
Clinical Potential of YY1-Hypoxia Axis for Vascular Normalization and to Improve Immunotherapy
Department of Precision Medicine, School of Medicine, University of Campania “Luigi Vanvitelli”, 80138 Naples, Italy
*
Author to whom correspondence should be addressed.
Cancers 2024, 16(3), 491; https://doi.org/10.3390/cancers16030491
Submission received: 8 December 2023
/
Revised: 12 January 2024
/
Accepted: 19 January 2024
/
Published: 23 January 2024
(This article belongs to the Special Issue Pleiotropic Pro-tumor Activities Regulated by Overexpressed Yin Yang 1 (YY1) in Human Cancers: Clinical Implications)
Abstract
:Simple Summary
Solid tumors create a hostile hypoxic microenvironment characterized by abnormal vascularization and immune suppression. Target tumor vessels and the activation of immunostimulatory cells have shown promising results in suppressing tumor growth, leading to the development of a variety of strategies. In this review, we dissect the concept of vessel and immune normalization and discuss the YY1-Hypoxia axis as a potential target.
Abstract
Abnormal vasculature in solid tumors causes poor blood perfusion, hypoxia, low pH, and immune evasion. It also shapes the tumor microenvironment and affects response to immunotherapy. The combination of antiangiogenic therapy and immunotherapy has emerged as a promising approach to normalize vasculature and unlock the full potential of immunotherapy. However, the unpredictable and redundant mechanisms of vascularization and immune suppression triggered by tumor-specific hypoxic microenvironments indicate that such combination therapies need to be further evaluated to improve patient outcomes. Here, we provide an overview of the interplay between tumor angiogenesis and immune modulation and review the function and mechanism of the YY1-HIF axis that regulates the vascular and immune tumor microenvironment. Furthermore, we discuss the potential of targeting YY1 and other strategies, such as nanocarrier delivery systems and engineered immune cells (CAR-T), to normalize tumor vascularization and re-establish an immune-permissive microenvironment to enhance the efficacy of cancer therapy.
1. Introduction
In solid tumors, a hypoxic microenvironment is one of the most common pathogenic conditions that promotes the formation of new vessels and favors immune escape [1]. Neovascularization is considered a milestone in tumor growth as it is the key mechanism supporting the proliferation of cancer cell clones and their dissemination to distant sites. The sprouting of existing vessels is thought to be the main initial event through which angiogenic growth factors promote new vessel formation from pre-existing vessels. However, several other vasculogenic mechanisms strongly associated with tumor hypoxia have been recognized, including intussusceptions, co-option, and vasculogenic mimicry [2,3,4]. Consequently, the vasculature within tumors is heterogeneous, abnormal, dysfunctional, and hyperpermeable. This condition determines insufficient nutrient and oxygen perfusion in selected tumor areas, such as the core far from the vessel, or in the high-proliferation cohort of cells. Under reduced oxygen tension, tumor cells adapt to hypoxia and activate several survival pathways, including hypoxia factor 1α (HIF-1α) and vascular endothelial growth factor (VEGF), able to generate a vicious cycle with immune cells.
VEGF, a key driver of angiogenesis, has also been demonstrated to be an immunosuppressive factor, interfering with the maturation of dendritic cells (DC) and suppressing T cell priming [5]. Moreover, cell components of the hypoxic tumor microenvironment (TME) increase chemokine secretion, including stromal cell-derived factor 1-α (SDF1-α) and C-C Motif Chemokine Ligand 28 (CCL28), and recruit regulatory T helper cells (Tregs), a heterogeneous population of immature myeloid cells (MDSCs), and M2-type macrophages. These immune cells contribute to an immunosuppressive microenvironment and promote abnormal vessel growth. In particular, M2-like macrophages secrete pro-angiogenic factors, mainly VEGF, which promote the development of excessive and immature vessels [6,7]. Therefore, such a hostile microenvironment also limits CD8+ T and CD4+ Th1 cell infiltration [8,9,10] (Figure 1). Consequently, strategies to induce a TME permissive to immune cell infiltration are of major interest to improve the efficacy of immune therapy and reduce tumor growth. However, therapeutic targeting of vascularization mechanisms has proven to be inefficient and, in some instances, exacerbates tumor progression [11,12]. This unexpected outcome, together with better knowledge of the contribution of immune cells to tumor vascularization mechanisms, has led to the concept of vascular normalization [13]. Its goal is to modulate abnormal vascular development, favoring blood perfusion within the tumor and opening the way to immunotherapy and other drugs [14,15]. Recent research has used various therapeutic approaches, ranging from optimal doses of antiangiogenic drugs alone or in combination with immune checkpoint inhibitors (ICIs) [16]. However, data emerging from trials have revealed the poor efficacy of this treatment because multiple mechanisms of immune suppression and vasculature heterogeneity, independent of VEGF, coexist in tumors. For clinical applications, additional exploration of hypoxia-response pathways is warranted to optimize vascular normalization, improve antitumor immunity, and avoid tumor resistance. In the following section, we describe the current vascular and immune normalization strategies and highlight the clinical potential of the HIF-YY1 signaling axis.
2. Vascular and Immune Crosstalk Normalization
Antiangiogenic therapy, initially developed to eliminate tumor blood vessels, results in increased hypoxia and tumor aggressiveness [11]. Growing evidence has demonstrated a complicated interconnection between vessels and immune cells, in which pro-angiogenic molecules have an immunosuppressive role and immune cells promote the proliferation of regulatory T cells and vessel immaturity. These observations indicate the need for therapeutic strategies to normalize the tumor vascular blood supply and immune modulate the TME permitting the infiltration of activated immune effector cells [17,18].
Therefore, ICI treatment administered concomitantly with antiangiogenic drugs has been shown to be more effective than single [13,17,19]. Indeed, targeting both the immune microenvironment and tumor vessels has been approved by the FDA for the treatment of liver, kidney, lung, and uterine cancer [20]. However, the efficacy of these combinatorial regimes differs between cancer types and is not free of adverse effects [21]. Trials have indicated that the duration of treatment, sequencing of therapies, and tumor-specific hypoxia pathways influence efficacy. All these aspects need to be better evaluated to improve the treatment benefits and patient outcomes. The future of antiangiogenic regimens seems to consist of tumor-specific treatments to normalize vascular and immune-cross talk and novel immune-cell technologies [21] (Figure 2).
3. Vascular Signaling by YY1
3.1. HIF-YY1 Vascular Signaling Axis
A plethora of mechanisms triggered by hypoxia affect tumor vascularization. Hypoxia is a hallmark of the TME of various cancer types, generated by the activation and stabilization of hypoxia-inducible factor-1 (HIF-1) [22], and its signaling is well known to promote the angiogenic switch [23]. Overexpression of HIF-1α has been shown to be closely associated with poor prognosis, increasing tumor growth, vascularization, and metastasis [15]. The general mechanism of HIF-1α activity includes its heterodimerization with p300 and the formation of a transcriptional activation complex. This can recognize the HRE-DNA site and activate the transcription of target genes, such as VEGF, angiopoietin, fibroblast growth factor 2 (FGF2), and platelet-derived growth factor (PDGFB) [16]. HIF-1α also regulates genes involved in cell survival and metabolism, such as phosphoglycerate kinase (PGK), carbonic anhydrase 9 (CA9), Bcl-2 interacting protein 3 (BNIP3), and glucose transporter 1 (GLUT1), all of which contribute to tumor progression [24]. Many binding partners can stabilize HIF-1α on specific promoters of genes, including the Yin Yang 1 protein (YY1) protein. This is a multifunctional zinc finger protein, a member of the GLI-Kruppel family, and is part of the polycomb complex [25]. The expression and function of YY1 have been reported in both normal and cancerous cells [26]. YY1 may activate or deactivate gene expression, depending on the binding partner, functional interactions with corepressors or activators, and chromatin status, resulting in different phenotypic effects [26,27,28]. YY1 is highly expressed in numerous cancer types, and increased levels correlate with poor clinical outcomes. Increased expression of YY1 was reported in several cancer types, including prostate cancer [29], osteosarcoma [30], breast cancer [31], glioma and meningioma [32], gastrointestinal cancer [33], pancreatic ductal adenocarcinoma [34], melanoma [35], and hepatocellular carcinoma (HCC) [36]. In many types of cancer, oncogenic tumor signaling triggered by YY1 involves the activation of the c-Myc oncogene [37], AKT [38], and inhibition of tumor suppression by p53 [39,40].
YY1 also shapes the TME under hypoxia, thereby promoting neoangiogenesis. Data demonstrated that YY1 cooperates with HIF-1α as a binding partner on the VEGF promoter, stabilizing HIF-1 transcriptional activity and increasing VEGF expression [41,42,43,44].
YY1 also regulates HIF-1α post-translationally because their interaction prevents proteasomal degradation of HIF-1α in hypoxic conditions [45]. This was confirmed by the observation that under hypoxia, YY1 knockdown disrupts the stability of HIF-1α and decreases the expression of target genes (e.g., VEGF, Transforming Growth Factor (TGF-α), PGK, CA9, BNIP3, and GLUT1) in a p53-independent manner [43]. Consistent with this, YY1 silencing impairing tumor vascularization reduced cancer growth and metastatic lung colonization in p53-deficient tumors [43]. Indeed, it has been demonstrated that p53 is negatively regulated by YY1, which facilitates direct binding to E3 ligase mouse double minute 2 (MDM2), thereby enhancing its ubiquitination and proteasomal degradation [39].
In tumor hypoxia, many chemokines can be released, including C-X-C motif chemokine 4 (CXCR4), which affects angiogenesis, immune surveillance, and the ability of tumor cells to metastasize [46]. Studies on osteosarcoma have shown that downregulation of YY1 negatively regulates the VEGF/CXCR4 axis pathway, thereby inhibiting angiogenesis and tumor cell migration by reducing the transcription and activity of MMPs [41,42]. These findings suggest that YY1 is a target in tumor therapy independent of p53 status. This is important because p53 is frequently mutated or downregulated in tumor cells. Figure 3 illustrates the angiogenic mechanism mediated by YY1.
3.2. YY1 Promotes Tumor Angiogenesis in HIF-1α-Independent Manners
YY1 may promote tumor angiogenesis in a HIF-1-independent manner via the DEK oncogene [47]. The oncoprotein DEK, a non-histone chromosomal factor, is overexpressed in various cancers, such as lung and prostate cancers [48,49], and is correlated with poor clinical outcomes [50]. DEK has been implicated in multiple cellular processes, including transcriptional regulation, mRNA processing, chromatin remodeling [51,52], cell proliferation, differentiation, and apoptosis [53,54]. It was originally identified as a fusion protein with the CAN nucleoporin in a subtype of acute myeloid leukemias [55]. Zhang et al. demonstrated a novel function of DEK following transcriptional activation via YY1 [47]. In the model proposed, DEK enhanced VEGF transcription by directly binding its responsive element (DRE) to the VEGF promoter or indirectly by stabilizing HIF-1α [56]. The authors demonstrated that DEK physically interacts with HIF-1α and p300, forming a complex with the VEGF promoter in breast cancer cells [47].
Tumor angiogenic mechanisms can be triggered also by proinflammatory cytokines (such as IL-8) [57], chemokines such as CXC motif chemokine ligand 1 (CXCL1) [58], or thrombospondin-1 (TSP-1) and TSP-2 [59] via sonic hedgehog (Shh) signaling. In lung cancer carrying Kras mutations, it has been demonstrated that the YY1 transcription factor is a crucial mediator between the mutated isoform of Kras and Shh signaling [60]. When Kras mutations were present, YY1 promoted neovascularization by increasing the ZNF322A expression that activated Shh pro-angiogenic signaling. The same group identified the Kras/YY1/ZNF322A/Shh transcriptional axis as part of an important mechanism underlying neo-angiogenesis and migration of lung cancer cells in in vitro/vivo models [60].
YY1 in endothelial cells (EC) is important for vascular development and angiogenesis. Its knockout determines vascular deficiency and embryonal lethality. Mechanistically, YY1 functions as a modulator protein of Notch signaling, controlling vascular development and EC functions, such as proliferation, migration, lumen formation, and vessel stability [61,62]. In particular, YY1 directly interacts with the N-terminal domain of the recombination signal binding protein for the Ig Kappa J region (RBPJ), competing with the Notch coactivator mastermind-like protein 1 (MAML1) for binding to RBPJ, and thereby impairs formation of the NICD/MAML1/RBPJ complex [63]. YY1 could therefore be a molecular target for the treatment of angiogenesis-related diseases (Figure 3).
3.3. YY1-Related Non-Coding RNAs (ncRNAs) in Angiogenic Mechanisms
In malignancies and under chronic hypoxia, YY1 also acts as the critical interface between epigenetic code and noncoding-RNA regulatory mechanism [45].
In some tumors, e.g., acute leukemia, YY1 acting as a transcription factor promotes the overexpression of small nucleolar RNA host gene 5 (SNHG5), which complements and binds miR-26b and reduces angiogenesis [64]. Reciprocal regulation between YY1 and miRNAs has also been reported. In acute myeloid leukemia, YY1 negatively regulates let-7a, which inhibits the expression of the anti-apoptotic protein B-cell lymphoma-extra-large (BCL-xL) [65]; instead, miR-7 may silence YY1 and KLF4 mRNAs, contributing to chemoresistance [66]. Several classes of RNAs, such as long non-coding RNA and circ-RNAs, are under the transcriptional control of YY1. In lung cancer, YY1 overexpresses the non-coding RNA MCM3AP-AS1, which promotes cancer progression by binding to and degradation of miR-340-5p, a negative regulator of angiogenesis [67]. However, YY1 can also be regulated by circ-RNAs. In particular, in cholangiocarcinoma (CCA), circ-CCAC1 promotes CCA progression and angiogenesis by degradation (sponging) miR-514a-5p, a negative regulator of YY1. Collectively, these data indicate that the circ-CCAC1/miR-514a-5p/YY1/CAMLG axis plays an important role in angiogenesis [68] (Figure 3).
Thus, many studies have confirmed that YY1 and non-coding RNAs are involved in a complex crosstalk that influences tumor progression. However, these mechanisms need to be further elucidated in order to fully understand their role and importance in malignance.
4. Immune Cells Regulate Tumor Angiogenesis
A complex regulatory network between the TME and innate and adaptive immune cells influences the pro and antiangiogenic phenotype of immune cells [10], in particular tumor-associated macrophages (TAM). Macrophages with the M1 phenotype suppress endothelial sprouting and can induce vessel maturation by secreting TNFα [69]. In contrast, macrophages with the M2 phenotype and TAMs linking angiopoietin-2 promote angiogenesis [70]. Consistently, the knockout of macrophages expressing Tie-2 (the angiopoietin receptor) induces normalization of the tumor vasculature [71]. CD11+ immune-suppressing myeloid-derived cells from the macrophage lineage may promote angiogenesis, forming vessel-like tubuli containing CD31 antigens (endothelial cell phenotype) or by their integration into the tumor vasculature [72]. Adaptive immune cells and CD8+ T cells also influence the EC characteristics and myeloid cell phenotypes. In particular, CD8+ cells secreting IFN-γ inhibit the proliferation and migration of ECs [73], stimulate vascular maturation by recruiting pericytes [73], and reprogram TAMs from the M2- to M1-like phenotype [74]. In addition, CD4+ T helper 1 (TH1) cells promote vessel normalization by secreting additional IFN-γ. Indeed, selective depletion of CD4+ TH1 cells increased vessel malformation, whereas CD4+ T cell activation improved vessel normalization [75]. However, in many solid tumors, hypoxic conditions result in ineffective vascularization and the expression of immune inhibitory antigens, determining resistance to ICI therapy [76]. In particular, the activation of HIF-1α stimulates both VEGF-A and inhibitory checkpoints, such as the PD-1/PDL1 pathways, which favor immunosuppressive phenotypes [77]. Additionally, hypoxia factor in fibroblast-associated cancer cell populations (CAFs) positively regulates the expression of transforming growth factor-beta (TGF-β), which promotes the differentiation and proliferation of Tregs with pro-tumorigenic myeloid cell phenotypes [78].
Normalization of immune cell profiles within the TME is a promising aim of immunotherapy.
5. YY1 in B and T Cells
A correlation between YY1 and the immunosuppressive TME has been reported in several tumor types [27,34]. Moreover, studies have reported YY1 as a regulator of B and T cell development, activation, differentiation, and immune function [79,80] (Figure 3).
B cell development and function depend on the YY1 downstream transcriptional program [80,81,82]. Deletion of the YY1 gene prevents the transition of pro-B cells into pre-B cells at early developmental stages [79,80].
Furthermore, YY1 was identified as a controller of nuclear deaminase protein levels (AID), an enzyme required for class switch recombination (CSR) and somatic hypermutation (SHM), the processes responsible for antibody maturation, and the expression of different immunoglobulin isotypes [80,81,83]. Increased levels of YY1 are correlated with poor survival prognosis in patients with diffuse large B-cell lymphoma (DLBCL) [83,84], high-grade DLBCL, or Burkitt’s lymphoma [85], suggesting an oncogenic function of YY1 in human B-cell lymphoma genesis [86]. Indeed, YY1 binds to the Kruppel-like factor 4 (KLF4) promoter, inducing proliferation and differentiation of B-cell neoplasm in B-NHL [87].
Notch signaling is an important pathway regulating binary cell-fate choices at crucial checkpoints, including T-cell- versus B-cell-specific gene expression, αβ versus γδ T-cell-receptor expression, and CD4+ versus CD8+ lineage decisions. During physiological T cell development, YY1 has been shown to regulate Notch1 expression by binding to the enhancer locus to promote signaling in T cells. YY1-deficient T cells have reduced Notch1 expression and signaling [88]. However, in the context of an immune suppressive tumor microenvironment, YY1 regulates the proliferation and function of Tregs through remodeling of the Foxp3 locus. Targeting YY1 could be useful to modulate Treg cell proliferation and the beneficial functions of CD8+ T cells [89].
On the other hand, YY1 seems to promote T cell exhaustion, a phenomenon that affects CD8+ T cells, in which persistent antigenic stimulation renders the cells hyporesponsive and incapable of eliminating tumor cells. Prolonged T cell activation upregulates both YY1 and EZH2 proteins that repress IL-2, a cytokine involved in their capability to kill tumor cells [90]. Another mechanism by which YY1 is involved is tumor resistance to ICI immunotherapy, achieved through the positive regulation of programmed death receptor-1 (PD-1)/and LAG3. The mechanism describes the activation of CD8+ T cells expressing PD-1 [91] and the binding to the ligand PD-L1 on different cells of the TME, promoting survival pathway instead of apoptosis [92,93]. The presence of PD-1-positive T cells and YY1 protein was correlated with disease progression in melanoma [90,94] and immune resistance to ICIs in lung adenocarcinoma [95]. Therefore, YY1 regulates several signaling pathways, including the inhibition of p53 and miR34a, the activation of the PI3K/Akt/mTOR pathway, as well as c-Myc and COX-2 [96] involved in tumor resistance to ICIs.
6. Preclinical Model and Clinical Findings of Vascular Normalization and Immune Modulation
Combinations of immune and antiangiogenic therapies have been tested in several preclinical models. In pancreatic neuroendocrine tumor (RT2-PNET), mammary carcinoma (MMTV-PyMT), and glioblastoma (NFpp10-GBM) [97] mouse models, therapy based on anti-VEGFR2 and anti-PD-L1 normalized tumor vessels, favoring the intertumoral infiltration of activated T cells. On the other hand, antiangiogenic therapy was also able to reduce the TOX-mediated T cell exhaustion program in the TME, influencing the proliferation of cytotoxic T lymphocytes [98].
The most successful preclinical results have been reported in renal cell carcinoma (RCC) and hepatic cell carcinoma (HCC), which showed improved anticancer immunity and overall survival in a mouse model (OS) [99,100]. Additional promising preclinical efficacy was also demonstrated by the combination of ANGPT2/Tie2 inhibitors with anti-VEGF and anti-PD-1 therapy in both genetically engineered and transplant tumor mouse models [101]. Recently, encouraging data were provided by a multicenter, open-label, multicohort phase Ib/II KEYNOTE-146 trial (NCT02501096) [102], which reported an increase ORR of 39.6–45.3% at 24 and 48 weeks, respectively, after lenvatinib and pembrolizumab (tyrosine kinase inhibitor and anti-PD-1, respectively) treatment in endometrial cancer. In addition to the above-mentioned trials, numerous studies on various malignancies have demonstrated the efficacy of combining PD-1/PD-L1 inhibitors and anti-VEGF agents [16,103]. Nonetheless, some challenges remain, as the effective dose, timing, and duration of treatment need to be established, but studies evaluating the effects of different strategies are beginning to emerge [104].
7. Novel Strategies: Targeted Therapy
An alternative strategy to normalize tumor vessels and improve immunotherapy efficacy is based on novel systems to deliver nitric oxide (NO), which regulates angiogenesis, using nanocarriers as a vehicle [105]. The NO approach is of great value not only because NO is a physiological molecule but also because nitrosylation is a regulative post-translational modification of proteins. In particular, NO may inhibit YY1 function. Indeed, DETA-NONOate, a NO donor inducing S-nitrosylation of YY1, inhibits its DNA-binding activity and triggers Fas to induce apoptosis [106,107]. DETA-NONOate is still in the early stages of clinical development, but its combination with cisplatin-induced apoptosis in otherwise therapy-resistant prostate cancer cell lines [106]. Additionally, NO donors could be useful in improving the efficacy of immunotherapy by reducing YY1 activity and Treg proliferation.
Hypoxia is an integral part of the TME, and HIF-1 pathways play a central role in tumor growth by affecting multiple interconnected mechanisms, rendering a challenge to target HIF-1. In this context, given the central role of YY1 in the stability of HIF-1α on the VEGF promoter, it is conceivable that YY1 targeting might offer an innovative therapeutic approach to normalize vascularization [108]. In this regard, it is tempting to speculate that targeting tumor YY1 may normalize tumor perfusion, favoring the recruitment of effector T cells and antitumoral immunotherapy. Given that it is possible to directly target hypoxia by using HIF-1 inhibitors, one can further stimulate the production of pro-angiogenic factors and cytokines to foster vascular endothelial cell proliferation [109]. Destabilizing HIF binding and reducing YY1 expression could be alternative possibilities [109,110,111,112].
Another approach to reduce YY1 mRNA and target tumor angiogenesis could be the use of RNA interference. As demonstrated in lung tumors, MiR-29a downregulates DNA methyltransferases (DNMT) 3A and 3B and suppresses YY1 mRNA [113,114]. MiR-186 reduces YY1 expression in lung and prostate tumor cells by binding to sequences at the 3′ UTR region of YY1 mRNA, leading to decreased cell migration and invasion [115]. Several other miRNAs, as well as regulation mediated by lncRNA [116], can target YY1 in various cancer types; however, although promising results have been obtained from preclinical studies, there are currently no clinical trials.
Preclinical studies using gene editing techniques have recently yielded promising results by inhibiting YY1. Xu et al. used CRISPR/Cas9 to downregulate YY1 in prostate cancer. Lowering YY1 expression reduced tumor cell metabolism and promoted apoptosis of prostate cancer cells [117]. Above all, YY1-targeted therapy has good potential in combination with immunotherapy to improve patient response rates and resistance compared to immunotherapy alone.
An alternative approach to improve the immune response is to target the immune microenvironment. Recent advances have been achieved by CAR T cell therapy [118]. This approach involves: 1-the the molecular modification of human T cells, introducing synthetic receptors capable of recognizing selected antigens as VEGF receptor 2 or tumor-associated antigens (TAAs), 2-autologous infusion of CAR-T cells in patients where they can recognize and kill cells expressing selected antigens [119,120]. This personalized approach could address the heterogeneity of vessel origin targeting with CAR T cell antigen-specific cells. Additionally, the improvement in vessel perfusion instead of vessel destruction could be helpful in recruiting a stable immune response against the tumor. CAR T-cell therapy has shown promising results in lymphoma, inducing durable antitumor immune responses, but has exhibited low efficacy in solid tumors due to limited capability to infiltrate the TME [121]. Nevertheless, considering the overall advantages and effectiveness of CAR T-cell therapy, many studies have evaluated their use in combination with vascular normalization treatment [122], as reported in Table 1.
8. Conclusions
Resistance to antiangiogenic therapies is mainly attributable to the complex interactions between vessels, immune cells, and cellular components of the microenvironment that allow tumor growth [134]. It has been proven that HIF-1 and protein kinase B (AKT) are capable of driving tumor growth despite mTOR and VEGF inhibition [135]. HIF-1-driven neovascularization can also be initiated independently of VEGF and thus be resistant to angiogenesis inhibitors [136]. One mechanism of VEGF tyrosine kinase inhibitor resistance is through the direct selection of cell subpopulations that can rapidly upregulate alternative pro-angiogenic pathways [137]. Several studies in RCCs and HCCs demonstrated that following treatment with tyrosine kinase inhibitors such as sunitinib [138] and sorafenib [139], these tumors increase the release of pro-angiogenic cytokines to foster vascular endothelial cell proliferation [140].
In addition, tumor hypoxia, generated by blocking angiogenesis, activates resistance to additional target receptors, such as epidermal growth factor receptor (EGFR) inhibitors [141] in non-small cell lung cancer and in a pancreatic cancer model [140]. With regard to the immune normalization of the TME, recent data from single-cell omics have demonstrated their diversity. Therefore, it is not surprising that immune cells showed different functions in different tumor types. However, as mentioned above, targeting vessels influences immune components and vice versa. Strategies combining vessel and immune normalization may yield greater efficacy, and studies are currently evaluating the effects of different combinations and dosages. In this context, YY1 may be an opportunity to prevent HIF function, inducing longer-term vascular normalization and enhancing the trafficking of T cells and immunotherapy efficiency. However, a better exploration of YY1-hypoxia-signaling pathways for clinical application, particularly in solid tumors, will be essential [142]. Nevertheless, YY1-targeting is a promising antitumor strategy, particularly in combination with other treatments.
Author Contributions
Conceptualization, F.d.N.; writing, resources, and data curation, C.M. and F.d.N. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Folkman, J. Tumor angiogenesis: Therapeutic implications. N. Engl. J. Med. 1971, 285, 1182–1186. [Google Scholar]
- Welti, J.; Loges, S.; Dimmeler, S.; Carmeliet, P. Recent molecular discoveries in angiogenesis and antiangiogenic therapies in cancer. J. Clin. Invest. 2013, 123, 3190–3200. [Google Scholar] [CrossRef]
- Liu, Z.L.; Chen, H.H.; Zheng, L.L.; Sun, L.P.; Shi, L. Angiogenic signaling pathways and anti-angiogenic therapy for cancer. Signal Transduct. Target. Ther. 2023, 8, 198. [Google Scholar] [CrossRef] [PubMed]
- Hanahan, D.; Weinberg, R.A. Hallmarks of cancer: The next generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef]
- Gabrilovich, D.I.; Chen, H.L.; Girgis, K.R.; Cunningham, H.T.; Meny, G.M.; Nadaf, S.; Kavanaugh, D.; Carbone, D.P. Production of vascular endothelial growth factor by human tumors inhibits the functional maturation of dendritic cells. Nat. Med. 1996, 2, 1096–1103. [Google Scholar] [CrossRef]
- Albini, A.; Bruno, A.; Noonan, D.M.; Mortara, L. Contribution to Tumor Angiogenesis From Innate Immune Cells within the Tumor Microenvironment: Implications for Immunotherapy. Front. Immunol. 2018, 9, 527. [Google Scholar] [CrossRef] [PubMed]
- Zhao, C.C.; Han, Q.J.; Ying, H.Y.; Gu, X.X.; Yang, N.; Li, L.Y.; Zhang, Q.Z. TNFSF15 facilitates differentiation and polarization of macrophages toward M1 phenotype to inhibit tumor growth. Oncoimmunology 2022, 11, 2032918. [Google Scholar] [CrossRef] [PubMed]
- Ge, Z.; Zhang, Q.; Lin, W.; Jiang, X.; Zhang, Y. The role of angiogenic growth factors in the immune microenvironment of glioma. Front. Oncol. 2023, 13, 1254694. [Google Scholar] [CrossRef]
- Facciabene, A.; Motz, G.T.; Coukos, G. T-regulatory cells: Key players in tumor immune escape and angiogenesis. Cancer Res. 2012, 72, 2162–2171. [Google Scholar] [CrossRef]
- Bruno, A.; Pagani, A.; Pulze, L.; Albini, A.; Dallaglio, K.; Noonan, D.M.; Mortara, L. Orchestration of angiogenesis by immune cells. Front. Oncol. 2014, 4, 131. [Google Scholar] [CrossRef]
- Cao, Y.; Langer, R.; Ferrara, N. Targeting angiogenesis in oncology, ophthalmology and beyond. Nat. Rev. Drug Discov. 2023, 22, 476–495. [Google Scholar] [CrossRef]
- Jayson, G.C.; Kerbel, R.; Ellis, L.M.; Harris, A.L. Antiangiogenic therapy in oncology: Current status and future directions. Lancet 2016, 388, 518–529. [Google Scholar] [CrossRef]
- Jain, R.K. Normalizing tumor vasculature with anti-angiogenic therapy: A new paradigm for combination therapy. Nat. Med. 2001, 7, 987–989. [Google Scholar] [CrossRef]
- Fang, J.; Lu, Y.; Zheng, J.; Jiang, X.; Shen, H.; Shang, X.; Lu, Y.; Fu, P. Exploring the crosstalk between endothelial cells, immune cells, and immune checkpoints in the tumor microenvironment: New insights and therapeutic implications. Cell Death Dis. 2023, 14, 586. [Google Scholar] [CrossRef]
- Fan, P.; Zhang, N.; Candi, E.; Agostini, M.; Piacentini, M.; Centre, T.O.R.; Shi, Y.; Huang, Y.; Melino, G. Alleviating hypoxia to improve cancer immunotherapy. Oncogene 2023, 42, 3591–3604. [Google Scholar] [CrossRef]
- Choi, Y.; Jung, K. Normalization of the tumor microenvironment by harnessing vascular and immune modulation to achieve enhanced cancer therapy. Exp. Mol. Med. 2023, 55, 2308–2319. [Google Scholar] [CrossRef]
- Binnewies, M.; Roberts, E.W.; Kersten, K.; Chan, V.; Fearon, D.F.; Merad, M.; Coussens, L.M.; Gabrilovich, D.I.; Ostrand-Rosenberg, S.; Hedrick, C.C.; et al. Understanding the tumor immune microenvironment (TIME) for effective therapy. Nat. Med. 2018, 24, 541–550. [Google Scholar] [CrossRef]
- Fukumura, D.; Kloepper, J.; Amoozgar, Z.; Duda, D.G.; Jain, R.K. Enhancing cancer immunotherapy using antiangiogenics: Opportunities and challenges. Nat. Rev. Clin. Oncol. 2018, 15, 325–340. [Google Scholar] [CrossRef] [PubMed]
- Mpekris, F.; Voutouri, C.; Baish, J.W.; Duda, D.G.; Munn, L.L.; Stylianopoulos, T.; Jain, R.K. Combining microenvironment normalization strategies to improve cancer immunotherapy. Proc. Natl. Acad. Sci. USA 2020, 117, 3728–3737. [Google Scholar] [CrossRef]
- Lee, W.S.; Yang, H.; Chon, H.J.; Kim, C. Combination of anti-angiogenic therapy and immune checkpoint blockade normalizes vascular-immune crosstalk to potentiate cancer immunity. Exp. Mol. Med. 2020, 52, 1475–1485. [Google Scholar] [CrossRef]
- Patel, S.A.; Nilsson, M.B.; Le, X.; Cascone, T.; Jain, R.K.; Heymach, J.V. Molecular Mechanisms and Future Implications of VEGF/VEGFR in Cancer Therapy. Clin. Cancer Res. 2023, 29, 30–39. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.W.; Bae, S.H.; Jeong, J.W.; Kim, S.H.; Kim, K.W. Hypoxia-inducible factor (HIF-1)alpha: Its protein stability and biological functions. Exp. Mol. Med. 2004, 36, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Hashimoto, T.; Shibasaki, F. Hypoxia-inducible factor as an angiogenic master switch. Front. Pediatr. 2015, 3, 33. [Google Scholar] [CrossRef]
- Infantino, V.; Santarsiero, A.; Convertini, P.; Todisco, S.; Iacobazzi, V. Cancer Cell Metabolism in Hypoxia: Role of HIF-1 as Key Regulator and Therapeutic Target. Int. J. Mol. Sci. 2021, 22, 5703. [Google Scholar] [CrossRef] [PubMed]
- Thomas, M.J.; Seto, E. Unlocking the mechanisms of transcription factor YY1: Are chromatin modifying enzymes the key? Gene 1999, 236, 197–208. [Google Scholar] [CrossRef] [PubMed]
- Gordon, S.; Akopyan, G.; Garban, H.; Bonavida, B. Transcription factor YY1: Structure, function, and therapeutic implications in cancer biology. Oncogene 2006, 25, 1125–1142. [Google Scholar] [CrossRef]
- Nicholson, S.; Whitehouse, H.; Naidoo, K.; Byers, R.J. Yin Yang 1 in human cancer. Crit. Rev. Oncog. 2011, 16, 245–260. [Google Scholar] [CrossRef] [PubMed]
- Weintraub, A.S.; Li, C.H.; Zamudio, A.V.; Sigova, A.A.; Hannett, N.M.; Day, D.S.; Abraham, B.J.; Cohen, M.A.; Nabet, B.; Buckley, D.L.; et al. YY1 Is a Structural Regulator of Enhancer-Promoter Loops. Cell 2017, 171, 1573–1588.e28. [Google Scholar] [CrossRef]
- Seligson, D.; Horvath, S.; Huerta-Yepez, S.; Hanna, S.; Garban, H.; Roberts, A.; Shi, T.; Liu, X.; Chia, D.; Goodglick, L.; et al. Expression of transcription factor Yin Yang 1 in prostate cancer. Int. J. Oncol. 2005, 27, 131–141. [Google Scholar] [CrossRef] [PubMed]
- De Nigris, F.; Botti, C.; de Chiara, A.; Rossiello, R.; Apice, G.; Fazioli, F.; Fiorito, C.; Sica, V.; Napoli, C. Expression of transcription factor Yin Yang 1 in human osteosarcomas. Eur. J. Cancer 2006, 42, 2420–2424. [Google Scholar] [CrossRef]
- Allouche, A.; Nolens, G.; Tancredi, A.; Delacroix, L.; Mardaga, J.; Fridman, V.; Winkler, R.; Boniver, J.; Delvenne, P.; Begon, D.Y. The combined immunodetection of AP-2alpha and YY1 transcription factors is associated with ERBB2 gene overexpression in primary breast tumors. Breast Cancer Res. 2008, 10, R9. [Google Scholar] [CrossRef] [PubMed]
- Baritaki, S.; Chatzinikola, A.M.; Vakis, A.F.; Soulitzis, N.; Karabetsos, D.A.; Neonakis, I.; Bonavida, B.; Spandidos, D.A. YY1 Over-expression in human brain gliomas and meningiomas correlates with TGF-beta1, IGF-1 and FGF-2 mRNA levels. Cancer Invest. 2009, 27, 184–192. [Google Scholar] [CrossRef] [PubMed]
- Chinnappan, D.; Xiao, D.; Ratnasari, A.; Andry, C.; King, T.C.; Weber, H.C. Transcription factor YY1 expression in human gastrointestinal cancer cells. Int. J. Oncol. 2009, 34, 1417–1423. [Google Scholar] [PubMed]
- Zhang, J.J.; Zhu, Y.; Xie, K.L.; Peng, Y.P.; Tao, J.Q.; Tang, J.; Li, Z.; Xu, Z.K.; Dai, C.C.; Qian, Z.Y.; et al. Yin Yang-1 suppresses invasion and metastasis of pancreatic ductal adenocarcinoma by downregulating MMP10 in a MUC4/ErbB2/p38/MEF2C-dependent mechanism. Mol. Cancer 2014, 13, 130. [Google Scholar] [CrossRef] [PubMed]
- Zhao, G.; Li, Q.; Wang, A.; Jiao, J. YY1 regulates melanoma tumorigenesis through a miR-9~RYBP axis. J. Exp. Clin. Cancer Res. 2015, 34, 66. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Li, D.; Sui, G. YY1 Is an Inducer of Cancer Metastasis. Crit. Rev. Oncog. 2017, 22, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Riggs, K.J.; Saleque, S.; Wong, K.K.; Merrell, K.T.; Lee, J.S.; Shi, Y.; Calame, K. Yin-yang 1 activates the c-myc promoter. Mol. Cell. Biol. 1993, 13, 7487–7495. [Google Scholar] [PubMed]
- Zhang, Q.; Wan, M.; Shi, J.; Horita, D.A.; Miller, L.D.; Kute, T.E.; Kridel, S.J.; Kulik, G.; Sui, G. Yin Yang 1 promotes mTORC2-mediated AKT phosphorylation. J. Mol. Cell Biol. 2016, 8, 232–243. [Google Scholar] [CrossRef] [PubMed]
- Sui, G.; Affar, B.; Shi, Y.; Brignone, C.; Wall, N.R.; Yin, P.; Donohoe, M.; Luke, M.P.; Calvo, D.; Grossman, S.R.; et al. Yin Yang 1 is a negative regulator of p53. Cell 2004, 117, 859–872. [Google Scholar] [CrossRef]
- Gronroos, E.; Terentiev, A.A.; Punga, T.; Ericsson, J. YY1 inhibits the activation of the p53 tumor suppressor in response to genotoxic stress. Proc. Natl. Acad. Sci. USA 2004, 101, 12165–12170. [Google Scholar] [CrossRef]
- De Nigris, F.; Rossiello, R.; Schiano, C.; Arra, C.; Williams-Ignarro, S.; Barbieri, A.; Lanza, A.; Balestrieri, A.; Giuliano, M.T.; Ignarro, L.J.; et al. Deletion of Yin Yang 1 protein in osteosarcoma cells on cell invasion and CXCR4/angiogenesis and metastasis. Cancer Res. 2008, 68, 1797–1808. [Google Scholar] [CrossRef] [PubMed]
- De Nigris, F.; Crudele, V.; Giovane, A.; Casamassimi, A.; Giordano, A.; Garban, H.J.; Cacciatore, F.; Pentimalli, F.; Marquez-Garban, D.C.; Petrillo, A.; et al. CXCR4/YY1 inhibition impairs VEGF network and angiogenesis during malignancy. Proc. Natl. Acad. Sci. USA 2010, 107, 14484–14489. [Google Scholar] [CrossRef]
- Wu, S.; Kasim, V.; Kano, M.R.; Tanaka, S.; Ohba, S.; Miura, Y.; Miyata, K.; Liu, X.; Matsuhashi, A.; Chung, U.I.; et al. Transcription factor YY1 contributes to tumor growth by stabilizing hypoxia factor HIF-1alpha in a p53-independent manner. Cancer Res. 2013, 73, 1787–1799. [Google Scholar] [CrossRef]
- Yang, W.; Li, Z.; Qin, R.; Wang, X.; An, H.; Wang, Y.; Zhu, Y.; Liu, Y.; Cai, S.; Chen, S.; et al. YY1 Promotes Endothelial Cell-Dependent Tumor Angiogenesis in Hepatocellular Carcinoma by Transcriptionally Activating VEGFA. Front. Oncol. 2019, 9, 1187. [Google Scholar] [CrossRef]
- Infante, T.; Mancini, F.P.; Lanza, A.; Soricelli, A.; de Nigris, F.; Napoli, C. Polycomb YY1 is a critical interface between epigenetic code and miRNA machinery after exposure to hypoxia in malignancy. Biochim. Biophys. Acta 2015, 1853, 975–986. [Google Scholar] [CrossRef]
- Korbecki, J.; Kojder, K.; Kapczuk, P.; Kupnicka, P.; Gawronska-Szklarz, B.; Gutowska, I.; Chlubek, D.; Baranowska-Bosiacka, I. The Effect of Hypoxia on the Expression of CXC Chemokines and CXC Chemokine Receptors—A Review of Literature. Int. J. Mol. Sci. 2021, 22, 843. [Google Scholar] [CrossRef]
- Zhang, Y.; Liu, J.; Wang, S.; Luo, X.; Li, Y.; Lv, Z.; Zhu, J.; Lin, J.; Ding, L.; Ye, Q. The DEK oncogene activates VEGF expression and promotes tumor angiogenesis and growth in HIF-1alpha-dependent and -independent manners. Oncotarget 2016, 7, 23740–23756. [Google Scholar] [CrossRef]
- Han, S.; Xuan, Y.; Liu, S.; Zhang, M.; Jin, D.; Jin, R.; Lin, Z. Clinicopathological significance of DEK overexpression in serous ovarian tumors. Pathol. Int. 2009, 59, 443–447. [Google Scholar] [CrossRef]
- Shibata, T.; Kokubu, A.; Miyamoto, M.; Hosoda, F.; Gotoh, M.; Tsuta, K.; Asamura, H.; Matsuno, Y.; Kondo, T.; Imoto, I.; et al. DEK oncoprotein regulates transcriptional modifiers and sustains tumor initiation activity in high-grade neuroendocrine carcinoma of the lung. Oncogene 2010, 29, 4671–4681. [Google Scholar] [CrossRef]
- Wang, X.; Lin, L.; Ren, X.; Lin, Z.; Li, Z.; Li, C.; Jin, T. High expression of oncoprotein DEK predicts poor prognosis of small cell lung cancer. Int. J. Clin. Exp. Pathol. 2014, 7, 5016–5023. [Google Scholar]
- Deutzmann, A.; Ganz, M.; Schonenberger, F.; Vervoorts, J.; Kappes, F.; Ferrando-May, E. The human oncoprotein and chromatin architectural factor DEK counteracts DNA replication stress. Oncogene 2015, 34, 4270–4277. [Google Scholar] [CrossRef]
- McGarvey, T.; Rosonina, E.; McCracken, S.; Li, Q.; Arnaout, R.; Mientjes, E.; Nickerson, J.A.; Awrey, D.; Greenblatt, J.; Grosveld, G.; et al. The acute myeloid leukemia-associated protein, DEK, forms a splicing-dependent interaction with exon-product complexes. J. Cell Biol. 2000, 150, 309–320. [Google Scholar] [CrossRef]
- Ageberg, M.; Gullberg, U.; Lindmark, A. The involvement of cellular proliferation status in the expression of the human proto-oncogene DEK. Haematologica 2006, 91, 268–269. [Google Scholar]
- Wise-Draper, T.M.; Allen, H.V.; Jones, E.E.; Habash, K.B.; Matsuo, H.; Wells, S.I. Apoptosis inhibition by the human DEK oncoprotein involves interference with p53 functions. Mol. Cell. Biol. 2006, 26, 7506–7519. [Google Scholar] [CrossRef]
- Von Lindern, M.; Fornerod, M.; van Baal, S.; Jaegle, M.; de Wit, T.; Buijs, A.; Grosveld, G. The translocation (6;9), associated with a specific subtype of acute myeloid leukemia, results in the fusion of two genes, dek and can, and the expression of a chimeric, leukemia-specific dek-can mRNA. Mol. Cell. Biol. 1992, 12, 1687–1697. [Google Scholar]
- Sitwala, K.V.; Adams, K.; Markovitz, D.M. YY1 and NF-Y binding sites regulate the transcriptional activity of the dek and dek-can promoter. Oncogene 2002, 21, 8862–8870. [Google Scholar] [CrossRef]
- Sparmann, A.; Bar-Sagi, D. Ras-induced interleukin-8 expression plays a critical role in tumor growth and angiogenesis. Cancer Cell 2004, 6, 447–458. [Google Scholar] [CrossRef]
- Ancrile, B.B.; O’Hayer, K.M.; Counter, C.M. Oncogenic ras-induced expression of cytokines: A new target of anti-cancer therapeutics. Mol. Interv. 2008, 8, 22–27. [Google Scholar] [CrossRef]
- Watnick, R.S.; Cheng, Y.N.; Rangarajan, A.; Ince, T.A.; Weinberg, R.A. Ras modulates Myc activity to repress thrombospondin-1 expression and increase tumor angiogenesis. Cancer Cell 2003, 3, 219–231. [Google Scholar] [CrossRef]
- Lin, C.C.; Kuo, I.Y.; Wu, L.T.; Kuan, W.H.; Liao, S.Y.; Jen, J.; Yang, Y.E.; Tang, C.W.; Chen, Y.R.; Wang, Y.C. Dysregulated Kras/YY1/ZNF322A/Shh transcriptional axis enhances neo-angiogenesis to promote lung cancer progression. Theranostics 2020, 10, 10001–10015. [Google Scholar] [CrossRef]
- Boareto, M.; Jolly, M.K.; Ben-Jacob, E.; Onuchic, J.N. Jagged mediates differences in normal and tumor angiogenesis by affecting tip-stalk fate decision. Proc. Natl. Acad. Sci. USA 2015, 112, E3836–E3844. [Google Scholar] [CrossRef]
- Benedito, R.; Roca, C.; Sorensen, I.; Adams, S.; Gossler, A.; Fruttiger, M.; Adams, R.H. The notch ligands Dll4 and Jagged1 have opposing effects on angiogenesis. Cell 2009, 137, 1124–1135. [Google Scholar] [CrossRef]
- Zhang, S.; Kim, J.Y.; Xu, S.; Liu, H.; Yin, M.; Koroleva, M.; Guo, J.; Pei, X.; Jin, Z.G. Endothelial-specific YY1 governs sprouting angiogenesis through directly interacting with RBPJ. Proc. Natl. Acad. Sci. USA 2020, 117, 4792–4801. [Google Scholar] [CrossRef]
- Li, Z.J.; Cheng, J.; Song, Y.; Li, H.H.; Zheng, J.F. LncRNA SNHG5 upregulation induced by YY1 contributes to angiogenesis via miR-26b/CTGF/VEGFA axis in acute myelogenous leukemia. Lab. Invest. 2021, 101, 341–352. [Google Scholar] [CrossRef]
- Chen, Y.; Jacamo, R.; Konopleva, M.; Garzon, R.; Croce, C.; Andreeff, M. CXCR4 downregulation of let-7a drives chemoresistance in acute myeloid leukemia. J. Clin. Invest. 2013, 123, 2395–2407. [Google Scholar] [CrossRef]
- Morales-Martinez, M.; Vega, G.G.; Neri, N.; Nambo, M.J.; Alvarado, I.; Cuadra, I.; Duran-Padilla, M.A.; Huerta-Yepez, S.; Vega, M.I. MicroRNA-7 Regulates Migration and Chemoresistance in Non-Hodgkin Lymphoma Cells Through Regulation of KLF4 and YY1. Front. Oncol. 2020, 10, 588893. [Google Scholar] [CrossRef]
- Li, X.; Yu, M.; Yang, C. YY1-mediated overexpression of long noncoding RNA MCM3AP-AS1 accelerates angiogenesis and progression in lung cancer by targeting miR-340-5p/KPNA4 axis. J. Cell. Biochem. 2020, 121, 2258–2267. [Google Scholar] [CrossRef]
- Xu, Y.; Leng, K.; Yao, Y.; Kang, P.; Liao, G.; Han, Y.; Shi, G.; Ji, D.; Huang, P.; Zheng, W.; et al. A Circular RNA, Cholangiocarcinoma-Associated Circular RNA 1, Contributes to Cholangiocarcinoma Progression, Induces Angiogenesis, and Disrupts Vascular Endothelial Barriers. Hepatology 2021, 73, 1419–1435. [Google Scholar] [CrossRef]
- Ji, Z.Z.; Chan, M.K.; Chan, A.S.; Leung, K.T.; Jiang, X.; To, K.F.; Wu, Y.; Tang, P.M. Tumour-associated macrophages: Versatile players in the tumour microenvironment. Front. Cell Dev. Biol. 2023, 11, 1261749. [Google Scholar] [CrossRef]
- Qian, B.Z.; Pollard, J.W. Macrophage diversity enhances tumor progression and metastasis. Cell 2010, 141, 39–51. [Google Scholar] [CrossRef]
- De Palma, M.; Murdoch, C.; Venneri, M.A.; Naldini, L.; Lewis, C.E. Tie2-expressing monocytes: Regulation of tumor angiogenesis and therapeutic implications. Trends Immunol. 2007, 28, 519–524. [Google Scholar] [CrossRef]
- Yang, L.; DeBusk, L.M.; Fukuda, K.; Fingleton, B.; Green-Jarvis, B.; Shyr, Y.; Matrisian, L.M.; Carbone, D.P.; Lin, P.C. Expansion of myeloid immune suppressor Gr+CD11b+ cells in tumor-bearing host directly promotes tumor angiogenesis. Cancer Cell 2004, 6, 409–421. [Google Scholar] [CrossRef]
- Kammertoens, T.; Friese, C.; Arina, A.; Idel, C.; Briesemeister, D.; Rothe, M.; Ivanov, A.; Szymborska, A.; Patone, G.; Kunz, S.; et al. Tumour ischaemia by interferon-gamma resembles physiological blood vessel regression. Nature 2017, 545, 98–102. [Google Scholar] [CrossRef]
- Bromley, S.K.; Mempel, T.R.; Luster, A.D. Orchestrating the orchestrators: Chemokines in control of T cell traffic. Nat. Immunol. 2008, 9, 970–980. [Google Scholar] [CrossRef]
- Tian, L.; Goldstein, A.; Wang, H.; Ching Lo, H.; Sun Kim, I.; Welte, T.; Sheng, K.; Dobrolecki, L.E.; Zhang, X.; Putluri, N.; et al. Mutual regulation of tumour vessel normalization and immunostimulatory reprogramming. Nature 2017, 544, 250–254. [Google Scholar] [CrossRef]
- Mosca, L.; de Angelis, A.; Ronchi, A.; De Chiara, A.; Fazioli, F.; Ruosi, C.; Altucci, L.; Conte, M.; de Nigris, F. Sarcoma Common MHC-I Haplotype Restricts Tumor-Specific CD8+ T Cell Response. Cancers 2022, 14, 3414. [Google Scholar] [CrossRef]
- Voron, T.; Colussi, O.; Marcheteau, E.; Pernot, S.; Nizard, M.; Pointet, A.L.; Latreche, S.; Bergaya, S.; Benhamouda, N.; Tanchot, C.; et al. VEGF-A modulates expression of inhibitory checkpoints on CD8+ T cells in tumors. J. Exp. Med. 2015, 212, 139–148. [Google Scholar] [CrossRef]
- Sanmamed, M.F.; Chen, L. A Paradigm Shift in Cancer Immunotherapy: From Enhancement to Normalization. Cell 2018, 175, 313–326. [Google Scholar] [CrossRef]
- Kleiman, E.; Jia, H.; Loguercio, S.; Su, A.I.; Feeney, A.J. YY1 plays an essential role at all stages of B-cell differentiation. Proc. Natl. Acad. Sci. USA 2016, 113, E3911–E3920. [Google Scholar] [CrossRef]
- Banerjee, A.; Sindhava, V.; Vuyyuru, R.; Jha, V.; Hodewadekar, S.; Manser, T.; Atchison, M.L. YY1 Is Required for Germinal Center B Cell Development. PLoS ONE 2016, 11, e0155311. [Google Scholar] [CrossRef]
- Liu, H.; Schmidt-Supprian, M.; Shi, Y.; Hobeika, E.; Barteneva, N.; Jumaa, H.; Pelanda, R.; Reth, M.; Skok, J.; Rajewsky, K.; et al. Yin Yang 1 is a critical regulator of B-cell development. Genes Dev. 2007, 21, 1179–1189. [Google Scholar] [CrossRef] [PubMed]
- Green, M.R.; Monti, S.; Dalla-Favera, R.; Pasqualucci, L.; Walsh, N.C.; Schmidt-Supprian, M.; Kutok, J.L.; Rodig, S.J.; Neuberg, D.S.; Rajewsky, K.; et al. Signatures of murine B-cell development implicate YY1 as a regulator of the germinal center-specific program. Proc. Natl. Acad. Sci. USA 2011, 108, 2873–2878. [Google Scholar] [CrossRef] [PubMed]
- Arribas Arranz, J.; Winter, D.N.; Drexler, H.G.; Eberth, S. Suitability of Yin Yang 1 transcript and protein levels for biomarker studies in B cell non-Hodgkin lymphoma. Biomark Res. 2018, 6, 11. [Google Scholar] [CrossRef]
- Sakhinia, E.; Glennie, C.; Hoyland, J.A.; Menasce, L.P.; Brady, G.; Miller, C.; Radford, J.A.; Byers, R.J. Clinical quantitation of diagnostic and predictive gene expression levels in follicular and diffuse large B-cell lymphoma by RT-PCR gene expression profiling. Blood 2007, 109, 3922–3928. [Google Scholar] [CrossRef]
- Castellano, G.; Torrisi, E.; Ligresti, G.; Nicoletti, F.; Malaponte, G.; Traval, S.; McCubrey, J.A.; Canevari, S.; Libra, M. Yin Yang 1 overexpression in diffuse large B-cell lymphoma is associated with B-cell transformation and tumor progression. Cell Cycle 2010, 9, 557–563. [Google Scholar] [CrossRef]
- Ramkumar, C.; Cui, H.; Kong, Y.; Jones, S.N.; Gerstein, R.M.; Zhang, H. Smurf2 suppresses B-cell proliferation and lymphomagenesis by mediating ubiquitination and degradation of YY1. Nat. Commun. 2013, 4, 2598. [Google Scholar] [CrossRef]
- Morales-Martinez, M.; Valencia-Hipolito, A.; Vega, G.G.; Neri, N.; Nambo, M.J.; Alvarado, I.; Cuadra, I.; Duran-Padilla, M.A.; Martinez-Maza, O.; Huerta-Yepez, S.; et al. Regulation of Kruppel-Like Factor 4 (KLF4) expression through the transcription factor Yin-Yang 1 (YY1) in non-Hodgkin B-cell lymphoma. Oncotarget 2019, 10, 2173–2188. [Google Scholar] [CrossRef] [PubMed]
- Assumpcao, A.; Fu, G.; Singh, D.K.; Lu, Z.; Kuehnl, A.M.; Welch, R.; Ong, I.M.; Wen, R.; Pan, X. A lineage-specific requirement for YY1 Polycomb Group protein function in early T cell development. Development 2021, 148, dev197319. [Google Scholar] [CrossRef]
- Hwang, S.S.; Jang, S.W.; Kim, M.K.; Kim, L.K.; Kim, B.S.; Kim, H.S.; Kim, K.; Lee, W.; Flavell, R.A.; Lee, G.R. YY1 inhibits differentiation and function of regulatory T cells by blocking Foxp3 expression and activity. Nat. Commun. 2016, 7, 10789. [Google Scholar] [CrossRef]
- Balkhi, M.Y.; Wittmann, G.; Xiong, F.; Junghans, R.P. YY1 Upregulates Checkpoint Receptors and Downregulates Type I Cytokines in Exhausted, Chronically Stimulated Human T Cells. iScience 2018, 2, 105–122. [Google Scholar] [CrossRef]
- Li, C.; Jiang, P.; Wei, S.; Xu, X.; Wang, J. Regulatory T cells in tumor microenvironment: New mechanisms, potential therapeutic strategies and future prospects. Mol. Cancer 2020, 19, 116. [Google Scholar] [CrossRef]
- Butterfield, L.H.; Najjar, Y.G. Immunotherapy combination approaches: Mechanisms, biomarkers and clinical observations. Nat. Rev. Immunol. 2023. [Google Scholar] [CrossRef]
- Schiano, C.; Soricelli, A.; De Nigris, F.; Napoli, C. New challenges in integrated diagnosis by imaging and osteo-immunology in bone lesions. Expert Rev. Clin. Immunol. 2019, 15, 289–301. [Google Scholar] [CrossRef]
- Kwiatkowska, D.; Mazur, E.; Reich, A. YY1 Is a Key Player in Melanoma Immunotherapy/Targeted Treatment Resistance. Front. Oncol. 2022, 12, 856963. [Google Scholar] [CrossRef]
- Tang, X.Y.; Xiong, Y.L.; Zhao, Y.B.; Yang, J.; Shi, A.P.; Zheng, K.F.; Liu, Y.J.; Shu, C.; Jiang, T.; Ma, N.; et al. Dual immunological and proliferative regulation of immune checkpoint FGL1 in lung adenocarcinoma: The pivotal role of the YY1-FGL1-MYH9 axis. Front. Immunol. 2022, 13, 1014053. [Google Scholar] [CrossRef]
- Hays, E.; Bonavida, B. YY1 regulates cancer cell immune resistance by modulating PD-L1 expression. Drug Resist. Updates 2019, 43, 10–28. [Google Scholar] [CrossRef]
- Shigeta, K.; Datta, M.; Hato, T.; Kitahara, S.; Chen, I.X.; Matsui, A.; Kikuchi, H.; Mamessier, E.; Aoki, S.; Ramjiawan, R.R.; et al. Dual Programmed Death Receptor-1 and Vascular Endothelial Growth Factor Receptor-2 Blockade Promotes Vascular Normalization and Enhances Antitumor Immune Responses in Hepatocellular Carcinoma. Hepatology 2020, 71, 1247–1261. [Google Scholar] [CrossRef]
- Zheng, X.; Fang, Z.; Liu, X.; Deng, S.; Zhou, P.; Wang, X.; Zhang, C.; Yin, R.; Hu, H.; Chen, X.; et al. Increased vessel perfusion predicts the efficacy of immune checkpoint blockade. J. Clin. Invest. 2018, 128, 2104–2115. [Google Scholar] [CrossRef]
- Motzer, R.J.; Penkov, K.; Haanen, J.; Rini, B.; Albiges, L.; Campbell, M.T.; Venugopal, B.; Kollmannsberger, C.; Negrier, S.; Uemura, M.; et al. Avelumab plus Axitinib versus Sunitinib for Advanced Renal-Cell Carcinoma. N. Engl. J. Med. 2019, 380, 1103–1115. [Google Scholar] [CrossRef]
- Cheng, A.L.; Qin, S.; Ikeda, M.; Galle, P.R.; Ducreux, M.; Kim, T.Y.; Lim, H.Y.; Kudo, M.; Breder, V.; Merle, P.; et al. Updated efficacy and safety data from IMbrave150: Atezolizumab plus bevacizumab vs. sorafenib for unresectable hepatocellular carcinoma. J. Hepatol. 2022, 76, 862–873. [Google Scholar] [CrossRef] [PubMed]
- Schmittnaegel, M.; Rigamonti, N.; Kadioglu, E.; Cassara, A.; Wyser Rmili, C.; Kiialainen, A.; Kienast, Y.; Mueller, H.J.; Ooi, C.H.; Laoui, D.; et al. Dual angiopoietin-2 and VEGFA inhibition elicits antitumor immunity that is enhanced by PD-1 checkpoint blockade. Sci. Transl. Med. 2017, 9, eaak9670. [Google Scholar] [CrossRef]
- Makker, V.; Rasco, D.; Vogelzang, N.J.; Brose, M.S.; Cohn, A.L.; Mier, J.; Di Simone, C.; Hyman, D.M.; Stepan, D.E.; Dutcus, C.E.; et al. Lenvatinib plus pembrolizumab in patients with advanced endometrial cancer: An interim analysis of a multicentre, open-label, single-arm, phase 2 trial. Lancet Oncol. 2019, 20, 711–718. [Google Scholar] [CrossRef]
- Cleveland, A.H.; Fan, Y. Reprogramming endothelial cells to empower cancer immunotherapy. Trends Mol. Med. 2023. [Google Scholar] [CrossRef]
- Huang, Y.; Yuan, J.; Righi, E.; Kamoun, W.S.; Ancukiewicz, M.; Nezivar, J.; Santosuosso, M.; Martin, J.D.; Martin, M.R.; Vianello, F.; et al. Vascular normalizing doses of antiangiogenic treatment reprogram the immunosuppressive tumor microenvironment and enhance immunotherapy. Proc. Natl. Acad. Sci. USA 2012, 109, 17561–17566. [Google Scholar] [CrossRef]
- Sung, Y.C.; Jin, P.R.; Chu, L.A.; Hsu, F.F.; Wang, M.R.; Chang, C.C.; Chiou, S.J.; Qiu, J.T.; Gao, D.Y.; Lin, C.C.; et al. Delivery of nitric oxide with a nanocarrier promotes tumour vessel normalization and potentiates anti-cancer therapies. Nat. Nanotechnol. 2019, 14, 1160–1169. [Google Scholar] [CrossRef]
- Huerta-Yepez, S.; Vega, M.; Jazirehi, A.; Garban, H.; Hongo, F.; Cheng, G.; Bonavida, B. Nitric oxide sensitizes prostate carcinoma cell lines to TRAIL-mediated apoptosis via inactivation of NF-kappa B and inhibition of Bcl-xl expression. Oncogene 2004, 23, 4993–5003. [Google Scholar] [CrossRef]
- Hongo, F.; Garban, H.; Huerta-Yepez, S.; Vega, M.; Jazirehi, A.R.; Mizutani, Y.; Miki, T.; Bonavida, B. Inhibition of the transcription factor Yin Yang 1 activity by S-nitrosation. Biochem. Biophys. Res. Commun. 2005, 336, 692–701. [Google Scholar] [CrossRef]
- Bonavida, B. Therapeutic YY1 Inhibitors in Cancer: ALL in ONE. Crit. Rev. Oncog. 2017, 22, 37–47. [Google Scholar] [CrossRef]
- Kao, T.W.; Bai, G.H.; Wang, T.L.; Shih, I.M.; Chuang, C.M.; Lo, C.L.; Tsai, M.C.; Chiu, L.Y.; Lin, C.C.; Shen, Y.A. Novel cancer treatment paradigm targeting hypoxia-induced factor in conjunction with current therapies to overcome resistance. J. Exp. Clin. Cancer Res. 2023, 42, 171. [Google Scholar] [CrossRef]
- Erin, N.; Grahovac, J.; Brozovic, A.; Efferth, T. Tumor microenvironment and epithelial mesenchymal transition as targets to overcome tumor multidrug resistance. Drug Resist. Updates 2020, 53, 100715. [Google Scholar] [CrossRef]
- Liu, X.; Jutooru, I.; Lei, P.; Kim, K.; Lee, S.O.; Brents, L.K.; Prather, P.L.; Safe, S. Betulinic acid targets YY1 and ErbB2 through cannabinoid receptor-dependent disruption of microRNA-27a:ZBTB10 in breast cancer. Mol. Cancer Ther. 2012, 11, 1421–1431. [Google Scholar] [CrossRef] [PubMed]
- Huerta-Yepez, S.; Baritaki, S.; Baay-Guzman, G.; Hernandez-Luna, M.A.; Hernandez-Cueto, A.; Vega, M.I.; Bonavida, B. Contribution of either YY1 or BclXL-induced inhibition by the NO-donor DETANONOate in the reversal of drug resistance, both in vitro and in vivo. YY1 and BclXL are overexpressed in prostate cancer. Nitric Oxide 2013, 29, 17–24. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; He, S.; Mei, R.; Kang, Y.; Duan, J.; Wei, R.; Xiang, C.; Wu, Y.; Lu, X.; Cai, Z.; et al. miR-29a suppresses IL-13-induced cell invasion by inhibiting YY1 in the AKT pathway in lung adenocarcinoma A549 cells. Oncol. Rep. 2018, 39, 2613–2623. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Garzon, R.; Sun, H.; Ladner, K.J.; Singh, R.; Dahlman, J.; Cheng, A.; Hall, B.M.; Qualman, S.J.; Chandler, D.S.; et al. NF-kappaB-YY1-miR-29 regulatory circuitry in skeletal myogenesis and rhabdomyosarcoma. Cancer Cell 2008, 14, 369–381. [Google Scholar] [CrossRef]
- Huang, T.; Wang, G.; Yang, L.; Peng, B.; Wen, Y.; Ding, G.; Wang, Z. MiR-186 inhibits proliferation, migration, and invasion of non-small cell lung cancer cells by downregulating Yin Yang 1. Cancer Biomark. 2017, 21, 221–228. [Google Scholar] [CrossRef] [PubMed]
- Su, H.; Liu, L.; Zhang, Y.; Wang, J.; Zhao, Y. Long noncoding RNA NPCCAT1 promotes nasopharyngeal carcinoma progression via upregulating YY1. Biochimie 2019, 157, 184–194. [Google Scholar] [CrossRef] [PubMed]
- Xu, C.; Tsai, Y.H.; Galbo, P.M.; Gong, W.; Storey, A.J.; Xu, Y.; Byrum, S.D.; Xu, L.; Whang, Y.E.; Parker, J.S.; et al. Cistrome analysis of YY1 uncovers a regulatory axis of YY1:BRD2/4-PFKP during tumorigenesis of advanced prostate cancer. Nucleic Acids Res. 2021, 49, 4971–4988. [Google Scholar] [CrossRef]
- Rosenberg, S.A.; Restifo, N.P.; Yang, J.C.; Morgan, R.A.; Dudley, M.E. Adoptive cell transfer: A clinical path to effective cancer immunotherapy. Nat. Rev. Cancer 2008, 8, 299–308. [Google Scholar] [CrossRef]
- Sterner, R.C.; Sterner, R.M. CAR-T cell therapy: Current limitations and potential strategies. Blood Cancer J. 2021, 11, 69. [Google Scholar] [CrossRef]
- Benmebarek, M.R.; Karches, C.H.; Cadilha, B.L.; Lesch, S.; Endres, S.; Kobold, S. Killing Mechanisms of Chimeric Antigen Receptor (CAR) T Cells. Int. J. Mol. Sci. 2019, 20, 1283. [Google Scholar] [CrossRef]
- Safarzadeh Kozani, P.; Safarzadeh Kozani, P.; Ahmadi Najafabadi, M.; Yousefi, F.; Mirarefin, S.M.J.; Rahbarizadeh, F. Recent Advances in Solid Tumor CAR-T Cell Therapy: Driving Tumor Cells From Hero to Zero? Front. Immunol. 2022, 13, 795164. [Google Scholar] [CrossRef] [PubMed]
- Kochenderfer, J.N.; Wilson, W.H.; Janik, J.E.; Dudley, M.E.; Stetler-Stevenson, M.; Feldman, S.A.; Maric, I.; Raffeld, M.; Nathan, D.A.; Lanier, B.J.; et al. Eradication of B-lineage cells and regression of lymphoma in a patient treated with autologous T cells genetically engineered to recognize CD19. Blood 2010, 116, 4099–4102. [Google Scholar] [CrossRef] [PubMed]
- T Cell Receptor Immunotherapy Targeting VEGFR2 for Patients with Metastatic Cancer. 2022. Available online: https://clinicaltrials.gov/ct2/show/NCT01218867CAR (accessed on 6 December 2023).
- Chinnasamy, D.; Yu, Z.; Theoret, M.R.; Zhao, Y.; Shrimali, R.K.; Morgan, R.A.; Feldman, S.A.; Restifo, N.P.; Rosenberg, S.A. Gene therapy using genetically modified lymphocytes targeting VEGFR-2 inhibits the growth of vascularized syngenic tumors in mice. J. Clin. Investig. 2010, 120, 3953–3968. [Google Scholar] [CrossRef]
- Wang, W.; Ma, Y.; Li, J.; Shi, H.S.; Wang, L.Q.; Guo, F.C.; Zhang, J.; Li, D.; Mo, B.H.; Wen, F.; et al. Specificity redirection by CAR with human VEGFR-1 affinity endows T lymphocytes with tumor-killing ability and anti-angiogenic potency. Gene Ther. 2013, 20, 970–978. [Google Scholar] [CrossRef] [PubMed]
- Xing, H.; Yang, X.; Xu, Y.; Tang, K.; Tian, Z.; Chen, Z.; Zhang, Y.; Xue, Z.; Rao, Q.; Wang, M.; et al. Anti-tumor effects of vascular endothelial growth factor/vascular endothelial growth factor receptor binding domain-modified chimeric antigen receptor T cells. Cytotherapy 2021, 23, 810–819. [Google Scholar] [CrossRef] [PubMed]
- Byrd, T.T.; Fousek, K.; Pignata, A.; Szot, C.; Samaha, H.; Seaman, S.; Dobrolecki, L.; Salsman, V.S.; Oo, H.Z.; Bielamowicz, K.; et al. TEM8/ANTXR1-Specific CAR T Cells as a Targeted Therapy for Triple-Negative Breast Cancer. Cancer Res. 2018, 78, 489–500. [Google Scholar] [CrossRef] [PubMed]
- Fierle, J.K.; Brioschi, M.; de Tiani, M.; Wetterwald, L.; Atsaves, V.; Abram-Saliba, J.; Petrova, T.V.; Coukos, G.; Dunn, S.M. Soluble trivalent engagers redirect cytolytic T cell activity toward tumor endothelial marker 1. Cell Rep. Med. 2021, 2, 100362. [Google Scholar] [CrossRef]
- Zhuang, X.; Maione, F.; Robinson, J.; Bentley, M.; Kaul, B.; Whitworth, K.; Jumbu, N.; Jinks, E.; Bystrom, J.; Gabriele, P.; et al. CAR T cells targeting tumor endothelial marker CLEC14A inhibit tumor growth. JCI Insight 2020, 5, e138808. [Google Scholar] [CrossRef]
- Wagner, J.; Wickman, E.; Shaw, T.I.; Anido, A.A.; Langfitt, D.; Zhang, J.; Porter, S.N.; Pruett-Miller, S.M.; Tillman, H.; Krenciute, G.; et al. Antitumor Effects of CAR T Cells Redirected to the EDB Splice Variant of Fibronectin. Cancer Immunol. Res. 2021, 9, 279–290. [Google Scholar] [CrossRef]
- Xie, Y.J.; Dougan, M.; Jailkhani, N.; Ingram, J.; Fang, T.; Kummer, L.; Momin, N.; Pishesha, N.; Rickelt, S.; Hynes, R.O.; et al. Nanobody-based CAR T cells that target the tumor microenvironment inhibit the growth of solid tumors in immunocompetent mice. Proc. Natl. Acad. Sci. USA 2019, 116, 7624–7631. [Google Scholar] [CrossRef]
- Wallstabe, L.; Mades, A.; Frenz, S.; Einsele, H.; Rader, C.; Hudecek, M. CAR T cells targeting alpha(v)beta(3) integrin are effective against advanced cancer in preclinical models. Adv. Cell Gene Ther. 2018, 1, e11. [Google Scholar] [CrossRef] [PubMed]
- Fu, X.; Rivera, A.; Tao, L.; Zhang, X. Genetically modified T cells targeting neovasculature efficiently destroy tumor blood vessels, shrink established solid tumors and increase nanoparticle delivery. Int. J. Cancer 2013, 133, 2483–2492. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Li, Y.; Tan, B.; Zhao, Q.; Fan, L.; Li, F.; Zhao, X. Progress and current status of molecule-targeted therapy and drug resistance in gastric cancer. Drugs Today 2020, 56, 469–482. [Google Scholar] [CrossRef] [PubMed]
- Rini, B.I.; Atkins, M.B. Resistance to targeted therapy in renal-cell carcinoma. Lancet Oncol. 2009, 10, 992–1000. [Google Scholar] [CrossRef] [PubMed]
- Oladipupo, S.S.; Hu, S.; Santeford, A.C.; Yao, J.; Kovalski, J.R.; Shohet, R.V.; Maslov, K.; Wang, L.V.; Arbeit, J.M. Conditional HIF-1 induction produces multistage neovascularization with stage-specific sensitivity to VEGFR inhibitors and myeloid cell independence. Blood 2011, 117, 4142–4153. [Google Scholar] [CrossRef] [PubMed]
- Bottsford-Miller, J.N.; Coleman, R.L.; Sood, A.K. Resistance and escape from antiangiogenesis therapy: Clinical implications and future strategies. J. Clin. Oncol. 2012, 30, 4026–4034. [Google Scholar] [CrossRef] [PubMed]
- Lai, X.M.; Liu, S.Y.; Tsai, Y.T.; Sun, G.H.; Chang, S.Y.; Huang, S.M.; Cha, T.L. HAF mediates the evasive resistance of anti-angiogenesis TKI through disrupting HIF-1alpha and HIF-2alpha balance in renal cell carcinoma. Oncotarget 2017, 8, 49713–49724. [Google Scholar] [CrossRef] [PubMed]
- Zhao, D.; Zhai, B.; He, C.; Tan, G.; Jiang, X.; Pan, S.; Dong, X.; Wei, Z.; Ma, L.; Qiao, H.; et al. Upregulation of HIF-2alpha induced by sorafenib contributes to the resistance by activating the TGF-alpha/EGFR pathway in hepatocellular carcinoma cells. Cell. Signal. 2014, 26, 1030–1039. [Google Scholar] [CrossRef]
- Shi, X.; Wang, M.; Zhang, Y.; Guo, X.; Liu, M.; Zhou, Z.; Zhao, Y.; He, R.; Gao, Y.; Liu, Y.; et al. Hypoxia activated HGF expression in pancreatic stellate cells confers resistance of pancreatic cancer cells to EGFR inhibition. EBioMedicine 2022, 86, 104352. [Google Scholar] [CrossRef]
- Lu, Y.; Liu, Y.; Oeck, S.; Zhang, G.J.; Schramm, A.; Glazer, P.M. Hypoxia Induces Resistance to EGFR Inhibitors in Lung Cancer Cells via Upregulation of FGFR1 and the MAPK Pathway. Cancer Res. 2020, 80, 4655–4667. [Google Scholar] [CrossRef]
- Sarvagalla, S.; Kolapalli, S.P.; Vallabhapurapu, S. The Two Sides of YY1 in Cancer: A Friend and a Foe. Front. Oncol. 2019, 9, 1230. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Crosstalk between tumor vasculature and immune cells in the tumor microenvironment. The TME plays a key role in tumor growth by influencing stromal and immune cells. Hypoxia TME promotes tumor vascularization by activating HIF-1α, VEGF, and many chemokines as indicated. Hypoxia increases immune suppression via proinflammatory molecules. Immune cells contribute to both the immunosuppressive tumor microenvironment and abnormal vascularization. Figure created with a Biorender program.
Figure 1.
Crosstalk between tumor vasculature and immune cells in the tumor microenvironment. The TME plays a key role in tumor growth by influencing stromal and immune cells. Hypoxia TME promotes tumor vascularization by activating HIF-1α, VEGF, and many chemokines as indicated. Hypoxia increases immune suppression via proinflammatory molecules. Immune cells contribute to both the immunosuppressive tumor microenvironment and abnormal vascularization. Figure created with a Biorender program.

Figure 2.
Hypoxia in the tumor microenvironment increases the secretion of VEGF, leading to the generation of leaky and abnormal vasculature limiting the access of drugs to the tumor. Hypoxia also creates a niche for cancer stem cells (CSCs). Antiangiogenic agents normalize the vasculature and create appropriate blood perfusion changes in the immune cell components present in the tumor microenvironment (TME). Tumors with normalized vessels are enriched in tumor-associated macrophages (TAM) with an M1 phenotype, immune protective cells (CD8+ T, CD4+ T, and dendritic cells), and present low infiltration of immunosuppressive cells (Treg, CCR2+ cells). The image is produced using the Biorender program.
Figure 2.
Hypoxia in the tumor microenvironment increases the secretion of VEGF, leading to the generation of leaky and abnormal vasculature limiting the access of drugs to the tumor. Hypoxia also creates a niche for cancer stem cells (CSCs). Antiangiogenic agents normalize the vasculature and create appropriate blood perfusion changes in the immune cell components present in the tumor microenvironment (TME). Tumors with normalized vessels are enriched in tumor-associated macrophages (TAM) with an M1 phenotype, immune protective cells (CD8+ T, CD4+ T, and dendritic cells), and present low infiltration of immunosuppressive cells (Treg, CCR2+ cells). The image is produced using the Biorender program.
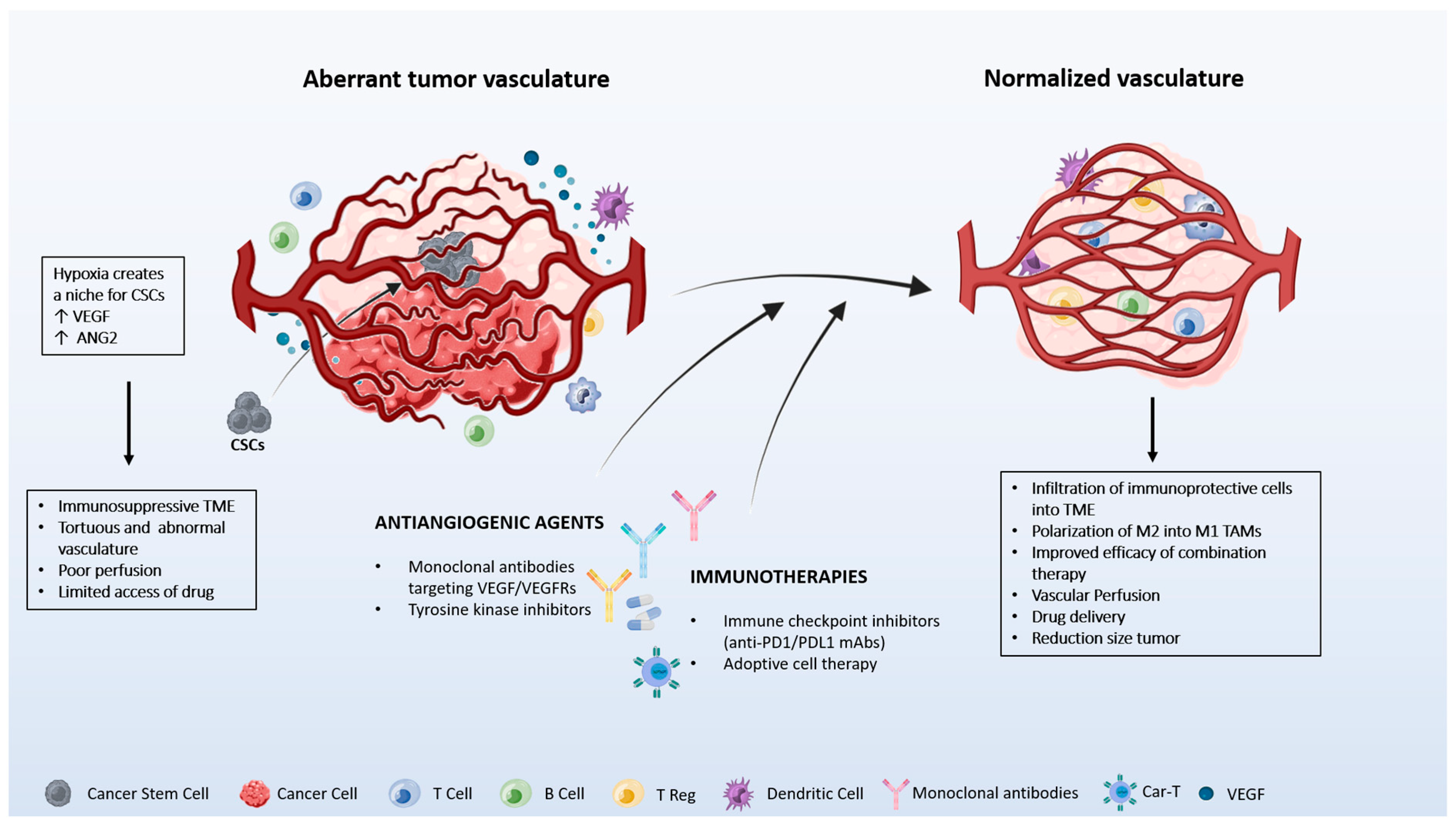
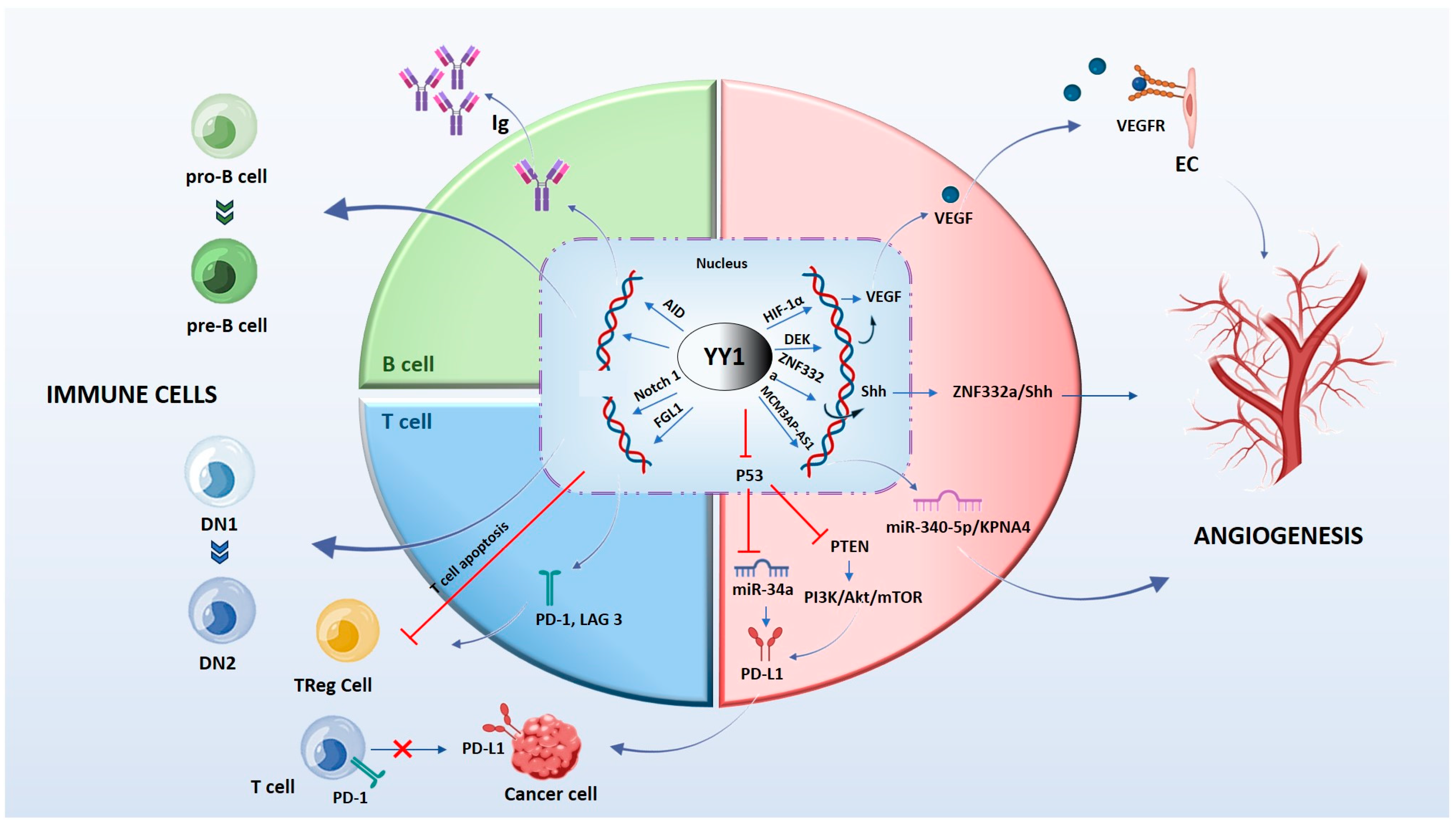
Figure 3.
The molecular mechanism by which YY1 regulates immune cells and angiogenesis in the tumor microenvironment. YY1 influences B cells and T cells through several signaling pathways. YY1 is essential for all stages of B cell development and controls processes responsible for antibody maturation and expression of different immunoglobulin. YY1 inhibits the function of T and Treg cells and favors the expression binding to promoter regions of PD-1 and LAG3. YY1 activates the Notch1 pathway, promoting the DN1-to-DN2 T cell transition. YY1 regulates the expression of crucial proteins associated with tumor neovascularization through the interaction with their promoters. Images were prepared using the Biorender program.
Figure 3.
The molecular mechanism by which YY1 regulates immune cells and angiogenesis in the tumor microenvironment. YY1 influences B cells and T cells through several signaling pathways. YY1 is essential for all stages of B cell development and controls processes responsible for antibody maturation and expression of different immunoglobulin. YY1 inhibits the function of T and Treg cells and favors the expression binding to promoter regions of PD-1 and LAG3. YY1 activates the Notch1 pathway, promoting the DN1-to-DN2 T cell transition. YY1 regulates the expression of crucial proteins associated with tumor neovascularization through the interaction with their promoters. Images were prepared using the Biorender program.

{kind=link}
{kind=link}
{kind=link}
Table 1.
Studies of CAR T cells in the tumor vasculature.
| Study | CAR T Cell Design | Study Data | Ref. | |||
|---|---|---|---|---|---|---|
| Vessel Target | Tumor Type | Study Type/Model | Ag Recognition; Clone | Construct | End Points | |
| VEGFR2 | Metastatic cancer | Phase I/II | scFv anti-human VEGFR-2; KDR1121 | CD8/CD28/4–1-BB/CD3ζ | No response/Progression Disease | [123] |
| VEGFR2 | Melanoma (B16F10) | Syngeneic | scFv anti-mouse VEGFR-2; DC101 | CD8/CD28/4–1-BB/CD3ζ | Inhibition of tumor growth; increased survival | [124] |
| Fibrosarcoma (MCA205) | ||||||
| Colon (MC38) | ||||||
| Colon (CT26) | ||||||
| kidney (RENCA) | ||||||
| VEGFR1 | Lung carcinoma (A549) | Xenograft | scFv anti-human VEGFR1; IMC-18F1 | IgG-Fc/CD4/CD3ζ | Inhibition of tumor growth; increased survival; metastasis inhibition | [125] |
| VEGFR2/3 | Breast (MDA-MB-231) | Xenograft | N-terminus VEGF-C | CD8/CD28/CD3ζ | Inhibition of tumor growth; metastasis inhibition | [126] |
| Breast (HCC1806) | ||||||
| TEM8 | Breast (MDA-MB-468) | Xenograft | scFv anti-human TEM8; L2 | IgG-Fc/CD28/4–1-BB/CD3ζ | Inhibition of tumor growth; increased survival; vascular disruption | [127] |
| Breast (LM231) | ||||||
| Breast (BCM-2665) | Patient-derived xenograft | |||||
| Breast (WHIM12) | ||||||
| TEM1 | Ewing sarcoma (A673) | Xenograft | scFv anti-human and anti-mouse TEM1; L1C1m | Trilobite engager (CD3/TEM1) | Inhibition of tumor growth | [128] |
| CLEC14a | Healthy mice | N/A Transgenic | scFV anti-mouse and anti-human CLEC14a; CRT3, CRT5 | CD28/CD3ζ | Inhibition of tumor growth; increased survival | [129] |
| RipTag2 | ||||||
| Pancreas (mPDAC) | Syngeneic | |||||
| Lung (LLC) | Syngeneic | |||||
| ED-B | Glioma (U87) | Xenograft | scFv anti-human ED-B; L19 | CD28/CD3ζ | Inhibition of tumor growth; increased survival | [130] |
| Lung (A549) | ||||||
| Ewing sarcoma (A673) | ||||||
| ED-B | Melanoma (B16F10) | Syngeneic | VHH; NJB2; camelid | CD8/CD28/CD3ζ | Inhibition of tumor growth; increased survival | [131] |
| Colon (MC38) | ||||||
| Integrin αvβ3 | Melanoma (A375) | Xenograft | scFv anti-human αvβ3; LM609 | CD28/CD3ζ | Tumor regression; increased survival | [132] |
| Integrin αvβ3 | Melanoma (B16F10) | Syngeneic | Echistatin (Disintegrin in snake venom) | CD28/CD3ζ | Inhibition of tumor growth; vascular disruption | [133] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Meo, C.; de Nigris, F. Clinical Potential of YY1-Hypoxia Axis for Vascular Normalization and to Improve Immunotherapy. Cancers 2024, 16, 491. https://doi.org/10.3390/cancers16030491
AMA Style
Meo C, de Nigris F. Clinical Potential of YY1-Hypoxia Axis for Vascular Normalization and to Improve Immunotherapy. Cancers. 2024; 16(3):491. https://doi.org/10.3390/cancers16030491
Chicago/Turabian StyleMeo, Concetta, and Filomena de Nigris. 2024. "Clinical Potential of YY1-Hypoxia Axis for Vascular Normalization and to Improve Immunotherapy" Cancers 16, no. 3: 491. https://doi.org/10.3390/cancers16030491
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.