Amorphous Phase Mediated Crystallization: Fundamentals of Biomineralization



Abstract
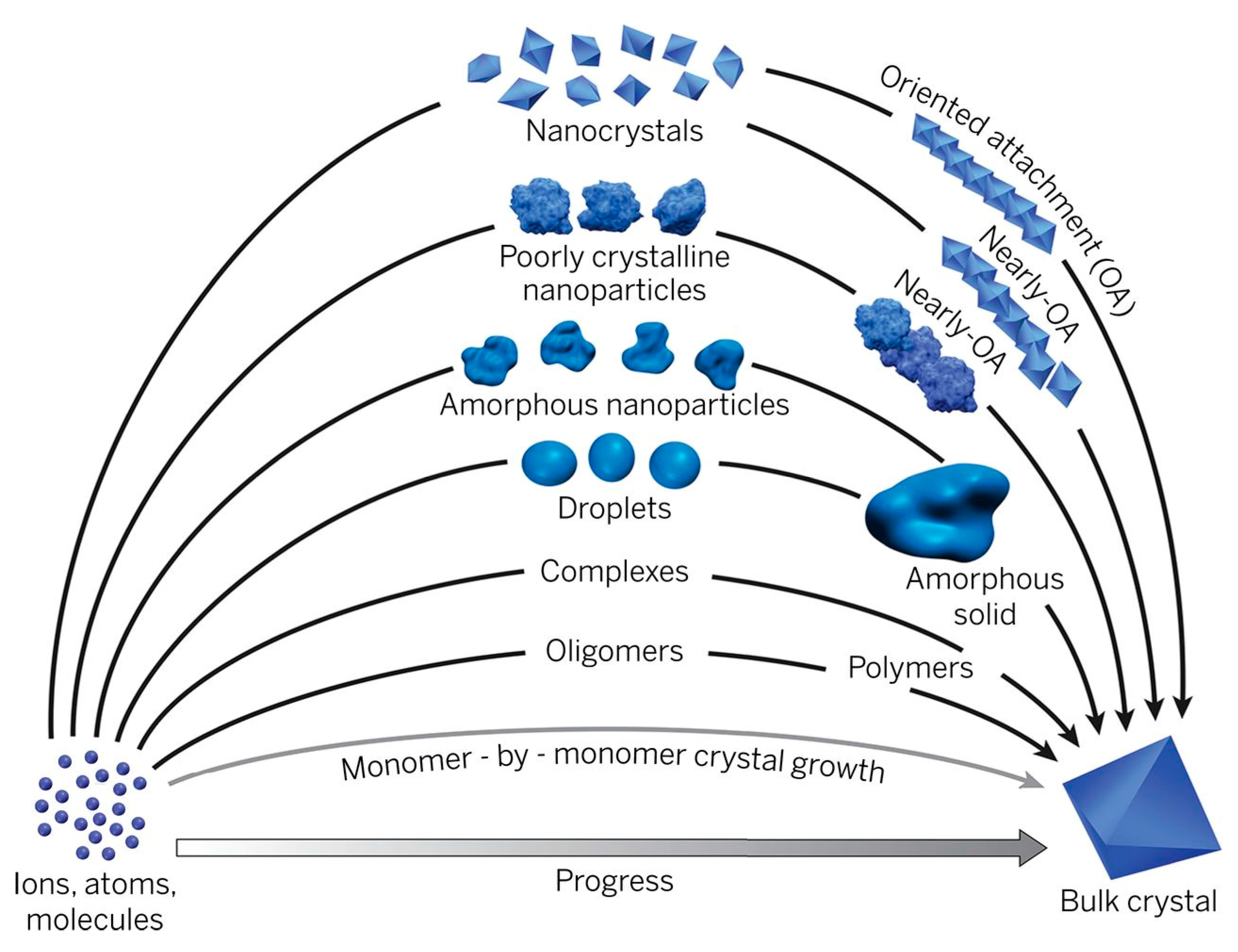
:1. Introduction
2. Amorphous Phase—Mediated Crystallization Pathways in Bio- and Biomimetic Mineralization Systems
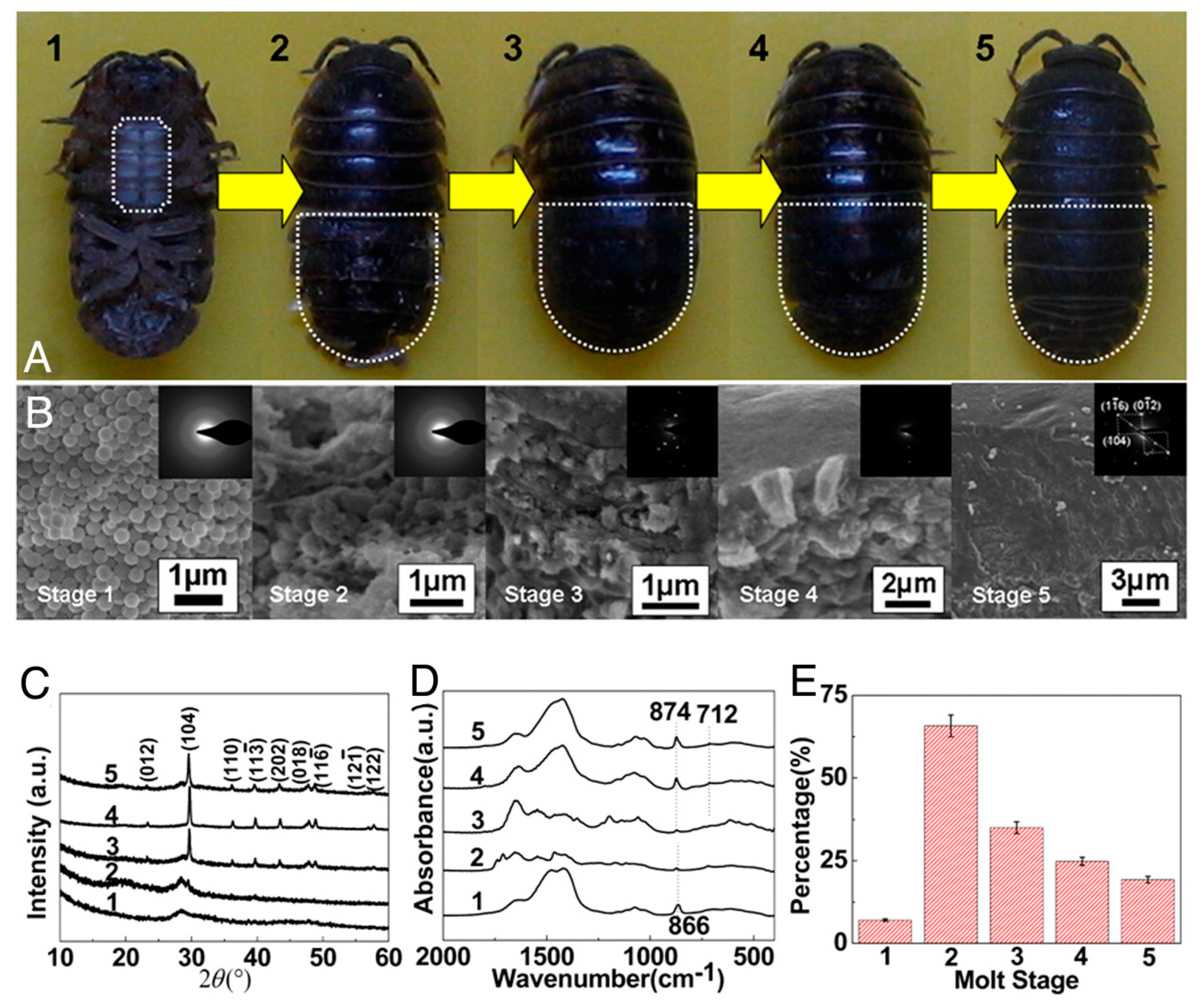
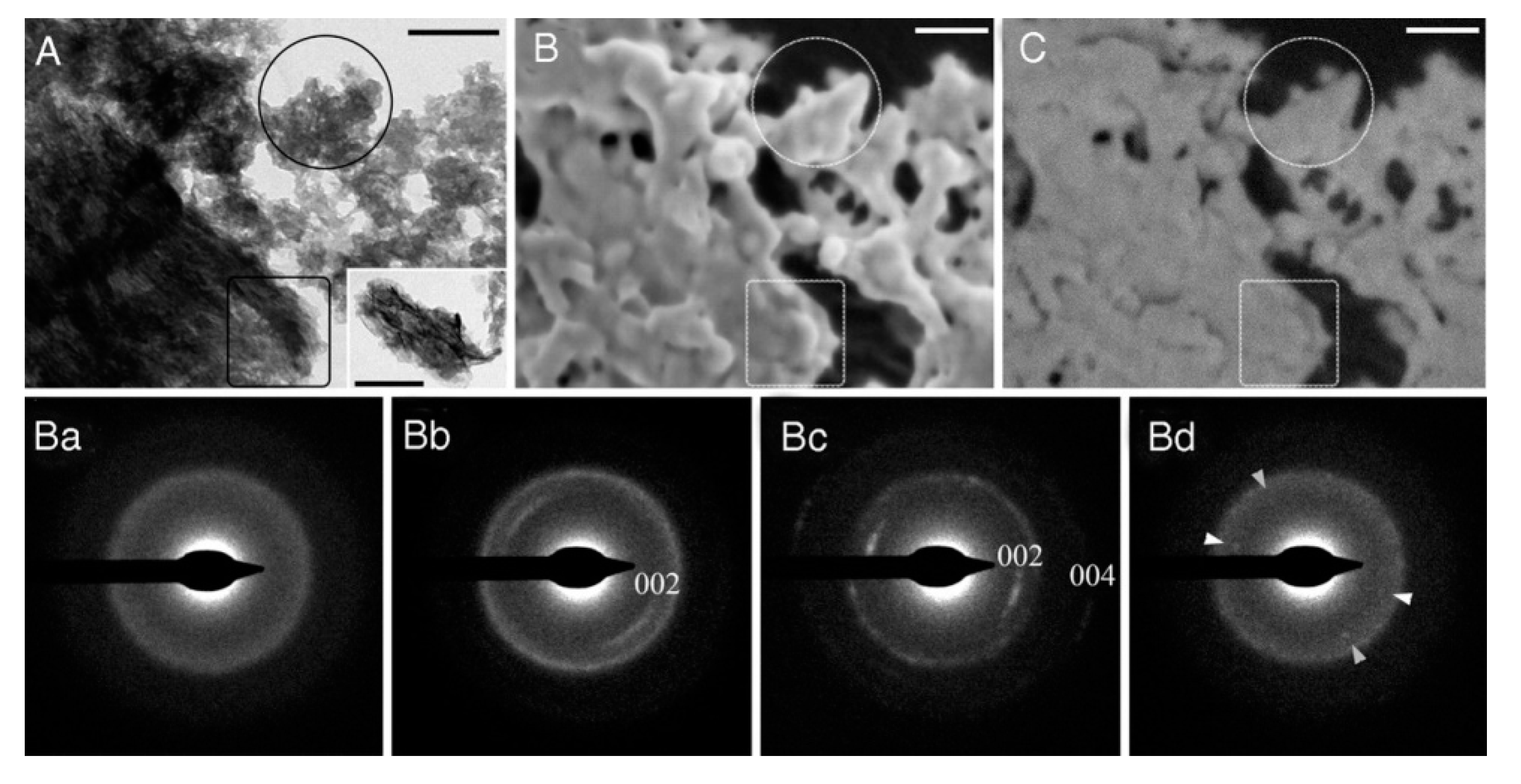
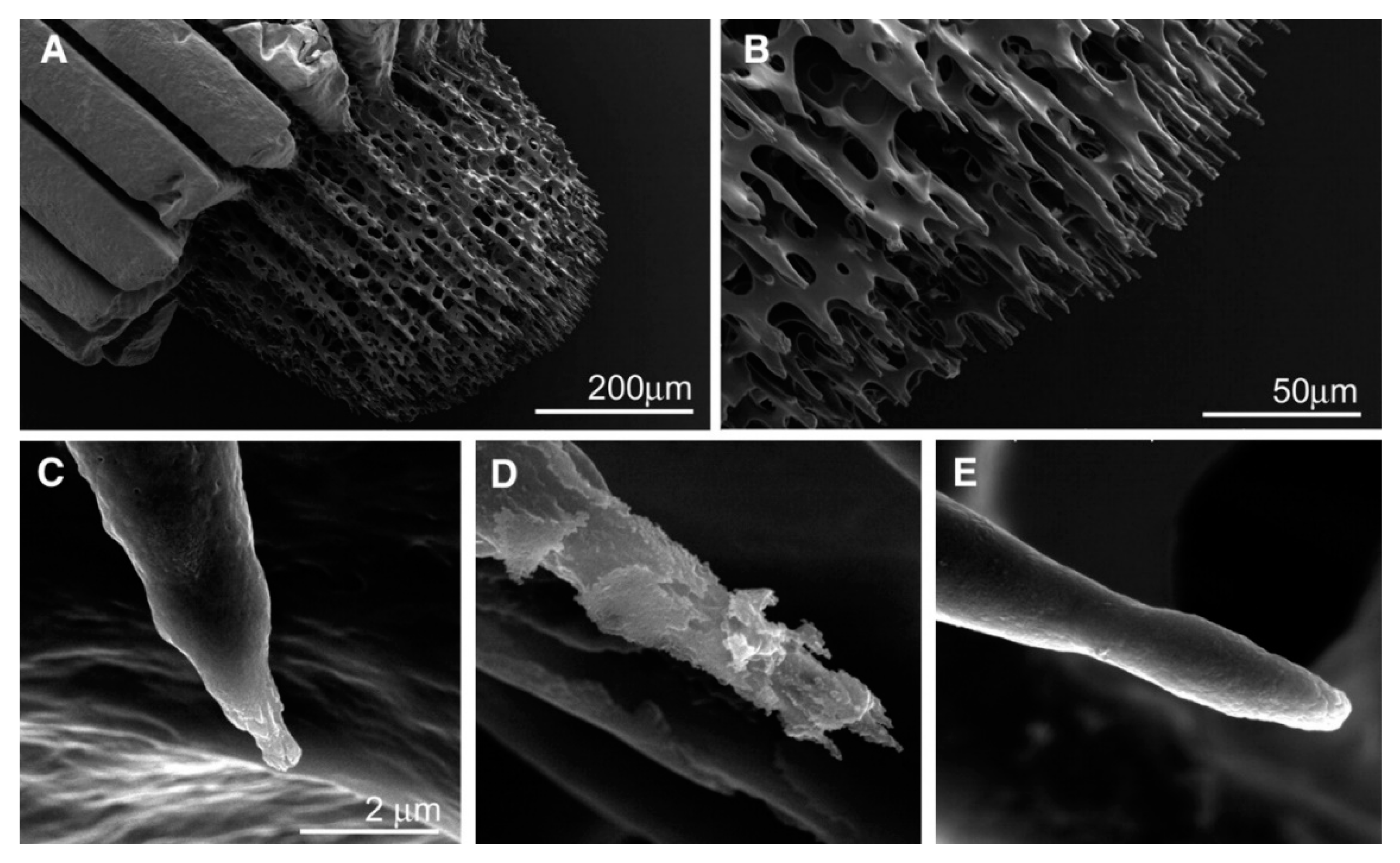
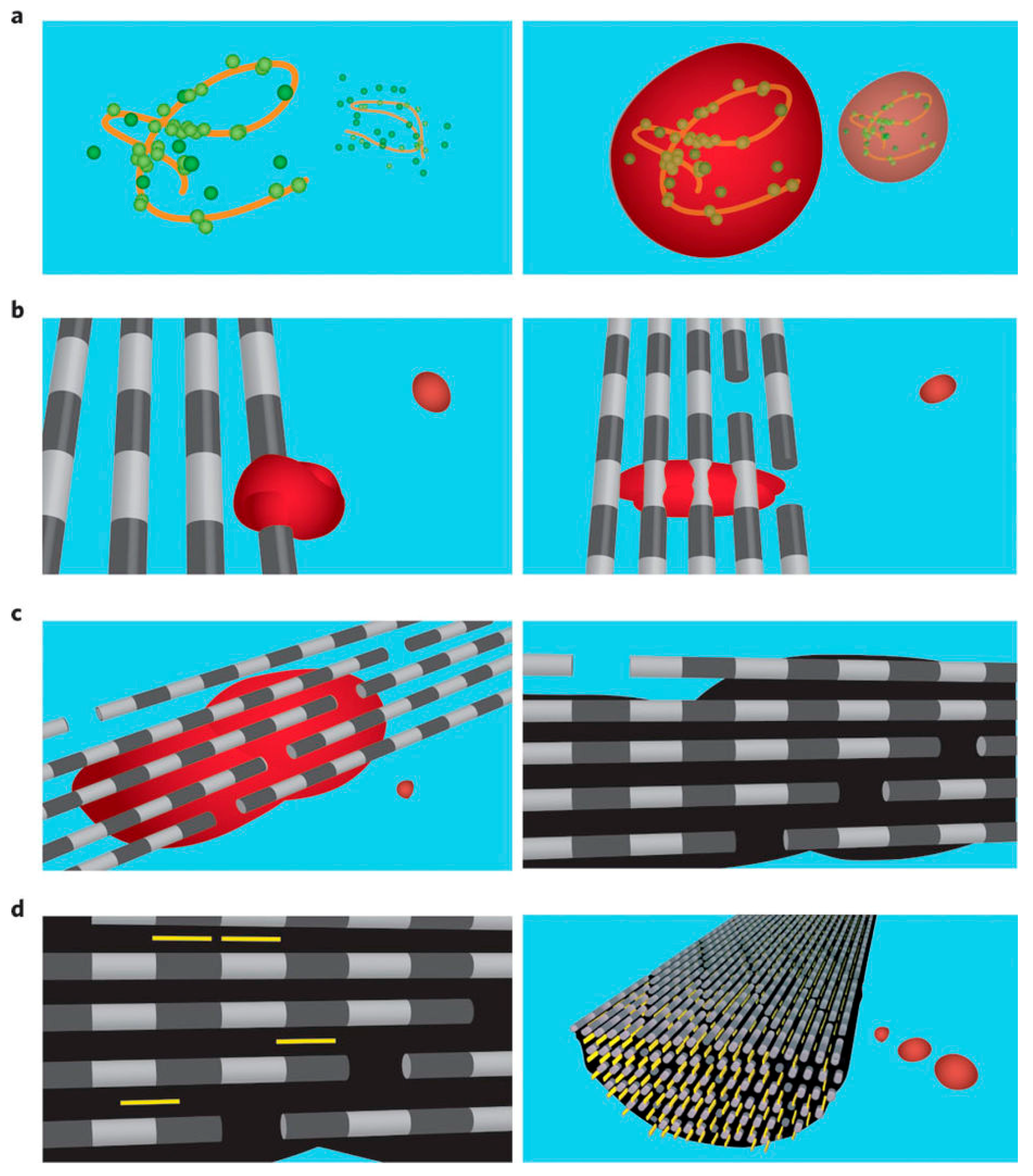
2.1. Biomineralization Systems
2.2. Biomimetic Mineralization Systems
2.2.1. Mineralization Medium
2.2.2. Biomimetic Organic Matrix
2.2.3. Biomimetic Mineralization in the Presence of Non-Matrix Proteins and Their Analogues
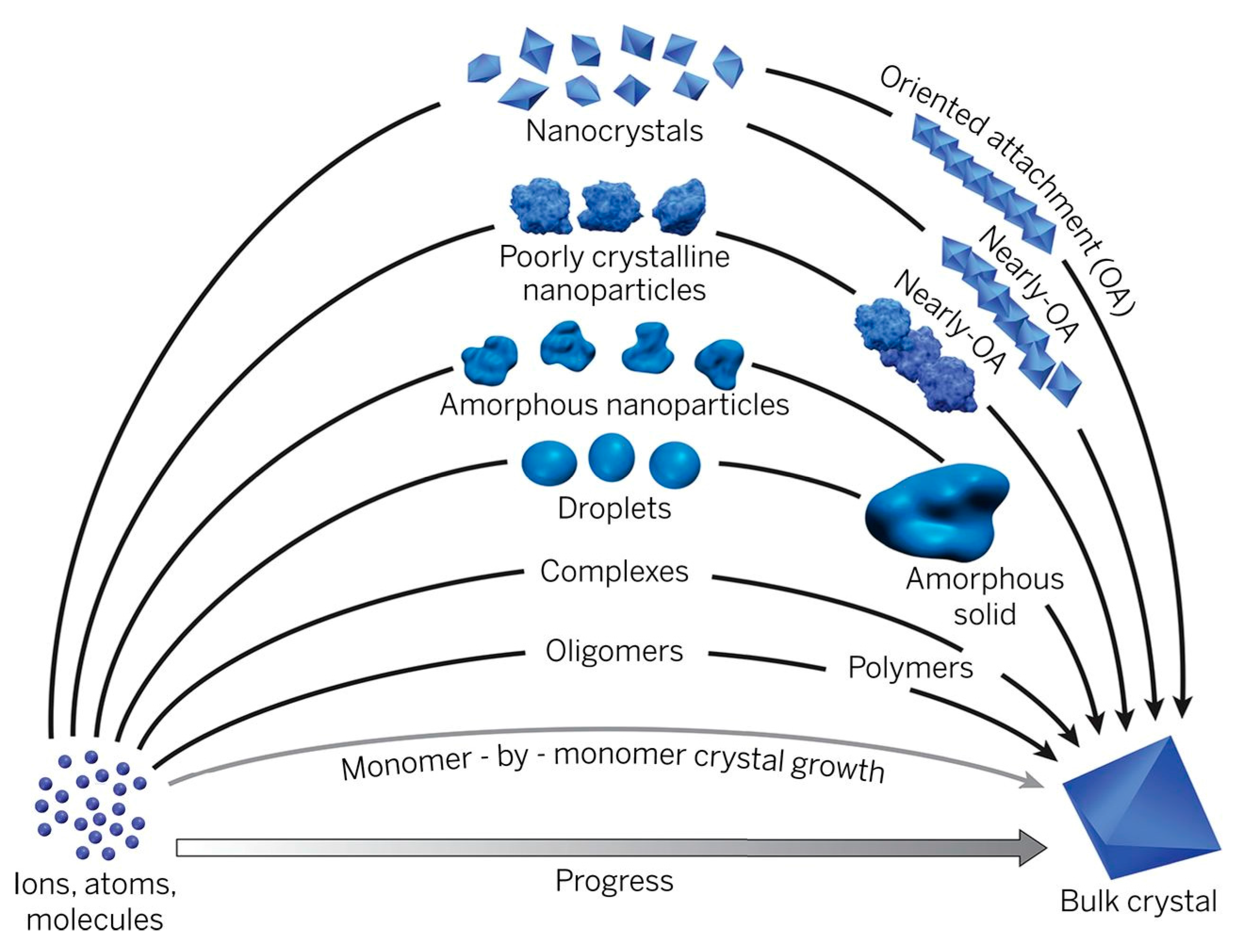
3. Kinetics of Amorphous Mineral Phase-Mediated Crystallization
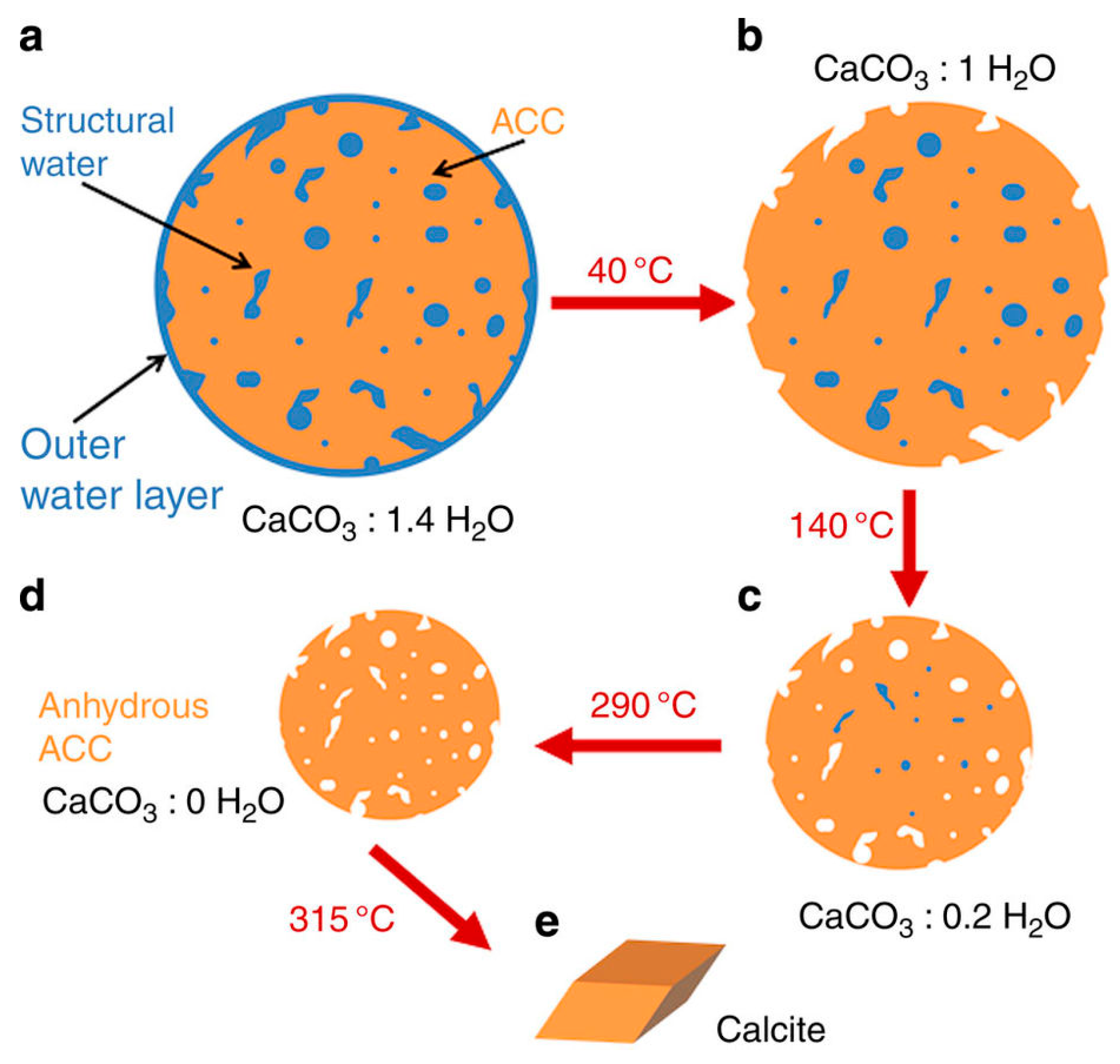
3.1. Determination of Amorphous Phases and Their Solubility
3.2. Classical Homogeneous and Heterogeneous Nucleation Theory
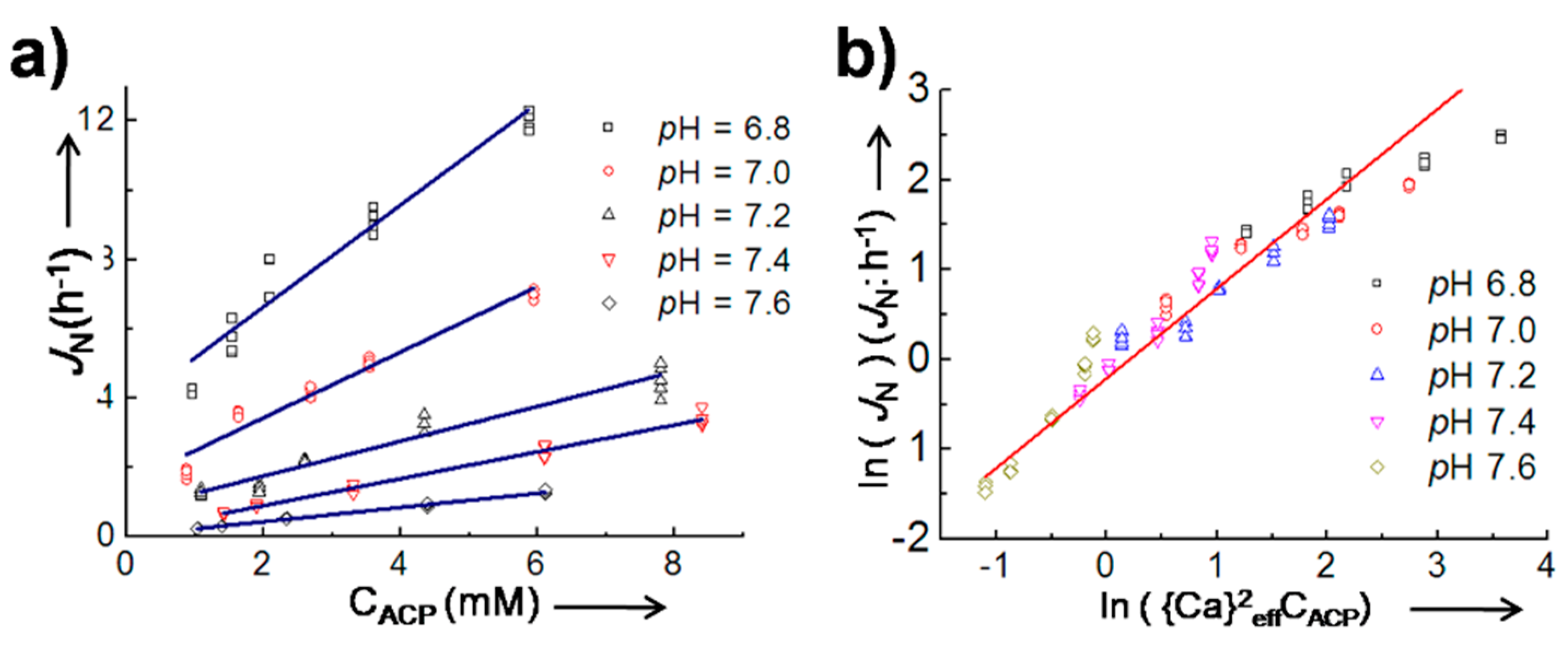
3.3. Amorphous Phase Mediated Nucleation
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Mann, S. General principles of biomineralization. In Biominer. Princ. Concepts Bioinorg. Mater. Chem; Oxford University Press: New York, NY, USA, 2002. [Google Scholar]
- Lowenstam, H.A.; Weiner, S. On Biomineralization; Oxford University Press: New York, NY, USA, 1989. [Google Scholar]
- De Yoreo, J.; Vekilov, P.G. Principles of Crystal Nucleation and Growth. Rev. Miner. Geochem. 2003, 54, 57–93. [Google Scholar] [CrossRef]
- Taylor, M.G.; Simkiss, K.; Greaves, G.N.; Okazaki, M.; Mann, S. An X-ray absorption spectroscopy study of the structure and transformation of amorphous calcium carbonate from plant cystoliths. Proc. R. Soc. Lond. Ser. B-Biol. Sci. 1993, 252, 75–80. [Google Scholar] [CrossRef]
- Bauer, P.; Elbaum, R.; Weiss, I.M. Calcium and silicon mineralization in land plants: Transport, structure and function. Plant. Sci. 2011, 180, 746–756. [Google Scholar] [CrossRef] [PubMed]
- Gal, A.; Hirsch, A.; Siegel, S.; Li, C.; Aichmayer, B.; Politi, Y.; Fratzl, P.; Weiner, S.; Addadi, L. Plant cystoliths: A complex functional biocomposite of four distinct silica and amorphous calcium carbonate phases. Chem.—A Eur. J. 2012, 18, 10262–10270. [Google Scholar] [CrossRef] [PubMed]
- He, H.; Veneklaas, E.J.; Kuo, J.; Lambers, H. Physiological and ecological significance of biomineralization in plants. Trends Plant. Sci. 2014, 19, 166–174. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.Y.; Lim, S.W. Templating and supersaturation-driven anti-templating: Principles of biomineral architecture. J. Am. Chem. Soc. 2003, 125, 888–895. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Nancollas, G.H. Calcium orthophosphates: Crystallization and dissolution. Chem. Rev. 2008, 108, 4628–4669. [Google Scholar] [CrossRef] [PubMed]
- Jiang, S.; Chen, Y.; Pan, H.; Zhang, Y.-J.; Tang, R. Faster nucleation at lower pH: Amorphous phase mediated nucleation kinetics. Phys. Chem. Chem. Phys. 2013, 15, 12530. [Google Scholar] [CrossRef] [PubMed]
- Jiang, S.; Pan, H.; Chen, Y.; Xu, X.; Tang, R. Amorphous calcium phosphate phase-mediated crystal nucleation kinetics and pathway. Faraday Discuss. 2015, 179, 451–461. [Google Scholar] [CrossRef] [PubMed]
- Blue, C.R.; Giuffre, A.; Mergelsberg, S.; Han, N.; De Yoreo, J.J.; Dove, P.M. Chemical and physical controls on the transformation of amorphous calcium carbonate into crystalline CaCO3 polymorphs. Geochim. Cosmochim. Acta 2017, 196, 179–196. [Google Scholar] [CrossRef]
- Liu, X.Y.; De Yoreo, J. Nanoscale Structure and Assembly at Solid-Fluid Interfaces: Assembly in Hybrid and Biological Systems; Kluwer Academic Press: Dordrecht, The Netherlands, 2004. [Google Scholar]
- De Yoreo, J.J.; Gilbert, P.U.P.A.; Sommerdijk, N.A.J.M.; Penn, R.L.; Whitelam, S.; Joester, D.; Zhang, H.; Rimer, J.D.; Navrotsky, A.; Banfield, J.F.; et al. Crystallization by particle attachment in synthetic, biogenic, and geologic environments. Science 2015, 349, aaa6760. [Google Scholar] [CrossRef] [PubMed]
- Banfield, J.F. Aggregation-Based Crystal growth and microstructure development in natural iron oxyhydroxide biomineralization products. Science 2000, 289, 751–754. [Google Scholar] [CrossRef] [PubMed]
- Cölfen, H.; Mann, S. Higher-order organization by mesoscale self-assembly and transformation of hybrid nanostructures. Angew. Chem.—Int. Ed. 2003, 42, 2350–2365. [Google Scholar] [CrossRef] [PubMed]
- Gebauer, D.; Volkel, A.; Colfen, H. Stable prenucleation calcium carbonate clusters. Science 2008, 322, 1819–1822. [Google Scholar] [CrossRef] [PubMed]
- Gebauer, D.; Cölfen, H. Prenucleation clusters and non-classical nucleation. Nano Today 2011, 6, 564–584. [Google Scholar] [CrossRef]
- Pouget, E.M.; Bomans, P.H.H.; Goos, J.A.C.M.; Frederik, P.M.; de With, G.; Sommerdijk, N.A.J.M. The initial stages of template-controlled CaCO3 formation revealed by cryo-TEM. Science 2009, 323, 1455–1458. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dey, A.; Bomans, P.H.H.; Müller, F.A.; Will, J.; Frederik, P.M.; de With, G.; Sommerdijk, N.A.J.M. The role of prenucleation clusters in surface-induced calcium phosphate crystallization. Nat. Mater. 2010, 9, 1010–1014. [Google Scholar] [CrossRef] [PubMed]
- Gower, L.B. Biomimetic model systems for investigating the amorphous precursor pathway and its role in biomineralization. Chem. Rev. 2008, 108, 4551–4627. [Google Scholar] [CrossRef] [PubMed]
- Faatz, M.; Gröhn, F.; Wegner, G. Amorphous calcium carbonate: Synthesis and potential intermediate in biomineralization. Adv. Mater. 2004, 16, 996–1000. [Google Scholar] [CrossRef]
- Addadi, L.; Raz, S.; Weiner, S. Taking advantage of disorder: Amorphous calcium carbonate and its roles in biomineralization. Adv. Mater. 2003, 15, 959–970. [Google Scholar] [CrossRef]
- Dorozhkin, S.V. Amorphous calcium (ortho)phosphates. Acta Biomater. 2010, 6, 4457–4475. [Google Scholar] [CrossRef] [PubMed]
- Weiner, S.; Mahamid, J.; Politi, Y.; Ma, Y.; Addadi, L. Overview of the amorphous precursor phase strategy in biomineralization. Front. Mater. Sci. China 2009, 3, 104–108. [Google Scholar] [CrossRef]
- Cartwright, J.H.E.; Checa, A.G.; Gale, J.D.; Gebauer, D.; Sainz-Díaz, C.I. Calcium carbonate polyamorphism and its role in biomineralization: How many amorphous calcium carbonates are there? Angew. Chem.—Int. Ed. 2012, 51, 11960–11970. [Google Scholar] [CrossRef] [PubMed]
- Termine, J.D.; Posner, A.S. Infrared analysis of rat bone: Age dependency of amorphous and crystalline mineral fractions. Science 1966, 153, 1523–1525. [Google Scholar] [CrossRef] [PubMed]
- Boskey, A.L. Amorphous calcium phosphate: The contention of bone. J. Dent. Res. 1997, 76, 1433–1436. [Google Scholar] [CrossRef] [PubMed]
- Mahamid, J.; Aichmayer, B.; Shimoni, E.; Ziblat, R.; Li, C.; Siegel, S.; Paris, O.; Fratzl, P.; Weiner, S.; Addadi, L. Mapping amorphous calcium phosphate transformation into crystalline mineral from the cell to the bone in zebrafish fin rays. Proc. Natl. Acad. Sci. USA 2010, 107, 6316–6321. [Google Scholar] [CrossRef] [PubMed]
- Nudelman, F.; Bomans, P.H.H.; George, A.; de With, G.; Sommerdijk, N.A.J.M. The role of the amorphous phase on the biomimetic mineralization of collagen. Faraday Discuss. 2012, 159, 357. [Google Scholar] [CrossRef] [PubMed]
- Olszta, M.J.; Cheng, X.; Jee, S.S.; Kumar, R.; Kim, Y.Y.; Kaufman, M.J.; Douglas, E.P.; Gower, L.B. Bone structure and formation: A new perspective. Mater. Sci. Eng. R Rep. 2007, 58, 77–116. [Google Scholar] [CrossRef]
- Sailaja, G.S.; Ramesh, P.; Varma, H.K. Ultrastructural evaluation of in vitro mineralized calcium phosphate phase on surface phosphorylated poly(hydroxy ethyl methacrylate-co-methyl methacrylate). J. Mater. Sci. Mater. Med. 2010, 21, 1183–1193. [Google Scholar] [CrossRef] [PubMed]
- Onuma, K. Recent research on pseudobiological hydroxyapatite crystal growth and phase transition mechanisms. Prog. Cryst. Growth Charact. Mater. 2006, 52, 223–245. [Google Scholar] [CrossRef]
- Barrere, F.; Snel, M.M.E.; Van Blitterswijk, C.A.; De Groot, K.; Layrolle, P. Nano-scale study of the nucleation and growth of calcium phosphate coating on titanium implants. Biomaterials 2004, 25, 2901–2910. [Google Scholar] [CrossRef] [PubMed]
- Politi, Y.; Metzler, R.A.; Abrecht, M.; Gilbert, B.; Wilt, F.H.; Sagi, I.; Addadi, L.; Weiner, S.; Gilbert, P.U.P.A. Transformation mechanism of amorphous calcium carbonate into calcite in the sea urchin larval spicule. Proc. Natl. Acad. Sci. USA 2008, 105, 17362–17366. [Google Scholar] [CrossRef] [PubMed]
- Politi, Y. Sea Urchin spine calcite forms via a transient amorphous calcium carbonate phase. Science 2004, 306, 1161–1164. [Google Scholar] [CrossRef] [PubMed]
- Addadi, L.; Joester, D.; Nudelman, F.; Weiner, S. Mollusk shell formation: A source of new concepts for understanding biomineralization processes. Chem.—A Eur. J. 2006, 12, 980–987. [Google Scholar] [CrossRef] [PubMed]
- Nassif, N.; Pinna, N.; Gehrke, N.; Antonietti, M.; Jager, C.; Colfen, H. Amorphous layer around aragonite platelets in nacre. Proc. Natl. Acad. Sci. USA 2005, 102, 12653–12655. [Google Scholar] [CrossRef] [PubMed]
- Weiner, S. An Overview of biomineralization processes and the problem of the vital effect. Rev. Miner. Geochem. 2003, 54, 1–29. [Google Scholar] [CrossRef]
- Mass, T.; Giuffre, A.J.; Sun, C.-Y.; Stifler, C.A.; Frazier, M.J.; Neder, M.; Tamura, N.; Stan, C.V.; Marcus, M.A.; Gilbert, P.U.P.A. Amorphous calcium carbonate particles form coral skeletons. Proc. Natl. Acad. Sci. USA 2017, 114, 7670–7678. [Google Scholar] [CrossRef] [PubMed]
- Combes, C.; Rey, C. Amorphous calcium phosphates: Synthesis, properties and uses in biomaterials. Acta Biomater. 2010, 6, 3362–3378. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, X.R.; Cai, A.H.; Liu, R.; Pan, H.H.; Tang, R.K.; Cho, K. The roles of water and polyelectrolytes in the phase transformation of amorphous calcium carbonate. J. Cryst. Growth 2008, 310, 3779–3787. [Google Scholar] [CrossRef]
- Gal, A.; Habraken, W.; Gur, D.; Fratzl, P.; Weiner, S.; Addadi, L. Calcite crystal growth by a solid-state transformation of stabilized amorphous calcium carbonate nanospheres in a hydrogel. Angew. Chem.—Int. Ed. 2013, 52, 4867–4870. [Google Scholar] [CrossRef] [PubMed]
- Tester, C.C.; Wu, C.H.; Krejci, M.R.; Mueller, L.; Park, A.; Lai, B.; Chen, S.; Sun, C.; Balasubramanian, M.; Joester, D. Time-resolved evolution of short- and long-range order during the transformation of amorphous calcium carbonate to calcite in the sea urchin embryo. Adv. Funct. Mater. 2013, 23, 4185–4194. [Google Scholar] [CrossRef]
- Nielsen, M.H.; Aloni, S.; De Yoreo, J.J. In situ TEM imaging of CaCO3 nucleation reveals coexistence of direct and indirect pathways. Science 2014, 345, 1158–1162. [Google Scholar] [CrossRef] [PubMed]
- Puech, J.; Heughebaert, J.-C.; Montel, G. A new mode of growing apatite crystallites. J. Cryst. Growth 1982, 56, 20–25. [Google Scholar] [CrossRef]
- Chatzipanagis, K.; Iafisco, M.; Roncal-Herrero, T.; Bilton, M.; Tampieri, A.; Kröger, R.; Delgado-López, J.M. Crystallization of citrate-stabilized amorphous calcium phosphate to nanocrystalline apatite: A surface-mediated transformation. CrystEngComm 2016, 18, 3170–3173. [Google Scholar] [CrossRef]
- Pan, H.; Liu, X.Y.; Tang, R.; Xu, H.Y. Mystery of the transformation from amorphous calcium phosphate to hydroxyapatite. Chem. Commun. 2010, 46, 7415–7417. [Google Scholar] [CrossRef] [PubMed]
- Posner, A.; Betts, F. Synthetic amorphous calcium-phosphate and its relation to bone-mineral structure. Acc. Chem. Res. 1975, 8, 273–281. [Google Scholar] [CrossRef]
- Lazić, S. Microcrystalline hydroxyapatite formation from alkaline solutions. J. Cryst. Growth 1995, 147, 147–154. [Google Scholar] [CrossRef]
- Boskey, A.; Posner, A. Conversion of amorphous calcium phosphate to microcrystalline hydroxyapatite. A pH-dependent, solution-mediated, solid-solid conversion. J. Phys. Chem. 1973, 77, 2313–2317. [Google Scholar] [CrossRef]
- Somrani, S.; Banu, M.; Jemal, M.; Rey, C. Physico-chemical and thermochemical studies of the hydrolytic conversion of amorphous tricalcium phosphate into apatite. J. Solid State Chem. 2005, 178, 1337–1348. [Google Scholar] [CrossRef] [Green Version]
- Tung, M.S.; Brown, W.E. An intermediate state in hydrolysis of amorphous calcium phosphate. Calcif. Tissue Int. 1983, 35, 783–790. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Blanco, J.D.; Shaw, S.; Bots, P.; Roncal-Herrero, T.; Benning, L.G. The role of pH and Mg on the stability and crystallization of amorphous calcium carbonate. J. Alloys Compd. 2012, 536, 477–479. [Google Scholar] [CrossRef]
- Kwon, K.Y.; Wang, E.; Nofal, M.; Lee, S.W. Microscopic study of hydroxyapatite dissolution as affected by fluoride ions. Langmuir 2011, 27, 5335–5339. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.R.I.; Han, T.Y.J.; Willey, T.M.; Wang, D.; Meulenberg, R.W.; Nilsson, J.; Dove, P.M.; Terminello, L.J.; Van Buuren, T.; De Yoreo, J.J. Structural development of mercaptophenol self-assembled monolayers and the overlying mineral phase during templated CaCO3 crystallization from a transient amorphous film. J. Am. Chem. Soc. 2007, 129, 10370–10381. [Google Scholar] [CrossRef] [PubMed]
- Ihli, J.; Wong, W.C.; Noel, E.H.; Kim, Y.-Y.; Kulak, A.N.; Christenson, H.K.; Duer, M.J.; Meldrum, F.C. Dehydration and crystallization of amorphous calcium carbonate in solution and in air. Nat. Commun. 2014, 5, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Liao, J.; Gou, B.; Huang, J.; Tang, R.; Tao, J.; Zhang, T.; Wang, K. Crystallization at Multiple Sites inside Particles of Amorphous Calcium Phosphate. Cryst. Growth Des. 2009, 9, 2620–2626. [Google Scholar] [CrossRef]
- Kim, S.; Ryu, H.S.; Shin, H.; Jung, H.S.; Hong, K.S. In situ observation of hydroxyapatite nanocrystal formation from amorphous calcium phosphate in calcium-rich solutions. Mater. Chem. Phys. 2005, 91, 500–506. [Google Scholar] [CrossRef]
- Abbona, F.; Baronnet, A. A XRD and TEM study on the transformation of amorphous calcium phosphate in the presence of magnesium. J. Cryst. Growth 1996, 165, 98–105. [Google Scholar] [CrossRef]
- Onuma, K.; Ito, A. Cluster growth model for hydroxyapatite. Chem. Mater. 1998, 10, 3346–3351. [Google Scholar] [CrossRef]
- Amos, F.F.; Sharbaugh, D.M.; Talham, D.R.; Gower, L.B.; Fricke, M.; Volkmer, D. Formation of single-crystalline aragonite tablets/films via an amorphous precursor. Langmuir 2007, 23, 1988–1994. [Google Scholar] [CrossRef] [PubMed]
- Weiner, S.; Lowenstam, H. Organization of Extracellularly Mineralized Tissues: A Comparative Study of Biological Crystal Growt. Crit. Rev. Biochem. 1986, 20, 365–408. [Google Scholar] [CrossRef]
- Mahamid, J.; Sharir, A.; Addadi, L.; Weiner, S. Amorphous calcium phosphate is a major component of the forming fin bones of zebrafish: Indications for an amorphous precursor phase. Proc. Natl. Acad. Sci. USA 2008, 105, 12748–12753. [Google Scholar] [CrossRef] [PubMed]
- Boonrungsiman, S.; Gentleman, E.; Carzaniga, R.; Evans, N.D.; McComb, D.W.; Porter, A.E.; Stevens, M.M. The role of intracellular calcium phosphate in osteoblast-mediated bone apatite formation. Proc. Natl. Acad. Sci. USA 2012, 109, 14170–14175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beniash, E.; Metzler, R.A.; Lam, R.S.K.; Gilbert, P.U.P.A. Transient amorphous calcium phosphate in forming enamel. J. Struct. Biol. 2009, 166, 133–143. [Google Scholar] [CrossRef] [PubMed]
- Lowenstam, H.A.; Weiner, S. Transformation of amorphous calcium phosphate to crystalline dahllite in the radular teeth of chitons. Science 1985, 227, 51–54. [Google Scholar] [CrossRef] [PubMed]
- Lévêque, I.; Cusack, M.; Davis, S.A.; Mann, S. Promotion of fluorapatite crystallization by soluble-matrix proteins from Lingula anatina shells. Angew. Chem. Int. Ed. 2004, 43, 885–888. [Google Scholar] [CrossRef] [PubMed]
- Fricke, M.; Volkmer, D. Crystallization of calcium carbonate beneath insoluble monolayers: Suitable models of mineral-matrix interactions in biomineralization? Top. Curr. Chem. 2006, 270, 1–41. [Google Scholar]
- Weiner, S.; Addadi, L. Design strategies in mineralized biological materials. J. Mater. Chem. 1997, 7, 689–702. [Google Scholar] [CrossRef]
- Weiss, I.M.; Tuross, N.; Addadi, L.; Weiner, S. Mollusc larval shell formation: Amorphous calcium carbonate is a precursor phase for aragonite. J. Exp. Zool. Part. A Ecol. Genet. Physiol. 2002, 293, 478–491. [Google Scholar] [CrossRef] [PubMed]
- Oyane, A.; Kim, H.-M.; Furuya, T.; Kokubo, T.; Miyazaki, T.; Nakamura, T. Preparation and assessment of revised simulated body fluids. J. Biomed. Mater. Res. 2003, 65, 188–195. [Google Scholar] [CrossRef] [PubMed]
- Kokubo, T.; Takadama, H. How useful is SBF in predicting in vivo bone bioactivity? Biomaterials 2006, 27, 2907–2915. [Google Scholar] [CrossRef] [PubMed]
- Kokubo, T.; Kushitani, H.; Ebisawa, Y.; Kitsugi, T.; Kotani, S.; Oura, K.; Yamamuro, T. Apatite formation on bioactive ceramics in body environment. Bioceramics 1989, 1, 157–162. [Google Scholar]
- Lu, X.; Leng, Y. Theoretical analysis of calcium phosphate precipitation in simulated body fluid. Biomaterials 2005, 26, 1097–1108. [Google Scholar] [CrossRef] [PubMed]
- Oyane, A.; Onuma, K.; Kokubo, T.; Ito, A. Clustering of Calcium Phosphate in the System CaCl2—H3PO4−KCl−H2O. J. Phys. Chem. B 1999, 103, 8230–8235. [Google Scholar] [CrossRef]
- Kim, H.-M.; Himeno, T.; Kawashita, M.; Kokubo, T.; Nakamura, T. The mechanism of biomineralization of bone-like apatite on synthetic hydroxyapatite: An in vitro assessment. J. R. Soc. Interface 2004, 1, 17–22. [Google Scholar] [CrossRef] [PubMed]
- Rhee, S.H.; Tanaka, J. Effect of citric acid on the nucleation of hydroxyapatite in a simulated body fluid. Biomaterials 1999, 20, 2155–2160. [Google Scholar] [CrossRef]
- Bleek, K.; Taubert, A. New developments in polymer-controlled, bioinspired calcium phosphate mineralization from aqueous solution. Acta Biomater. 2013, 9, 6283–6321. [Google Scholar] [CrossRef] [PubMed]
- Müller, L.; Müller, F.A. Preparation of SBF with different HCO3− content and its influence on the composition of biomimetic apatites. Acta Biomater. 2006, 2, 181–189. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.T.Y.; Leng, Y.; Chow, K.L.; Ren, F.; Ge, X.; Wang, K.; Lu, X. Cell culture medium as an alternative to conventional simulated body fluid. Acta Biomater. 2011, 7, 2615–2622. [Google Scholar] [CrossRef] [PubMed]
- Tao, J.; Pan, H.; Wang, J.; Wu, J.; Wang, B.; Xu, X.; Tang, R. Evolution of amorphous calcium phosphate to hydroxyapatite probed by gold nanoparticles. J. Phys. Chem. C 2008, 112, 14929–14933. [Google Scholar] [CrossRef]
- Blumenthal, N.C.; Posner, A.S. Hydroxyapatite: Mechanism of formation and properties. Calcif. Tissue Int. 1973, 13, 235–243. [Google Scholar] [CrossRef]
- Chen, Y.; Gu, W.; Pan, H.; Jiang, S.; Tang, R. Stabilizing amorphous calcium phosphate phase by citrate adsorption. CrystEngComm 2014, 16, 1864–1867. [Google Scholar] [CrossRef]
- Wang, Y.-N.; Jiang, S.; Pan, H.; Tang, R. Less is more: Silicate in the crystallization of hydroxyapatite in simulated body fluids. CrystEngComm 2016, 18, 379–383. [Google Scholar] [CrossRef]
- Jiang, S.; Jin, W.; Wang, Y.-N.; Pan, H.; Sun, Z.; Tang, R. Effect of the aggregation state of amorphous calcium phosphate on hydroxyapatite nucleation kinetics. RSC Adv. 2017, 7, 25497–25503. [Google Scholar] [CrossRef]
- Kim, S.; Ryu, H.S.; Shin, H.; Jung, H.S.; Hong, K.S. Direct observation of hydroxyapatite nucleation from amorphous phase in a stoichiometric calcium/phosphate aqueous solution. Chem. Lett. 2004, 33, 1292–1293. [Google Scholar] [CrossRef]
- Gattuso, J.-P.; Frankignoulle, M.; Bourge, I.; Romaine, S.; Buddemeier, R.W. Effect of calcium carbonate saturation of seawater on coral calcification. Glob. Planet. Chang. 1998, 18, 37–46. [Google Scholar] [CrossRef]
- Stanley, S.M. Effects of global seawater chemistry on biomineralization: Past, present, and future. Chem. Rev. 2008, 108, 4483–4498. [Google Scholar] [CrossRef] [PubMed]
- Ries, J.B.; Stanley, S.M.; Hardie, L.A. Scleractinian corals produce calcite, and grow more slowly, in artificial Cretaceous seawater. Geology 2006, 34, 525–528. [Google Scholar] [CrossRef]
- Reddy, M.M.; Nancollas, G.H. The crystallization of calcium carbonate: IV. The effect of magnesium, strontium and sulfate ions. J. Cryst. Growth 1976, 35, 33–38. [Google Scholar] [CrossRef]
- Berner, R.A. The role of magnesium in the crystal growth of calcite and aragonite from sea water. Geochim. Cosmochim. Acta 1975, 39, 489IN3495–494504. [Google Scholar] [CrossRef]
- Mucci, A.; Morse, J.W. The incorporation of Mg2+ and Sr2+ into calcite overgrowths: Influences of growth rate and solution composition. Geochim. Cosmochim. Acta 1983, 47, 217–233. [Google Scholar] [CrossRef]
- Stanley, S.M. Influence of seawater chemistry on biomineralization throughout Phanerozoic time: Paleontological and experimental evidence. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2006, 232, 214–236. [Google Scholar] [CrossRef]
- Purgstaller, B.; Konrad, F.; Dietzel, M.; Immenhauser, A.; Mavromatis, V. Control of Mg2+/Ca2+ activity ratio on the formation of crystalline carbonate minerals via an amorphous precursor. Cryst. Growth Des. 2017, 17, 1069–1078. [Google Scholar] [CrossRef]
- Shen, X.; Belcher, A.M.; Hansma, P.K.; Stucky, G.D.; Morse, D.E. Molecular cloning and characterization of lustrin A, a matrix protein from shell and pearl nacre of Haliotis rufescens. J. Biol. Chem. 1997, 272, 32472–32481. [Google Scholar] [CrossRef] [PubMed]
- Pereira-Mouriès, L.; Almeida, M.; Ribeiro, C.; Peduzzi, J.; Barthélemy, M.; Milet, C.; Lopez, E. Soluble silk-like organic matrix in the nacreous layer of the bivalve Pinctada maxima. FEBS J. 2002, 269, 4994–5003. [Google Scholar] [CrossRef]
- Time-Lapse, S.C.S.; Franz, C.M.; Muller, D.J. Atomic force microscopy in biomedical research. Methods 2011, 736, 97–107. [Google Scholar]
- Orgel, J.P.R.O.; Irving, T.C.; Miller, A.; Wess, T.J. Microfibrillar structure of type I collagen in situ. Proc. Natl. Acad. Sci. USA 2006, 103, 9001–9005. [Google Scholar] [CrossRef] [PubMed]
- Perumal, S.; Antipova, O.; Orgel, J.P.R.O. Collagen fibril architecture, domain organization, and triple-helical conformation govern its proteolysis. Proc. Natl. Acad. Sci. USA 2008, 105, 2824–2829. [Google Scholar] [CrossRef] [PubMed]
- Shoulders, M.D.; Raines, R.T. Collagen structure and stability. Annu. Rev. Biochem. 2009, 78, 929–958. [Google Scholar] [CrossRef] [PubMed]
- Addadi, L.; Weiner, S. Interactions between acidic proteins and crystals: Stereochemical requirements in biomineralization. Proc. Natl. Acad. Sci. USA 1985, 82, 4110–4114. [Google Scholar] [CrossRef] [PubMed]
- Mann, S.; Archibald, D.D.; Didymus, J.M.; Douglas, T.; Heywood, B.R.; Meldrum, F.C.; Reeves, N.J. Organic Interfaces: Biominerals. Science 1993, 261, 1286–1292. [Google Scholar] [CrossRef] [PubMed]
- Berman, A.; Ahn, D.J.; Lio, A.; Salmeron, M.; Reichert, A.; Charych, D. Total alignment of calcite at acidic polydiacetylene films: Cooperativity at the organic-inorganic Interface. Science 1995, 269, 515–518. [Google Scholar] [CrossRef] [PubMed]
- Aizenberg, J.; Black, A.J.; Whitesides, G.M. Oriented growth of calcite controlled by self-assembled monolayers of functionalized alkanethiols supported on gold and silver. J. Am. Chem. Soc. 1999, 121, 4500–4509. [Google Scholar] [CrossRef]
- Travaille, A.M.; Kaptijn, L.; Verwer, P.; Hulsken, B.; Elemans, J.A.A.W.; Nolte, R.J.M.; Van Kempen, H. Highly oriented self-assembled monolayers as templates for epitaxial calcite growth. J. Am. Chem. Soc. 2003, 125, 11571–11577. [Google Scholar] [CrossRef] [PubMed]
- Duffy, D.M.; Harding, J.H. Simulation of organic monolayers as templates for the nucleation of calcite crystals. Langmuir 2004, 20, 7630–7636. [Google Scholar] [CrossRef] [PubMed]
- DiMasi, E.; Olszta, M.J.; Patel, V.M.; Gower, L.B. When is template directed mineralization really template directed? CrystEngComm 2003, 5, 346. [Google Scholar] [CrossRef]
- Gehrke, N.; Nassif, N.; Pinna, N.; Antonietti, M.; Gupta, H.S.; Cölfen, H. Retrosynthesis of nacre via amorphous precursor particles. Chem. Mater. 2005, 17, 6514–6516. [Google Scholar] [CrossRef]
- Nudelman, F.; Pieterse, K.; George, A.; Bomans, P.H.H.; Friedrich, H.; Brylka, L.J.; Hilbers, P.A.J.; de With, G.; Sommerdijk, N.A.J.M. The role of collagen in bone apatite formation in the presence of hydroxyapatite nucleation inhibitors. Nat. Mater. 2010, 9, 1004–1009. [Google Scholar] [CrossRef] [PubMed]
- Niu, L.; Jee, S.E.; Jiao, K.; Tonggu, L.; Li, M.; Wang, L.; Yang, Y.; Bian, J.; Breschi, L.; Jang, S.S.; et al. Collagen intrafibrillar mineralization as a result of the balance between osmotic equilibrium and electroneutrality. Nat. Mater. 2016, 16, 370–378. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Chen, Y.; Li, L.; Sun, J.; Gu, X.; Xu, X.; Pan, H.; Tang, R. Remineralization of dentin collagen by meta-stabilized amorphous calcium phosphate. CrystEngComm 2013, 15, 6151–6158. [Google Scholar] [CrossRef]
- Wang, Y.; Azaïs, T.; Robin, M.; Vallée, A.; Catania, C.; Legriel, P.; Pehau-Arnaudet, G.; Babonneau, F.; Giraud-Guille, M.-M.; Nassif, N. The predominant role of collagen in the nucleation, growth, structure and orientation of bone apatite. Nat. Mater. 2012, 11, 724–733. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Kim, Y.K.; Dai, L.; Li, N.; Khan, S.O.; Pashley, D.H.; Tay, F.R. Hierarchical and non-hierarchical mineralisation of collagen. Biomaterials 2011, 32, 1291–1300. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Li, N.; Qi, Y.P.; Dai, L.; Bryan, T.E.; Mao, J.; Pashley, D.H.; Tay, F.R. Intrafibrillar collagen mineralization produced by biomimetic hierarchical nanoapatite assembly. Adv. Mater. 2011, 23, 975–980. [Google Scholar] [CrossRef] [PubMed]
- Dai, L.; Qi, Y.P.; Niu, L.N.; Liu, Y.; Pucci, C.R.; Looney, S.W.; Ling, J.Q.; Pashley, D.H.; Tay, F.R. Inorganic-organic nanocomposite assembly using collagen as a template and sodium tripolyphosphate as a biomimetic analog of matrix phosphoprotein. Cryst. Growth Des. 2011, 11, 3504–3511. [Google Scholar] [CrossRef] [PubMed]
- Niu, L.N.; Jiao, K.; Ryou, H.; Yiu, C.K.Y.; Chen, J.H.; Breschi, L.; Arola, D.D.; Pashley, D.H.; Tay, F.R. Multiphase intrafibrillar mineralization of collagen. Angew. Chem.—Int. Ed. 2013, 52, 5762–5766. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Luo, X.; Niu, L.; Yang, H.; Yiu, C.K.Y.; Wang, T.; Zhou, L.; Mao, J.; Huang, C.; Pashley, D.H.; et al. Biomimetic intrafibrillar mineralization of type i collagen with intermediate precursors-loaded mesoporous carriers. Sci. Rep. 2015, 5, 11199. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, J.; Chen, C.; Pan, H.; Chen, Y.; Mao, C.; Wang, W.; Tang, R.; Gu, X. Biomimetic promotion of dentin remineralization using l-glutamic acid: Inspiration from biomineralization proteins. J. Mater. Chem. B 2014, 2, 4544–4553. [Google Scholar] [CrossRef]
- Chen, C.; Mao, C.; Sun, J.; Chen, Y.; Wang, W.; Pan, H.; Tang, R.; Gu, X. Glutaraldehyde-induced remineralization improves the mechanical properties and biostability of dentin collagen. Mater. Sci. Eng. C 2016, 67, 657–665. [Google Scholar] [CrossRef] [PubMed]
- Ubukata, T. Architectural, constraints on the morphogenesis of prismatic structure in bivalvia. Palaeontology 1994, 37, 241–261. [Google Scholar]
- Checa, A.G.; Rodriguez-Navarro, A. Geometrical and crystallographic constraints determine the self-organization of shell microstructures in Unionidae (Bivalvia: Mollusca). Proc. R. Soc. B Biol. Sci. 2001, 268, 771–778. [Google Scholar] [CrossRef] [PubMed]
- Gilbert, P.; Metzler, R.A.; Zhou, D.; Scholl, A.; Doran, A.; Young, A.; Kunz, M.; Tamura, N.; Coppersmith, S.N. Gradual ordering in red abalone nacre. J. Am. Chem. Soc. 2008, 130, 17519–17527. [Google Scholar] [CrossRef] [PubMed]
- Ping, H.; Xie, H.; Wan, Y.; Zhang, Z.; Zhang, J.; Xiang, M.; Xie, J.; Wang, H.; Wang, W.; Fu, Z. Confinement controlled mineralization of calcium carbonate within collagen fibrils. J. Mater. Chem. B 2016, 4, 880–886. [Google Scholar] [CrossRef]
- Cantaert, B.; Beniash, E.; Meldrum, F.C. Nanoscale confinement controls the crystallization of calcium phosphate: Relevance to bone formation. Chemistry 2013, 19, 14918–14924. [Google Scholar] [CrossRef] [PubMed]
- Xiao, C.; Li, M.; Wang, B.; Liu, M.-F.; Shao, C.; Pan, H.; Lu, Y.; Xu, B.-B.; Li, S.; Zhan, D.; et al. Total morphosynthesis of biomimetic prismatic-type CaCO3 thin films. Nat. Commun. 2017, 8, 1398. [Google Scholar] [CrossRef] [PubMed]
- Gower, L.B.; Odom, D.J. Deposition of calcium carbonate films by a polymer-induced liquid-precursor (PILP) process. J. Cryst. Growth 2000, 210, 719–734. [Google Scholar] [CrossRef]
- Wei, H.; Ma, N.; Shi, F.; Wang, Z.; Zhang, X. Artificial nacre by alternating preparation of layer-by-layer polymer films and CaCO3 strata. Chem. Mater. 2007, 19, 1974–1978. [Google Scholar] [CrossRef]
- Finnemore, A.; Cunha, P.; Shean, T.; Vignolini, S.; Guldin, S.; Oyen, M.; Steiner, U. Biomimetic layer-by-layer assembly of artificial nacre. Nat. Commun. 2012, 3, 966. [Google Scholar] [CrossRef] [PubMed]
- Mao, L.-B.; Gao, H.-L.; Yao, H.-B.; Liu, L.; Colfen, H.; Liu, G.; Chen, S.-M.; Li, S.-K.; Yan, Y.-X.; Liu, Y.-Y.; et al. Synthetic nacre by predesigned matrix-directed mineralization. Science 2016, 354, 107–110. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Han, J.T.; Cho, K. Formation of amorphous calcium carbonate thin films and their role in biomineralization. Chem. Mater. 2004, 16, 1740–1746. [Google Scholar] [CrossRef]
- Jiang, J.; Gao, M.-R.; Qiu, Y.-H.; Wang, G.-S.; Liu, L.; Cai, G.-B.; Yu, S.-H. Confined crystallization of polycrystalline high-magnesium calcite from compact Mg-ACC precursor tablets and its biological implications. CrystEngComm 2011, 13, 952–956. [Google Scholar] [CrossRef]
- Rajasekharan, A.K.; Andersson, M. Role of nanoscale confinement on calcium phosphate formation at high supersaturation. Cryst. Growth Des. 2015, 15, 2775–2780. [Google Scholar] [CrossRef]
- Loste, E.; Meldrum, F.C. Control of calcium carbonate morphology by transformation of an amorphous precursor in a constrained volume. Chem. Commun. 2001, 10, 901–902. [Google Scholar] [CrossRef]
- Park, R.J.; Meldrum, F.C. Synthesis of single crystals of calcite with complex morphologies. Adv. Mater. 2002, 14, 1167–1169. [Google Scholar] [CrossRef]
- Loste, E.; Park, R.J.; Warren, J.; Meldrum, F.C. Precipitation of calcium carbonate in confinement. Adv. Funct. Mater. 2004, 14, 1211–1220. [Google Scholar] [CrossRef]
- Stephens, C.J.; Ladden, S.F.; Meldrum, F.C.; Christenson, H.K. Amorphous calcium carbonate is stabilized in confinement. Adv. Funct. Mater. 2010, 20, 2108–2115. [Google Scholar] [CrossRef]
- He, G.; Dahl, T.; Veis, A.; George, A. Nucleation of apatite crystals in vitro by self-assembled dentin matrix protein 1. Nat. Mater. 2003, 2, 552–558. [Google Scholar] [CrossRef] [PubMed]
- Cölfen, H. Biomineralization: A crystal-clear view. Nat. Mater. 2010, 9, 960–961. [Google Scholar] [CrossRef] [PubMed]
- Dimasi, E.; Kwak, S.Y.; Amos, F.F.; Olszta, M.J.; Lush, D.; Gower, L.B. Complementary control by additives of the kinetics of amorphous CaCO3 mineralization at an organic interface: In-situ synchrotron X-ray observations. Phys. Rev. Lett. 2006, 97, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Cheng, X.; Varona, P.L.; Olszta, M.J.; Gower, L.B. Biomimetic synthesis of calcite films by a polymer-induced liquid-precursor (PILP) process. 1. Influence and incorporation of magnesium. J. Cryst. Growth 2007, 307, 395–404. [Google Scholar] [CrossRef]
- Homeijer, S.J.; Olszta, M.J.; Barrett, R.A.; Gower, L.B. Growth of nanofibrous barium carbonate on calcium carbonate seeds. J. Cryst. Growth 2008, 310, 2938–2945. [Google Scholar] [CrossRef]
- Olszta, M.J.; Gajjeraman, S.; Kaufman, M.; Gower, L.B. Nanofibrous calcite synthesized via a solution–precursor–solid mechanism. Chem. Mater. 2004, 16, 2355–2362. [Google Scholar] [CrossRef]
- Bergström, L.; Sturm, E.V.; Salazar-Alvarez, G.; Cölfen, H. Mesocrystals in biominerals and colloidal arrays. Acc. Chem. Res. 2015, 48, 1391–1402. [Google Scholar]
- Revealed, I.; Yuwono, V.M.; Burrows, N.D.; Soltis, J.A.; Lee Penn, R. Oriented aggregation: Formation and transformation of mesocrystal. J. Am. Chem. Soc. 2010, 132, 2163–2165. [Google Scholar]
- Niederberger, M.; Cölfen, H. Oriented attachment and mesocrystals: Non-classical crystallization mechanisms based on nanoparticle assembly. Phys. Chem. Chem. Phys. 2006, 8, 3271–3287. [Google Scholar] [CrossRef] [PubMed]
- Cölfen, H.; Antonietti, M. Mesocrystals: Inorganic superstructures made by highly parallel crystallization and controlled alignment. Angew. Chem.—Int. Ed. 2005, 44, 5576–5591. [Google Scholar] [CrossRef] [PubMed]
- Long, X.; Ma, Y.; Cho, K.R.; Li, D.; De Yoreo, J.J.; Qi, L. Oriented calcite micropillars and prisms formed through aggregation and recrystallization of poly(acrylic acid) stabilized nanoparticles. Cryst. Growth Des. 2013, 13, 3856–3863. [Google Scholar] [CrossRef]
- Agthe, M.; Plivelic, T.S.; Labrador, A.; Bergström, L.; Salazar-Alvarez, G. Following in real time the two-step assembly of nanoparticles into mesocrystals in levitating drops. Nano Lett. 2016, 16, 6838–6843. [Google Scholar] [CrossRef] [PubMed]
- Zhu, G.; Yao, S.; Zhai, H.; Liu, Z.; Li, Y.; Pan, H.; Tang, R. Evolution from classical to non-classical aggregation-based crystal growth of calcite by organic additive control. Langmuir 2016, 32, 8999–9004. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.-Y.; Schenk, A.S.; Ihli, J.; Kulak, A.N.; Hetherington, N.B.J.; Tang, C.C.; Schmahl, W.W.; Griesshaber, E.; Hyett, G.; Meldrum, F.C. A critical analysis of calcium carbonate mesocrystals. Nat. Commun. 2014, 5, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Tao, J.; Pan, H.; Zeng, Y.; Xu, R.; Tang, R. Roles of amorphous calcium phosphate and biological additives in the assembly of hydroxyapatite nanoparticles. J. Phys. Chem. B 2007, 111, 13410–13418. [Google Scholar] [CrossRef] [PubMed]
- Zhai, H.; Jiang, W.; Tao, J.; Lin, S.; Chu, X.; Xu, X.; Tang, R. Self-assembled organic-inorganic hybrid elastic crystal via biomimetic mineralization. Adv. Mater. 2010, 22, 3729–3734. [Google Scholar] [CrossRef] [PubMed]
- Zhai, H.; Quan, Y.; Li, L.; Liu, X.-Y.; Xu, X.; Tang, R. Spontaneously amplified homochiral organic–inorganic nano-helix complexes via self-proliferation. Nanoscale 2013, 5, 3006–3012. [Google Scholar] [CrossRef] [PubMed]
- Tao, J.; Zhou, D.; Zhang, Z.; Xu, X.; Tang, R. Magnesium-aspartate-based crystallization switch inspired from shell molt of crustacean. Proc. Natl. Acad. Sci. USA 2009, 106, 22096–22101. [Google Scholar] [CrossRef] [PubMed]
- Orme, C.A.; Noy, A; Wierzbicki, A; McBride, M.T.; Grantham, M.; Teng, H.H.; Dove, P.M.; DeYoreo, J.J. Formation of chiral morphologies through selective binding of amino acids to calcite surface steps. Nature 2001, 411, 775–779. [Google Scholar] [CrossRef] [PubMed]
- Chung, J.; Granja, I.; Taylor, M.G.; Mpourmpakis, G.; Asplin, J.R.; Rimer, J.D. Molecular modifiers reveal a mechanism of pathological crystal growth inhibition. Nature 2016, 536, 446–450. [Google Scholar] [CrossRef] [PubMed]
- De Yoreo, J.J.; Zepeda-Ruiz, L.A.; Friddle, R.W.; Qiu, S.R.; Wasylenki, L.E.; Chernov, A.A.; Gilmer, G.H.; Dove, P.M. Rethinking classical crystal growth models through molecular scale insights: Consequences of kink-limited kinetics. Cryst. Growth Des. 2009, 9, 5135–5144. [Google Scholar] [CrossRef]
- Yang, X.; Xie, B.; Wang, L.; Qin, Y.; Henneman, Z.J.; Nancollas, G.H. Influence of magnesium ions and amino acids on the nucleation and growth of hydroxyapatite. CrystEngComm 2011, 13, 1153–1158. [Google Scholar] [CrossRef]
- Song, R.-Q.; Cölfen, H. Additive controlled crystallization. CrystEngComm 2011, 13, 1249. [Google Scholar] [CrossRef]
- Qiu, S.R.; Orme, C. A Dynamics of biomineral formation at the near molecular level. Chem. Rev. 2008, 108, 4784–4822. [Google Scholar] [CrossRef] [PubMed]
- Giocondi, J.L.; El-Dasher, B.S.; Nancollas, G.H.; Orme, C.A. Molecular mechanisms of crystallization impacting calcium phosphate cements. Philos. Trans. R. Soc. A Math. Phys. Eng. Sci. 2010, 368, 1937–1961. [Google Scholar] [CrossRef] [PubMed]
- Sikirić, M.D.; Füredi-Milhofer, H. The influence of surface active molecules on the crystallization of biominerals in solution. Adv. Colloid Interface Sci. 2006, 128–130, 135–158. [Google Scholar] [CrossRef] [PubMed]
- Nancollas, G.H.; Wu, W. Biomineralization mechanisms: A kinetics and interfacial energy approach. J. Cryst. Growth 2000, 211, 137–142. [Google Scholar] [CrossRef]
- Liu, X.Y. From solid-fluid interfacial structure to nucleation kinetics: principles and strategies for micro/nanostructure engineering. In Nanoscale Structure and Assembly at Solid-Fluid Interfaces: Interfacial Structures versus Dynamics; Liu, X.Y., De Yoreo, J., Eds.; Springer: London, UK, 2004; Volume 1, pp. 109–175. ISBN 978-1-4020-7810-1. [Google Scholar]
- Demichelis, R.; Raiteri, P.; Gale, J.D.; Quigley, D.; Gebauer, D. Stable prenucleation mineral clusters are liquid-like ionic polymers. Nat. Commun. 2011, 2, 590. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Y.-T.; Johnson, W.L. Disordered Materials: A Survey of Amorphous Solids. Science 1987, 235, 997–1002. [Google Scholar] [CrossRef] [PubMed]
- Betts, F.; Posner, A. Structural model for amorphous calcium phosphate. Trans. Am. Crystallogr. Assoc. 1974, 10, 73–84. [Google Scholar]
- Betts, F.; Posner, A.S. An X-ray radial distribution study of amorphous calcium phosphate. Mater. Res. Bull. 1974, 9, 353–360. [Google Scholar] [CrossRef]
- Treboux, G.; Layrolle, P.; Kanzaki, N.; Onuma, K.; Ito, A. Existence of Posner’s Cluster in Vacuum. J. Phys. Chem. A 2000, 104, 5111–5114. [Google Scholar] [CrossRef]
- Kanzaki, N.; Treboux, G.; Onuma, K.; Tsutsumi, S.; Ito, A. Calcium phosphate clusters. Biomaterials 2001, 22, 2921–2929. [Google Scholar] [CrossRef]
- Dorozhkin, S.V. Calcium orthophosphates. J. Mater. Sci. 2007, 42, 1061–1095. [Google Scholar] [CrossRef]
- Sedlak, J.M.; Beebe, R.A. Temperature programmed dehydration of amorphous calcium phosphate. J. Colloid Interface Sci. 1974, 47, 483–489. [Google Scholar] [CrossRef]
- Kojima, Y.; Sakama, K.; Toyama, T.; Yasue, T.; Arai, Y. Dehydration of water molecules in amorphous calcium phosphate. Phosphorus Res. Bull. 1994, 4, 47–52. [Google Scholar] [CrossRef]
- Eanes, E.D.; Posner, A. Division of biophysics: Kinetics and mechanism of conversion of noncrystalline calcium phosphate to crystalline hydroxyapatite. Trans. N. Y. Acad. Sci. 1965, 28, 233–241. [Google Scholar] [CrossRef]
- Blumenthal, N.C.; Posner, A.S.; Holmes, J.M. Effect of preparation conditions on the properties and transformation of amorphous calcium phosphate. Mater. Res. Bull. 1972, 7, 1181–1189. [Google Scholar] [CrossRef]
- Termine, J.D.; Posner, A.S. Infra-red determination of the percentage of crystallinity in apatitic calcium phosphates. Nature 1966, 211, 268–270. [Google Scholar] [CrossRef] [PubMed]
- Harries, J.E.; Hukins, D.W.L.; Holt, C.; Hasnain, S.S. Conversion of amorphous calcium phosphate into hydroxyapatite investigated by EXAFS spectroscopy. J. Cryst. Growth 1987, 84, 563–570. [Google Scholar] [CrossRef]
- Feng, C.F.; Khor, K.A.; Kweh, S.W.K.; Cheang, P. Thermally induced crystallization of amorphous calcium phosphate in plasma-spheroidised hydroxyapatite powders. Mater. Lett. 2000, 46, 229–233. [Google Scholar] [CrossRef]
- Jaeger, C.; Maltsev, S.K.A. Progress of structural elucidation of amorphous calcium phosphate (ACP) and hydroxyapatite (HAP): Disorder and surfaces as seen by solid state NMR. Key Eng. Mater. 2006, 69–72. [Google Scholar] [CrossRef]
- Rodriguez-Blanco, J.D.; Shaw, S.; Benning, L.G. The kinetics and mechanisms of amorphous calcium carbonate (ACC) crystallization to calcite, viavaterite. Nanoscale 2011, 3, 265–271. [Google Scholar] [CrossRef] [PubMed]
- Hu, Q.; Nielsen, M.H.; Freeman, C.L.; Hamm, L.M.; Tao, J.; Lee, J.R.I.; Han, T.Y.J.; Becker, U.; Harding, J.H.; Dove, P.M.; et al. The thermodynamics of calcite nucleation at organic interfaces: Classical vs. non-classical pathways. Faraday Discuss. 2012, 159, 509–523. [Google Scholar] [CrossRef]
- Hamm, L.M.; Giuffre, A.J.; Han, N.; Tao, J.; Wang, D.; De Yoreo, J.J.; Dove, P.M. Reconciling disparate views of template-directed nucleation through measurement of calcite nucleation kinetics and binding energies. Proc. Natl. Acad. Sci. USA 2014, 111, 1304–1309. [Google Scholar] [CrossRef] [PubMed]
- Jahromi, M.T.; Yao, G.; Cerruti, M. The importance of amino acid interactions in the crystallization of hydroxyapatite. J. R. Soc. Interface 2013, 10, 20120906. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Xu, X.; Tang, R. Improvement of Biological Organisms Using Functional Material Shells. Adv. Funct. Mater. 2016, 26, 1862–1880. [Google Scholar] [CrossRef]
- Satyanarayana, T.; Prakash, A.; Johri, B.N. Microorganisms in environmental management: Microbes and environment. In Microorg. Environ. Manag. Microbes Environ; Springer Science & Business Media Press: Kluwer, The Netherlands, 2012. [Google Scholar]
- Smeets, P.J.M.; Cho, K.R.; Kempen, R.G.E.; Sommerdijk, N.A.J.M.; De Yoreo, J.J. Calcium carbonate nucleation driven by ion binding in a biomimetic matrix revealed by in situ electron microscopy. Nat. Mater. 2015, 14, 394–399. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Han, J.T.; Kim, D.H.; Cho, K. Two modes of transformation of amorphous calcium carbonate films in air. J. Phys. Chem. B 2006, 110, 2764–2770. [Google Scholar] [CrossRef] [PubMed]
- Ihli, J.; Kulak, A.N.; Meldrum, F.C. Freeze-drying yields stable and pure amorphous calcium carbonate (ACC). Chem. Commun. 2013, 49, 3134–3136. [Google Scholar] [CrossRef] [PubMed]
- De Yoreo, J. Research Methods in Biomineralization Science; Elsevier Press: Amsterdam, The Netherlands, 2013. [Google Scholar]
- Stephenson, A.E.; DeYoreo, J.J.; Wu, L.; Wu, K.J.; Hoyer, J.; Dove, P.M. Peptides enhance magnesium signature in calcite: Insights into origins of vital effects. Science 2008, 322, 724–727. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Csetenyi, L.; Gadd, G.M. Biomineralization of metal carbonates by Neurospora crassa. Environ. Sci. Technol. 2014, 48, 14409–14416. [Google Scholar] [CrossRef] [PubMed]
- Cai, G.-B.; Zhao, G.-X.; Wang, X.-K.; Yu, S.-H. Synthesis of polyacrylic acid stabilized amorphous calcium carbonate nanoparticles and their application for removal of toxic heavy metal ions in water. J. Phys. Chem. C 2010, 114, 12948–12954. [Google Scholar] [CrossRef]
- Xiong, W.; Tang, Y.; Shao, C.; Zhao, Y.; Jin, B.; Huang, T.; Miao, Y.; Shu, L.; Ma, W.; Xu, X. Prevention of cyanobacterial blooms using nanosilica: A biomineralization-inspired strategy. Environ. Sci. Technol. 2017, 51, 12717–12726. [Google Scholar] [CrossRef] [PubMed]
- Nam, K.T.; Kim, D.-W.; Yoo, P.J.; Chiang, C.-Y.; Meethong, N.; Hammond, P.T.; Chiang, Y.-M.; Belcher, A.M. Virus-enabled synthesis and assembly of nanowires for lithium ion battery electrodes. Science 2006, 312, 885–888. [Google Scholar] [CrossRef] [PubMed]
- Uchiyama, H.; Hosono, E.; Zhou, H.; Imai, H. Three-dimensional architectures of spinel-type LiMn2O4 prepared from biomimetic porous carbonates and their application to a cathode for lithium-ion batteries. J. Mater. Chem. 2009, 19, 4012–4016. [Google Scholar] [CrossRef]
- Ryu, J.; Kim, S.W.; Kang, K.; Park, C.B. Mineralization of self-assembled peptide nanofibers for rechargeable lithium ion batteries. Adv. Mater. 2010, 22, 5537–5541. [Google Scholar] [CrossRef] [PubMed]
- Xiong, W.; Zhao, X.; Zhu, G.; Shao, C.; Li, Y.; Ma, W.; Xu, X.; Tang, R. Silicification-induced cell aggregation for the sustainable production of H2 under aerobic conditions. Angew. Chem. 2015, 127, 12129–12133. [Google Scholar] [CrossRef]
- Yao, S.; Jin, B.; Liu, Z.; Shao, C.; Zhao, R.; Wang, X.; Tang, R. Biomineralization: From material tactics to biological strategy. Adv. Mater. 2017, 29. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Sun, C.; Li, P.; Wu, T.; Zhou, H.; Yang, D.; Liu, Y.; Ma, X.; Song, Z.; Nian, Q.; et al. Vaccine engineering with dual-functional mineral shell: A promising strategy to overcome preexisting immunity. Adv. Mater. 2016, 28, 694–700. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Li, X.; Mo, L.; Song, Z.; Chen, W.; Deng, Y.; Zhao, H.; Qin, E.; Qin, C.; Tang, R. Eggshell-inspired biomineralization generates vaccines that do not require refrigeration. Angew. Chem.–Int. Ed. 2012, 51, 10576–10579. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Liu, P.; Liu, Z.; Pan, H.; Xu, X.; Tang, R. Biomimetic construction of cellular shell by adjusting the interfacial energy. Biotechnol. Bioeng. 2014, 111, 386–395. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Phase | Formula | Solubility a -log(Ks) |
|---|---|---|
| Dicalcium phosphate dihydrate (DCPD) b | CaHPO4•2H2O | 6.59 |
| Octacalcium phosphate (OCP) b | Ca8(HPO4)2(PO4)4•5H2O | 96.6 |
| α-Tricalcium phosphate (α-TCP) b | α-Ca3(PO4)2 | 25.5 |
| β-Tricalcium phosphate (β-TCP) b | β-Ca3(PO4)2 | 28.9 |
| Hydroxyapatite (HAP) b | Ca10(PO4)6(OH)2 | 116.8 |
| Vaterite c | CaCO3 | 7.91 |
| Aragonite c | CaCO3 | 8.34 |
| Calcite c | CaCO3 | 8.48 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jin, W.; Jiang, S.; Pan, H.; Tang, R. Amorphous Phase Mediated Crystallization: Fundamentals of Biomineralization. Crystals 2018, 8, 48. https://doi.org/10.3390/cryst8010048
Jin W, Jiang S, Pan H, Tang R. Amorphous Phase Mediated Crystallization: Fundamentals of Biomineralization. Crystals. 2018; 8(1):48. https://doi.org/10.3390/cryst8010048
Chicago/Turabian StyleJin, Wenjing, Shuqin Jiang, Haihua Pan, and Ruikang Tang. 2018. "Amorphous Phase Mediated Crystallization: Fundamentals of Biomineralization" Crystals 8, no. 1: 48. https://doi.org/10.3390/cryst8010048