Expected Changes to Alpine Pastures in Extent and Composition under Future Climate Conditions

 ,
,  , , , , and
, , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Methodology
2.2.1. Predictor Variables: Topographic, Soil, and Climate Dataset
2.2.2. Response Variables: Pasturelands and Pasture Macro-Types Dataset
2.2.3. Random Forest: Model Set Up, Calibration, and Validation
3. Results
3.1. RF Calibration and Classification Accuracy
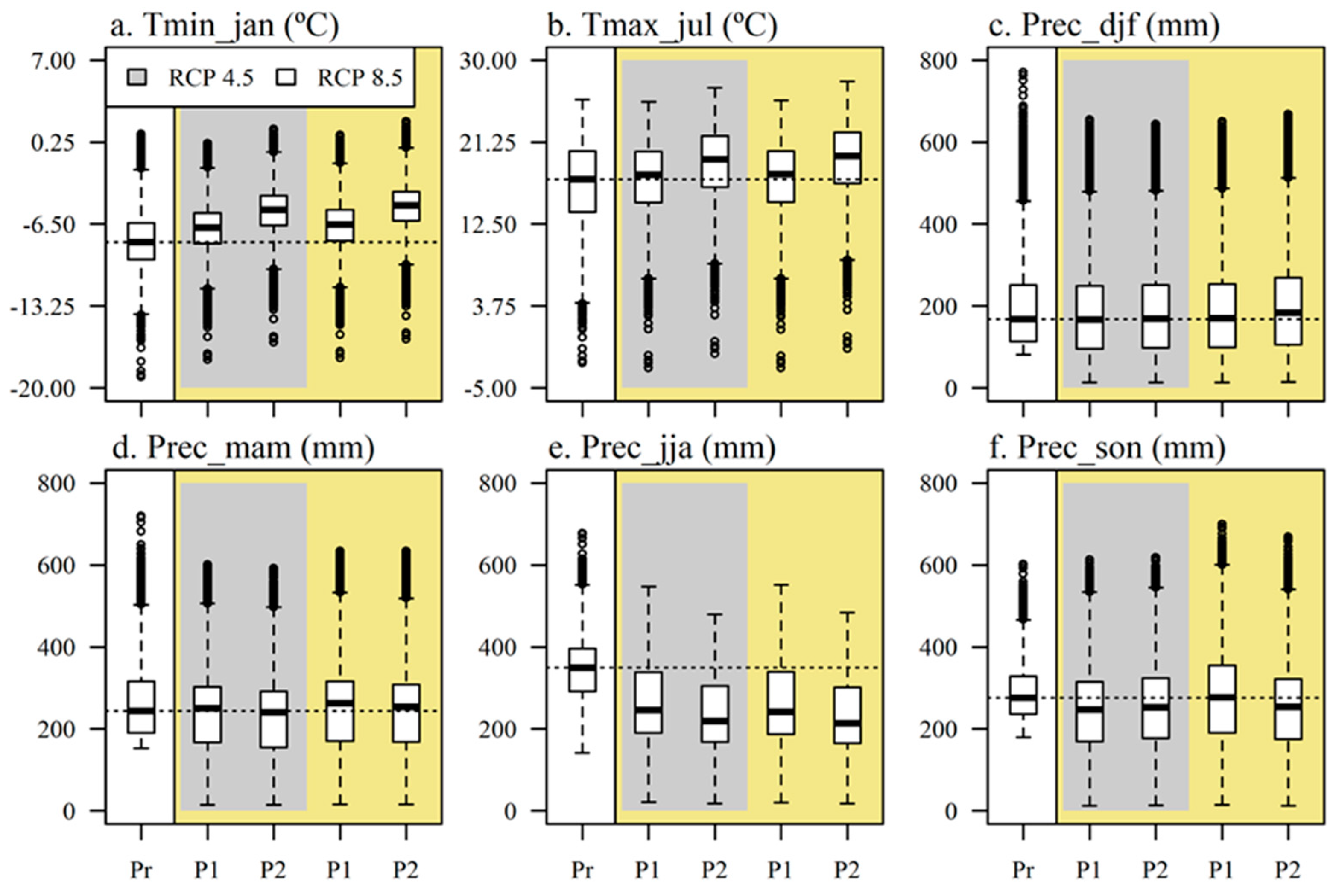
3.2. Climate Analysis
3.3. Altitude Pattern Dynamics of Pasture Macro-Types Suitability
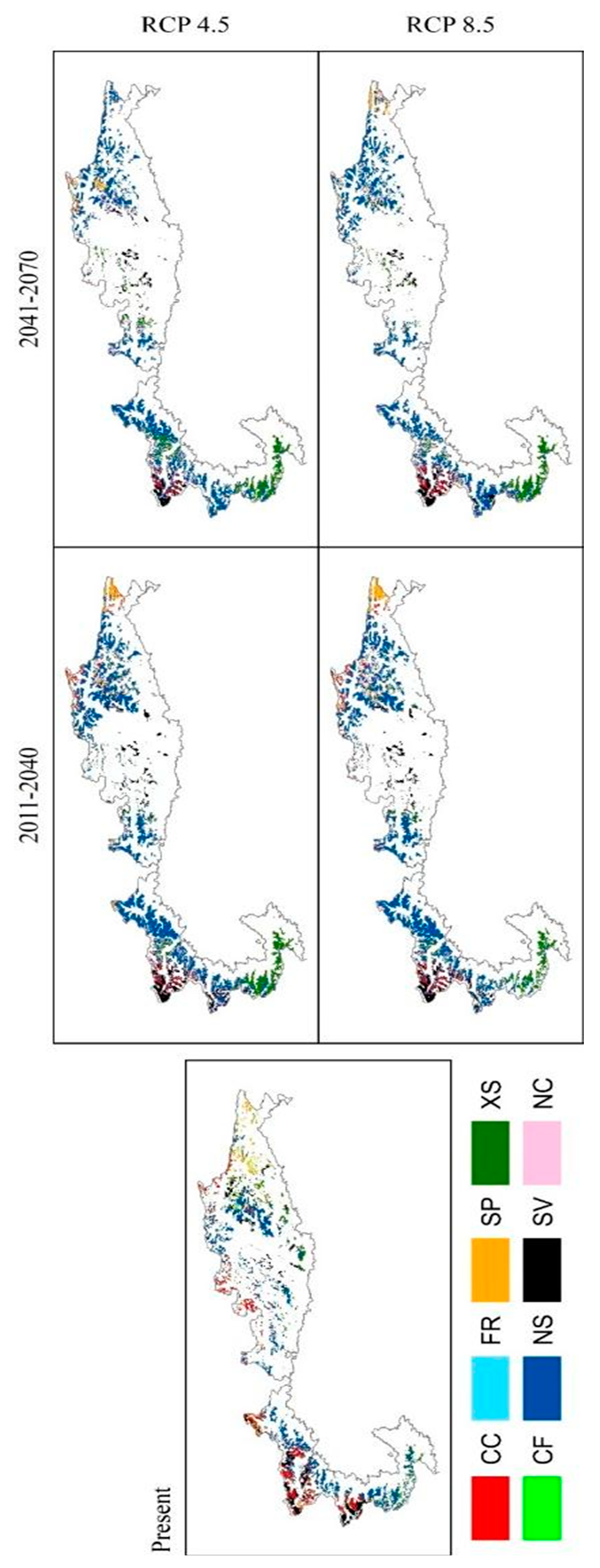
3.4. Global Dynamics of Alpine Pasturelands Suitability
3.5. Pasture Macro-Type Analysis under Future Conditions
3.5.1. Changes of Pasture Macro-Type Suitability
3.5.2. Changes in Pasture Macro-Types Composition
4. Discussion
5. Concluding Remarks
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Organisation for Economic Co-Operation and Development. Climate Change in the European Alps; OECD Publishing: Paris, France, 2007; ISBN 9789264031685. [Google Scholar]
- Sundseth, K. Natura 2000 in the Alpine Region; European Communities: Luxembourg, 2009; p. 15. [Google Scholar]
- Bengtsson, J.; Bullock, J.M.; Egoh, B.; Everson, C.; Everson, T.; O’Connor, T.; O’Farrell, P.J.; Smith, H.G.; Lindborg, R. Grasslands—More important for ecosystem services than you might think. Ecosphere 2019, 10, e02582. [Google Scholar] [CrossRef]
- The Economics of Ecosystems and Biodiversity. The Economics of Ecosystems and Biodiversity: Mainstreanubg the Economics of Nature: A Synthesis of the Approach, Conclusions and Recommendations of TEEB; Progress Press: Birkirkara, Malta, 2010; ISBN 9783981341034. [Google Scholar]
- Grêt-Regamey, A.; Brunner, S.H.; Kienast, F. Mountain Ecosystem Services: Who Cares? Mt. Res. Dev. 2012, 32, S23–S34. [Google Scholar] [CrossRef]
- Bernués, A.; Rodríguez-Ortega, T.; Ripoll-Bosch, R.; Alfnes, F. Socio-cultural and economic valuation of ecosystem services provided by Mediterranean mountain agroecosystems. PLoS ONE 2014, 9, e102479. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Millenium Ecosystem Assesment. Eccosystems and Human Well-Being: Synthesis; Island Press: Washington, DC, USA, 2005; ISBN 1-59726-040-1. [Google Scholar]
- Sturaro, E.; Cocca, G.; Gallo, L.; Mrad, M.; Ramanzin, M. Sistemi zootecnici e stili aziendali sulle Alpi Orientali Italiane: Indagine su un campione di allevamenti. Ital. J. Anim. Sci. 2009, 8, 541–554. [Google Scholar] [CrossRef]
- Mazzocchi, C.; Sali, G. Assessing the value of pastoral farming in the Alps using choice experiments: Evidence for public policies and management. J. Environ. Plan. Manag. 2019, 62, 552–567. [Google Scholar] [CrossRef]
- Argenti, G.; Lombardi, G. The pasture-type approach for mountain pasture description and management. Ital. J. Agron. 2012, 7, 293–299. [Google Scholar] [CrossRef] [Green Version]
- Baumont, R.; Prache, S.; Meuret, M.; Morand-Fehr, P. How forage characteristics influence behaviour and intake in small ruminants: A review. Livest. Prod. Sci. 2000, 64, 15–28. [Google Scholar] [CrossRef]
- Poore, M.; Scaglia, G.; Andrae, J.; Harmon, D.; Rogers, J.; Blacklin, S. Pasture-Finished Beef Production in the South; Elsevier: Amsterdam, The Netherlands, 2019; ISBN 9780128144756. [Google Scholar]
- Wilkinson, J.M.; Lee, M.R.F.; Rivero, M.J.; Chamberlain, A.T. Some challenges and opportunities for grazing dairy cows on temperate pastures. Grass Forage Sci. 2020, 75, 1–17. [Google Scholar] [CrossRef]
- Tardella, F.M.; Bricca, A.; Goia, I.G.; Catorci, A. How mowing restores montane Mediterranean grasslands following cessation of traditional livestock grazing. Agric. Ecosyst. Environ. 2020, 295, 106880. [Google Scholar] [CrossRef]
- Boscariol, G.P. La strategia per le aree interne quale strumento di sviluppo dei territori montani. Riv. Giuridica Mezzog. 2017, 3, 675–702. [Google Scholar]
- Hinojosa, L.; Lambin, E.F.; Mzoughi, N.; Napoléone, C. Place attachment as a factor of mountain farming permanence: A survey in the French Southern Alps. Ecol. Econ. 2016, 130, 308–315. [Google Scholar] [CrossRef]
- Hinojosa, L.; Napoléone, C.; Moulery, M.; Lambin, E.F. The “mountain effect” in the abandonment of grasslands: Insights from the French Southern Alps. Agric. Ecosyst. Environ. 2016, 221, 115–124. [Google Scholar] [CrossRef]
- Lamprecht, A.; Semenchuk, P.R.; Steinbauer, K.; Winkler, M.; Pauli, H. Climate change leads to accelerated transformation of high-elevation vegetation in the central Alps. New Phytol. 2018, 220, 447–459. [Google Scholar] [CrossRef] [PubMed]
- Gobiet, A.; Kotlarski, S.; Beniston, M.; Heinrich, G.; Rajczak, J.; Stoffel, M. 21st century climate change in the European Alps—A review. Sci. Total Environ. 2014, 493, 1138–1151. [Google Scholar] [CrossRef]
- Heinrich, G.; Gobiet, A.; Truhetz, H.; Mendlik, T. Expected Climate Change and Its Uncertainty in the Alpine Region Extended Uncertainty Assessment of the Reclip: Century and ENSEMBLES Multi-Model Dataset; Scientific Report No. 50-2013; Wegener Center for Climate and Global Change: Graz, Austria, 2013. [Google Scholar]
- Calanca, P. Climate change and drought occurrence in the Alpine region: How severe are becoming the extremes? Glob. Planet. Chang. 2007, 57, 151–160. [Google Scholar] [CrossRef]
- Deléglise, C.; Meisser, M.; Mosimann, E.; Spiegelberger, T.; Signarbieux, C.; Jeangros, B.; Buttler, A. Drought-induced shifts in plants traits, yields and nutritive value under realistic grazing and mowing managements in a mountain grassland. Agric. Ecosyst. Environ. 2015, 213, 94–104. [Google Scholar] [CrossRef]
- Hock, R.; Rasul, G.; Adler, C.; Cáceres, B.; Gruber, S.; Hirabayashi, Y.; Jackson, M.; Kääb, A.; Kang, S.; Kutuzov, S.; et al. Chapter 2: High Mountain Areas. In IPCC Special Report on the Ocean and Cryosphere in a Changing Climate; Pörtner, H.-O., Roberts, D.C., Masson-Delmotte, V., Zhai, P., Tignor, M., Poloczanska, E., Mintenbeck, K., Alegría, A., Nicolai, M., Okem, A., et al., Eds.; Intergovernmental Panel on Climate Change (IPCC): Geneva, Switzerland, 2019; pp. 1–83. [Google Scholar]
- Auer, I.; Böhm, R.; Jurkovic, A.; Lipa, W.; Orlik, A.; Potzmann, R.; Schöner, W.; Ungersböck, M.; Matulla, C.; Briffa, K.; et al. HISTALP—Historical instrumental climatological surface time series of the Greater Alpine Region. Int. J. Climatol. 2007, 27, 17–46. [Google Scholar] [CrossRef]
- Gavazov, K.S.; Peringer, A.; Buttler, A.; Gillet, F.; Spiegelberger, T. Dynamics of forage production in pasture-woodlands of the Swiss Jura mountains under projected climate change scenarios. Ecol. Soc. 2013, 18, 38. [Google Scholar] [CrossRef] [Green Version]
- Matteodo, M.; Wipf, S.; Stöckli, V.; Rixen, C.; Vittoz, P. Elevation gradient of successful plant traits for colonizing alpine summits under climate change. Environ. Res. Lett. 2013, 8, 024043. [Google Scholar] [CrossRef]
- Pauli, H.; Gottfried, M.; Dullinger, S.; Abdaladze, O.; Akhalkatsi, M.; Alonso, J.L.B.; Coldea, G.; Dick, J.; Erschbamer, B.; Calzado, R.F.; et al. Recent Plant Diversity Changes on Europe’s Mountain Summits. Science 2012, 336, 353–355. [Google Scholar] [CrossRef] [Green Version]
- Carlson, B.Z.; Corona, M.C.; Dentant, C.; Bonet, R.; Thuiller, W.; Choler, P. Observed long-term greening of alpine vegetation—A case study in the French Alps. Environ. Res. Lett. 2017, 12, 114006. [Google Scholar] [CrossRef]
- Schwager, P.; Berg, C. Global warming threatens conservation status of alpine EU habitat types in the European Eastern Alps. Reg. Environ. Chang. 2019, 19, 2411–2421. [Google Scholar] [CrossRef] [Green Version]
- Hoegh-Guldberg, O.; Jacob, D.; Taylor, M.; Bindi, M.; Brown, S.; Camilloni, I.; Diedhiou, A.; Djalante, R.; Ebi, K.L.; Engelbrecht, F.; et al. Chapter 3: Impacts of 1.5 °C global warming on natural and human systems. In Global Warming of 1.5 °C: An IPCC Special Report; Intergovernmental Panel on Climate Change (IPCC): Geneva, Switzerland, 2018. [Google Scholar]
- Dibari, C.; Bindi, M.; Moriondo, M.; Staglianò, N.; Targetti, S.; Argenti, G. Spatial data integration for the environmental characterization of pasture macrotypes in the Italian Alps. Grass Forage Sci. 2016, 71, 219–234. [Google Scholar] [CrossRef]
- Peel, M.C.; Finlayson, B.L.; McMahon, T.A. Updated world map of the Köppen-Geiger climate classification. Hydrol. Earth Syst. Sci. 2007, 11, 1633–1644. [Google Scholar] [CrossRef] [Green Version]
- Schär, C.; Davies, T.D.; Frei, C.; Wanner, H.; Widmann, M.; Wild, M.; Davies, H.C. Current Alpine Climate; The MIT Press: Cambridge, MA, USA, 1998. [Google Scholar]
- Beniston, M. Mountain weather and climate: A general overview and a focus on climatic change in the Alps. Hydrobiologia 2006, 562, 3–16. [Google Scholar] [CrossRef] [Green Version]
- Baruck, J.; Nestroy, O.; Sartori, G.; Baize, D.; Traidl, R.; Vrščaj, B.; Bräm, E.; Gruber, F.E.; Heinrich, K.; Geitner, C. Soil classification and mapping in the Alps: The current state and future challenges. Geoderma 2016, 264, 312–331. [Google Scholar] [CrossRef]
- Orlandi, S.; Probo, M.; Sitzia, T.; Trentanovi, G.; Garbarino, M.; Lombardi, G.; Lonati, M. Environmental and land use determinants of grassland patch diversity in the western and eastern Alps under agro-pastoral abandonment. Biodivers. Conserv. 2016, 25, 275–293. [Google Scholar] [CrossRef]
- Lüth, C.; Tasser, E.; Niedrist, G.; Via, J.D.; Tappeiner, U. Plant communities of mountain grasslands in a broad cross-section of the Eastern Alps. Flora Morphol. Distrib. Funct. Ecol. Plants 2011, 206, 433–443. [Google Scholar] [CrossRef]
- Argenti, G.; Bottai, L.; Chiesi, M.; Maselli, F.; Stagliano, N.; Targetti, S. Analysis and assessment of mountain pastures by integration of multispectral and ancillary data. Riv. Ital. Telerilevamento 2011, 43, 45–57. [Google Scholar] [CrossRef]
- Breiman, L. Random Forests. Mach. Learn. 2001, 45, 5–32. [Google Scholar] [CrossRef] [Green Version]
- Nachtergaele, F.; van Velthuizen, H.; Verelst, L. The Harmonized World Soil Database; Version 1.0.; FAO: Rome, Italy, 2008; pp. 34–37. [Google Scholar]
- Hijmans, R.J.; Cameron, S.E.; Parra, J.L.; Jones, P.G.; Jarvis, A. Very high resolution interpolated climate surfaces for global land areas. Int. J. Climatol. 2005, 25, 1965–1978. [Google Scholar] [CrossRef]
- Dibari, C.; Argenti, G.; Catolfi, F.; Moriondo, M.; Staglianò, N.; Bindi, M. Pastoral suitability driven by future climate change along the apennines. Ital. J. Agron. 2015, 10, 109–116. [Google Scholar] [CrossRef] [Green Version]
- Ruti, P.M.; Somot, S.; Giorgi, F.; Dubois, C.; Flaounas, E.; Obermann, A.; Dell’Aquila, A.; Pisacane, G.; Harzallah, A.; Lombardi, E.; et al. Med-CORDEX initiative for Mediterranean climate studies. Bull. Am. Meteorol. Soc. 2016, 97, 1187–1208. [Google Scholar] [CrossRef] [Green Version]
- Targetti, S.; Staglianò, N.; Messeri, A.; Argenti, G. A state-and-transition approach to alpine grasslands under abandonment. IForest 2010, 3, 44–51. [Google Scholar] [CrossRef] [Green Version]
- Cavallero, A.; Aceto, P.; Gorlier, A.; Lombardi, G.; Lonati, M.; Martinasso, B.; Tagliatori, C. I Tipi Pastorali delle Alpi Piemontesi; Alberto Perdisa Editore: Bologna, Italy, 2007. [Google Scholar]
- Istituto Superiore per la Protezione e la Ricerca Ambientale. Il Progetto Carta della Natura: Linee Guida per la Cartografia e la Valutazione degli Habitat alla Scala 1:50.000; ISPRA: Rome, Italy, 2009; ISBN 9788844803810.
- Bissardon, M.; Guibal, L. CORINE Biotopes: French Habitat Types [Typs d’Habitats Français]; École Nationale du Génie Rural, des Eaux et des Forêts: Nancy, France, 1997; p. 179. [Google Scholar]
- Pignatti, S. Flora d’Italia; Edagricole: Bologna, Italy, 1982; ISBN 8850652429. [Google Scholar]
- Raunkiaer, C. The Life Forms of Plants and Statistical Plant Geography; The Clarendon Press: Oxford, UK, 1934. [Google Scholar]
- Attorre, F.; Alfò, M.; De Sanctis, M.; Francesconi, F.; Valenti, R.; Vitale, M.; Bruno, F. Evaluating the effects of climate change on tree species abundance and distribution in the Italian peninsula. Appl. Veg. Sci. 2011, 14, 242–255. [Google Scholar] [CrossRef]
- Gaál, M.; Moriondo, M.; Bindi, M. Modelling the impact of climate change on the Hungarian wine regions using Random Forest. Appl. Ecol. Environ. Res. 2012, 10, 121–140. [Google Scholar] [CrossRef]
- Moriondo, M.; Jones, G.V.; Bois, B.; Dibari, C.; Ferrise, R.; Trombi, G.; Bindi, M. Projected shifts of wine regions in response to climate change. Clim. Chang. 2013, 119, 825–839. [Google Scholar] [CrossRef]
- Moriondo, M.; Trombi, G.; Ferrise, R.; Brandani, G.; Dibari, C.; Ammann, C.M.; Lippi, M.M.; Bindi, M. Olive trees as bio-indicators of climate evolution in the Mediterranean Basin. Glob. Ecol. Biogeogr. 2013, 22, 818–833. [Google Scholar] [CrossRef]
- Cutler, D.R.; Edwards, T.C.; Beard, K.H.; Cutler, A.; Kyle, T.; Gibson, J.; Lawler, J.J.; Beard, H.; Hess, T. Random Forests for Classification in Ecology Published by: Ecological Society of America. Ecology 2007, 88, 2783–2792. [Google Scholar] [CrossRef]
- Iverson, L.R.; Prasad, A.M.; Matthews, S.N.; Peters, M. Estimating potential habitat for 134 eastern US tree species under six climate scenarios. For. Ecol. Manag. 2008, 254, 390–406. [Google Scholar] [CrossRef]
- Dibari, C.; Argenti, G.; Moriondo, M.; Staglianò, N.; Targetti, S.; Bindi, M. Climate change impacts on distribution and composition of the Alpine Natural Pasturelands. In Proceedings of the 1st Annual Conference Climate Change and Its Implications on Ecosystem and Society, Lecce, Italy, 23–24 September 2013; pp. 578–586. [Google Scholar]
- Schroth, G.; Läderach, P.; Martinez-Valle, A.I.; Bunn, C.; Jassogne, L. Vulnerability to climate change of cocoa in West Africa: Patterns, opportunities and limits to adaptation. Sci. Total Environ. 2016, 556, 231–241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oshiro, T.M.; Perez, P.S.; Baranauskas, J.A. How Many Trees in a Random Forest? In Proceedings of the Lecture Notes in Computer Science; Perner, P., Ed.; Springer: Berlin/Heidelberg, Germany, 2012; pp. 154–168. [Google Scholar]
- Allouche, O.; Tsoar, A.; Kadmon, R. Assessing the accuracy of species distribution models: Prevalence, kappa and the true skill statistic (TSS). J. Appl. Ecol. 2006, 43, 1223–1232. [Google Scholar] [CrossRef]
- Harrison, M.T.; Cullen, B.R.; Rawnsley, R.P. Modelling the sensitivity of agricultural systems to climate change and extreme climatic events. Agric. Syst. 2016, 148, 135–148. [Google Scholar] [CrossRef]
- Thornton, P.K.; Ericksen, P.J.; Herrero, M.; Challinor, A.J. Climate variability and vulnerability to climate change: A review. Glob. Chang. Biol. 2014, 20, 3313–3328. [Google Scholar] [CrossRef] [PubMed]
- Scherrer, D.; Kãrner, C. Infra-red thermometry of alpine landscapes challenges climatic warming projections. Glob. Chang. Biol. 2010, 16, 2602–2613. [Google Scholar] [CrossRef]
- Choler, P. Winter soil temperature dependence of alpine plant distribution: Implications for anticipating vegetation changes under a warming climate. Perspect. Plant Ecol. Evol. Syst. 2018, 30, 6–15. [Google Scholar] [CrossRef]
- Cannone, N.; Sgorbati, S.; Guglielmin, M. Unexpected impacts of climate change on alpine vegetation. Front. Ecol. Environ. 2007, 7, 360–364. [Google Scholar] [CrossRef] [Green Version]
- Unterluggauer, P.; Mallaun, M.; Erschbamer, B. The higher the summit, the higher the diversity changes—Results of a long-term monitoring project in the Dolomites. Gredleriana 2016, 16, 5–34. [Google Scholar]
- Gehrig-Fasel, J.; Guisan, A.; Zimmermann, N.E. Tree line shifts in the Swiss Alps: Climate change or land abandonment? J. Veg. Sci. 2007, 18, 571–582. [Google Scholar] [CrossRef]
- Motta, R.; Nola, P. Growth trends and dynamics in sub-alpine forest stands in the Varaita Valley (Piedmont, Italy) and their relationships with human activities and global change. J. Veg. Sci. 2001, 12, 219–230. [Google Scholar] [CrossRef]
- Chauchard, S.; Beilhe, F.; Denis, N.; Carcaillet, C. An increase in the upper tree-limit of silver fir (Abies alba Mill.) in the Alps since the mid-20th century: A land-use change phenomenon. For. Ecol. Manag. 2010, 259, 1406–1415. [Google Scholar] [CrossRef]
- Rogora, M.; Frate, L.; Carranza, M.L.; Freppaz, M.; Stanisci, A.; Bertani, I.; Bottarin, R.; Brambilla, A.; Canullo, R.; Carbognani, M.; et al. Assessment of climate change effects on mountain ecosystems through a cross-site analysis in the Alps and Apennines. Sci. Total Environ. 2018, 624, 1429–1442. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klos, P.Z.; Link, T.E.; Abatzoglou, J.T. Extent of the rain-snow transition zone in the western U.S. under historic and projected climate. Geophys. Res. Lett. 2014, 41, 4560–4568. [Google Scholar] [CrossRef]
- Petersky, R.S.; Shoemaker, K.T.; Weisberg, P.J.; Harpold, A.A. The sensitivity of snow ephemerality to warming climate across an arid to montane vegetation gradient. Ecohydrology 2019, 12, e2060. [Google Scholar] [CrossRef]
- Ziliotto, U.; Andrich, O.; Lasen, C.; Ramanzin, M. Tratti Essenziali della Tipologia Veneta dei Pascoli di Monte e Dintorni; University of Padua: Padua, Italy, 2004; p. 253. [Google Scholar]
- Güsewell, S.; Jewell, P.L.; Edwards, P.J. Effects of heterogeneous habitat use by cattle on nutrient availability and litter decomposition in soils of an Alpine pasture. Plant Soil 2005, 268, 135–149. [Google Scholar] [CrossRef]
- Frate, L.; Carranza, M.L.; Evangelista, A.; Stinca, A.; Schaminée, J.H.J.; Stanisci, A. Climate and land use change impacts on mediterranean high-mountain vegetation in the apennines since the 1950s. Plant Ecol. Divers. 2018, 11, 85–96. [Google Scholar] [CrossRef]
- Liu, Q.; Fu, Y.H.; Zhu, Z.; Liu, Y.; Liu, Z.; Huang, M.; Janssens, I.A.; Piao, S. Delayed autumn phenology in the Northern Hemisphere is related to change in both climate and spring phenology. Glob. Chang. Biol. 2016, 22, 3702–3711. [Google Scholar] [CrossRef]
- Xu, C.; Liu, H.; Williams, A.P.; Yin, Y.; Wu, X. Trends toward an earlier peak of the growing season in Northern Hemisphere mid-latitudes. Glob. Chang. Biol. 2016, 22, 2852–2860. [Google Scholar] [CrossRef]
- Park, T.; Chen, C.; Macias-Fauria, M.; Tømmervik, H.; Choi, S.; Winkler, A.; Bhatt, U.S.; Walker, D.A.; Piao, S.; Brovkin, V.; et al. Changes in timing of seasonal peak photosynthetic activity in northern ecosystems. Glob. Chang. Biol. 2019, 25, 2382–2395. [Google Scholar] [CrossRef] [Green Version]
- Buitenwerf, R.; Rose, L.; Higgins, S.I. Three decades of multi-dimensional change in global leaf phenology. Nat. Clim. Chang. 2015, 5, 364–368. [Google Scholar] [CrossRef]
- Gonsamo, A.; Chen, J.M.; Ooi, Y.W. Peak season plant activity shift towards spring is reflected by increasing carbon uptake by extratropical ecosystems. Glob. Chang. Biol. 2018, 24, 2117–2128. [Google Scholar] [CrossRef] [PubMed]
- Pittarello, M.; Probo, M.; Perotti, E.; Lonati, M.; Lombardi, G.; Ravetto Enri, S. Grazing Management Plans improve pasture selection by cattle and forage quality in sub-alpine and alpine grasslands. J. Mt. Sci. 2019, 16, 2126–2135. [Google Scholar] [CrossRef]
- Celaya, R.; Oliván, M.; Ferreira, L.M.M.; Martínez, A.; García, U.; Osoro, K. Comparison of grazing behaviour, dietary overlap and performance in non-lactating domestic ruminants grazing on marginal heathland areas. Livest. Sci. 2007, 106, 271–281. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Legend | Number of Pixels |
|---|---|
| SP: pastures encroached by shrub species | 4101 |
| CC: pastures dominated by Carex curvula | 2661 |
| CF: pastures dominated by Carex firma | 1185 |
| NS: pastures dominated by Nardus stricta | 5030 |
| FR: pastures dominated by Festuca gr. rubra | 503 |
| SV: pastures dominated by Sesleria varia | 3560 |
| XS: pastures dominated by xeric species | 1560 |
| Un-classified pastures | 8866 |
| Other land use | 29,657 |
| Simulated | RF Accuracy (%) | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| SP | CC | CF | NS | FR | XS | SV | Observed | |||
| Observed | SP | 1531 | 8 | 1 | 16 | 0 | 4 | 9 | 1569 | 97.6 |
| CC | 2 | 1651 | 0 | 6 | 0 | 0 | 5 | 1664 | 99.2 | |
| CF | 6 | 0 | 508 | 3 | 0 | 1 | 5 | 523 | 97.1 | |
| NS | 3 | 3 | 0 | 3516 | 0 | 5 | 2 | 3529 | 99.6 | |
| FR | 3 | 1 | 0 | 6 | 211 | 0 | 3 | 224 | 94.2 | |
| XS | 0 | 1 | 0 | 19 | 0 | 1070 | 1 | 1091 | 98.1 | |
| SV | 0 | 4 | 6 | 13 | 1 | 5 | 2253 | 2282 | 98.7 | |
| Simulated | 1545 | 1668 | 515 | 3579 | 212 | 1085 | 2278 | 98.7 | ||
| Number of Pixels | % | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| RCM | Present | RCP 4.5 | RCP 8.5 | RCP 4.5 | RCP 8.5 | ||||
| - - - | - - - | 2011–2040 | 2041–2070 | 2011–2040 | 2041–2070 | 2011–2040 | 2041–2070 | 2011–2040 | 2041–2070 |
| Aladin | 20,874 | 19,557 | 16,704 | 18,931 | 16,712 | −6% | −20% | −9% | −20% |
| CMCC | 20,874 | 13,130 | 11,225 | 15,037 | 11,289 | −37% | −46% | −28% | −46% |
| ICTP | 20,874 | 15,507 | 93,92 | 16,497 | 12,548 | −26% | −55% | −21% | −40% |
| Ensemble | 20,874 | 16,065 | 12,440 | 16,822 | 13,516 | −23% | −40% | −19% | −35% |
| Scenario | RCP 4.5 | RCP 8.5 | RCP 4.5 | RCP 8.5 | RCP Ensemble | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| RCM | Present | 2011–2040 | 2041–2070 | 2011–2040 | 2041–2070 | 2011–2040 | 2041–2070 | 2011–2040 | 2041–2070 | 2011–2040 | 2041–2070 | |
| - - - | - - - | Nº of Pixels | % | % | ||||||||
| Shrub species (SP) | Ensemble | 2332 | 1168 | 498 | 1490 | 789 | −50 | −79 | −36 | −66 | −43 | −72.5 |
| Carex curvula (CC) | 3026 | 988 | 748 | 1188 | 790 | −67 | −75 | −61 | −74 | −64 | −74.5 | |
| Carex firma (CF) | 617 | 120 | 7 | 150 | 2 | −81 | −99 | −76 | −100 | −78.5 | −99.5 | |
| Nardus stricta (NS) | 9303 | 9183 | 7355 | 10058 | 7842 | −1 | −21 | 8 | −16 | 3.5 | −18.5 | |
| Festuca rubra (FR) | 256 | 104 | 5 | 36 | 18 | −59 | −98 | −86 | −93 | −72.5 | −95.5 | |
| Xeric species (XS) | 2219 | 2217 | 2499 | 2123 | 2367 | 0 | 13 | −4 | 7 | −2 | 10 | |
| Sesleria varia (SV) | 3121 | 2284 | 1329 | 1777 | 1708 | −27 | −57 | −43 | −45 | −35 | −51 | |
| Macro-Types | RCM | RCP45 | RCP85 | |||
|---|---|---|---|---|---|---|
| - - - | Present | 2011–2040 | 2041–2070 | 2011–2040 | 2041–2070 | |
| Shrub species (SP) | Ensemble | 11% | 7% (−4%) | 4% (−7%) | 9% (−2%) | 6% (−5%) |
| Carex curvula (CC) | 14% | 6% (−8%) | 6% (−8%) | 7% (−7%) | 6% (−8%) | |
| Carex firma (CF) | 3% | 1% (−2%) | 0% (−3%) | 1% (−2%) | 0% (−3%) | |
| Nardus stricta (NS) | 45% | 57% (+12%) | 59% (+14%) | 60% (+15%) | 58% (+13%) | |
| Festuca rubra (FR) | 1% | 1% (0%) | 0% (−1%) | 0% (−1%) | 0% (−1%) | |
| Xeric species (XS) | 11% | 14% (+3%) | 20% (+9%) | 13% (+2%) | 18% (+7%) | |
| Sesleria varia (SV) | 15% | 14% (−1%) | 11% (−4%) | 11% (−4%) | 13% (−2%) | |
| Total | 100% | 100% | 100% | 100% | 100% | |
| Time-Slice | SP | CC | CF | NS | FR | XS | SV | Total | |
|---|---|---|---|---|---|---|---|---|---|
| Occurrence | |||||||||
| Shrub species (SP) | 2011–2040 | 31.7% | 9.0% | 9.5% | 29.2% | 1.6% | 1.3% | 17.9% | 100% |
| Carex curvula (CC) | 19.1% | 42.6% | 10.0% | 13.1% | 0.5% | 0.4% | 14.5% | 100% | |
| Carex firma (CF) | 2.8% | 6.2% | 39.1% | 12.1% | 3.5% | 0.0% | 36.5% | 100% | |
| Nardus stricta (NS) | 14.3% | 12.4% | 3.5% | 56.8% | 1.5% | 2.1% | 9.6% | 100% | |
| Festuca rubra (FR) | 2.3% | 7.5% | 0.0% | 50.9% | 13.0% | 21.8% | 4.6% | 100% | |
| Xeric species (XS) | 4.0% | 5.1% | 0.2% | 59.1% | 1.1% | 27.5% | 3.0% | 100% | |
| Sesleria varia (SV) | 10.1% | 19.9% | 5.1% | 21.7% | 1.8% | 1.5% | 40.2% | 100% | |
| Shrub species (SP) | 2041–2070 | 17.1% | 18.1% | 17.2% | 18.7% | 1.7% | 0.8% | 26.6% | 100% |
| Carex curvula (CC) | 20.2% | 44.5% | 6.0% | 12.6% | 0.7% | 0.2% | 16.1% | 100% | |
| Carex firma (CF) | 3.2% | 3.2% | 78.2% | 0.0% | 0.0% | 0.0% | 15.7% | 100% | |
| Nardus stricta (NS) | 15.9% | 16.5% | 6.6% | 42.3% | 1.8% | 1.8% | 15.3% | 100% | |
| Festuca rubra (FR) | 6.0% | 7.2% | 0.0% | 57.0% | 2.4% | 16.1% | 11.5% | 100% | |
| Xeric species (XS) | 6.7% | 11.9% | 0.6% | 58.4% | 1.0% | 14.9% | 6.6% | 100% | |
| Sesleria varia (SV) | 11.0% | 25.2% | 2.3% | 18.5% | 0.8% | 1.3% | 41.2% | 100% |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dibari, C.; Costafreda-Aumedes, S.; Argenti, G.; Bindi, M.; Carotenuto, F.; Moriondo, M.; Padovan, G.; Pardini, A.; Staglianò, N.; Vagnoli, C.; et al. Expected Changes to Alpine Pastures in Extent and Composition under Future Climate Conditions. Agronomy 2020, 10, 926. https://doi.org/10.3390/agronomy10070926
Dibari C, Costafreda-Aumedes S, Argenti G, Bindi M, Carotenuto F, Moriondo M, Padovan G, Pardini A, Staglianò N, Vagnoli C, et al. Expected Changes to Alpine Pastures in Extent and Composition under Future Climate Conditions. Agronomy. 2020; 10(7):926. https://doi.org/10.3390/agronomy10070926
Chicago/Turabian StyleDibari, Camilla, Sergi Costafreda-Aumedes, Giovanni Argenti, Marco Bindi, Federico Carotenuto, Marco Moriondo, Gloria Padovan, Andrea Pardini, Nicolina Staglianò, Carolina Vagnoli, and et al. 2020. "Expected Changes to Alpine Pastures in Extent and Composition under Future Climate Conditions" Agronomy 10, no. 7: 926. https://doi.org/10.3390/agronomy10070926