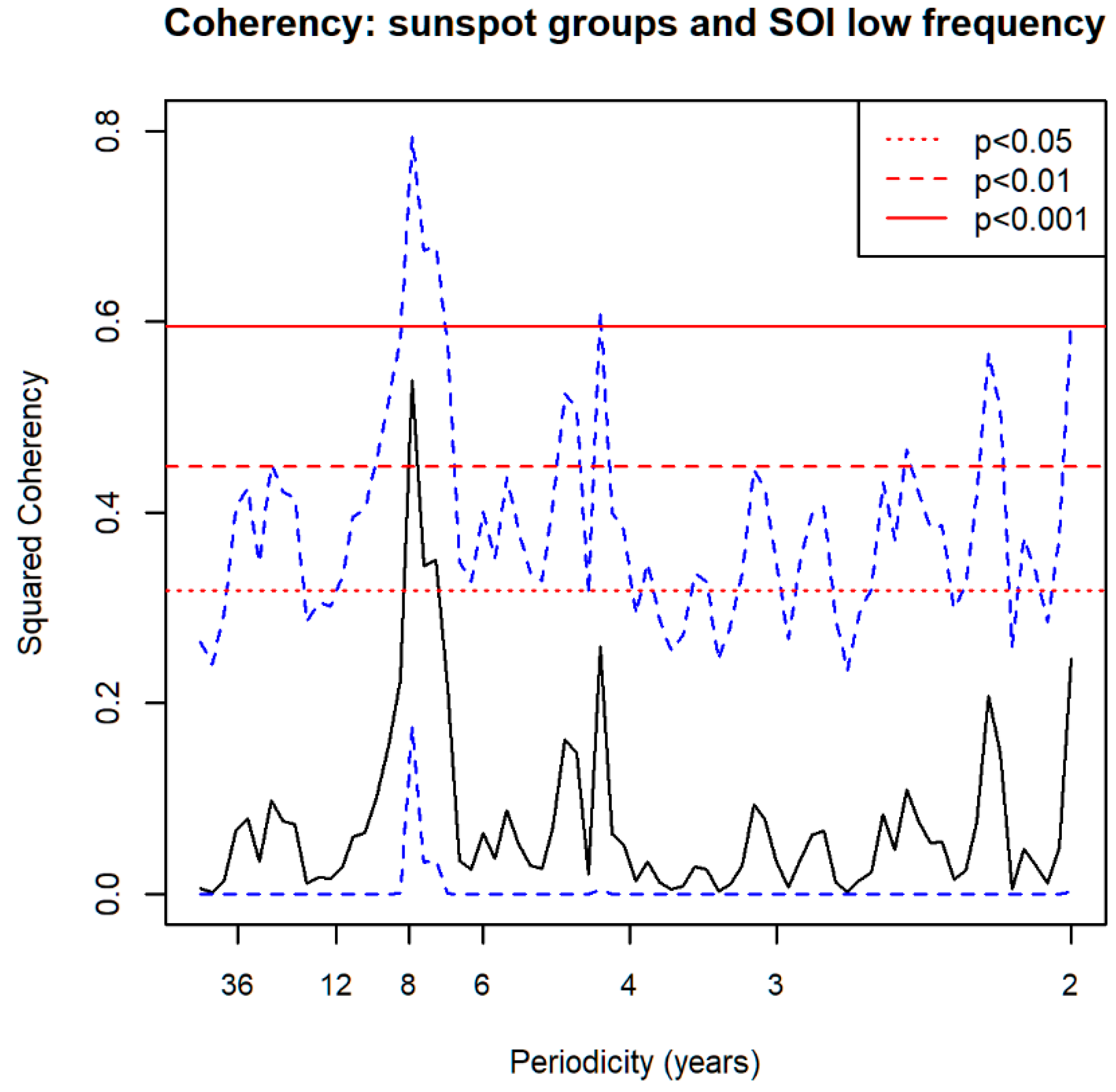
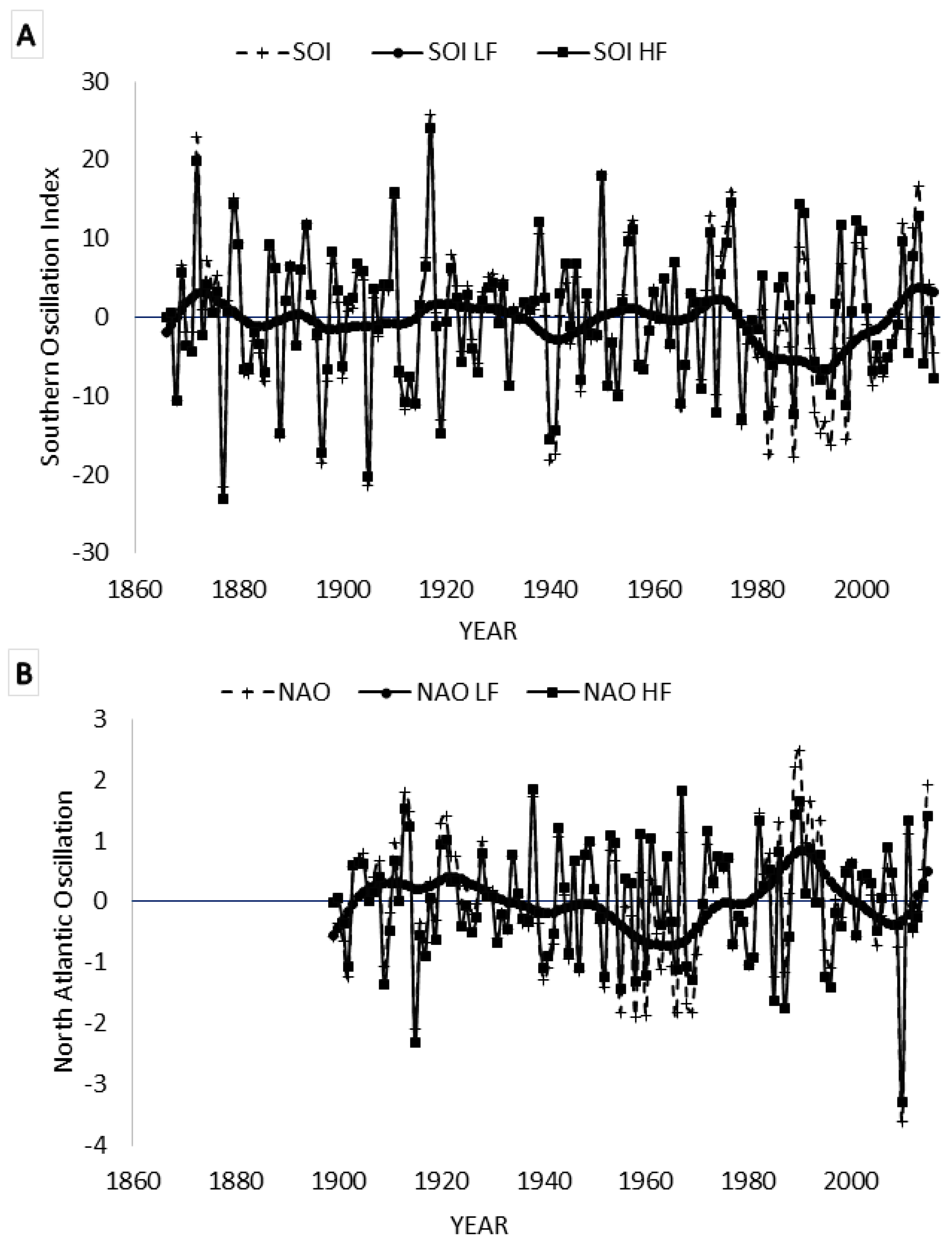
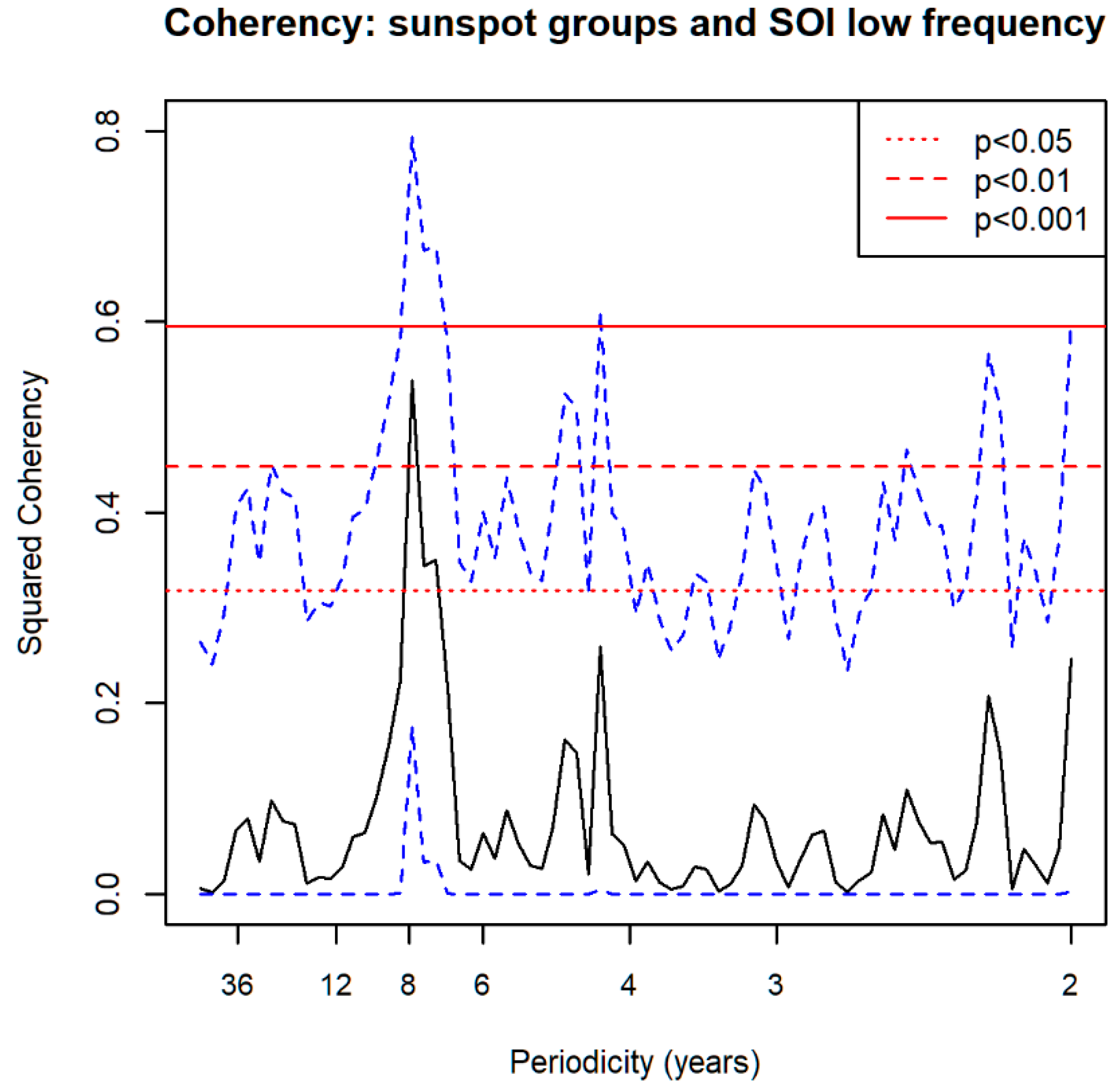
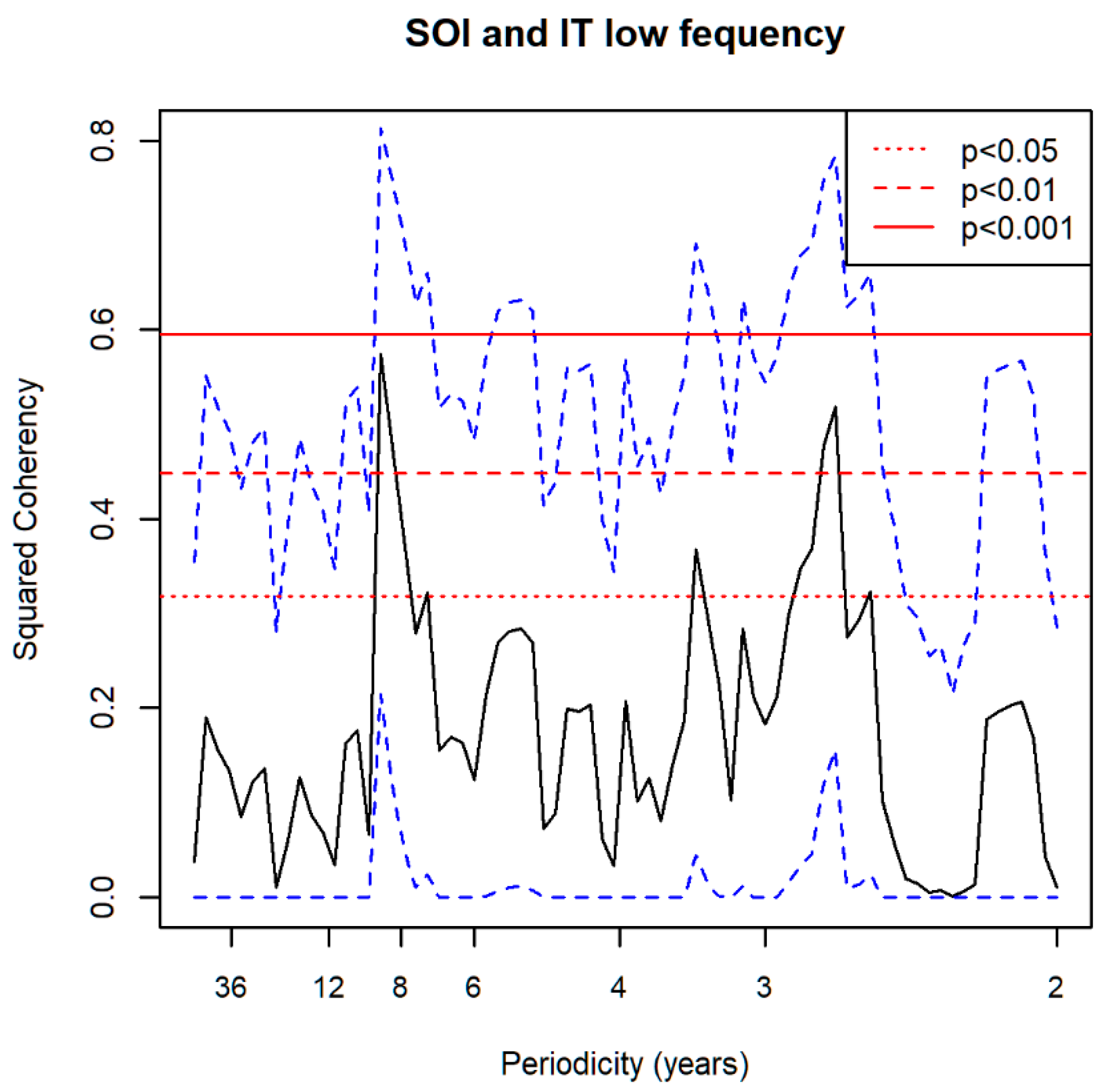
3.5. Spectral Coherence: Sunspots and Oceanic Systems
3.6. Spectral Coherence: Oriental Migratory Locusts
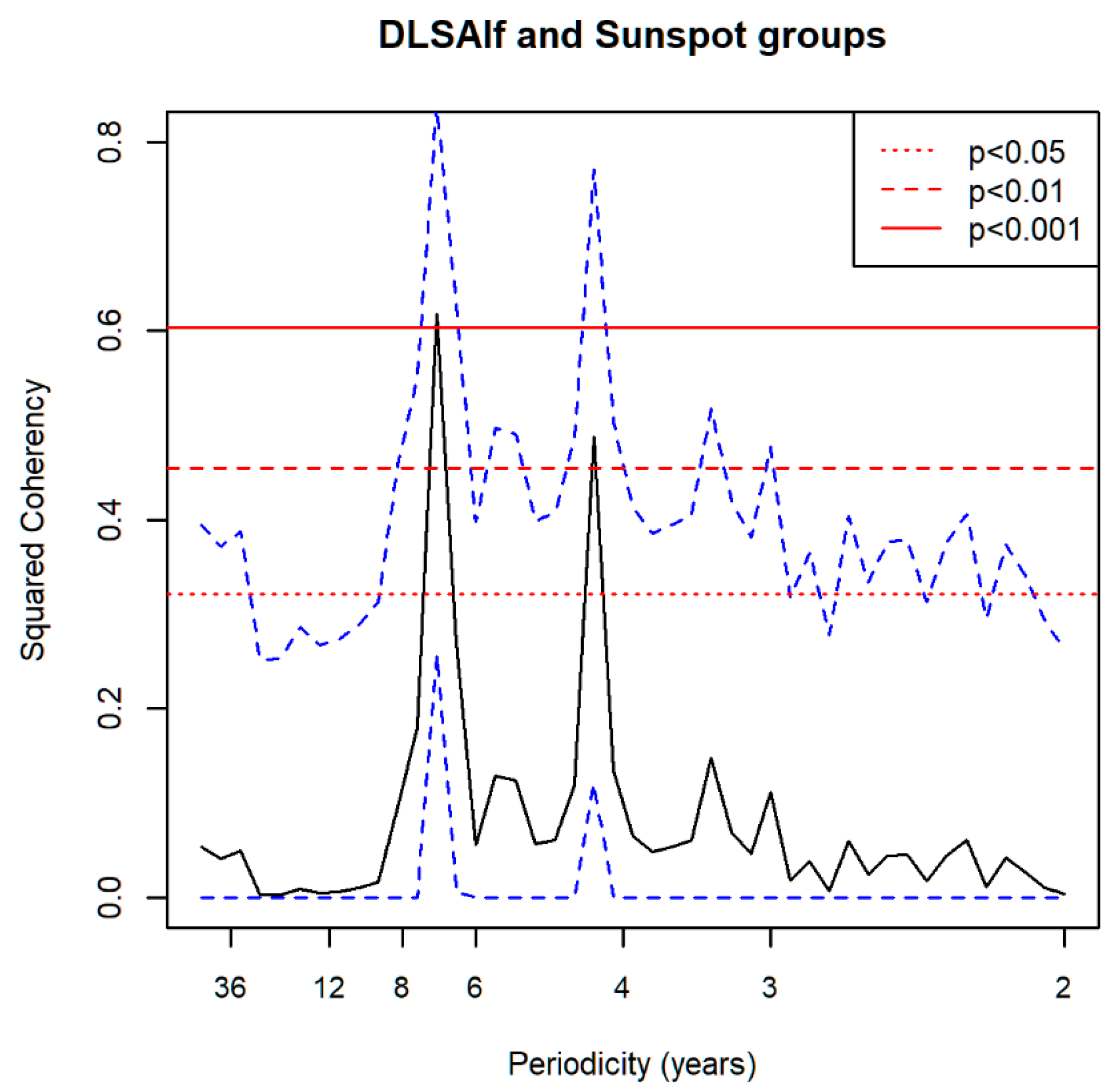
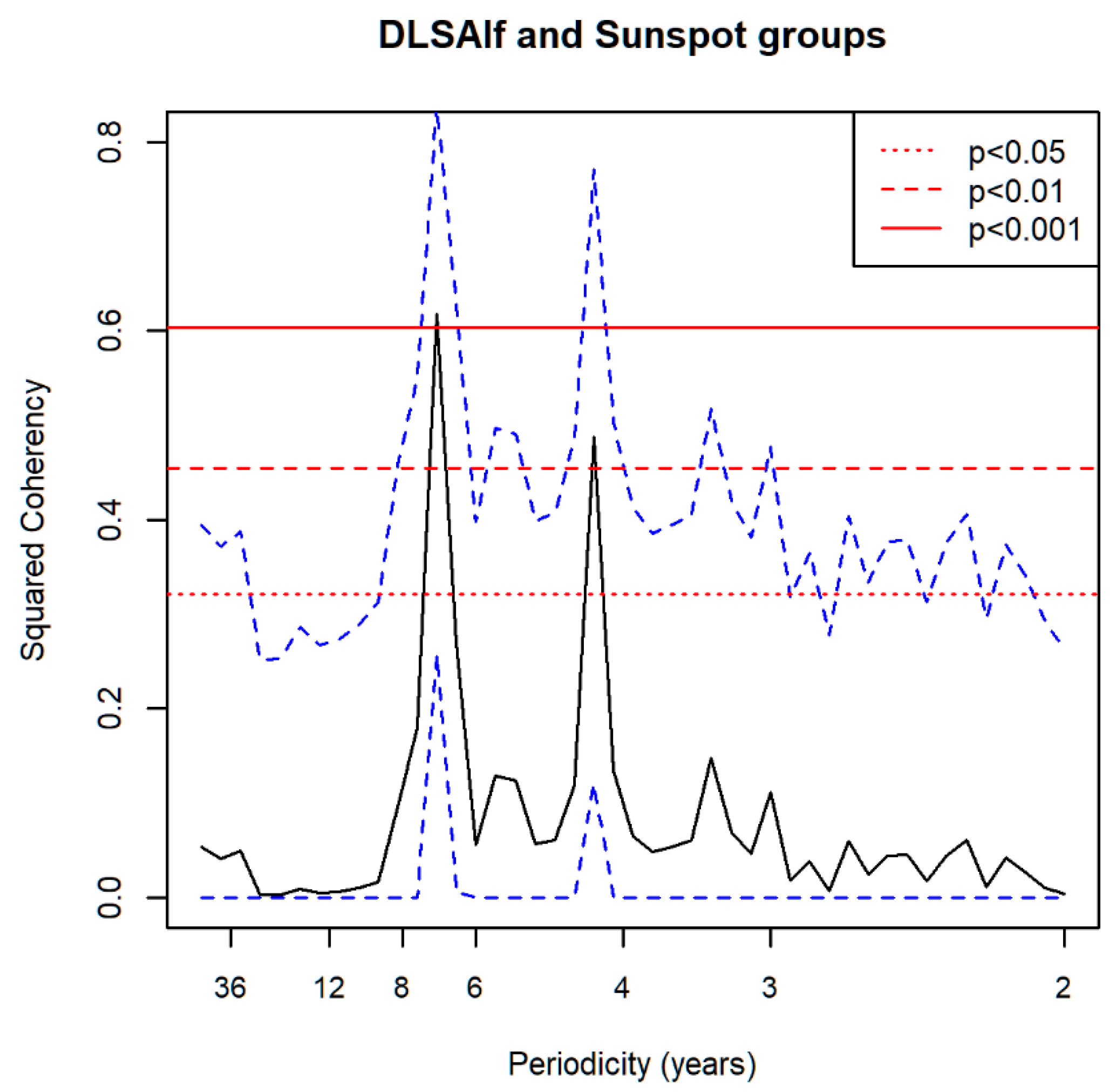
3.7. Spectral Coherence: Numbers of Territories Infested with Desert Locust Swarms, 1866–2014 (1866 to 2010 for Sunspot Groups Data)
3.8. Spectral Coherence: Numbers of 1° Grid Squares Infested with Desert Locust Swarms, 1930–2014
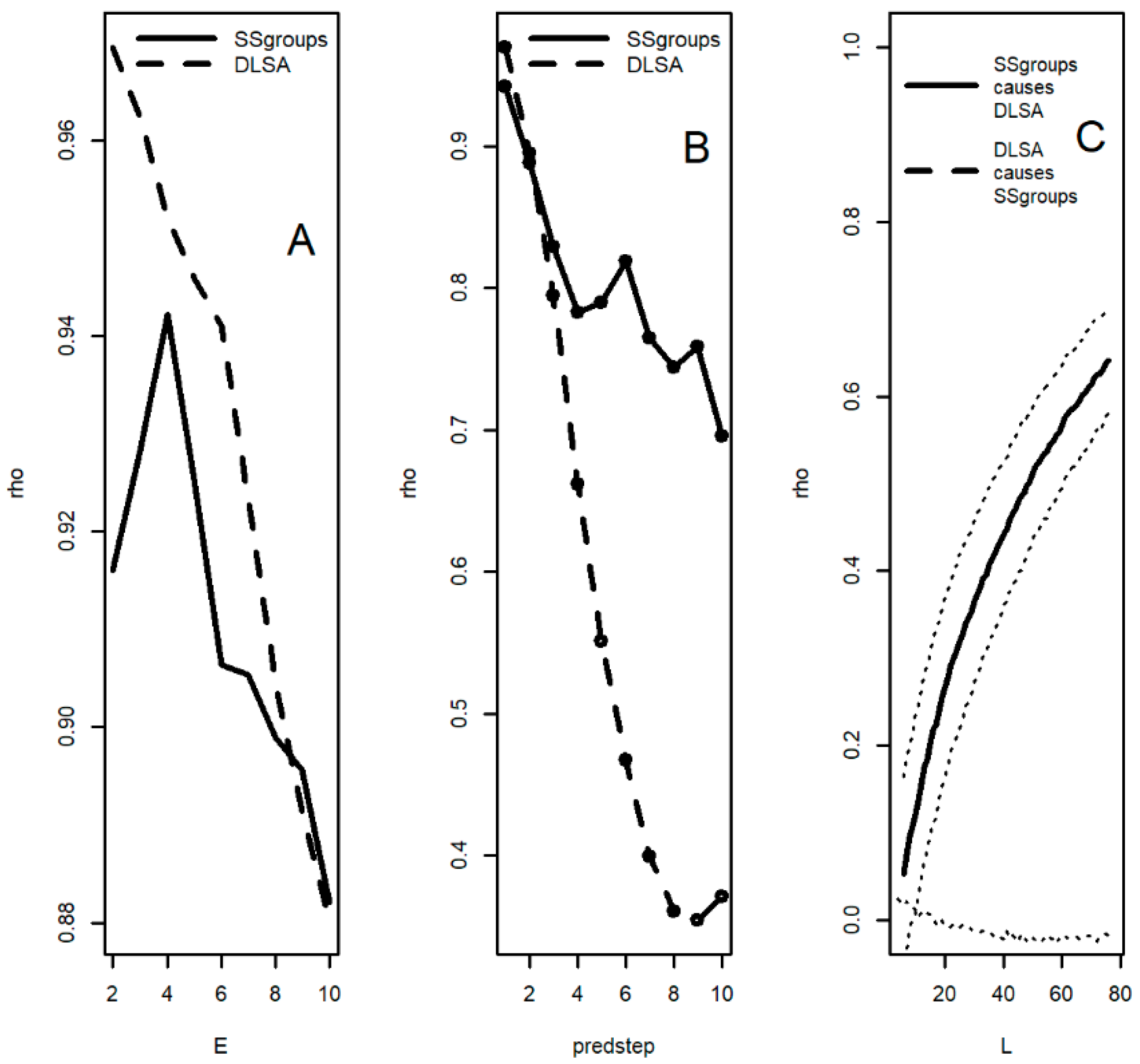
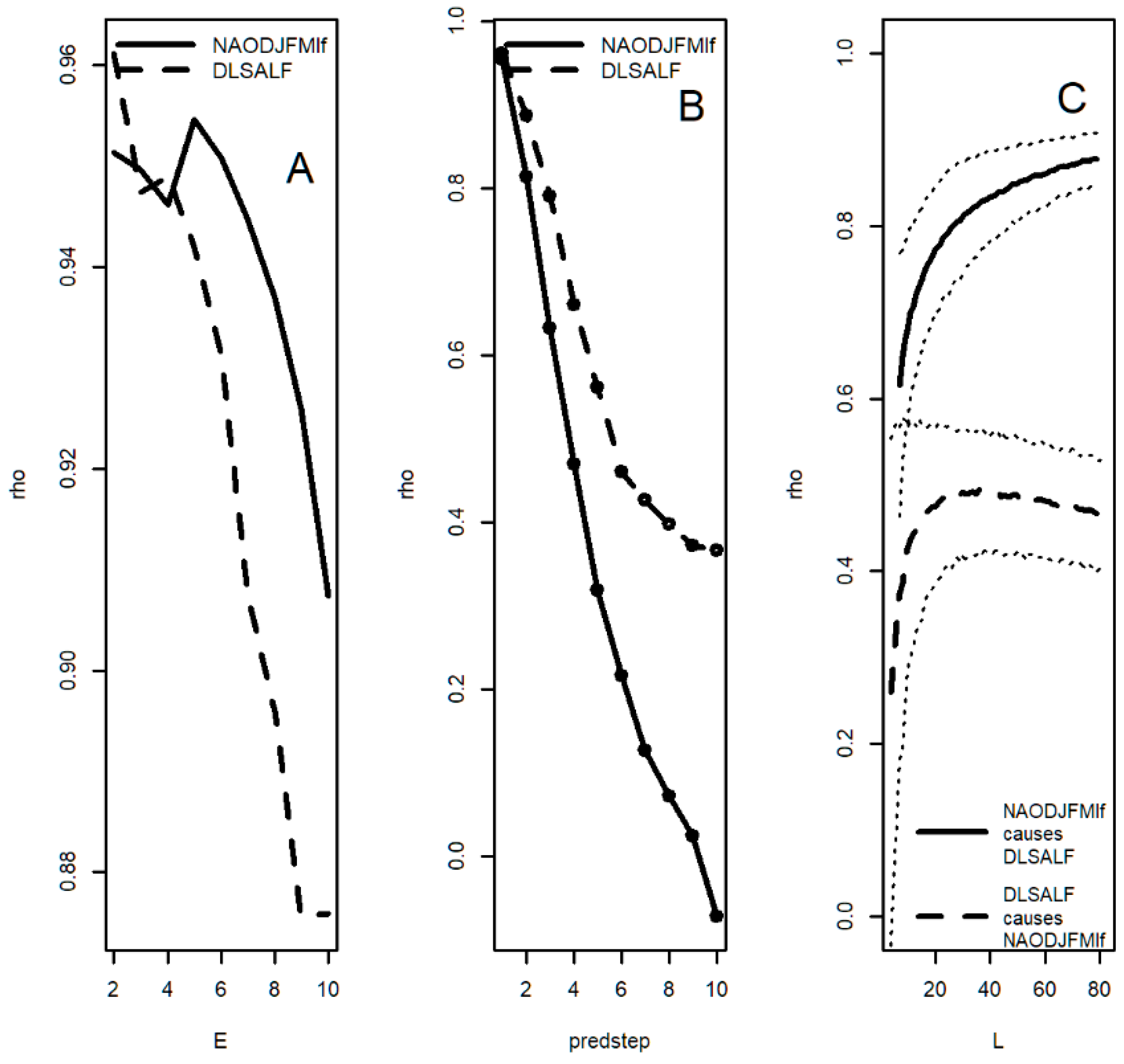
3.9. Convergent Cross Mapping: Sunspot Groups and Oceanic Systems
3.10. Convergent Cross Mapping: Oriental Migratory Locusts
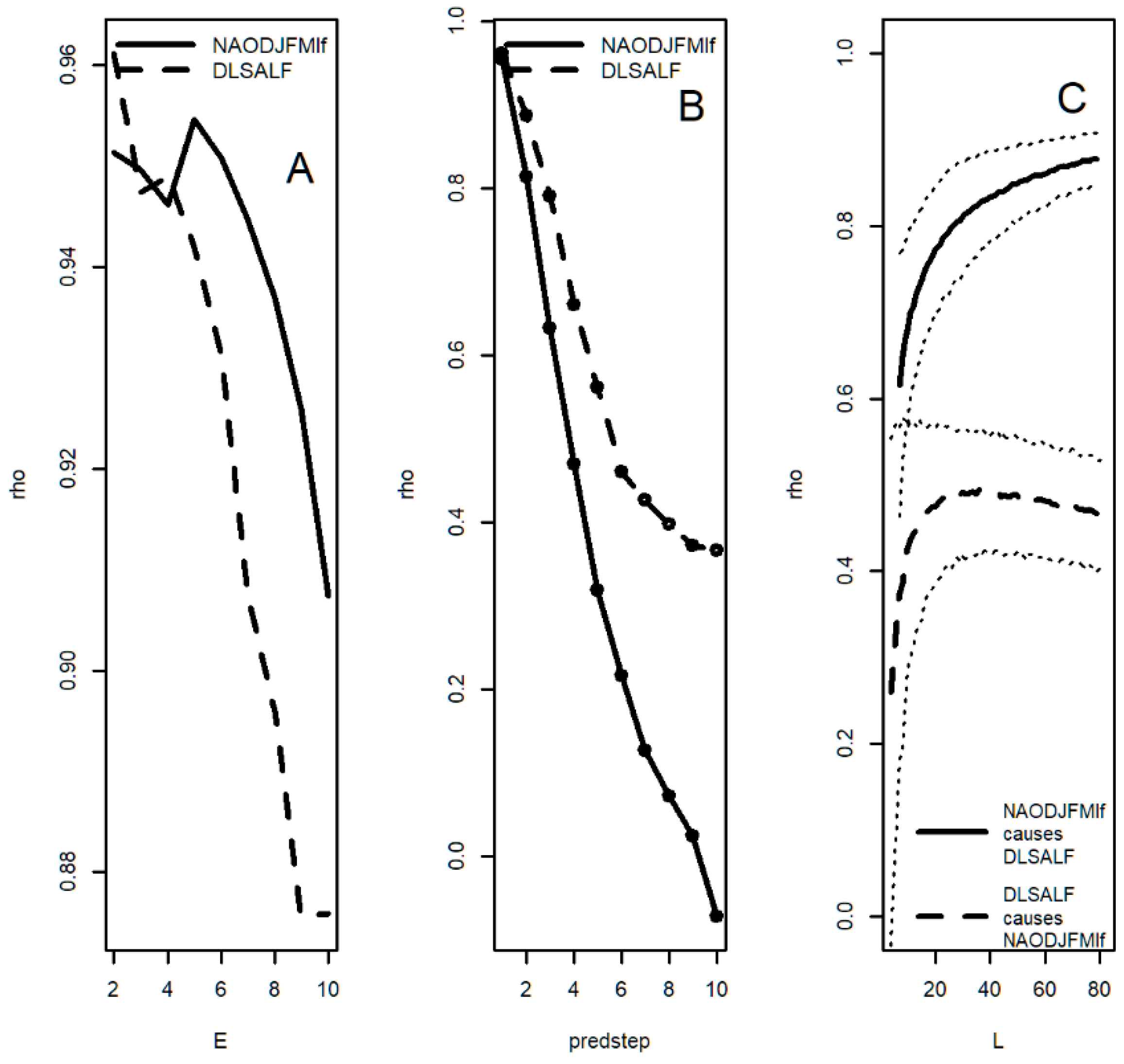
3.11. Convergent Cross Mapping (CCM): Desert Locusts
4. Discussion
5. Conclusions
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cisse, S.; Ghaout, S.; Mazih, A.; Ebbe, M.A.O.B.; Piou, C. Effect of vegetation on density thresholds of adult desert locust gregarization from survey data in Mauritania. Entomol. Exp. Appl. 2013, 149, 159–165. [Google Scholar] [CrossRef]
- Cisse, S.; Ghaout, S.; Mazih, A.; Ebbe, M.A.O.B.; Benahi, A.; Piou, C. Estimation of density threshold of gregarization of desert locust hoppers from field sampling in Mauritania. Entomol. Exp. Appl. 2015, 156, 136–148. [Google Scholar] [CrossRef]
- Uvarov, B. Grasshoppers and Locusts. A Handbook of General Acridology; Cambridge University Press: Cambridge, UK, 1966; Volume 1. [Google Scholar]
- Uvarov, B. Grasshoppers and Locusts. A Handbook of General Acridology; Centre for Overseas Pest Research: London, UK, 1977; Volume 2. [Google Scholar]
- Simpson, S.J.; McCaffery, A.R.; Hägele, B.F. A behavioural analysis of phase change in the desert locust. Biol. Rev. Camb. Philos. Soc. 1999, 74, 461–480. [Google Scholar] [CrossRef]
- Simpson, S.J.; Despland, E.; Hägele, B.F.; Dodgson, T. Gregarious behavior in desert locusts is evoked by touching their back legs. Proc. Natl. Acad. Sci. USA 2001, 98, 3895–3897. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Desert Locust Guidelines, 2nd ed.; FAO: Rome, Italy, 2001.
- Cullen, D.A.; Cease, A.J.; Latchininsky, A.V.; Ayali, A.; Berry, K.; Buhl, J.; De Keyser, R.; Foquet, B.; Hadrich, J.C.; Matheson, T.; et al. From molecules to management: Mechanisms and consequences of locust phase polyphenism. Adv. Insect Physiol. 2017, 53, 167–285. [Google Scholar] [CrossRef]
- Cheke, R.A.; Holt, J. Complex dynamics of desert locust plagues. Ecol. Entomol. 1993, 18, 109–115. [Google Scholar] [CrossRef]
- Tratalos, J.A.; Cheke, R.A.; Healey, R.; Stenseth, N.-C. Desert locust populations, rainfall and climate change: Insights from phenomenological models using gridded monthly data. Clim. Res. 2010, 43, 229–239. [Google Scholar] [CrossRef]
- Pedgley, D.E. (Ed.) The Desert Locust Forecasting Manual; Centre for Overseas Pest Research: London, UK, 1981. [Google Scholar]
- Cressman, K. Information and forecasting. In Desert Locust Guidelines, 2nd ed.; Food and Agriculture Organization of the United Nations: Rome, Italy, 2001; Chapter 3. [Google Scholar]
- Vallebona, C.; Genesio, L.; Crisci, A.; Pasqui, M.; Di Vecchia, A.; Maracchi, G. Large-scale climatic patterns forcing desert locust upsurges in West Africa. Clim. Res. 2008, 37, 35–41. [Google Scholar] [CrossRef]
- Todd, M.C.; Washington, R.; Cheke, R.A.; Kniveton, D. Brown locust outbreaks and climate variability in southern Africa. J. Appl. Ecol. 2002, 39, 31–42. [Google Scholar] [CrossRef]
- Swinton, A.H. Data obtained from solar physics and earthquake commotions applied to elucidate locust multiplication and migration. In Third Report of the United States Entomological Commission, Relating to the Rocky Mountain Locust, the Western Cricket, the Armyworm Canker-Worms, and the Hessian-Fly; Riley, C.V., Packard, A.S., Thomas, C., Eds.; Government Printing Office: Washington, DC, USA, 1883; pp. 65–85. [Google Scholar]
- Ramchandra Rao, Y. A preliminary note on a study of locust infestations in North-West India since 1863 in relation to sunspot cycles. In Proceedings of the 5th International Locust Conference, Brussels, Belgium, 25 August–1 September 1938; pp. 252–257. [Google Scholar]
- Ramchandra Rao, Y. The Desert Locust in India; No. 21; Indian Council for Agricultural Research: New Delhi, India, 1960; p. 721.
- Pasquier, R. Les sauterelles pèlerines. L’invasion actuelle, les recherches, la lutte. L’Agria 1942, 99, 1–12. [Google Scholar]
- Shcherbinovskiǐ, N.S. The Desert Locust, Schistocerca gregaria. The Problem of Defending the Southern Territories of the U.S.S.R. Against Invasions by Swarms of Schistocerca; Gosudarstvennoe Izdatel’stvo Sel’skohozâjstvennoj Literatury: Moscow, Russia, 1952. [Google Scholar]
- Shumakov, E.M. Study of Locust Ecology in the U.S.S.R. with Special Reference to the Development of Opinions on Phase Variation; No. 114; Colloques Internationaux du Centre National de la Recherche Scientifique: Paris, France, 1962; pp. 217–228. [Google Scholar]
- Meehl, G.A.; Arblaster, J.M.; Matthes, K.; Sassi, F.; van Loon, H. Amplifying the Pacific climate system response to a small 11-year solar cycle forcing. Science 2009, 325, 1114–1118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsonis, A.A.; Deyle, E.R.; May, R.M.; Sugihara, G.; Swanson, K.; Verbeten, J.D.; Wang, G. Dynamical evidence for causality between galactic cosmic rays and interannual variation in global temperature. Proc. Natl. Acad. Sci. USA 2015, 112, 3253–3256. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Visbeck, M.H.; Hurrell, J.W.; Polvani, L.; Cullen, H.M. The North Atlantic Oscillation: Past, present, and future. Proc. Natl. Acad. Sci. USA 2001, 98, 12876–12877. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tian, H.; Stige, L.C.; Cazelles, B.; Kausrud, K.L.; Svarverud, R.; Stenseth, N.-C.; Zhang, Z. Reconstruction of a 1910-y-long locust series reveals consistent associations with climate fluctuations in China. Proc. Natl. Acad. Sci. USA 2011, 108, 14521–14526. [Google Scholar] [CrossRef] [Green Version]
- Waloff, Z.; Green, S.M. Some temporal characteristics of Desert Locust plagues with a statistical analysis. In Anti-Locust Memoir; Centre for Overseas Pest Research: London, UK, 1976. [Google Scholar]
- Cheke, R.A.; Tratalos, J.A. Migration, patchiness and population processes illustrated by two migrant pests. BioScience 2007, 57, 145–154. [Google Scholar] [CrossRef] [Green Version]
- Hoyt, D.V.; Schatten, K.H. Group Sunspot Numbers: A New Solar Activity Reconstruction. Sol. Phys. 1998, 179, 189–219. [Google Scholar] [CrossRef]
- Hoyt, D.V.; Schatten, K.H. Group Sunspot Numbers: A New Solar Activity Reconstruction. Sol. Phys. 1998, 181, 491–512. [Google Scholar] [CrossRef]
- Vaquero, J.M.; Svalgaard, L.; Carrasco, V.M.S.; Clette, F.; Lefèvre, L.; Gallego, M.C.; Arlt, R.; Aparicio, A.J.P.; Richard, J.-G.; Howe, R. Revised collection of the number of sunspot groups from 1610 to 2010. Sol. Phys. 2016, 291, 3061–3074. [Google Scholar] [CrossRef] [Green Version]
- Meehl, G.A.; van Loon, H. The sea-saw in winter temperatures between Greenland and Northern Europe, Part 3: Teleconnections with lower latitudes. Mon. Weather Rev. 1979, 107, 1095–1106. [Google Scholar] [CrossRef] [Green Version]
- Abram, N.J.; Gagan, M.K.; Cole, J.E.; Hantoro, W.S.; Mudelsee, M. Recent intensification of tropical climate variability in the Indian Ocean. Nat. Geosci. 2008, 1, 849–853. [Google Scholar] [CrossRef]
- R Core Team. The R Project for Statistical Computing. Available online: https://www.R-project.org/ (accessed on 9 August 2020).
- Louca, S.; Doebeli, M. Detecting cyclicity in ecological time series. Ecology 2015, 96, 1724–1732. [Google Scholar] [CrossRef]
- Chatfield, C. The Analysis of Time Series: An Introduction; Chapman & Hall: New York, NY, USA, 1992. [Google Scholar]
- Shumway, R.H.; Stoffer, D.S. Time Series: A Data Analysis Approach Using R; CRC Press: Boca Raton, FL, USA, 2019. [Google Scholar]
- Sugihara, G.; May, R.; Ye, H.; Hsieh, C.; Deyle, E.; Fogarty, M.; Munch, S. Detecting causality in complex ecosystems. Science 2012, 338, 496–500. [Google Scholar] [CrossRef] [PubMed]
- Bendat, J.S.; Piersol, A.G. Random Data: Analysis and Measurement Procedures; John Wiley and Sons, Inc.: New York, NY, USA, 2000. [Google Scholar]
- Granger, C.W.J. Investigating causal relations by econometric models and cross-spectral methods. Econometrica 1969, 37, 424–438. [Google Scholar] [CrossRef]
- Clark, A.T.; Ye, H.; Isbell, F.; Deyle, E.R.; Cowles, J.; Tilman, G.D.; Sugihara, G. Spatial convergent cross mapping to detect causal relationships from short time series. Ecology 2015, 96, 1174–1181. [Google Scholar] [CrossRef]
- Chang, C.-W.; Ushio, M.; Hseih, C.-H. Empirical dynamic modeling for beginners. Ecol. Res. 2017, 32, 785–796. [Google Scholar] [CrossRef]
- Takens, F. Detecting strange attractors in turbulence. In Dynamical Systems and Turbulence; Rand, D.A., Young, L.-S., Eds.; Springer: Berlin, Germany, 1981; pp. 366–381. [Google Scholar]
- Sugihara, G.; May, R.M. Nonlinear forecasting as a way of distinguishing chaos from measurement error in time series. Nature 1990, 344, 734–741. [Google Scholar] [CrossRef]
- Ye, H.; Deyle, E.R.; Gilarranz, L.J.; Sugihara, G. Distinguishing time-delayed causal interactions using convergent cross mapping. Sci. Rep. 2015, 5, 14750. [Google Scholar] [CrossRef] [Green Version]
- Chen, D.; Xiao, Y.; Cheke, R.A. Early warning signals for an environmental driver of respiratory infections. Bull. Math. Biol. 2021. In Review. [Google Scholar]
- Chiodo, G.; Oehrlein, J.; Polvani, L.M.; Fyfe, J.C.; Smith, A.K. Insignificant influence of the 11-year solar cycle on the North Atlantic Oscillation. Nat. Geosci. 2019, 12, 94–99. [Google Scholar] [CrossRef]
- Waloff, Z.; Green, S.M. Regularities in duration of regional desert locust plagues. Nature 1975, 256, 484–485. [Google Scholar] [CrossRef]
- Waloff, Z. The upsurges and recessions of the Desert Locust plague: An historical survey. In Anti-Locust Memoir no. 8; Anti-Locust Research Centre: London, UK, 1966. [Google Scholar]
- Lamb, P.J. Case studies of tropical Atlantic circulation patterns during recent sub-Saharan weather anomalies: 1967 and 1968. Mon. Weather Rev. 1978, 106, 482–491. [Google Scholar] [CrossRef] [Green Version]
- Lamb, P.J.; Peppler, R.A. North Atlantic oscillation: Concept and an application. Bull. Am. Meteorol. Soc. 1987, 68, 1218–1225. [Google Scholar] [CrossRef]
- Kühberger, A.; Fritz, A.; Lermer, E.; Scherndl, T. The significance fallacy in inferential statistics. BMC Res. Notes 2015, 8, 84. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Data Set | Unit | Interval | Start Date | End Date | Source |
|---|---|---|---|---|---|
| Oriental Migratory Locust series | Counties with locusts multiplied by outbreak intensity (graded 1–3) adjusted for recording effort (Ladj in [9]) | Annual | 1610 | 1911 | Tian et al. (2011) |
| Total Desert Locust (DL) | Territories | Annual | 1866 | 2015 | FAO |
| Total Desert Locust Inferred Series (IT) | Territories | Annual | 1866 | 2015 | FAO |
| Total Desert Locust 1° grid square series (DLSA) | 1° grid square | Annual | 1930 | 2014 | Converted by JAT from FAO data |
| Regional versions of DLSA for the Western (DLSAW), North Central (DLSANC), South Central (DLSASC) and Eastern Regions (DLSAE) | |||||
| Southern Oscillation Index (SOI) | Annual | 1866 | 2015 | http://www.esrl.noaa.gov/psd/gcos_wgsp/Timeseries/SOI/ | |
| North Atlantic Oscillation (NAO) | Annual, DJF, MAM, JJA, SON & DJFM series | 1899 | 2016 | https://climatedataguide.ucar.edu/climate-data/hurrell-north-atlantic-oscillation-nao-index-pc-based. (Principal Components based time series of the leading Empirical Orthogonal Function of Sea Level Pressure anomalies over the Atlantic sector, 20°–80° N, 90° W–40° E.) | |
| Indian Ocean Dipole | Coral reconstruction of mean unfiltered dipole mode index | Annual | 1846 | 1994 | https://www.ncdc.noaa.gov/paleo-search/study/8607; [31] |
| Sunspot Group Numbers | Annual | 1610 | 2010 | http://www.sidc.be/silso/groupnumberv3 | |
| Time Series 2 (Response) | |||||
|---|---|---|---|---|---|
| NAO | NAOlf | IOD | SOI | SOIlf | |
| Time Series 1 (Driver) | |||||
| Spectral Coherence | |||||
| Sunspot Groups | 4.05 ** | 4.05 ***, 5.79 * | 4.50 *, 2.37 ** | NS | 7.89 ** |
| Convergence Cross Mapping | |||||
| Sunspot Groups | NS | p < 0.0001 | NS | NS | p = 0.04 |
| Time Series 2 (Response) | ||||||||
|---|---|---|---|---|---|---|---|---|
| IT | ITlf | DLSA | DLSAlf | DLSAWlf | DLSANClf | DLSASClf | DLSAElf | |
| Time Series 1 (Potential Driver) | ||||||||
| Sunspot groups | 22.86 *, 2.32 ** | 7.14 *, 2.27 * | 6.23 * | 6.75 ***, 4.26* | 6.75 * | 6.75 **, 4.26 * | 6.75 * | 6.75 *** |
| SOI | 8.82 **, 7.89 *, 4.29 *, 3.95 *, 2.68 ***, 2.54 ** | 8.83 **, 8.33 *, 3.41 *, 2.67 **, 2.54 * | NS | NS | NS | NS | NS | NS |
| SOIlf | 10.71 *, 7.14 *, 5.17 *, 4.0 **, 3.75 *, 2.68 **, 2.54 ** | 15.0 *, 10.71 *, 8.82 ***, 7.14 **, 5.0*, 3.41 *, 3.09 * 2.68 **, 2.54 * | NS | 6.43 **, 4.74 ***, 2.90 * | 12.86 *, 6.92 ***, 4.09 **, 3.21 ** | 6.42 *, 5.0 ***, 3.10 *, 2.90 * | 5.0 ***, 3.21 *, 2.90 * | 6.43 **, 5.0 **, 2.90 *, 2.14 ** |
| NAO | 2.10 ** | 3.24 *, 2.10 ** | NS | 4.5 * | 4.5 * | 4.5 *, 4.09 *, 3.75 *, 3.33 * | NS | NS |
| NAOlf | 9.23 ***, 6.32 ***, 2.10 *** | 60.0 *, 17.14 **, 9.23 ***, 6.0 **, 3.16 *, 2.10 *** | NS | 15.0 *, 8.18 ***, 4.5 *** | 18.0 *, 8.18 *, 6.0 *, 4.74 ** | 15.0 *, 8.18 ***, 4.5 * | 8.18 ***, 5.29 *, 4.74 *, 4.5 *, 2.19 *, 2.0 * | 20 *, 7.5 *, 6.92 **, 4.5 ***, 3.12 *, 2.18 * |
| NAODJFlf (winter) | 9.23 *, 6.0 ** | 17.14 ***, 9.23 ***, 6.0 ***, 5.22 * | NS | 15.0 *, 8.18 ***, 4.5 *** | 15.0 *, 8.18 *, 6.0 *, 4.74 **, 4.29 * | 15.0 *, 8.18 *** 4.74 *** | 8.18 ***, 5.29 *, 4.74 * | 15.0 *, 6.92 **, 4.5 ***, 3.21 *, 2.90 * |
| NAOMAMlf (spring) | 9.23 *, 6.0 * | 9.23 ***, 6.0 ***, 3.24 * | 3.21* | 15.0 *, 10.0 *, 6.43 *, 4.74 *** | 18.0 *, 9.0 **, 6.43 ***, 4.5 * | 15.0 **, 10.0 *, 6.43 **, 4.74 *** | 15.0 **, 9.0 *, 7.5 *, 4.74 *** | 18.0 **, 6.43 ***, 4.74 ***, 2.81 * |
| NAOJJAlf (summer) | 20 *,10 *, 6.67 *, 4.8 * | 20 **, 13.33 **, 10 **, 8 **, 6.67 **, 4.8 **, 3.53 *** | NS | 30.0 **, 15.0 **, 9.0 ***, 6.43 *, 5.29 ***, 4.28 *, 3.10 * | 30.0 **, 15.0 **, 11.25 **, 9.0*, 6.92 **, 5.29 *, 4.29 **, 3.21 *, 2.09 * | 15.0 **, 9.0 ***, 5.29 ** | 30.0 **, 15.0 **, 9.0 ***, 5.29 ***, 3.21 ** | 15.0 **, 9.0 **, 6.0 **, 5.29 **, 4.29 ***, 3.10 **, 2.09 ** |
| NAOSONlf (autumn) | 9.23 *, 3.64 *, 2.40 ** | 15.0 *, 9.23 ***, 6.0 *, 4.8 *, 3.33 * 2.40 *** | 9.0 *, 5.62 * | 9.0 ***, 6.43 **, 5.0 ** | 12.86 *, 9.0 ***, 6.92 ***, 3.33 * | 9.0 ***, 6.43 **, 5.0 *** | 11.25 ***, 9.0 ***, 5.0 *** | 9.0 *, 6.43 ***, 5.0 * |
| IOD (Mean DMI) | 12.27 *, 3.21 * | 3.37 ** | NS | 7.20 * | NS | NS | NS | NS |
| Time Series 2 (Response) | |||||||
|---|---|---|---|---|---|---|---|
| ITlf | DLSA | DLSAlf | DLSAWlf | DLSANClf | DLSASClf | DLSAElf | |
| Time Series 1 (Driver) | |||||||
| Sunspot Groups | <0.0001 | <0.0001 | <0.0001 | <0.0001 | <0.0001 | <0.0001 | <0.0001 |
| SOIlf | <0.005 | NS | <0.0001 | NS | NS | 0.001 | 0.002 |
| NAOlf | NS | NS | <0.0001 | 0.001 | <0.005 | <0.0001 | 0.003 |
| NAODJFlf | NS | NS | 0.034 | NS | NS | 0.0007 | NS |
| NAOMAMlf | <0.0001 | NS | <0.0001 | NS | 0.009 | <0.0001 | 0.0007 |
| NAOJJAlf | 0.01 | NS | 0.038 | NS | 0.021 | <0.005 | <0.0001 |
| NAOSONlf | <0.0001 | NS | 0.048 | 0.05 | <0.003 | <0.003 | NS |
| NAODJFMlf | NS | NS | 0.0007 | 0.002 | 0.004 | <0.0001 | <0.003 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cheke, R.A.; Young, S.; Wang, X.; Tratalos, J.A.; Tang, S.; Cressman, K. Evidence for a Causal Relationship between the Solar Cycle and Locust Abundance. Agronomy 2021, 11, 69. https://doi.org/10.3390/agronomy11010069
Cheke RA, Young S, Wang X, Tratalos JA, Tang S, Cressman K. Evidence for a Causal Relationship between the Solar Cycle and Locust Abundance. Agronomy. 2021; 11(1):69. https://doi.org/10.3390/agronomy11010069
Chicago/Turabian StyleCheke, Robert A., Stephen Young, Xia Wang, Jamie A. Tratalos, Sanyi Tang, and Keith Cressman. 2021. "Evidence for a Causal Relationship between the Solar Cycle and Locust Abundance" Agronomy 11, no. 1: 69. https://doi.org/10.3390/agronomy11010069
APA StyleCheke, R. A., Young, S., Wang, X., Tratalos, J. A., Tang, S., & Cressman, K. (2021). Evidence for a Causal Relationship between the Solar Cycle and Locust Abundance. Agronomy, 11(1), 69. https://doi.org/10.3390/agronomy11010069