A Comparative Study of Organic and Conventional Management on the Rhizosphere Microbiome, Growth and Grain Quality Traits of Tritordeum

 , ,
, ,  ,
,  ,
,
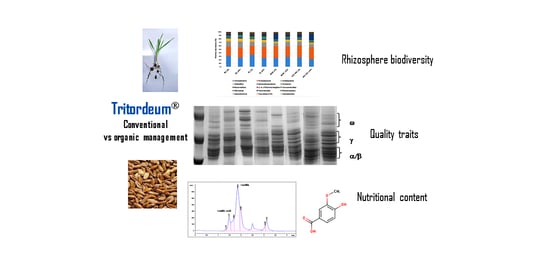
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Field Trial Set-up and Agronomic Parameters
2.2. Rhizosphere Soil Sampling and Root Growth Analyses
2.3. DNA Extraction from Rhizosphere Soil, Rhizobiome Identification by 16S rRNA Gene Sequencing, and Data Analysis
2.4. Determination of Micro- and Macronutrients in the Grain
2.5. Determination of Free Phenolic Acids
2.6. SDS-PAGE of Gluten Proteins, Separation and Densitometric Analysis of Gliadins, High Molecular Weight Glutenin Subunits (HMW-GS), and Low Molecular Weight Glutenin Subunits (LMW-GS)
2.7. Bioinformatics and Statistical Analysis
3. Results and Discussion
3.1. Root Growth and Rhizosphere Microbial Biodiversity
3.2. Grain Yield, and Nutrient and Phenolic Acid Contents
3.3. Grain Protein Content and Composition
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- FAOSTAT. Available online: http://www.fao.org/faostat/en/#home (accessed on 4 May 2020).
- Shewry, P.R.; Hey, S.J. The contribution of wheat to human diet and health. Food Energy Secur. 2015, 4, 178–202. [Google Scholar] [CrossRef] [PubMed]
- Kotschi, J. Agricultural biodiversity is essential for adapting to climate change. Ecol. Persp. Sci. Soc. 2007, 16, 98–101. [Google Scholar] [CrossRef]
- FAO. The Second Report on the State of the World’s Plant Genetic Resources for Food and Agriculture; FAO: Rome, Italy, 2010. [Google Scholar]
- Döring, T.F.; Knapp, S.; Kovacs, G.; Murphy, K.; Wolfe, M.S. Evolutionary plant breeding in cereals-into a new era. Sustainability 2011, 3, 1944–1971. [Google Scholar] [CrossRef] [Green Version]
- Tadesse, W.; Sanchez-Garcia, M.; Assefa, S.G.; Amri, A.; Bishaw, Z.; Ogbonnaya, F.C.; Baum, M. Genetic gains in wheat breeding and its role in feeding the world. Crop Breed. Gen. Genom. 2019, 1, e190005. [Google Scholar]
- Martín, A.; Sanchez-Monge Laguna, E.S.M. A hybrid between Hordeum chilense and Triticum turgidum. Cereal Res. Commun. 1980, 8, 349–353. [Google Scholar]
- Martín, A.; Sanchez-Monge Laguna, E. Cytology and morphology of the amphiploid Hordeum chilense × Triticum turgidum conv. Durum. Euphytica 1982, 31, 261–267. [Google Scholar] [CrossRef]
- Martín, A.; Alvarez, J.B.; Martín, L.M.; Barro, F.; Ballesteros, J. The development of Tritordeum: A novel cereal for food processing. J. Cereal Sci. 1999, 30, 85–95. [Google Scholar] [CrossRef]
- Agrasys. Available online: http://www.tritordeum.com (accessed on 25 January 2020).
- Villegas, A.; Casadesù, J.; Atienza, S.G.; Martos, V.; Maalouf, F.; Karam, F.; Aranjuelo, I.; Nogués, S. Tritordeum, wheat and triticale yield components under multi-local Mediterranean drought conditions. Field Crop. Res. 2010, 116, 68–74. [Google Scholar] [CrossRef] [Green Version]
- Atienza, S.G.; Ballesteros, J.; Martín, A.; Hornero-Méndez, D. Genetic variability of carotenoid concentration and degree of esterification among Tritordeum (×Tritordeum Ascherson et Graebner) and durum wheat accessions. J. Agric. Food Chem. 2007, 55, 4244–4251. [Google Scholar] [CrossRef]
- Pérez-Gálvez, A.; Mínguez-Mosquera, M.I. Esterification of xanthophylls and its effect on chemical behavior and bioavailability of carotenoids in the human. Nutr. Res. 2005, 25, 631–640. [Google Scholar] [CrossRef]
- Giordano, D.; Reyneri, A.; Locatelli, M.; Coïsson, J.D.; Blandino, M. Distribution of bioactive compounds in pearled fractions of Tritordeum. Food Chem. 2019, 301, 125228. [Google Scholar] [CrossRef]
- Erlandsson, A. Tritordeum. Evaluation of a New Food Cereal. Master’s Thesis, Swedish University of Agricultural Sciences, Uppsala, Sweden, 2010. [Google Scholar]
- Hack, H.; Bleiholder, H.; Buhr, L.; Meier, U.; Schnock-Fricke, U.; Weber, E.; Witzenberger, A. Einheitliche Codierung der phanologischen Entwicklungsstadien mono- und dikotyler Pflanzen—Erweiterte BBCH-Skala, Allgemein. Nachrichtenbl. Deut. Pflanzenschutzd. 1992, 44, 265–270. [Google Scholar]
- Pornaro, C.; Macolino, S.; Menegon, A.; Richardson, M. WinRHIZO technology for measuring morphological traits of bermudagrass stolons. Agron. J. 2017, 109, 3007–3010. [Google Scholar] [CrossRef] [Green Version]
- Milani, C.; Hevia, A.; Foroni, E.; Duranti, S.; Turroni, F.; Lugli, G.A.; Sanchez, B.; Martín, R.; Gueimonde, M.; van Sinderen, D.; et al. Assessing the fecal microbiota: An optimized ion torrent 16S rRNA gene-based analysis protocol. PLoS ONE 2013, 8, e68739. [Google Scholar] [CrossRef]
- Visioli, G.; Sanangelantoni, A.M.; Vamerali, T.; Dal Cortivo, C.; Blandino, M. 16S rDNA profiling to reveal the influence of seed-applied biostimulants on the rhizosphere of young maize plants. Molecules 2018, 23, 1461. [Google Scholar] [CrossRef] [Green Version]
- USEPA, EPA Method 3052. Microwave assisted acid digestion of siliceous and organically based matrices. In Test Methods for Evaluating Solid Waste, 3rd ed.; USEPA: Washington, DC, USA, 1995. [Google Scholar]
- Adom, K.K.; Sorrells, M.E.; Liu, R.H. Phytochemical profiles and antioxidant activity of wheat varieties. J. Agric. Food Chem. 2003, 51, 7825–7834. [Google Scholar] [CrossRef]
- Dal Cortivo, C.; Conselvan, G.B.; Carletti, P.; Barion, G.; Sella, L.; Vamerali, T. Biostimulant effects of seed-applied Sedaxane fungicide: Morphological and physiological changes in maize seedlings. Front. Plant Sci. 2017, 8, 2072. [Google Scholar] [CrossRef] [Green Version]
- Visioli, G.; Bonas, U.; Dal Cortivo, C.; Pasini, G.; Marmiroli, N.; Mosca, G.; Vamerali, T. Variations in yield and gluten proteins in durum wheat varieties under late-season foliar versus soil application of nitrogense fertilizer in a northern Mediterranean environment. J. Sci. Food Agric. 2018, 98, 2360–2369. [Google Scholar] [CrossRef] [Green Version]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Peña, A.G.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef] [Green Version]
- Edgar, R.C. Search and clustering orders of magnitude faster than BLAST. Bioinformatics 2010, 26, 2460–2461. [Google Scholar] [CrossRef] [Green Version]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA ribosomal RNA gene database project: Improved data processing and web-based tools. Nucleic Acids Res. 2013, 41, D590–D596. [Google Scholar] [CrossRef]
- Bulgarelli, D.; Garrido-Oter, R.; Münch, P.C.; Weiman, A.; Dröge, J.; Pan, Y.; McHardy, A.C.; Schulze-Lefert, P. Structure and function of the bacterial root microbiota in wild and domesticated barley. Cell Host Microbe 2015, 17, 392–403. [Google Scholar] [CrossRef] [Green Version]
- Li, F.; Chen, L.; Zhang, J.; Yin, J.; Huang, S. Bacterial community structure after long-term organic and inorganic fertilization reveals important associations between soil nutrients and specific taxa involved in nutrient transformations. Front. Microbiol. 2017, 8, 187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kolton, M.; Green, S.J.; Harel, Y.M.; Sela, N.; Elad, Y.; Cytryn, E. Draft genome sequence of Flavobacterium sp. Strain F52, isolated from the rhizosphere of bell pepper (Capsicum annuum L. cv. Maccabi). J. Bacteriol. 2012, 194, 5462–5463. [Google Scholar] [CrossRef] [Green Version]
- Wolińska, A.; Kuźniar, A.; Zielenkiewicz, U.; Izak, D.; Szafranek-Nakonieczna, A.; Banach, A.; Błaszczyk, M. Bacteroidetes as a sensitive biological indicator of agricultural soil usage revealed by a culture-independent approach. Appl. Soil Ecol. 2017, 119, 128–137. [Google Scholar] [CrossRef]
- Kanelli, M.; Mandic, M.; Kalakona, M.; Vasilakos, S.; Kekos, D.; Nikodinovic-Runic, J.; Topakas, E. Microbial production of violacein and process optimization for dyeing polyamide fabrics with acquired antimicrobial properties. Front. Microbiol. 2018, 9, 1495. [Google Scholar] [CrossRef] [Green Version]
- Pérez-Jaramillo, J.E.; Carrión, V.J.; de Hollander, M.; Raaijmakers, J.M. The wild side of plant microbiomes. Microbiome 2018, 6, 143. [Google Scholar] [CrossRef] [Green Version]
- Barro, F.; González-Fontes, A.; Maldonado, J.M. Nitrate uptake and reduction by durum wheat (Triticum turgidum) and Tritordeum (Hordeum chilense × Triticum turgidum). J. Plant Physiol. 1994, 143, 313–317. [Google Scholar] [CrossRef]
- Barro, F.; Fontes, A.G.; Maldonado, J.M. Organic nitrogen content and nitrate and nitrite reductase activities in Tritordeum and wheat grown under nitrate or ammonium. Plant Soil 1991, 135, 251–256. [Google Scholar] [CrossRef]
- Baruah, R. Towards the bioavailability of zinc in agricultural soils. In Role of Rhizospheric Microbes in Soil; Meena, V., Ed.; Springer: Singapore, 2018. [Google Scholar]
- Ozcan, T.; Akpinar-Bayizit, A.; Yilmaz-Ersan, L.; Delikanli, B. Phenolics in human health. Int. J. Chem. Eng. Appl. 2014, 5, 393–396. [Google Scholar] [CrossRef] [Green Version]
- Fernandez-Orozco, R.; Li, L.; Harflett, C.; Shewry, P.R.; Ward, J.L. Effects of environment and genotype on phenolic acids in wheat in the HEALTHGRAIN diversity screen. J. Agric. Food Chem. 2010, 58, 9341–9352. [Google Scholar] [CrossRef]
- Li, L.; Shewry, P.R.; Ward, J.L. Phenolic acids in wheat varieties in the HEALTHGRAIN Diversity Screen. J. Agric. Food Chem. 2008, 56, 9732–9739. [Google Scholar] [CrossRef]
- Navas-Lopez, J.F.; Ostos-Garrido, F.J.; Castillo, A.; Martíìn, A.; Gimenez, M.J.; Piston, F. Phenolic content variability and its chromosome location in Tritordeum. Front. Plant Sci. 2014, 5, 10. [Google Scholar] [CrossRef] [Green Version]
- Acreche, M.M.; Slafer, G.A. Variation of grain nitrogen content in relation with grain yield in old and modern Spanish wheats grown under a wide range of agronomic conditions in a mediterranean region. J. Agric. Sci. 2009, 147, 657–667. [Google Scholar] [CrossRef] [Green Version]
- Vaquero, L.; Comino, I.; Vivas, S.; Rodríguez-Martín, L.; Giménez, M.J.; Pastor, J.; Sousa, C.; Barro, F. Tritordeum: A novel cereal for food processing with good acceptability and significant reduction in gluten immunogenic peptides in comparison with wheat. J. Sci. Food Agric. 2018, 98, 2201–2209. [Google Scholar] [CrossRef] [PubMed]
- Altenbach, S.B.; Kothari, K.M.; Lieu, D. Environmental conditions during wheat grain development alter temporal regulation of major gluten protein genes. Cereal Chem. 2002, 79, 279–285. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Farming System | Cultivar | Root Parameters | ||||
|---|---|---|---|---|---|---|
| Length (cm Plant−1) | Surface Area (cm2 Plant−1) | Average Diameter (µm) | Tips (n Plant−1) | Branching Index (n Forks cm−1) | ||
| Organic | Iride | 72.9 ± 8.036 a | 11.9 ± 1.854 a | 514 ±24.17 b | 97.6 ± 27.08 a | 1.76 ± 0.385 a |
| Bulel | 38.5 ± 0.651 b | 6.90 ± 0.211 ab | 569 ± 14.17 a | 30.0 ± 2.835 b | 0.77 ± 0.029 b | |
| HTC-444 | 29.2 ± 2.554 b | 4.74 ± 0.158 b | 523 ± 34.48 ab | 26.1 ± 2.570 b | 1.10 ± 0.105 ab | |
| Conventional | Iride | 86.8 ± 33.55 a | 12.1 ± 5.017 a | 432 ± 9.948 c | 103 ± 45.12 a | 1.28 ± 0.287 a |
| Bulel | 32.3 ± 2.657 b | 5.05 ± 0.584 b | 495 ± 17.77 b | 25.1 ± 4.636 b | 1.11 ± 0.203 a | |
| HTC-444 | 39.3 ± 5.851 b | 7.30 ± 0.995 b | 592 ± 7.219 a | 36.7 ± 12.54 b | 1.32 ± 0.228 a | |
| Farming system | ns | ns | ns | ns | ns | |
| Cultivar | * | * | ** | * | ns | |
| Farming system × Cultivar | ns | ns | ** | ns | ns | |
| Farming System | Cultivar | Plant Height (cm) | Culm Lodging (%) | TKW (g) | TW (g L−1) | Yield (t ha−1 at 14% Humidity) |
|---|---|---|---|---|---|---|
| Organic | Iride | 73.9 ± 2.1 a | 8.33 ± 1.67 a | 48.6 ± 0.59 a | 81.7 ± 0.95 a | 2.47 ± 0.16 a |
| Bulel | 65.8 ± 0.9 b | 1.67 ± 1.67 a | 35.9 ± 0.58 b | 75.0 ± 0.41 b | 1.02 ± 0.09 b | |
| HTC-444 | 68.4 ± 3.5 ab | 8.33 ± 6.01 a | 37.4 ± 0.06 b | 72.8 ± 0.79 b | 0.86 ± 0.08 b | |
| Conventional | Iride | 78.6 ± 0.3 b | 1.67 ± 1.67 b | 53.1 ± 0.91 a | 81.9 ± 0.54 a | 5.35 ± 0.02 a |
| Bulel | 80.2 ± 0.5 a | 3.33 ± 1.67 b | 41.7 ± 0.23 b | 77.9 ± 0.27 b | 3.33 ± 0.10 b | |
| HTC-444 | 82.4 ± 1.3 a | 13.3 ± 1.67 a | 40.3 ± 0.25 b | 75.3 ± 0.32 c | 2.99 ± 0.12 c | |
| Farming System | *** | ns | *** | ** | *** | |
| Cultivar | ns | * | *** | *** | *** | |
| Farming System × Cultivar | * | ns | ns | ns | ** | |
| Farming System | Cultivar | Element | |||||||
|---|---|---|---|---|---|---|---|---|---|
| N (%) | Ca (mg kg−1) | Fe (mg kg−1) | K (mg kg−1) | Mg (mg kg−1) | P (mg kg−1) | S (mg kg−1) | Zn (mg kg−1) | ||
| Organic | Iride | 2.10 ± 0.02 c | 326 ± 5 d | 39.7 ± 2.3 c | 5107 ± 208 bc | 1353 ± 80 c | 5381 ± 246 abc | 1438 ± 34 b | 55.8 ± 5.3 bc |
| Bulel | 2.32 ± 0.03 b | 406 ± 32 bc | 47.0 ± 3.3 bc | 6386 ± 429 a | 1626 ± 88 ab | 5664 ± 343 ab | 1899 ± 111 a | 58.1 ± 5.2 b | |
| HTC-444 | 2.65 ± 0.12 a | 452 ± 10 ab | 57.5 ± 3.7 ab | 5845 ± 81 ab | 1699 ± 34 a | 6053 ± 95 a | 1978 ± 50 a | 72.1 ± 3.2 a | |
| Conventional | Iride | 2.27 ± 0.05 a | 354 ± 22 cd | 43.2 ± 3.5 c | 4721 ± 267 c | 1351 ± 96 c | 5161 ± 335 bc | 1572 ± 80 b | 44.2 ± 4.6 cd |
| Bulel | 2.26 ± 0.01 a | 367 ± 24 cd | 46.2 ± 2.6 c | 5240 ± 328 bc | 1420 ± 93 bc | 4633 ± 285 c | 1873 ± 123 a | 41.8 ± 2.3 d | |
| HTC-444 | 2.31 ± 0.05 a | 506 ± 36 a | 64.63 ± 4.8 a | 6028 ± 495 ab | 1702 ± 93 a | 5823 ± 325 ab | 2047 ± 112 a | 56.6 ± 3.7 bc | |
| Farming System | ns | ns | ns | ns | ns | ns | ns | ** | |
| Cultivar | ** | *** | *** | * | ** | * | *** | ** | |
| Farming System × Cultivar | ** | ns | ns | ns | n | ns | ns | ns | |
| Farming System | Cultivar | Phenolic Acid (µg g−1 d.w.) | |||||
|---|---|---|---|---|---|---|---|
| Vanillic | Caffeic | Syringic | p-Coumaric | t-Ferulic | Total Amount | ||
| Organic | Iride | 41.5 ± 20.0 b | 9.22 ± 2.11 b | 21.7 ± 3.4 a | 2.71 ± 1.32 a | 20.5 ± 4.8 a | 95.8 ± 15 b |
| Bulel | 46.1 ± 16.1 b | 13.22 ± 5.1 ab | 6.43 ± 2.59 c | 1.84 ± 0.82 a | 41.2 ± 26.58 a | 108 ± 47 ab | |
| HTC-444 | 97.1 ± 51.8 ab | 15.55 ± 3.8 ab | 10.2 ± 2.7 bc | 2.04 ± 0.18 a | 10.8 ± 1.03 a | 135 ± 50 ab | |
| Conventional | Iride | 147 ± 38.5 a | 7.46 ± 1.65 b | 20.1 ± 3.2 a | 2.07 ± 1.36 a | 10.9 ± 0.03 a | 188 ± 43 ab |
| Bulel | 140 ± 9.7 a | 15.96 ± 2.0 ab | 10.3 ± 2.9 bc | 1.83 ± 0.22 a | 10.3 ± 0.43 a | 179 ± 13 ab | |
| HTC-444 | 166 ± 14.6 a | 20.96 ± 3.2 a | 16.5 ± 1.0 ab | 1.99 ± 0.28 a | 9.75 ± 0.26 a | 215 ± 15 a | |
| Farming system | ** | ns | ns | ns | ns | * | |
| Cultivar | ns | * | ** | ns | ns | ns | |
| Farming system × Cultivar | ns | ns | ns | ns | ns | ns | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Visioli, G.; Lauro, M.; Vamerali, T.; Dal Cortivo, C.; Panozzo, A.; Folloni, S.; Piazza, C.; Ranieri, R. A Comparative Study of Organic and Conventional Management on the Rhizosphere Microbiome, Growth and Grain Quality Traits of Tritordeum. Agronomy 2020, 10, 1717. https://doi.org/10.3390/agronomy10111717
Visioli G, Lauro M, Vamerali T, Dal Cortivo C, Panozzo A, Folloni S, Piazza C, Ranieri R. A Comparative Study of Organic and Conventional Management on the Rhizosphere Microbiome, Growth and Grain Quality Traits of Tritordeum. Agronomy. 2020; 10(11):1717. https://doi.org/10.3390/agronomy10111717
Chicago/Turabian StyleVisioli, Giovanna, Marta Lauro, Teofilo Vamerali, Cristian Dal Cortivo, Anna Panozzo, Silvia Folloni, Cristina Piazza, and Roberto Ranieri. 2020. "A Comparative Study of Organic and Conventional Management on the Rhizosphere Microbiome, Growth and Grain Quality Traits of Tritordeum" Agronomy 10, no. 11: 1717. https://doi.org/10.3390/agronomy10111717