MicroRNAs in the Pathogenesis, Diagnosis, Prognosis and Targeted Treatment of Cutaneous T-Cell Lymphomas


 ,
,  ,
,  ,
,  ,
, 

Abstract
:1. Introduction
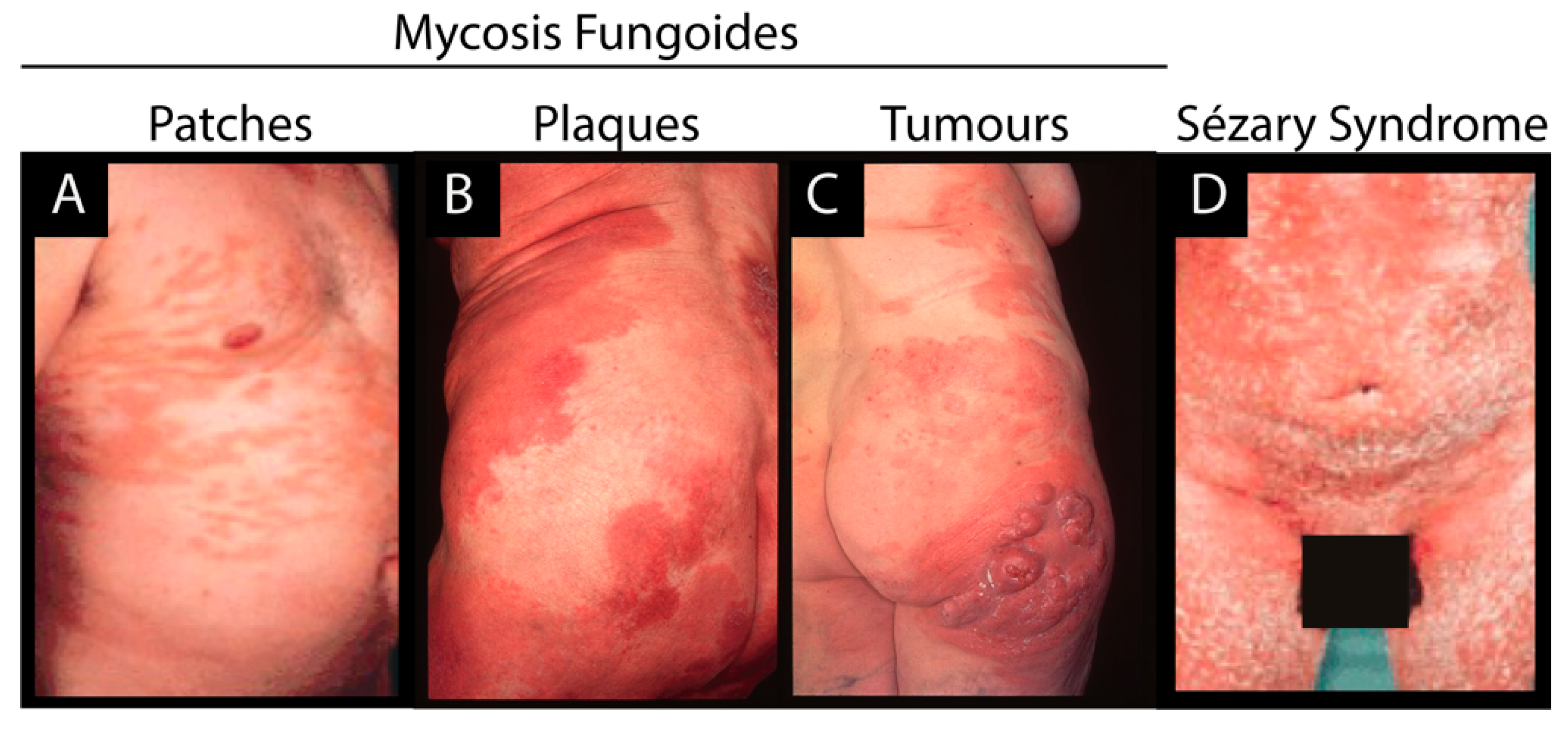
1.1. Etiology of Cutaneous T Cell Lymphoma
1.2. MicroRNAs in Health and Disease
2. Differentially Expressed miRs in CTCL
3. Dysregulation of miRs in CTCL
4. OncomiRs in CTCL Pathogenesis
4.1. Role of miR-21 in Malignant Proliferation
4.2. Role of miR-155 in Disease Progression
4.3. Role of miR-214 in Promoting Cell Survival in SS
4.4. The miR-17/92 Cluster—Role as oncomiRs or Tumor Suppressor miRs in CTCL?
5. Key Tumor Suppressive miRs Playing a Role in CTCL Pathogenesis
6. Non-Canonical Functions of miRs
7. miRs as Diagnostic, Prognostic and Treatment Predictive Biomarkers
8. Dual Interplay between miRs and Therapeutics Targeting the Epigenome
9. miR Targeted Therapy
10. Conclusions
Funding
Acknowledgments
Conflicts of Interest
References
- Hristov, A.C.; Tejasvi, T.; Wilcox, R.A. Mycosis fungoides and Sézary syndrome: 2019 update on diagnosis, risk-stratification, and management. Am. J. Hematol. 2019, 94, 1027–1041. [Google Scholar] [CrossRef] [Green Version]
- Kim, E.J.; Hess, S.; Richardson, S.K.; Newton, S.; Showe, L.C.; Benoit, B.M.; Ubriani, R.; Vittorio, C.C.; Junkins-Hopkins, J.M.; Wysocka, M.; et al. Immunopathogenesis and therapy of cutaneous T cell lymphoma. J. Clin. Investig. 2005, 115, 798–812. [Google Scholar] [CrossRef] [Green Version]
- Lindahl, L.M.; Besenbacher, S.; Rittig, A.H.; Celis, P.; Willerslev-Olsen, A.; Gjerdrum, L.M.R.; Krejsgaard, T.; Johansen, C.; Litman, T.; Woetmann, A.; et al. Prognostic miRNA classifier in early-stage mycosis fungoides: Development and validation in a Danish nationwide study. Blood 2018, 131, 759–770. [Google Scholar] [CrossRef] [Green Version]
- Litvinov, I.V.; Shtreis, A.; Kobayashi, K.; Glassman, S.; Tsang, M.; Woetmann, A.; Sasseville, D.; Ødum, N.; Duvic, M. Investigating potential exogenous tumor initiating and promoting factors for Cutaneous T-Cell Lymphomas (CTCL), a rare skin malignancy. Oncoimmunology 2016, 5, e1175799. [Google Scholar] [CrossRef] [Green Version]
- Ghazawi, F.M.; Alghazawi, N.; Le, M.; Netchiporouk, E.; Glassman, S.J.; Sasseville, D.; Litvinov, I.V. Environmental and Other Extrinsic Risk Factors Contributing to the Pathogenesis of Cutaneous T Cell Lymphoma (CTCL). Front. Oncol. 2019, 9. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.; Goh, G.; Walradt, T.; Hong, B.S.; Bunick, C.G.; Chen, K.; Bjornson, R.D.; Maman, Y.; Wang, T.; Tordoff, J.; et al. Genomic landscape of cutaneous T cell lymphoma. Nat. Genet. 2015, 47, 1011–1019. [Google Scholar] [CrossRef] [PubMed]
- Netchiporouk, E.; Gantchev, J.; Tsang, M.; Thibault, P.; Watters, A.K.; Hughes, J.-D.M.; Ghazawi, F.M.; Woetmann, A.; Ødum, N.; Sasseville, D.; et al. Analysis of CTCL cell lines reveals important differences between mycosis fungoides/Sézary syndrome vs. HTLV-1+ leukemic cell lines. Oncotarget 2017, 8, 95981–95998. [Google Scholar] [CrossRef] [PubMed]
- Ødum, N. Deregulated signalling and inflammation in cutaneous T-cell lymphoma. Br. J. Dermatol. 2020, 182, 16–17. [Google Scholar] [CrossRef]
- Kiel, M.J.; Sahasrabuddhe, A.A.; Rolland, D.C.M.; Velusamy, T.; Chung, F.; Schaller, M.; Bailey, N.G.; Betz, B.L.; Miranda, R.N.; Porcu, P.; et al. Genomic analyses reveal recurrent mutations in epigenetic modifiers and the JAK–STAT pathway in Sézary syndrome. Nat. Commun. 2015, 6, 8470. [Google Scholar] [CrossRef]
- Yumeen, S.; Girardi, M. Insights Into the Molecular and Cellular Underpinnings of Cutaneous T Cell Lymphoma. Yale J. Biol. Med. 2020, 93, 111–121. [Google Scholar]
- Seffens, A.; Herrera, A.; Tegla, C.; Buus, T.B.; Hymes, K.B.; Ødum, N.; Geskin, L.J.; Koralov, S.B. STAT3 Dysregulation in Mature T and NK Cell Lymphomas. Cancers 2019, 11, 1711. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Torres, A.N.B.; Cats, D.; Mei, H.; Szuhai, K.; Willemze, R.; Vermeer, M.H.; Tensen, C.P. Genomic analysis reveals recurrent deletion of JAK-STAT signaling inhibitors HNRNPK and SOCS1 in mycosis fungoides. Geneschromosomes Cancer 2018, 57, 653–664. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brender, C.; Nielsen, M.; Kaltoft, K.; Mikkelsen, G.; Zhang, Q.; Wasik, M.; Billestrup, N.; Odum, N. STAT3-mediated constitutive expression of SOCS-3 in cutaneous T-cell lymphoma. Blood 2001, 97, 1056–1062. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yi, S.; Sun, J.; Qiu, L.; Fu, W.; Wang, A.; Liu, X.; Yang, Y.; Kadin, M.E.; Tu, P.; Wang, Y. Dual Role of EZH2 in Cutaneous Anaplastic Large Cell Lymphoma: Promoting Tumor Cell Survival and Regulating Tumor Microenvironment. J. Investig. Dermatol. 2018, 138, 1126–1136. [Google Scholar] [CrossRef] [Green Version]
- Qiu, L.; Liu, F.; Yi, S.; Li, X.; Liu, X.; Xiao, C.; Lian, C.G.; Tu, P.; Wang, Y. Loss of 5-Hydroxymethylcytosine Is an Epigenetic Biomarker in Cutaneous T-Cell Lymphoma. J. Investig. Dermatol. 2018, 138, 2388–2397. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Wang, J.; Yu, M.; Wang, Y.; Zhang, H.; Yin, J.; Li, Z.; Li, T.; Yan, H.; Li, F.; et al. SNF5 deficiency induces apoptosis resistance by repressing SATB1 expression in Sézary syndrome. Leuk. Lymphoma 2018, 59, 2405–2413. [Google Scholar] [CrossRef]
- Lopez, A.T.; Bates, S.; Geskin, L. Current Status of HDAC Inhibitors in Cutaneous T-cell Lymphoma. Am. J. Clin. Derm. 2018. [Google Scholar] [CrossRef]
- Fantin, V.R.; Loboda, A.; Paweletz, C.P.; Hendrickson, R.C.; Pierce, J.W.; Roth, J.A.; Li, L.; Gooden, F.; Korenchuk, S.; Hou, X.S.; et al. Constitutive activation of signal transducers and activators of transcription predicts vorinostat resistance in cutaneous T-cell lymphoma. Cancer Res. 2008, 68, 3785–3794. [Google Scholar] [CrossRef] [Green Version]
- Si, W.; Shen, J.; Zheng, H.; Fan, W. The role and mechanisms of action of microRNAs in cancer drug resistance. Clin. Epigenetics 2019, 11, 25. [Google Scholar] [CrossRef]
- Lin, S.; Gregory, R.I. MicroRNA biogenesis pathways in cancer. Nat. Rev. Cancer 2015, 15, 321–333. [Google Scholar] [CrossRef]
- Ali Syeda, Z.; Langden, S.S.S.; Munkhzul, C.; Lee, M.; Song, S.J. Regulatory Mechanism of MicroRNA Expression in Cancer. Int. J. Mol. Sci. 2020, 21, 1723. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Calin, G.A.; Sevignani, C.; Dumitru, C.D.; Hyslop, T.; Noch, E.; Yendamuri, S.; Shimizu, M.; Rattan, S.; Bullrich, F.; Negrini, M.; et al. Human microRNA genes are frequently located at fragile sites and genomic regions involved in cancers. Proc. Natl. Acad. Sci. USA 2004, 101, 2999–3004. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maj, J.; Jankowska-Konsur, A.; Sadakierska-Chudy, A.; Noga, L.; Reich, A. Altered microRNA expression in mycosis fungoides. Br. J. Dermatol. 2012, 166, 331–336. [Google Scholar] [CrossRef] [PubMed]
- Marosvári, D.; Téglási, V.; Csala, I.; Marschalkó, M.; Bödör, C.; Timár, B.; Csomor, J.; Hársing, J.; Reiniger, L. Altered microRNA expression in folliculotropic and transformed mycosis fungoides. Pathol. Oncol. Res. 2015, 21, 821–825. [Google Scholar] [CrossRef]
- Benner, M.F.; Ballabio, E.; van Kester, M.S.; Saunders, N.J.; Vermeer, M.H.; Willemze, R.; Lawrie, C.H.; Tensen, C.P. Primary cutaneous anaplastic large cell lymphoma shows a distinct miRNA expression profile and reveals differences from tumor-stage mycosis fungoides. Exp. Dermatol. 2012, 21, 632–634. [Google Scholar] [CrossRef] [Green Version]
- Papadavid, E.; Braoudaki, M.; Bourdakou, M.; Lykoudi, A.; Nikolaou, V.; Tounta, G.; Ekonomidi, A.; Athanasiadis, E.; Spyrou, G.; Antoniou, C.; et al. Aberrant microRNA expression in tumor mycosis fungoides. Tumour Biol. 2016, 37, 14667–14675. [Google Scholar] [CrossRef]
- Van Kester, M.S.; Ballabio, E.; Benner, M.F.; Chen, X.H.; Saunders, N.J.; van der Fits, L.; van Doorn, R.; Vermeer, M.H.; Willemze, R.; Tensen, C.P.; et al. miRNA expression profiling of mycosis fungoides. Mol. Oncol. 2011, 5, 273–280. [Google Scholar] [CrossRef] [Green Version]
- Shen, X.; Wang, B.; Li, K.; Wang, L.; Zhao, X.; Xue, F.; Shi, R.; Zheng, J. MicroRNA Signatures in Diagnosis and Prognosis of Cutaneous T-Cell Lymphoma. J. Investig. Dermatol. 2018, 138, 2024–2032. [Google Scholar] [CrossRef] [Green Version]
- Narducci, M.G.; Arcelli, D.; Picchio, M.C.; Lazzeri, C.; Pagani, E.; Sampogna, F.; Scala, E.; Fadda, P.; Cristofoletti, C.; Facchiano, A.; et al. MicroRNA profiling reveals that miR-21, miR486 and miR-214 are upregulated and involved in cell survival in Sézary syndrome. Cell Death Dis. 2011, 2, e151. [Google Scholar] [CrossRef] [Green Version]
- Qin, Y.; Buermans, H.P.J.; van Kester, M.S.; van der Fits, L.; Out-Luiting, J.J.; Osanto, S.; Willemze, R.; Vermeer, M.H.; Tensen, C.P. Deep-Sequencing Analysis Reveals that the miR-199a2/214 Cluster within DNM3os Represents the Vast Majority of Aberrantly Expressed MicroRNAs in Sézary Syndrome. J. Investig. Dermatol. 2012, 132, 1520–1522. [Google Scholar] [CrossRef] [Green Version]
- Ballabio, E.; Mitchell, T.; van Kester, M.S.; Taylor, S.; Dunlop, H.M.; Chi, J.; Tosi, I.; Vermeer, M.H.; Tramonti, D.; Saunders, N.J.; et al. MicroRNA expression in Sezary syndrome: Identification, function, and diagnostic potential. Blood 2010, 116, 1105–1113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kohnken, R.; Kodigepalli, K.M.; Mishra, A.; Porcu, P.; Wu, L. MicroRNA-181 contributes to downregulation of SAMHD1 expression in CD4+ T-cells derived from Sèzary syndrome patients. Leuk. Res. 2017, 52, 58–66. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rittig, A.H.; Lindahl, L.M.; Johansen, C.; Celis, P.; Ødum, N.; Iversen, L.; Litman, T. The MicroRNA Expression Profile Differs Between Erythrodermic Mycosis Fungoides and Sézary Syndrome. Acta Derm. Venereol. 2019, 99, 1148–1153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sandoval, J.; Díaz-Lagares, A.; Salgado, R.; Servitje, O.; Climent, F.; Ortiz-Romero, P.L.; Pérez-Ferriols, A.; Garcia-Muret, M.P.; Estrach, T.; Garcia, M.; et al. MicroRNA expression profiling and DNA methylation signature for deregulated microRNA in cutaneous T-cell lymphoma. J. Investig. Dermatol. 2015, 135, 1128–1137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gallardo, F.; Sandoval, J.; Díaz-Lagares, A.; Garcia, R.; D’Altri, T.; González, J.; Alegre, V.; Servitje, O.; Crujeiras, A.-B.; Stefánsson, Ó.-A.; et al. Notch1 Pathway Activation Results from the Epigenetic Abrogation of Notch-Related MicroRNAs in Mycosis Fungoides. J. Investig. Dermatol. 2015, 135, 3144–3152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kamstrup, M.R.; Gjerdrum, L.M.R.; Biskup, E.; Thyssing Lauenborg, B.; Ralfkiaer, E.; Woetmann, A.; Ødum, N.; Gniadecki, R. Notch1 as a potential therapeutic target in cutaneous T-cell lymphoma. Blood 2010, 116, 2504–2512. [Google Scholar] [CrossRef]
- Garaicoa, F.H.; Roisman, A.; Arias, M.; Trila, C.; Fridmanis, M.; Abeldaño, A.; Vanzulli, S.; Narbaitz, M.; Slavutsky, I. Genomic imbalances and microRNA transcriptional profiles in patients with mycosis fungoides. Tumour Biol. 2016, 37, 13637–13647. [Google Scholar] [CrossRef]
- Gluud, M.; Fredholm, S.; Blümel, E.; Willerslev-Olsen, A.; Buus, T.B.; Nastasi, C.; Krejsgaard, T.; Bonefeld, C.M.; Woetmann, A.; Iversen, L.; et al. MicroRNA-93 Targets p21 and Promotes Proliferation in Mycosis Fungoides T Cells. Dermatology 2020, 1–6. [Google Scholar] [CrossRef]
- Lindahl, L.M.; Fredholm, S.; Joseph, C.; Nielsen, B.S.; Jønson, L.; Willerslev-Olsen, A.; Gluud, M.; Blümel, E.; Petersen, D.L.; Sibbesen, N.; et al. STAT5 induces miR-21 expression in cutaneous T cell lymphoma. Oncotarget 2016, 7, 45730–45744. [Google Scholar] [CrossRef] [Green Version]
- Kopp, K.L.; Ralfkiaer, U.; Gjerdrum, L.M.R.; Helvad, R.; Pedersen, I.H.; Litman, T.; Jønson, L.; Hagedorn, P.H.; Krejsgaard, T.; Gniadecki, R.; et al. STAT5-mediated expression of oncogenic miR-155 in cutaneous T-cell lymphoma. Cell Cycle 2013, 12, 1939–1947. [Google Scholar] [CrossRef]
- Van der Fits, L.; van Kester, M.S.; Qin, Y.; Out-Luiting, J.J.; Smit, F.; Zoutman, W.H.; Willemze, R.; Tensen, C.P.; Vermeer, M.H. MicroRNA-21 expression in CD4+ T cells is regulated by STAT3 and is pathologically involved in Sézary syndrome. J. Investig. Dermatol. 2011, 131, 762–768. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sibbesen, N.A.; Kopp, K.L.; Litvinov, I.V.; Jønson, L.; Willerslev-Olsen, A.; Fredholm, S.; Petersen, D.L.; Nastasi, C.; Krejsgaard, T.; Lindahl, L.M.; et al. Jak3, STAT3, and STAT5 inhibit expression of miR-22, a novel tumor suppressor microRNA, in cutaneous T-Cell lymphoma. Oncotarget 2015, 6, 20555–20569. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valencak, J.; Schmid, K.; Trautinger, F.; Wallnöfer, W.; Muellauer, L.; Soleiman, A.; Knobler, R.; Haitel, A.; Pehamberger, H.; Raderer, M. High expression of Dicer reveals a negative prognostic influence in certain subtypes of primary cutaneous T cell lymphomas. J. Dermatol. Sci. 2011, 64, 185–190. [Google Scholar] [CrossRef] [PubMed]
- Gambichler, T.; Salveridou, K.; Schmitz, L.; Käfferlein, H.U.; Brüning, T.; Stockfleth, E.; Sand, M.; Lang, K. Low Drosha protein expression in cutaneous T-cell lymphoma is associated with worse disease outcome. J. Eur. Acad. Derm. Venereol. 2019, 33, 1695–1699. [Google Scholar] [CrossRef]
- Odum, N.; Lindahl, L.M.; Wod, M.; Krejsgaard, T.; Skytthe, A.; Woetmann, A.; Iversen, L.; Christensen, K. Investigating heredity in cutaneous T-cell lymphoma in a unique cohort of Danish twins. Blood Cancer J. 2017, 7, e517. [Google Scholar] [CrossRef] [Green Version]
- Lindahl, L.M.; Willerslev-Olsen, A.; Gjerdrum, L.M.R.; Nielsen, P.R.; Blümel, E.; Rittig, A.H.; Celis, P.; Herpers, B.; Becker, J.C.; Stausbøl-Grøn, B.; et al. Antibiotics inhibit tumor and disease activity in cutaneous T cell lymphoma. Blood 2019. [Google Scholar] [CrossRef]
- Blümel, E.; Willerslev-Olsen, A.; Gluud, M.; Lindahl, L.M.; Fredholm, S.; Nastasi, C.; Krejsgaard, T.; Surewaard, B.G.J.; Koralov, S.B.; Hu, T.; et al. Staphylococcal alpha-toxin tilts the balance between malignant and non-malignant CD4+ T cells in cutaneous T-cell lymphoma. OncoImmunology 2019, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Fanok, M.H.; Sun, A.; Fogli, L.K.; Narendran, V.; Eckstein, M.; Kannan, K.; Dolgalev, I.; Lazaris, C.; Heguy, A.; Laird, M.E.; et al. Role of Dysregulated Cytokine Signaling and Bacterial Triggers in the Pathogenesis of Cutaneous T-Cell Lymphoma. J. Investig. Derm. 2018, 138, 1116–1125. [Google Scholar] [CrossRef] [Green Version]
- Willerslev-Olsen, A. Staphylococcus aureus enterotoxins induce FOXP3 in neoplastic T cells in Sézary syndrome. Blood Cancer J. in press.
- Rao, R.; Rieder, S.A.; Nagarkatti, P.; Nagarkatti, M. Staphylococcal enterotoxin B-induced microRNA-155 targets SOCS1 to promote acute inflammatory lung injury. Infect. Immun. 2014, 82, 2971–2979. [Google Scholar] [CrossRef] [Green Version]
- Santos, J.C.; Brianti, M.T.; Almeida, V.R.; Ortega, M.M.; Fischer, W.; Haas, R.; Matheu, A.; Ribeiro, M.L. Helicobacter pylori infection modulates the expression of miRNAs associated with DNA mismatch repair pathway. Mol. Carcinog. 2017, 56, 1372–1379. [Google Scholar] [CrossRef] [PubMed]
- Ou, Y.; Ren, H.; Zhao, R.; Song, L.; Liu, Z.; Xu, W.; Liu, Y.; Wang, S. Helicobacter pylori CagA promotes the malignant transformation of gastric mucosal epithelial cells through the dysregulation of the miR-155/KLF4 signaling pathway. Mol. Carcinog. 2019, 58, 1427–1437. [Google Scholar] [CrossRef] [PubMed]
- Floch, P.; Capdevielle, C.; Staedel, C.; Izotte, J.; Sifré, E.; Laur, A.M.; Giese, A.; Korolik, V.; Dubus, P.; Mégraud, F.; et al. Deregulation of MicroRNAs in Gastric Lymphomagenesis Induced in the d3Tx Mouse Model of Helicobacter pylori Infection. Front. Cell Infect. Microbiol. 2017, 7, 185. [Google Scholar] [CrossRef] [PubMed]
- Ralfkiaer, U.; Lindahl, L.M.; Lindal, L.; Litman, T.; Gjerdrum, L.-M.; Ahler, C.B.; Gniadecki, R.; Marstrand, T.; Fredholm, S.; Iversen, L.; et al. MicroRNA expression in early mycosis fungoides is distinctly different from atopic dermatitis and advanced cutaneous T-cell lymphoma. Anticancer Res. 2014, 34, 7207–7217. [Google Scholar] [PubMed]
- Cristofoletti, C.; Picchio, M.C.; Lazzeri, C.; Tocco, V.; Pagani, E.; Bresin, A.; Mancini, B.; Passarelli, F.; Facchiano, A.; Scala, E.; et al. Comprehensive analysis of PTEN status in Sezary syndrome. Blood 2013, 122, 3511–3520. [Google Scholar] [CrossRef] [PubMed]
- Hollander, M.C.; Blumenthal, G.M.; Dennis, P.A. PTEN loss in the continuum of common cancers, rare syndromes and mouse models. Nat. Rev. Cancer 2011, 11, 289–301. [Google Scholar] [CrossRef]
- Kopp, K.L.; Ralfkiaer, U.; Nielsen, B.S.; Gniadecki, R.; Woetmann, A.; Ødum, N.; Ralfkiaer, E. Expression of miR-155 and miR-126 in situ in cutaneous T-cell lymphoma. APMIS 2013, 121, 1020–1024. [Google Scholar] [CrossRef]
- Moyal, L.; Barzilai, A.; Gorovitz, B.; Hirshberg, A.; Amariglio, N.; Jacob-Hirsch, J.; Maron, L.; Feinmesser, M.; Hodak, E. miR-155 is involved in tumor progression of mycosis fungoides. Exp. Dermatol. 2013, 22, 431–433. [Google Scholar] [CrossRef] [Green Version]
- Fredholm, S.; Willerslev-Olsen, A.; Met, Ö.; Kubat, L.; Gluud, M.; Mathiasen, S.L.; Friese, C.; Blümel, E.; Petersen, D.L.; Hu, T.; et al. Special AT rich-binding1 protein (SATB1) in malignant T cells. J. Investig. Dermatol. 2018. [Google Scholar] [CrossRef] [Green Version]
- Herrera, A.; Fredholm, S.; Cheng, A.; Mimitou, E.P.; Seffens, A.; Bar-Natan, M.; Sun, A.; Latkowski, J.-A.; Willerslew-Olsen, A.; Buus, T.B.; et al. Low SATB1 Expression Promotes IL-5 and IL-9 Expression in Sézary Syndrome. J. Investig. Dermatol. 2020, 140, 713–716. [Google Scholar] [CrossRef]
- Moyal, L.; Yehezkel, S.; Gorovitz, B.; Keren, A.; Gilhar, A.; Lubin, I.; Sherman, S.; Hodak, E. Oncogenic role of microRNA-155 in mycosis fungoides: An in vitro and xenograft mouse model study. Br. J. Dermatol. 2017, 177, 791–800. [Google Scholar] [CrossRef] [PubMed]
- Litvinov, I.V.; Cordeiro, B.; Fredholm, S.; Ødum, N.; Zargham, H.; Huang, Y.; Zhou, Y.; Pehr, K.; Kupper, T.S.; Woetmann, A.; et al. Analysis of STAT4 expression in cutaneous T-cell lymphoma (CTCL) patients and patient-derived cell lines. Cell Cycle 2014, 13, 2975–2982. [Google Scholar] [CrossRef] [Green Version]
- Netchiporouk, E.; Litvinov, I.V.; Moreau, L.; Gilbert, M.; Sasseville, D.; Duvic, M. Deregulation in STAT signaling is important for cutaneous T-cell lymphoma (CTCL) pathogenesis and cancer progression. Cell Cycle 2014, 13, 3331–3335. [Google Scholar] [CrossRef] [PubMed]
- Wu, M.; Duan, Q.; Liu, X.; Zhang, P.; Fu, Y.; Zhang, Z.; Liu, L.; Cheng, J.; Jiang, H. MiR-155-5p promotes oral cancer progression by targeting chromatin remodeling gene ARID2. Biomed. Pharmacother. 2020, 122, 109696. [Google Scholar] [CrossRef]
- Seto, A.G.; Beatty, X.; Lynch, J.M.; Hermreck, M.; Tetzlaff, M.; Duvic, M.; Jackson, A.L. Cobomarsen, an oligonucleotide inhibitor of miR-155, co-ordinately regulates multiple survival pathways to reduce cellular proliferation and survival in cutaneous T-cell lymphoma. Br. J. Haematol. 2018, 183, 428–444. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Celik, H.; Koh, W.K.; Kramer, A.C.; Ostrander, E.L.; Mallaney, C.; Fisher, D.A.C.; Xiang, J.; Wilson, W.C.; Martens, A.; Kothari, A.; et al. JARID2 Functions as a Tumor Suppressor in Myeloid Neoplasms by Repressing Self-Renewal in Hematopoietic Progenitor Cells. Cancer Cell 2018, 34, 741–756.e8. [Google Scholar] [CrossRef] [Green Version]
- Su, C.-L.; Deng, T.-R.; Shang, Z.; Xiao, Y. JARID2 inhibits leukemia cell proliferation by regulating CCND1 expression. Int. J. Hematol. 2015, 102, 76–85. [Google Scholar] [CrossRef]
- Litvinov, I.V.; Netchiporouk, E.; Cordeiro, B.; Zargham, H.; Pehr, K.; Gilbert, M.; Zhou, Y.; Moreau, L.; Woetmann, A.; Ødum, N.; et al. Ectopic expression of embryonic stem cell and other developmental genes in cutaneous T-cell lymphoma. Oncoimmunology 2014, 3. [Google Scholar] [CrossRef]
- Nakagawa, R.; Leyland, R.; Meyer-Hermann, M.; Lu, D.; Turner, M.; Arbore, G.; Phan, T.G.; Brink, R.; Vigorito, E. MicroRNA-155 controls affinity-based selection by protecting c-MYC+ B cells from apoptosis. J. Clin. Investig. 2016, 126, 377–388. [Google Scholar] [CrossRef] [Green Version]
- Zargar, S.; Tomar, V.; Shyamsundar, V.; Vijayalakshmi, R.; Somasundaram, K.; Karunagaran, D. A Feedback Loop between MicroRNA 155 (miR-155), Programmed Cell Death 4, and Activation Protein 1 Modulates the Expression of miR-155 and Tumorigenesis in Tongue Cancer. Mol. Cell. Biol. 2019, 39. [Google Scholar] [CrossRef] [Green Version]
- Espadinha, A.-S.; Prouzet-Mauléon, V.; Claverol, S.; Lagarde, V.; Bonneu, M.; Mahon, F.-X.; Cardinaud, B. A tyrosine kinase-STAT5-miR21-PDCD4 regulatory axis in chronic and acute myeloid leukemia cells. Oncotarget 2017, 8, 76174–76188. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cristofoletti, C.; Bresin, A.; Picozza, M.; Picchio, M.C.; Monzo, F.; Helmer Citterich, M.; Passarelli, F.; Frezzolini, A.; Scala, E.; Monopoli, A.; et al. Blood and skin-derived Sezary cells: Differences in proliferation-index, activation of PI3K/AKT/mTORC1 pathway and its prognostic relevance. Leukemia 2019, 33, 1231–1242. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zeng, T.; Zhang, Q.; Yu, X.; Gao, X.; Qiu, Y. Inhibition of cell migration and invasion and promotion of cell apoptosis by overexpression of programmed cell death 4 (PDCD4) in cervical cancer Siha cells. Int. J. Clin. Exp. Pathol. 2018, 11, 4676–4683. [Google Scholar] [PubMed]
- Long, J.; Yin, Y.; Guo, H.; Li, S.; Sun, Y.; Zeng, C.; Zhu, W. The mechanisms and clinical significance of PDCD4 in colorectal cancer. Gene 2019, 680, 59–64. [Google Scholar] [CrossRef] [PubMed]
- Kumar, R.; Selth, L.A.; Schulz, R.B.; Tay, B.S.; Neilsen, P.M.; Callen, D.F. Genome-wide mapping of ZNF652 promoter binding sites in breast cancer cells. J. Cell. Biochem. 2011, 112, 2742–2747. [Google Scholar] [CrossRef] [PubMed]
- Neilsen, P.M.; Noll, J.E.; Mattiske, S.; Bracken, C.P.; Gregory, P.A.; Schulz, R.B.; Lim, S.P.; Kumar, R.; Suetani, R.J.; Goodall, G.J.; et al. Mutant p53 drives invasion in breast tumors through up-regulation of miR-155. Oncogene 2013, 32, 2992–3000. [Google Scholar] [CrossRef] [Green Version]
- Jiang, D.; Aguiar, R.C.T. MicroRNA-155 controls RB phosphorylation in normal and malignant B lymphocytes via the noncanonical TGF-β1/SMAD5 signaling module. Blood 2014, 123, 86–93. [Google Scholar] [CrossRef] [Green Version]
- Rai, D.; Kim, S.-W.; McKeller, M.R.; Dahia, P.L.M.; Aguiar, R.C.T. Targeting of SMAD5 links microRNA-155 to the TGF-beta pathway and lymphomagenesis. Proc. Natl. Acad. Sci. USA 2010, 107, 3111–3116. [Google Scholar] [CrossRef] [Green Version]
- Fuchs, O.; Simakova, O.; Klener, P.; Cmejlova, J.; Zivny, J.; Zavadil, J.; Stopka, T. Inhibition of Smad5 in human hematopoietic progenitors blocks erythroid differentiation induced by BMP4. Blood Cells Mol. Dis. 2002, 28, 221–233. [Google Scholar] [CrossRef]
- Zhang, Y.; Song, J.; Rutenberg, D.; Sokol, L. Hepatic Infiltration with Malignant T-cells Manifesting as Impending Acute Liver Failure in Sezary Syndrome. Mediterr. J. Hematol. Infect. Dis. 2020, 12, e2020007. [Google Scholar] [CrossRef]
- Jiang, H.; Cao, H.-J.; Ma, N.; Bao, W.-D.; Wang, J.-J.; Chen, T.-W.; Zhang, E.-B.; Yuan, Y.-M.; Ni, Q.-Z.; Zhang, F.-K.; et al. Chromatin remodeling factor ARID2 suppresses hepatocellular carcinoma metastasis via DNMT1-Snail axis. Proc. Natl. Acad. Sci. USA 2020, 117, 4770–4780. [Google Scholar] [CrossRef] [PubMed]
- Benoit, B.M.; Jariwala, N.; O’Connor, G.; Oetjen, L.K.; Whelan, T.M.; Werth, A.; Troxel, A.B.; Sicard, H.; Zhu, L.; Miller, C.; et al. CD164 identifies CD4+ T cells highly expressing genes associated with malignancy in Sézary syndrome: The Sézary signature genes, FCRL3, Tox, and miR-214. Arch. Dermatol. Res. 2017, 309, 11–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kohnken, R.; McNeil, B.; Wen, J.; McConnell, K.; Grinshpun, L.; Keiter, A.; Chen, L.; William, B.; Porcu, P.; Mishra, A. Preclinical Targeting of MicroRNA-214 in Cutaneous T-Cell Lymphoma. J. Investig. Dermatol. 2019, 139, 1966–1974.e3. [Google Scholar] [CrossRef] [PubMed]
- Fang, L.-L.; Wang, X.-H.; Sun, B.-F.; Zhang, X.-D.; Zhu, X.-H.; Yu, Z.-J.; Luo, H. Expression, regulation and mechanism of action of the miR-17-92 cluster in tumor cells (Review). Int. J. Mol. Med. 2017, 40, 1624–1630. [Google Scholar] [CrossRef] [Green Version]
- Moyal, L.; Gorovitz-Haris, B.; Yehezkel, S.; Jacob-Hirsch, J.; Bershtein, V.; Barzilai, A.; Rotem, C.; Sherman, S.; Amitay-Laish, I.; Feinmesser, M.; et al. Unilesional mycosis fungoides is associated with increased expression of microRNA-17~92 and T helper 1 skewing. Br. J. Dermatol. 2019, 180, 1123–1134. [Google Scholar] [CrossRef]
- Hanahan, D.; Weinberg, R.A. The Hallmarks of Cancer. Cell 2000, 100, 57–70. [Google Scholar] [CrossRef] [Green Version]
- Xia, L.; Wu, L.; Xia, H.; Bao, J.; Li, Q.; Chen, X.; Xia, R. miR-337 suppresses cutaneous T-cell lymphoma via the STAT3 pathway. Cell Cycle 2019, 18, 1635–1645. [Google Scholar] [CrossRef]
- Ito, M.; Teshima, K.; Ikeda, S.; Kitadate, A.; Watanabe, A.; Nara, M.; Yamashita, J.; Ohshima, K.; Sawada, K.; Tagawa, H. MicroRNA-150 inhibits tumor invasion and metastasis by targeting the chemokine receptor CCR6, in advanced cutaneous T-cell lymphoma. Blood 2014, 123, 1499–1511. [Google Scholar] [CrossRef] [Green Version]
- Kitadate, A.; Ikeda, S.; Teshima, K.; Ito, M.; Toyota, I.; Hasunuma, N.; Takahashi, N.; Miyagaki, T.; Sugaya, M.; Tagawa, H. MicroRNA-16 mediates the regulation of a senescence-apoptosis switch in cutaneous T-cell and other non-Hodgkin lymphomas. Oncogene 2016, 35, 3692–3704. [Google Scholar] [CrossRef]
- Gupta, S.; Silveira, D.A.; Barbé-Tuana, F.M.; Mombach, J.C.M. Integrative data modeling from lung and lymphatic cancer predicts functional roles for miR-34a and miR-16 in cell fate regulation. Sci. Rep. 2020, 10, 2511. [Google Scholar] [CrossRef] [Green Version]
- Wei, H.; Liu, R.; Guo, X.; Zhou, Y.; Sun, B.; Wang, J. miRNA-135a regulates Hut78 cell proliferation via the GATA-3/TOX signaling pathway. Mol. Med. Rep. 2019, 19, 2361–2367. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McGirt, L.Y.; Adams, C.M.; Baerenwald, D.A.; Zwerner, J.P.; Zic, J.A.; Eischen, C.M. miR-223 regulates cell growth and targets proto-oncogenes in mycosis fungoides/cutaneous T-cell lymphoma. J. Investig. Dermatol. 2014, 134, 1101–1107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ohmatsu, H.; Sugaya, M.; Suga, H.; Morimura, S.; Miyagaki, T.; Kai, H.; Kagami, S.; Fujita, H.; Asano, Y.; Tada, Y.; et al. Serum IL-31 levels are increased in patients with cutaneous T-cell lymphoma. Acta Derm. Venereol. 2012, 92, 282–283. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Möbs, M.; Gryzik, S.; Haidar, A.; Humme, D.; Beyer, M.; Vandersee, S. Analysis of the IL-31 pathway in Mycosis fungoides and Sézary syndrome. Arch. Dermatol. Res. 2015, 307, 479–485. [Google Scholar] [CrossRef] [PubMed]
- Malek, M.; Gleń, J.; Rębała, K.; Kowalczyk, A.; Sobjanek, M.; Nowicki, R.; Ruckemann-Dziurdzińska, K.; Sokołowska-Wojdyło, M. Il-31 does not correlate to pruritus related to early stage cutaneous T-cell lymphomas but is involved in pathogenesis of the disease. Acta Derm. Venereol. 2015, 95, 283–288. [Google Scholar] [CrossRef] [Green Version]
- Han, Q.; Liu, D.; Convertino, M.; Wang, Z.; Jiang, C.; Kim, Y.H.; Luo, X.; Zhang, X.; Nackley, A.; Dokholyan, N.V.; et al. miRNA-711 binds and activates TRPA1 extracellularly to evoke acute and chronic pruritus. Neuron 2018, 99, 449–463.e6. [Google Scholar] [CrossRef] [Green Version]
- Pluta, L.; Yousefi, B.; Damania, B.; Khan, A.A. Endosomal TLR-8 Senses microRNA-1294 Resulting in the Production of NFḱB Dependent Cytokines. Front. Immunol. 2019, 10. [Google Scholar] [CrossRef]
- Bayraktar, R.; Bertilaccio, M.T.S.; Calin, G.A. The Interaction between Two Worlds: MicroRNAs and Toll-Like Receptors. Front. Immunol. 2019, 10. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.-J.; Guo, J.-S.; Li, S.-S.; Wu, X.-B.; Cao, D.-L.; Jiang, B.-C.; Jing, P.-B.; Bai, X.-Q.; Li, C.-H.; Wu, Z.-H.; et al. TLR8 and its endogenous ligand miR-21 contribute to neuropathic pain in murine DRG. J. Exp. Med. 2018, 215, 3019–3037. [Google Scholar] [CrossRef] [Green Version]
- Hanna, J.; Hossain, G.S.; Kocerha, J. The Potential for microRNA Therapeutics and Clinical Research. Front. Genet. 2019, 10. [Google Scholar] [CrossRef] [Green Version]
- Ralfkiaer, U.; Hagedorn, P.H.; Bangsgaard, N.; Løvendorf, M.B.; Ahler, C.B.; Svensson, L.; Kopp, K.L.; Vennegaard, M.T.; Lauenborg, B.; Zibert, J.R.; et al. Diagnostic microRNA profiling in cutaneous T-cell lymphoma (CTCL). Blood 2011, 118, 5891–5900. [Google Scholar] [CrossRef] [PubMed]
- Marstrand, T.; Ahler, C.B.; Ralfkiaer, U.; Clemmensen, A.; Kopp, K.L.; Sibbesen, N.A.; Krejsgaard, T.; Litman, T.; Wasik, M.A.; Bonefeld, C.M.; et al. Validation of a diagnostic microRNA classifier in cutaneous T-cell lymphomas. Leuk. Lymphoma 2014, 55, 957–958. [Google Scholar] [CrossRef] [PubMed]
- Manso, R.; Martínez-Magunacelaya, N.; Eraña-Tomás, I.; Monsalvez, V.; Rodríguez-Peralto, J.L.; Ortiz-Romero, P.-L.; Santonja, C.; Cristóbal, I.; Piris, M.A.; Rodríguez-Pinilla, S.M. Mycosis fungoides progression could be regulated by microRNAs. PLoS ONE 2018, 13, e0198477. [Google Scholar] [CrossRef]
- Dusílková, N.; Bašová, P.; Polívka, J.; Kodet, O.; Kulvait, V.; Pešta, M.; Trněný, M.; Stopka, T. Plasma miR-155, miR-203, and miR-205 are Biomarkers for Monitoring of Primary Cutaneous T-Cell Lymphomas. Int. J. Mol. Sci. 2017, 18, 2136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McGirt, L.; Baerenwald, D.; Vonderheid, E.; Eischen, C. Early changes in miRNA expression are predictive of response to extracorporeal photopheresis in cutaneous T-cell lymphoma. J. Eur. Acad. Derm. Venereol. 2015, 29, 2269–2271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manfè, V.; Biskup, E.; Willumsgaard, A.; Skov, A.G.; Palmieri, D.; Gasparini, P.; Laganá, A.; Woetmann, A.; Ødum, N.; Croce, C.M.; et al. cMyc/miR-125b-5p signalling determines sensitivity to bortezomib in preclinical model of cutaneous T-cell lymphomas. PLoS ONE 2013, 8, e59390. [Google Scholar] [CrossRef]
- Manfè, V.; Biskup, E.; Rosbjerg, A.; Kamstrup, M.; Skov, A.G.; Lerche, C.M.; Lauenborg, B.T.; Odum, N.; Gniadecki, R. miR-122 regulates p53/Akt signalling and the chemotherapy-induced apoptosis in cutaneous T-cell lymphoma. PLoS ONE 2012, 7, e29541. [Google Scholar] [CrossRef] [Green Version]
- Abe, F.; Kitadate, A.; Ikeda, S.; Yamashita, J.; Nakanishi, H.; Takahashi, N.; Asaka, C.; Teshima, K.; Miyagaki, T.; Sugaya, M.; et al. Histone deacetylase inhibitors inhibit metastasis by restoring a tumor suppressive microRNA-150 in advanced cutaneous T-cell lymphoma. Oncotarget 2017, 8, 7572–7585. [Google Scholar] [CrossRef] [Green Version]
- Krejsgaard, T.; Lindahl, L.M.; Mongan, N.P.; Wasik, M.A.; Litvinov, I.V.; Iversen, L.; Langhoff, E.; Woetmann, A.; Odum, N. Malignant inflammation in cutaneous T-cell lymphoma—A hostile takeover. Semin. Immunopathol. 2017, 39, 269–282. [Google Scholar] [CrossRef]
- Mishra, A.; La Perle, K.; Kwiatkowski, S.; Sullivan, L.A.; Sams, G.H.; Johns, J.; Curphey, D.P.; Wen, J.; McConnell, K.; Qi, J.; et al. Mechanism, Consequences, and Therapeutic Targeting of Abnormal IL15 Signaling in Cutaneous T-cell Lymphoma. Cancer Discov. 2016, 6, 986–1005. [Google Scholar] [CrossRef] [Green Version]
- Kohnken, R.; Wen, J.; Mundy-Bosse, B.; McConnell, K.; Keiter, A.; Grinshpun, L.; Hartlage, A.; Yano, M.; McNeil, B.; Chakravarti, N.; et al. Diminished microRNA-29b level is associated with BRD4-mediated activation of oncogenes in cutaneous T-cell lymphoma. Blood 2018, 131, 771–781. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Querfeld, C.; Foss, F.M.; Kim, Y.H.; Pinter-Brown, L.; William, B.M.; Porcu, P.; Pacheco, T.; Haverkos, B.M.; DeSimone, J.; Guitart, J.; et al. Phase 1 Trial of Cobomarsen, an Inhibitor of Mir-155, in Cutaneous T Cell Lymphoma. Blood 2018, 132, 2903. [Google Scholar] [CrossRef]
- Querfeld, C.; Foss, F.M.; Pinter-Brown, L.C.; Porcu, P.; William, B.M.; Pacheco, T.; Haverkos, B.M.; Kim, Y.H.; Guitart, J.; Halwani, A.S.; et al. Phase 1 Study of the Safety and Efficacy of MRG-106, a Synthetic Inhibitor of microRNA-155, in CTCL Patients. Blood 2017, 130, 820. [Google Scholar]
- Nielsen, M.; Nissen, M.H.; Gerwien, J.; Zocca, M.-B.; Rasmussen, H.M.; Nakajima, K.; Röpke, C.; Geisler, C.; Kaltoft, K.; Ødum, N. Spontaneous interleukin-5 production in cutaneous T-cell lymphoma lines is mediated by constitutively activated Stat3. Blood 2002, 99, 973–977. [Google Scholar] [CrossRef]
- Verma, N.K.; Davies, A.M.; Long, A.; Kelleher, D.; Volkov, Y. STAT3 knockdown by siRNA induces apoptosis in human cutaneous T-cell lymphoma line Hut78 via downregulation of Bcl-xL. Cell. Mol. Biol. Lett. 2010, 15, 342–355. [Google Scholar] [CrossRef]
- Cremer, T.J.; Fatehchand, K.; Shah, P.; Gillette, D.; Patel, H.; Marsh, R.L.; Besecker, B.Y.; Rajaram, M.V.S.; Cormet-Boyaka, E.; Kanneganti, T.-D.; et al. MiR-155 induction by microbes/microbial ligands requires NF-κB-dependent de novo protein synthesis. Front. Cell Infect. Microbiol. 2012, 2, 73. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| miR-155 Targets | Role in Cancers | Validated Functions | Fold Change | References |
|---|---|---|---|---|
| JARID2 | Tumor suppressor in leukemia | Reduces proliferation and survival | 2.2 | [66,67,68,69] |
| PDCD4 | Tumor suppressor in leukemia, tongue-, colorectal- and cervical cancer, mutated in SS | Increases apoptosis Induces cell cycle arrest | 2.0 | [70,71,72,73,74] |
| ZNF652 | Tumor suppressor in breast cancer | Represses drivers of invasion and metastasis | 3.4 | [75,76] |
| SMAD5 | Tumor suppressor in leukemia and B-cell lymphoma | Plays a role in growth inhibition of malignant cells | 2.2 | [77,78,79] |
| ARID2 | Tumor suppressor in hepatocellular carcinoma and oral cancer, mutated in SS | Suppresses growth and progression of tumor cells | 2.1 | [64,80,81] |
| MicroRNA | Status in CTCL | Validated Functions |
|---|---|---|
| miR-16 | Downregulated in CTCL Upregulated by HDAC inhibition | Increases p21 and cell cycle arrest |
| miR-17/92 | Upregulated in tumor MF Reduced in SS | Influences cytokines in MF Tumor suppressive properties in SS |
| miR-21 | Upregulated in CTCL Driven by STAT signaling | Targets PTEN Increases cell survival |
| miR-22 | Downregulated in CTCL Repressed by STAT signaling | Suppresses c-Myc |
| miR-29b | Downregulated in CTCL Reduced by IL-15 | Suppresses BRD4 |
| miR-93 | Upregulated in tumor MF Reduced in SS | Represses p21 expression Induces proliferation in MF |
| miR-122 | Upregulated in CTCL | Decreases sensitivity to chemotherapy |
| miR-125b | OncomiR in CTCL Regulated by c-Myc | Represses sensitivity to Bortezomib |
| miR-135a | Downregulated in CTCL | Suppresses GATA3 |
| miR-150 | Downregulated in advanced CTCL Upregulated by HDAC inhibition | Shapes the inflammatory environment Inhibits migration |
| miR-155 | Upregulated in CTCL Driven by STAT signaling | Increases proliferation and survival Targets multiple pathways |
| miR-200c | Downregulated in CTCL Silenced by promotor methylation | Inhibits Notch signaling |
| miR-214 | Upregulated in SS Driven by TWIST1 and BRD4 | Promotes cell survival Increases disease severity in mouse models |
| miR-223 | Downregulated in CTCL | Suppresses E2F1 Reduces cell growth |
| miR-337 | Downregulated in CTCL | Targets STAT3 signaling Inhibits cell viability |
| miR-711 | Upregulated in CTCL | Causes pruritus |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gluud, M.; Willerslev-Olsen, A.; Gjerdrum, L.M.R.; Lindahl, L.M.; Buus, T.B.; Andersen, M.H.; Bonefeld, C.M.; Krejsgaard, T.; Litvinov, I.V.; Iversen, L.; et al. MicroRNAs in the Pathogenesis, Diagnosis, Prognosis and Targeted Treatment of Cutaneous T-Cell Lymphomas. Cancers 2020, 12, 1229. https://doi.org/10.3390/cancers12051229
Gluud M, Willerslev-Olsen A, Gjerdrum LMR, Lindahl LM, Buus TB, Andersen MH, Bonefeld CM, Krejsgaard T, Litvinov IV, Iversen L, et al. MicroRNAs in the Pathogenesis, Diagnosis, Prognosis and Targeted Treatment of Cutaneous T-Cell Lymphomas. Cancers. 2020; 12(5):1229. https://doi.org/10.3390/cancers12051229
Chicago/Turabian StyleGluud, Maria, Andreas Willerslev-Olsen, Lise Mette Rahbek Gjerdrum, Lise M. Lindahl, Terkild B. Buus, Mads Hald Andersen, Charlotte Menne Bonefeld, Thorbjorn Krejsgaard, Ivan V. Litvinov, Lars Iversen, and et al. 2020. "MicroRNAs in the Pathogenesis, Diagnosis, Prognosis and Targeted Treatment of Cutaneous T-Cell Lymphomas" Cancers 12, no. 5: 1229. https://doi.org/10.3390/cancers12051229
APA StyleGluud, M., Willerslev-Olsen, A., Gjerdrum, L. M. R., Lindahl, L. M., Buus, T. B., Andersen, M. H., Bonefeld, C. M., Krejsgaard, T., Litvinov, I. V., Iversen, L., Becker, J. C., Persson, J. L., Koralov, S. B., Litman, T., Geisler, C., Woetmann, A., & Odum, N. (2020). MicroRNAs in the Pathogenesis, Diagnosis, Prognosis and Targeted Treatment of Cutaneous T-Cell Lymphomas. Cancers, 12(5), 1229. https://doi.org/10.3390/cancers12051229