Construction of Efficient Platform Escherichia coli Strains for Polyhydroxyalkanoate Production by Engineering Branched Pathway

 ,
,  ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains, Media, Reagents, and Culture Conditions
2.2. Genome Editing
2.3. Analytical Methods
2.4. CoA Analysis Using Liquid Chromatography (LC)–Mass Spectrometry (MS)
3. Results
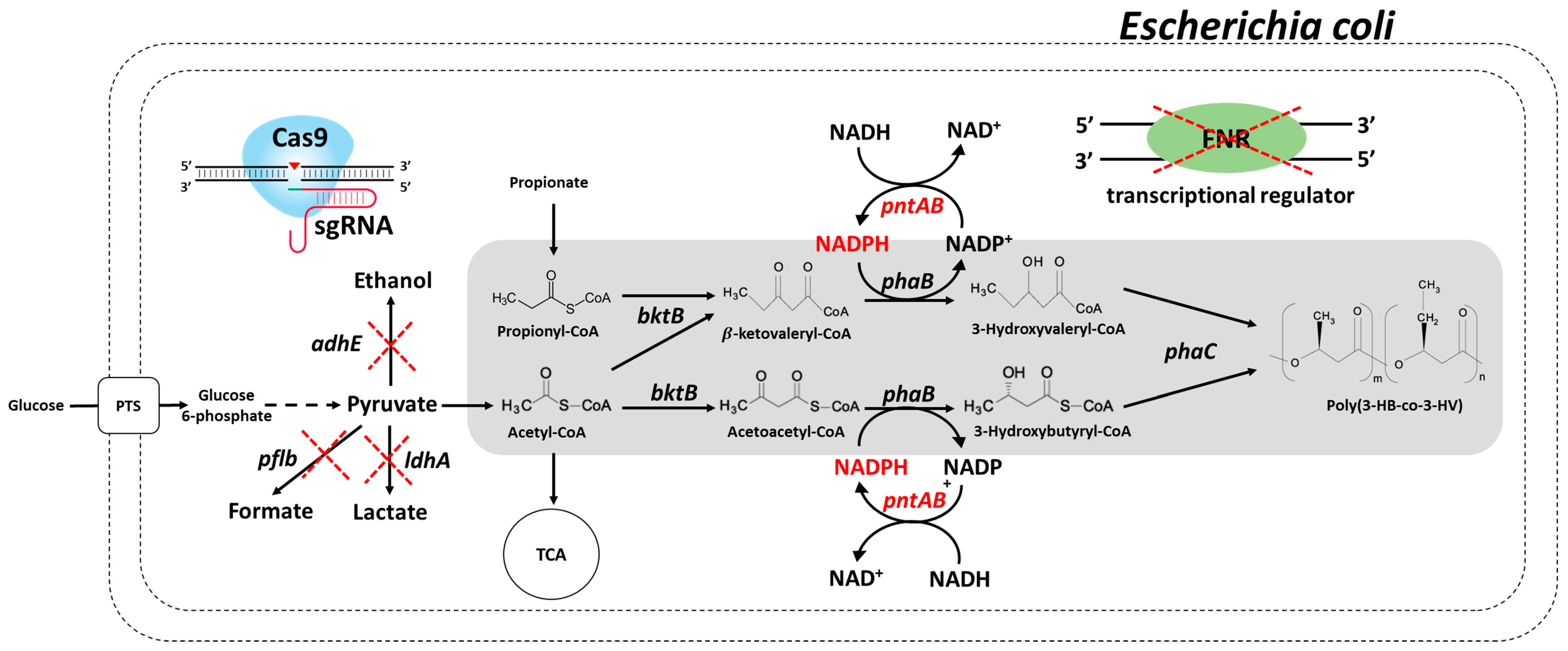
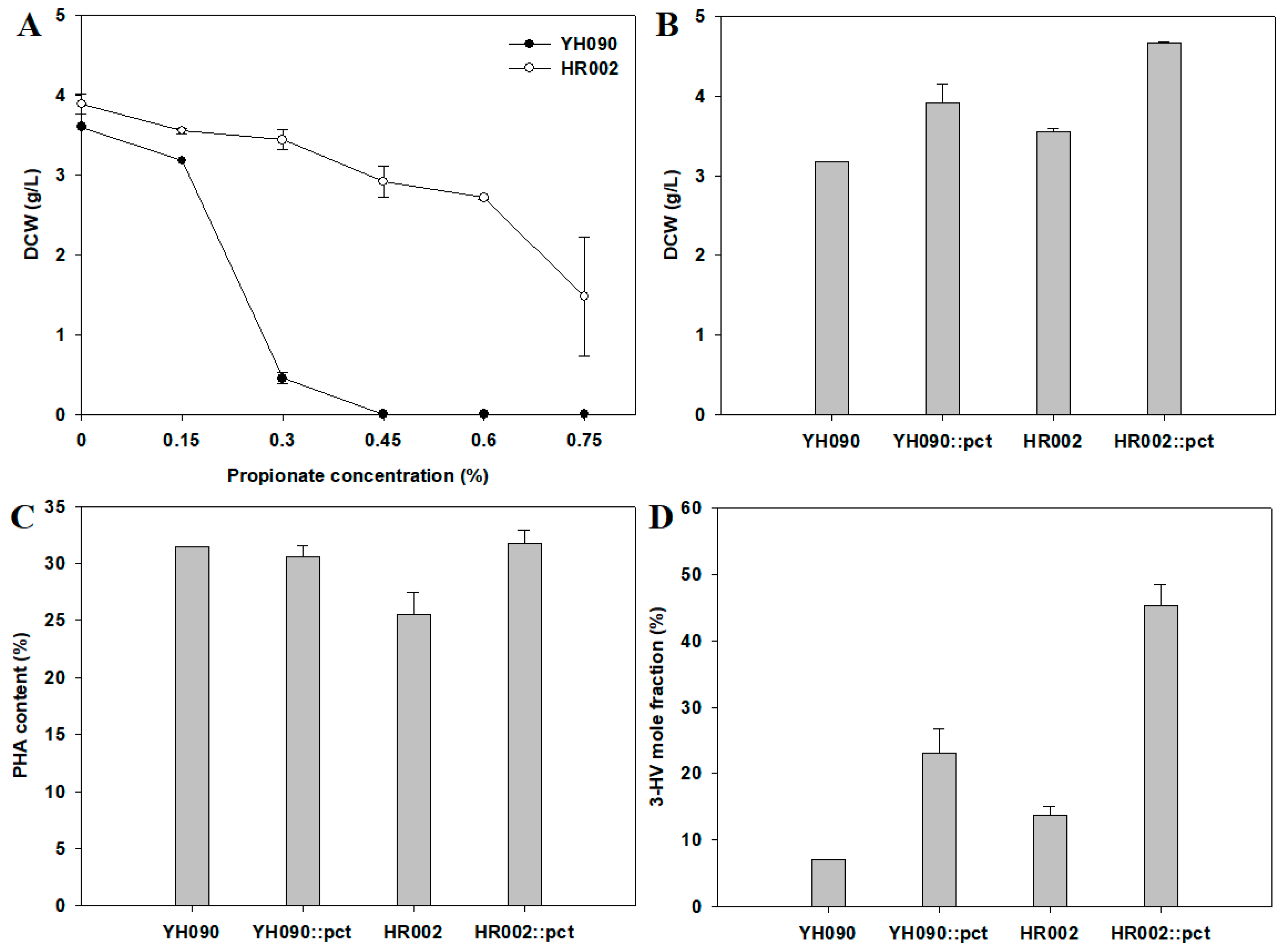
3.1. Construction of Engineered E. coli for Redirecting Metabolic Flux into PHA Production
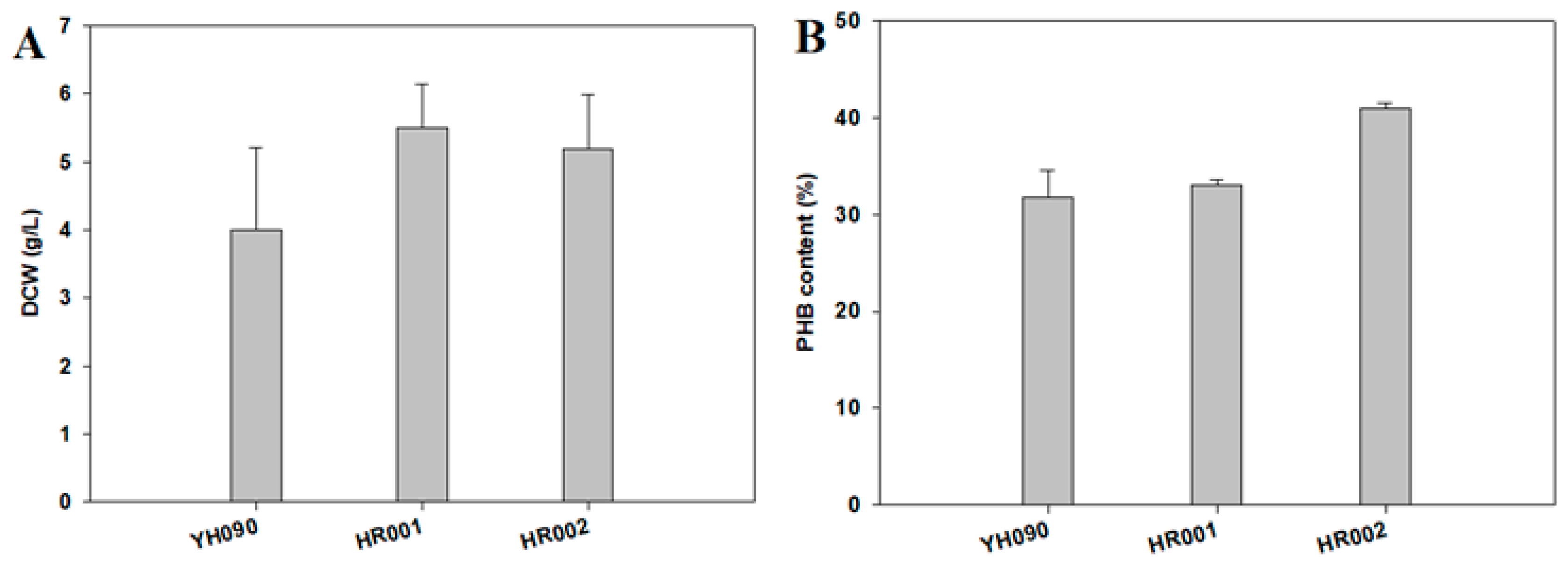
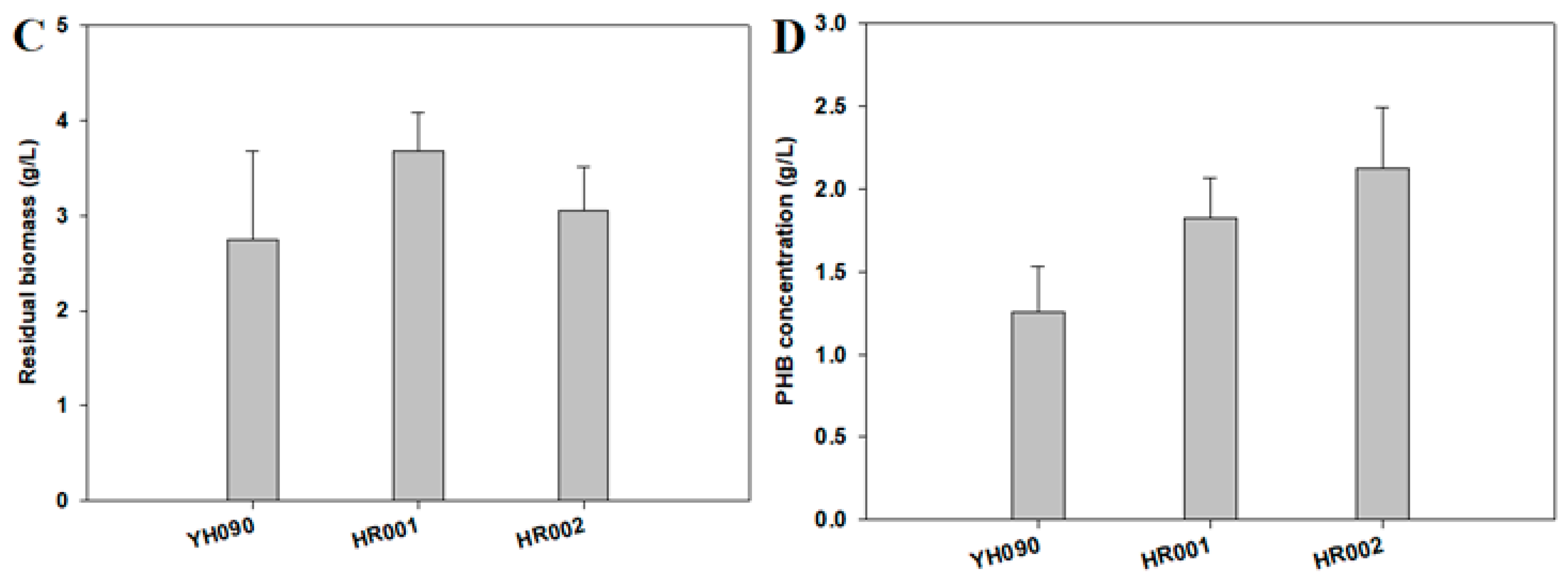
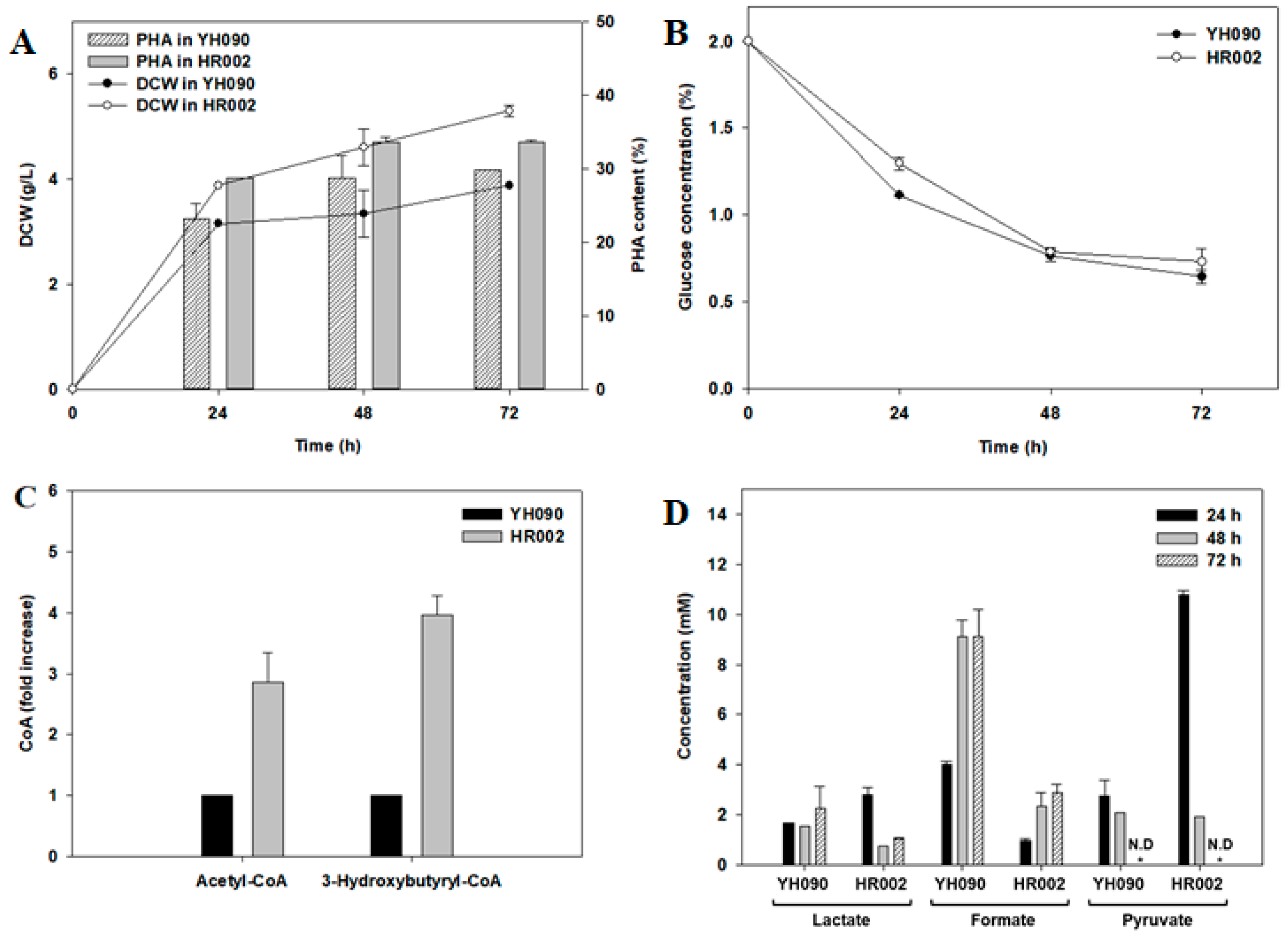
3.2. PHB Production and Metabolite Analysis in Engineered E. coli
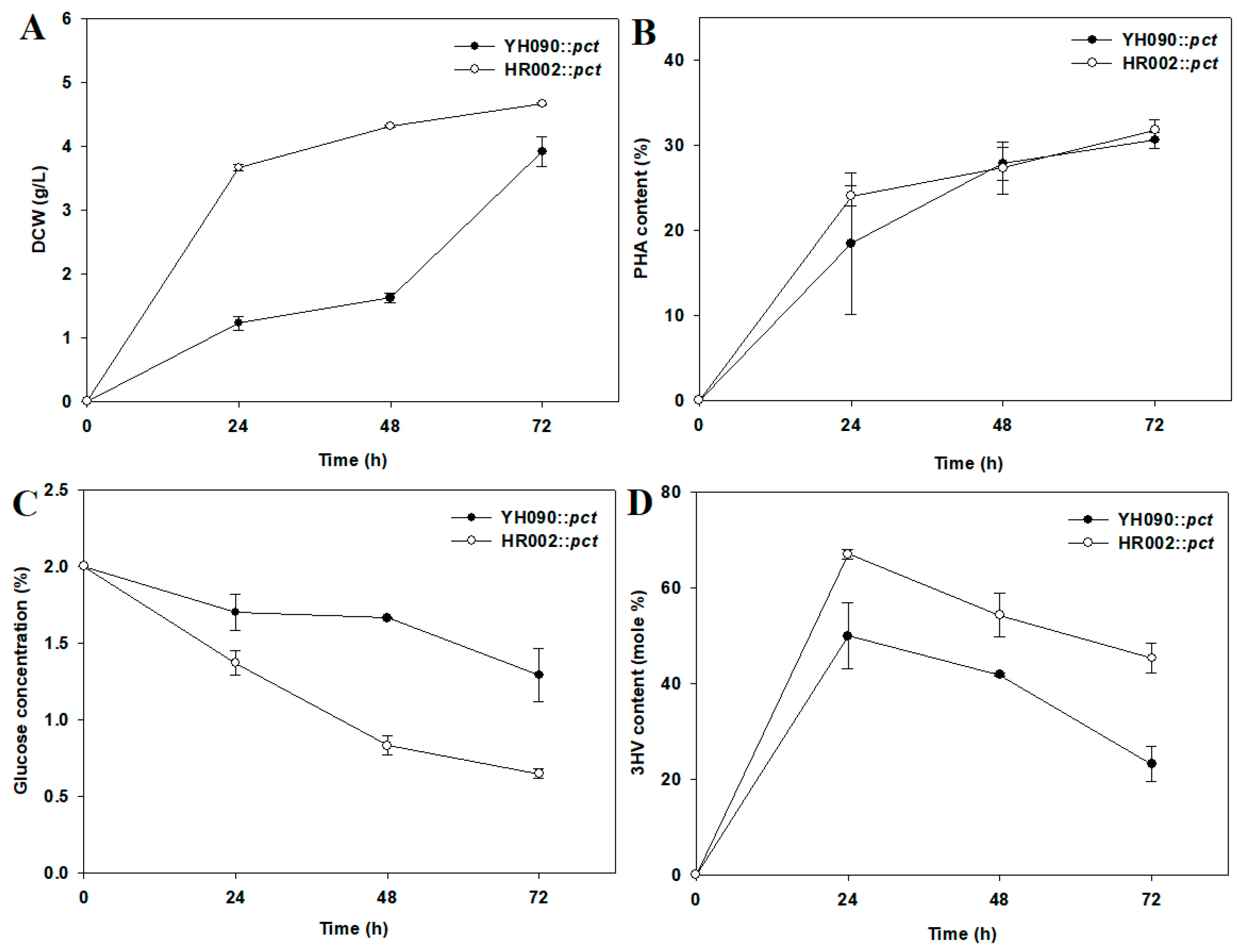
3.3. Application of the Engineered E. coli for PHBV Production
4. Discussion
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Madkour, M.H.; Heinrich, D. PHA recovery from biomass. Biomacromolecules 2013, 14, 2963–2972. [Google Scholar] [CrossRef]
- Steinbüchel, A.; Füchtenbusch, B. Bacterial and other biological systems for polyester production. Trends Biotechnol. 1998, 16, 419–427. [Google Scholar] [CrossRef]
- Steinbüchel, A. Non-biodegradable biopolymers from renewable resources: Perspectives and impacts. Curr. Opin. Biotechnol. 2005, 16, 607–613. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.Y. Bacterial polyhydroxyalkanoates. Biotechnol. Bioeng. 1996, 49, 1–14. [Google Scholar] [CrossRef]
- Liu, Q.; Lin, Z. Improved poly(3-hydroxybutyrate) production in Escherichia coli by inactivation of cytochrome bd-II oxidase or/and NDH-II dehydrogenase in low efficient respiratory chains. J. Biotechnol. 2014, 192, 170–176. [Google Scholar] [CrossRef] [PubMed]
- Du, C.; Sabirova, J. Polyhydroxyalkanoates production from low-cost sustainable raw materials. Curr. Chem. Biol. 2012, 6, 14–25. [Google Scholar]
- Li, R.; Zhang, H. The production of polyhydroxyalkanoates in recombinant Escherichia coli. Bioresour. Technol. 2007, 98, 2313–2320. [Google Scholar] [CrossRef]
- Li, Z.-J.; Cai, L. Overexpression of NAD kinase in recombinant Escherichia coli harboring the phbCAB operon improves poly(3-hydroxybutyrate) production. Appl. Microbiol. Biotechnol. 2009, 83, 939–947. [Google Scholar] [CrossRef]
- Bugnicourt, E.; Cinelli, P. Polyhydroxyalkanoate (PHA): Review of synthesis, characteristics, processing and potential applications in packaging. Express Polym. Lett. 2014, 8, 791–808. [Google Scholar] [CrossRef] [Green Version]
- Huo, G.; Zhu, Y. Metabolic engineering of an E. coli ndh knockout strain for PHB production from mixed glucose–xylose feedstock. J. Chem. Technol. Biotechnol. 2017, 92, 2739–2745. [Google Scholar] [CrossRef]
- Miyake, M.; Schnackenberg, J. Phosphotransacetylase as a key factor in biological production of polyhydroxybutyrate. Appl. Biochem. Biotechnol. 2000, 84, 1039–1044. [Google Scholar] [CrossRef]
- Lim, S.-J.; Jung, Y.-M. Amplification of the NADPH-related genes zwf and gnd for the oddball biosynthesis of PHB in an E. coli transformant harboring a cloned phbCAB operon. J. Biosci. Bioeng. 2002, 93, 543–549. [Google Scholar] [CrossRef]
- Kabir, M.M.; Shimizu, K. Gene expression patterns for metabolic pathway in pgi knockout Escherichia coli with and without phb genes based on RT-PCR. J. Biotechnol. 2003, 105, 11–31. [Google Scholar] [CrossRef]
- Mougiakos, I.; Bosma, E.F. Hijacking CRISPR-Cas for high-throughput bacterial metabolic engineering: Advances and prospects. Curr. Opin. Biotechnol. 2018, 50, 146–157. [Google Scholar] [CrossRef] [PubMed]
- Doudna, J.A.; Charpentier, E. The new frontier of genome engineering with CRISPR-Cas9. Science 2014, 346, 1258096. [Google Scholar] [CrossRef] [PubMed]
- Jiang, W.; Bikard, D. RNA-guided editing of bacterial genomes using CRISPR-Cas systems. Nat. Biotechnol. 2013, 31, 233. [Google Scholar] [CrossRef] [PubMed]
- Pyne, M.E.; Moo-Young, M. Coupling the CRISPR/Cas9 system to lambda Red recombineering enables simplified chromosomal gene replacement in Escherichia coli. Appl. Environ. Microbiol. 2015, 81, 5103–5114. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.; Chen, B. Multigene editing in the Escherichia coli genome using the CRISPR-Cas9 system. Appl. Environ. Microbiol. 2015. [Google Scholar] [CrossRef] [PubMed]
- Pettinari, M.J.; Vázquez, G.J. Poly(3-Hydroxybutyrate) Synthesis Genes in Azotobacter sp. Strain FA8. Appl. Environ. Microbiol. 2001, 67, 5331–5334. [Google Scholar] [CrossRef]
- Li, Z.-J.; Shi, Z.-Y. Production of poly(3-hydroxybutyrate-co-4-hydroxybutyrate) from unrelated carbon sources by metabolically engineered Escherichia coli. Metab. Eng. 2010, 12, 352–359. [Google Scholar] [CrossRef]
- Braunegg, G.; Sonnleitner, B. A rapid gas chromatographic method for the determination of poly-β-hydroxybutyric acid in microbial biomass. Appl. Microbiol. Biotechnol. 1978, 6, 29–37. [Google Scholar] [CrossRef]
- Bhatia, S.; Yi, D.H. Overexpression of succinyl-CoA synthase for poly(3-hydroxybutyrate-co-3-hydroxyvalerate) production in engineered Escherichia coli BL 21 (DE 3). J. Appl. Microbiol. 2015, 119, 724–735. [Google Scholar] [CrossRef]
- Bhatia, S.K.; Yi, D.-H. Development of semi-synthetic microbial consortia of Streptomyces coelicolor for increased production of biodiesel (fatty acid methyl esters). Fuel 2015, 159, 189–196. [Google Scholar] [CrossRef]
- Hong, Y.-G.; Moon, Y.-M. Production of glutaric acid from 5-aminovaleric acid using Escherichia coli whole cell bio-catalyst overexpressing GabTD from Bacillus subtilis. Enzyme Microb. Technol. 2018, 118, 57–65. [Google Scholar] [CrossRef] [PubMed]
- Park, J.W.; Jung, W.S. Analysis of intracellular short organic acid-coenzyme A esters from actinomycetes using liquid chromatography-electrospray ionization-mass spectrometry. J. Mass Spectrom. 2007, 42, 1136–1147. [Google Scholar] [CrossRef]
- Yang, Y.H.; Brigham, C. Biosynthesis of poly(3-hydroxybutyrate-co-3-hydroxyvalerate) containing a predominant amount of 3-hydroxyvalerate by engineered Escherichia coli expressing propionate-CoA transferase. J. Appl. Microbiol. 2012, 113, 815–823. [Google Scholar] [CrossRef] [PubMed]
- Jeon, J.-M.; Kim, H.-J. Application of acetyl-CoA acetyltransferase (AtoAD) in Escherichia coli to increase 3-hydroxyvalerate fraction in poly(3-hydroxybutyrate-co-3-hydroxyvalerate). Bioprocess. Biosyst. Eng. 2017, 40, 781–789. [Google Scholar] [CrossRef]
- Kim, S.K.; Seong, W. CRISPR interference-guided multiplex repression of endogenous competing pathway genes for redirecting metabolic flux in Escherichia coli. Microb. Cell Fact. 2017, 16, 188. [Google Scholar] [CrossRef] [PubMed]
- Jones, J.A.; Toparlak, Ö.D. Metabolic pathway balancing and its role in the production of biofuels and chemicals. Curr. Opin. Biotechnol. 2015, 33, 52–59. [Google Scholar] [CrossRef] [Green Version]
- Datsenko, K.A.; Wanner, B.L. One-step inactivation of chromosomal genes in Escherichia coli K-12 using PCR products. Proc. Natl. Acad. Sci. USA 2000, 97, 6640–6645. [Google Scholar] [CrossRef]
- Cherrington, C.A.; Hinton, M. Effect of short-chain organic acids on macromolecular synthesis in Escherichia coli. J. Appl. Bacteriol. 1990, 68, 69–74. [Google Scholar] [CrossRef]
- Jensen, E.B.; Carlsen, S. Production of recombinant human growth hormone in Escherichia coli: Expression of different precursors and physiological effects of glucose, acetate, and salts. Biotechnol. Bioeng. 1990, 36, 1–11. [Google Scholar] [CrossRef]
- Li, Y.; Lin, Z. Metabolic engineering of Escherichia coli using CRISPR–Cas9 meditated genome editing. Metab. Eng. 2015, 31, 13–21. [Google Scholar] [CrossRef] [PubMed]
- Ronda, C.; Pedersen, L.E. CRMAGE: CRISPR optimized mage recombineering. Sci. Rep. 2016, 6, 19452. [Google Scholar] [CrossRef]
- Wei, X.-X.; Zheng, W.-T. Metabolic engineering of Escherichia coli for poly(3-hydroxybutyrate) production under microaerobic condition. BioMed Res. Int. 2015, 2015, 789315. [Google Scholar] [CrossRef]
- Ma, W.; Wang, J. Poly(3-hydroxybutyrate-co-3-hydroxyvalerate) co-produced with l-isoleucine in Corynebacterium glutamicum WM001. Microb. Cell Fact. 2018, 17, 93. [Google Scholar] [CrossRef] [PubMed]
- Jian, J.; Zhang, S.-Q. Production of polyhydroxyalkanoates by Escherichia coli mutants with defected mixed acid fermentation pathways. Appl. Microbiol. Biotechnol. 2010, 87, 2247–2256. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.-W.; Wang, H.-H. Biosynthesis of poly(3-hydroxybutyrate-co-3-hydroxyvalerate) by recombinant Escherichia coli harboring propionyl-CoA synthase gene (prpE) or propionate permease gene (prpP). Biochem. Eng. J. 2009, 43, 72–77. [Google Scholar] [CrossRef]
- Yang, Y.-H.; Brigham, C. Improved detergent-based recovery of polyhydroxyalkanoates (PHAs). Biotechnol. Lett. 2011, 33, 937–942. [Google Scholar] [CrossRef] [Green Version]
- Seo, H.-M.; Jeon, J.-M. Combinatorial application of two aldehyde oxidoreductases on isobutanol production in the presence of furfural. J. Ind. Microbiol. Biotechnol. 2016, 43, 37–44. [Google Scholar] [CrossRef]
- Lynd, L.R.; Laser, M.S. How biotech can transform biofuels. Nat. Biotechnol. 2008, 26, 169. [Google Scholar] [CrossRef] [PubMed]
- Slater, S.; Gallaher, T. Production of poly-(3-hydroxybutyrate-co-3-hydroxyvalerate) in a recombinant Escherichia coli strain. Appl. Environ. Microbiol. 1992, 58, 1089–1094. [Google Scholar] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain or Plasmid | Description | Reference |
|---|---|---|
| E. coli strains | ||
| DH5α | General cloning strain | Invitrogen |
| BW25113 | lacIq, rrnB3, ∆lacZ4787, bsdR514, ∆araBAD, ∆rhaBAD | [21] |
| BW25113(DE3) | BW25113 derivative containing DE3 | [22] |
| YH090 | BW25113 (DE3) containing pLW487 | [22] |
| HR100 | BW25113 (DE3), ∆pflb, ∆ldhA, ∆adhE, ∆fnr | This study |
| HR200 | BW25113 (DE3), ∆pflb, ∆ldhA, ∆adhE, ∆fnr, containing pACYC::pntAB | This study |
| HR001 | BW25113 (DE3), ∆pflb, ∆ldhA, ∆adhE, ∆fnr, containing pLW487 | This study |
| HR002 | HR001 containing pACYC::pntAB | This study |
| Plasmids | ||
| pLW487 | Spectinomycin-resistant pEP2-based plasmid carrying PCR products of bktB, phaB, and phaC under the trc promoter from Ralstonia eutropha H16 | [23] |
| pACYCDuet-1 | A compatible chloramphenicol-resistant plasmid carrying the T7 promoter | Novagen |
| pACYC::pntAB | pACYCDuet-1 carrying the PCR product of pntAB from E. coli K12 MG1655 | [24] |
| pET-28a | A compatible kanamycin-resistant plasmid carrying the T7 promoter | Novagen |
| pET-28a::pct | pET-28a carrying the PCR product of pct from Ralstonia eutropha H16 | This study |
| pCas | repA101 (Ts), kanamycin-resistant, cas9 gene, Red recombination genes under an arabinose-inducible promoter, lacIq, Ptrc-sgRNA targeting pBR322 origin of pgRNA | This study |
| pgRNA | pBR322 origin, ampicillin-resistant, sgRNA with an N20 sequence for targeting gene | This study |
| Primers | ||
| N20_pflb_F | CTCT ACTAGT CATCGTATTCCGGAGTACGC GTTTTAGAGCTAGAAATAGC | This study |
| N20_ldhA_F | CTCT ACTAGT TTAAACCAGTTCGTTCGGGC GTTTTAGAGCTAGAAATAGC | This study |
| N20_adhE_F | CTCT ACTAGT CCGAAAGCACACAGGGACTT GTTTTAGAGCTAGAAATAGC | This study |
| N20_fnr_F | CTCT ACTAGT TTCCGCCTGACGATGACTCG GTTTTAGAGCTAGAAATAGC | This study |
| N20_R | CTCT ACTAGT ATTATACCTAGGAC | This study |
| E. coli Strain | Carbon Source | DCW (g/L) | PHA Content (wt %) | 3-HV (mol %) | Y3HV/Prop (g 3-HV/g Propionate) | Cultivation | Reference |
|---|---|---|---|---|---|---|---|
| YH090::pct (BW25113 (DE3) containing pLW487 (PHA operon) and pct from C. propionicum) | 1% glucose, 0.3% propionate | 2 | 50 | 80 | 0.30 | Batch (Flask) | [39] |
| YH090::sucCD (BW25113 (DE3) containing pLW487 (PHA operon) and sucCD) | 1% glucose, 0.3% propionate | 1.2 | 54.8 | 24.1 | 0.09 | Batch (Flask) | [28] |
| YH090::pct (BW25113 (DE3) containing pLW487 (PHA operon) and pctfrom R. eutropha H16) | 2% glucose, 0.15% propionate | 3.9 | 30.6 | 23.1 | 0.34 | Batch (Flask) | This study |
| HR002::pct (BW25113 (DE3) containing pLW487 (PHA operon), pntAB, pct from R. eutropha H16, ∆pflb, ∆ldhA, ∆adhE, ∆fnr) | 2% glucose, 0.15% propionate | 4.5 | 32 | 50 | 0.67 | Batch (Flask) | This study |
| LS5218 containing p4A (PHA operon), fadR, atoC | 0.4% glucose, 25 mM propionate | - | - | 39 | 0.10 | Batch (Flask) | [42] |
| BL21 containing pLW487 (PHA operon) and atoAD | 1% glucose, 0.3% propionate | 3 | 35 | 70 | 0.30 | Fed-batch (Flask) | [32] |
| XL10 containing pBHR68 (PHA operon) and prpP | 2% glucose, 0.2% propionate | 39.8 | 60.5 | 15.1 | 0.46 | Fed-batch (Fermentor) | [38] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jung, H.-R.; Yang, S.-Y.; Moon, Y.-M.; Choi, T.-R.; Song, H.-S.; Bhatia, S.K.; Gurav, R.; Kim, E.-J.; Kim, B.-G.; Yang, Y.-H. Construction of Efficient Platform Escherichia coli Strains for Polyhydroxyalkanoate Production by Engineering Branched Pathway. Polymers 2019, 11, 509. https://doi.org/10.3390/polym11030509
Jung H-R, Yang S-Y, Moon Y-M, Choi T-R, Song H-S, Bhatia SK, Gurav R, Kim E-J, Kim B-G, Yang Y-H. Construction of Efficient Platform Escherichia coli Strains for Polyhydroxyalkanoate Production by Engineering Branched Pathway. Polymers. 2019; 11(3):509. https://doi.org/10.3390/polym11030509
Chicago/Turabian StyleJung, Hye-Rim, Su-Yeon Yang, Yu-Mi Moon, Tae-Rim Choi, Hun-Suk Song, Shashi Kant Bhatia, Ranjit Gurav, Eun-Jung Kim, Byung-Gee Kim, and Yung-Hun Yang. 2019. "Construction of Efficient Platform Escherichia coli Strains for Polyhydroxyalkanoate Production by Engineering Branched Pathway" Polymers 11, no. 3: 509. https://doi.org/10.3390/polym11030509