Advantages of Using Paclitaxel in Combination with Oncolytic Adenovirus Utilizing RNA Destabilization Mechanism

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Construction of the Virus for Selective Replication in Cancer Cells
2.2. In Vitro Cytolytic Potential of Ad-fosARE
2.3. Comparison of the Oncolytic Effects of Ad-fosARE and dl1520
2.4. The Potential Synergistic Activity of Paclitaxel
2.5. Synergistic Effect of Oncolytic Adenovirus and Paclitaxel
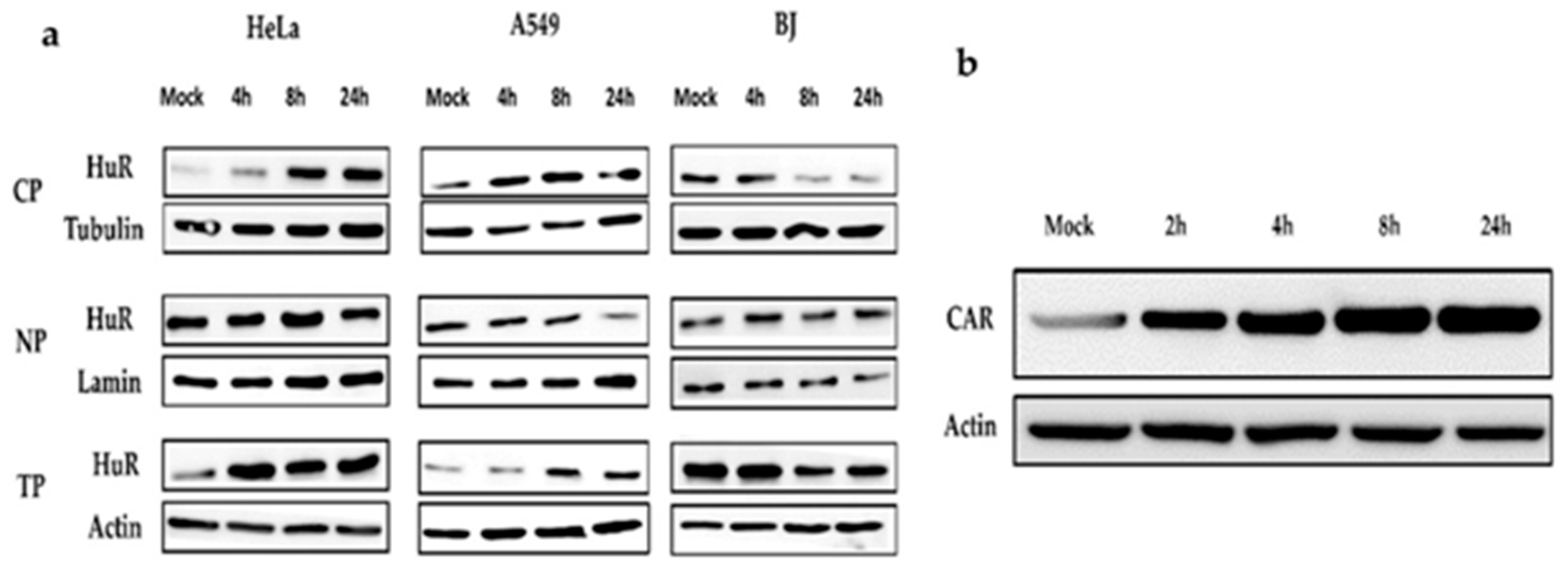
2.6. Effects of Paclitaxel Treatment on Ad-fosARE Viral Protein Synthesis and mRNA Stabilization
2.7. Combination Treatment Increases the Level of Post-Translationally Modified Tubulin
2.8. Effects of Paclitaxel on the Cell Lysis Activity of the Virus
2.9. In Vivo Synergistic Effect of Ad-fosARE and PTX in Murine Flank Tumor
3. Discussion
4. Materials and Methods
4.1. Cell Lines
4.2. Construction of Ad-fosARE
4.3. Drug, Reagents, and Antibodies
4.4. Immunocytochemistry
4.5. Preparation of Ad-fosARE, WT300 and dl1520 Lysates
4.6. Cytopathic Effect (CPE) Assay and Cell Viability Assay
4.7. In Vitro Virus Proliferation Assay
4.8. Western Blot Analysis
4.9. HuR Depletion
4.10. RNA Extraction and Quantitative Real-Time RT-PCR
4.11. In Vivo Analysis
4.12. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Alemany, R.; Balague, C.; Curiel, D.T. Replicative adenoviruses for cancer therapy. Nat. Biotechnol. 2000, 18, 723–727. [Google Scholar] [CrossRef]
- Kirn, D. Replication-selective Oncolytic Adenoviruses: Virotherapy aimed at genetic targets in cancer. Oncogene 2000, 19, 6660–6669. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abou El Hassan, M.A.; Van der Meulen-Muileman, I.; Abbas, S.; Kruyt, F.A. Conditionally replicating adenoviruses kill tumor cells via a basic apoptotic machinery—independent mechanism that resembles necrosis-like programmed cell death. J. Virol. 2004, 78, 12243–12251. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Espel, E. The Role of the AU-rich Elements of mRNAs in Controlling translation. Semin. Cell Dev. Biol. 2005, 16, 59–67. [Google Scholar] [CrossRef] [PubMed]
- Kruys, V.; Wathelet, M.; Poupart, P.; Contreras, R.; Fiers, W.; Content, J.; Huez, G. The 3 untranslated region of the human interferon-beta mRNA has an inhibitory effect on translation. Proc. Natl. Acad. Sci. USA 1987, 84, 6030–6034. [Google Scholar] [CrossRef] [Green Version]
- Kruys, V.; Marinx, O.; Shaw, G.; Deschamps, J.; Huez, G. Translational blockade imposed by cytokine-derived UA-rich sequences. Science 1989, 245, 852–855. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.Y.; Shyu, A.B. AU-rich elements: Characterization and importance in mRNA degradation. Trends Biochem. Sci. 1995, 20, 465–470. [Google Scholar] [CrossRef]
- Nadar, M.; Chan, M.Y.; Huang, S.W.; Huang, C.C.; Tseng, J.T.; Tsai, C.H. HuR binding to AU-rich elements present in the 3’ untranslated region of Classical swine fever virus. Virol. J. 2011, 8, 340. [Google Scholar] [CrossRef] [Green Version]
- DeMaria, C.T.; Brewer, G. AUF1 binding affinity to A+U-rich elements correlates with rapid mRNA degradation. J. Biol. Chem. 1996, 271, 12179–12184. [Google Scholar] [CrossRef] [Green Version]
- Yeh, C.H.; Hung, L.Y.; Hsu, C.; Le, S.Y.; Lee, P.T.; Liao, W.L.; Lin, Y.T.; Chang, W.C.; Tseng, J.T. RNA-binding protein HuR interacts with thrombomodulin 5’untranslated region and represses internal ribosome entry sitemediated translation under IL-1 beta treatment. Mol. Biol. Cell 2008, 19, 3812–3822. [Google Scholar] [CrossRef] [Green Version]
- Habiba, U.; Kitamura, T.; Yanagawa-Matsuda, A.; Higashino, F.; Hida, K.; Totsuka, Y.; Shindoh, M. HuR and podoplanin expression is associated with a high risk of malignant transformation in patients with oral preneoplastic lesion. Oncol. Lett. 2016, 5, 3199–3207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fan, X.C.; Steitz, J.A. HNS, a nuclear-cytoplasmic shuttling sequence in HuR. Proc. Natl. Acad. Sci. USA 1998, 95, 15293–15298. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brennan, C.M.; Steitz, J.A. HuR and mRNA stability. Cell. Mol. Life Sci. CMLS 2001, 58, 266–277. [Google Scholar] [CrossRef] [PubMed]
- De Silanes, I.L.; Lal, A.; Gorospe, M. HuR: Post-transcriptional paths to malignancy. RNA Boil. 2005, 2, 11–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Silanes, I.L.; Fan, J.S.; Yang, X.L.; Zonderman, A.B.; Potapova, O.; Pizer, E.S.; Gorospe, M. Role of the RNA-binding protein HuR in colon carcinogenesis. Oncogene 2003, 22, 7146–7154. [Google Scholar] [CrossRef]
- Ingemarsdotter, C.K.; Baird, S.K.; Connell, C.M.; Öberg, D.; Halldén, G.; McNeish, I.A. Low-dose paclitaxel synergizes with oncolytic adenoviruses via mitotic slippage and apoptosis in ovarian cancer. Oncogene 2010, 29, 6051–6606. [Google Scholar] [CrossRef] [Green Version]
- McGuire, W.P.; Hoskins, W.J.; Brady, M.F.; Kucera, P.R.; Partridge, E.E.; Look, K.Y.; Clarke-Pearson, D.L.; Davidson, M. Cyclophosphamide and cisplatin compared with paclitaxel and cisplatin in patients with stage III and stage IV ovarian cancer. N. Engl. J. Med. 1996, 334, 1–6. [Google Scholar] [CrossRef]
- Horwitz, S.B. Mechanism of action of Taxol. Trends Pharmacol. Sci. 1992, 13, 134–136. [Google Scholar] [CrossRef]
- Kelkar, S.A.; Pfister, K.K.; Crystal, R.G.; Leopold, P.L. Cytoplasmic dynein mediates adenovirus binding to microtubules. J. Virol. 2004, 78, 10122–10132. [Google Scholar] [CrossRef] [Green Version]
- Leopold, P.L.; Crystal, R.G. Intracellular trafficking of adenovirus: Many means to many ends. Adv. Drug Deliv. Rev. 2007, 59, 810–821. [Google Scholar] [CrossRef]
- Warren, J.C.; Rutkowski, A.; Cassimeris, L. Infection with replication-deficient adenovirus induces changes in the dynamic instability of host cell microtubules. Mol. Biol. Cell 2006, 17, 3557–3568. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seidman, M.A.; Hogan, S.M.; Wendland, R.L.; Worgall, S.; Crystal, R.G.; Leopold, P.L. Variation in adenovirus receptor expression and adenovirus vector-mediated transgene expression at defined stages of the cell cycle. Mol. Ther. 2001, 4, 13–21. [Google Scholar] [CrossRef] [PubMed]
- Fok, P.T.; Huang, K.C.; Holland, P.C.; Nalbantoglu, J. The Coxsackie and Adenovirus Receptor Binds Microtubules and Plays a Role in Cell Migration. J. Biol. Chem. 2007, 282, 7512–7521. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abdolazimi, Y.; Mojarrad, M.; Pedram, M.; Modarressi, M.H. Analysis of the expression of coxsackievirus and adenovirus receptor in fve colon cancer cell lines. World J. Gastroenterol. 2007, 13, 6365–6369. [Google Scholar] [CrossRef] [PubMed]
- Tomko, R.P.; Xu, R.; Philipson, L. HCAR and MCAR: The human and mouse cellular receptors for subgroup C adenoviruses and group B coxsackieviruses. Proc. Natl. Acad. Sci. USA 1997, 94, 3352–3356. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bergelson, J.M.; Cunningham, J.A.; Droguett, G.; Kurt-Jones, E.A.; Krithivas, A.; Hong, J.S.; Horwitz, M.S.; Crowell, R.L.; Finberg, R.W. Isolation of a common receptor for Coxsackie B viruses and adenoviruses 2 and 5. Science 1997, 275, 1320–1323. [Google Scholar] [CrossRef]
- Badawi, A.; Biyanee, A.; Nasrullah, U.; Winslow, S.; Schmid, T.; Pfeilschifter, J.; Eberhardt, W. Inhibition of IRES-dependent translation of caspase-2 by HuR confers chemotherapeutic drug resistance in colon carcinoma cells. Oncotarget 2018, 9, 18367–18385. [Google Scholar] [CrossRef] [Green Version]
- Cheng, Y.C.; Liou, J.P.; Kuo, C.C.; Lai, W.Y.; Shih, K.H.; Chang, C.Y.; Pan, W.Y.; Tseng, J.T.; Chang, J.Y. MPT0B098, a novel microtubule inhibitor that destabilizes the hypoxia-inducible factor-1α mRNA through decreasing nuclear-cytoplasmic translocation of RNA-binding protein HuR. Mol. Cancer Ther. 2013, 12, 1202–1212. [Google Scholar] [CrossRef] [Green Version]
- Doller, A.; Akool, E.-S.; Huwiler, A.; Müller, R.; Radeke, H.H.; Pfeilschifter, J.; Eberhardt, W. Posttranslational modification of the AU-rich element binding protein HuR by protein kinase Cdelta elicits angiotensin II-induced stabilization and nuclear export of cyclooxygenase 2 mRNA. Mol. Cell. Biol. 2008, 28, 2608–2625. [Google Scholar] [CrossRef] [Green Version]
- Holt, C.E.; Bullock, S.L. Subcellular mRNA localization in animal cells and why it matters. Science 2009, 326, 1212–1216. [Google Scholar] [CrossRef] [Green Version]
- Blower, M.D. Molecular insights into intracellular RNA localization. Int. Rev. Cell. Mol. Biol. 2013, 302, 1–39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eberhardt, W.; Badawi, A.; Biyanee, A.; Pfeilschifter, J. Cytoskeleton-Dependent Transport as a Potential Target for Interfering with Post-transcriptional HuR mRNA Regulons. Front. Pharmacol. 2016, 7, 251. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jehung, J.P.; Kitamura, T.; Matsuda, A.Y.; Kuroshima, T.; Towfik, A.; Yasuda, M.; Sano, H.; Kitagawa, Y.; Minowa, K.; Shindho, M.; et al. Adenovirus infection induces HuR relocalization to facilitate virus replication. BBRC 2018, 495, 1795–1800. [Google Scholar] [CrossRef] [PubMed]
- Tazawa, H.; Kuroda, S.; Hasei, J.; Kagawa, S.; Fujiwara, T. Impact of Autophagy in Oncolytic Adenoviral Therapy for Cancer. IJMS 2017, 18, 1479. [Google Scholar] [CrossRef] [Green Version]
- Bischoff, J.R.; Kirn, D.H.; Williams, A.; Heise, C.; Horn, S.; Muna, M.; Ng, L.; Nye, J.A.; Sampson-Johannes, A.; Fattaey, A.; et al. An adenovirus mutant that replicates selectively in p53-defcient human tumor cells. Science 1996, 274, 373–376. [Google Scholar] [CrossRef]
- Kasuya, H.; Takeda, S.; Nomoto, S.; Nakao, A. The potential of oncolytic virus therapy for pancreatic cancer. Cancer Gene Ther. 2005, 12, 725–736. [Google Scholar] [CrossRef]
- Chou, C.T. Drug Combination Studies and Their Synergy Quantification Using the Chou-Talalay Method. Cancer Res. 2010, 70, 440–446. [Google Scholar] [CrossRef] [Green Version]
- Doller, A.; Schulz, S.; Pfeilschifter, J.; Eberhardt, W. RNA-dependent association with myosin IIA promotes F-actin-guided trafficking of the ELAV-like protein HuR to polysomes. Nucleic Acids Res. 2013, 41, 9152–9167. [Google Scholar] [CrossRef] [Green Version]
- Habiba, U.; Kuroshima, T.; Yanagawa-Matsuda, A.; Kitamura, T.; Chowdhury, A.; Jehung, J.P.; Hossain, E.; Sano, H.; Kitagawa, Y.; Shindoh, M.; et al. HuR translocation to the cytoplasm of cancer cells in actin-independent manner. Exp. Cell Res. 2018, 369, 218–225. [Google Scholar] [CrossRef]
- Fujiwara, Y.; Kasashima, K.; Saito, K.; Fukuda, M.; Fukao, A.; Sasano, Y.; Inoue, K.; Fujiwara, T.; Sakamoto, H. Microtubule association of a neuronal RNA-binding protein HuD through its binding to the light chain of MAP1B. Biochimie 2011, 93, 817–822. [Google Scholar] [CrossRef]
- Subbaramaiah, K.; Marmo, T.P.; Dixon, D.A.; Dannenberg, A.J. Regulation of cyclooxgenase-2 mRNA stability by taxanes: Evidence for involvement of p38, MAPKAPK-2, and HuR. J. Biol. Chem. 2003, 278, 37637–37647. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kotta-Loizou, I.; Vasilopoulos, S.N.; Coutts, R.H.; Theocharis, S. Current Evidence and Future Perspectives on HuR and Breast Cancer Development, Prognosis, and Treatment. Neoplasia 2016, 18, 674–688. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsuoka, H.; Furusawa, M.; Tomoda, H.; Seo, Y. Difference in cytotoxicity of paclitaxel against neoplastic and normal cells. Anticancer Res. 1994, 14, 163–167. [Google Scholar] [PubMed]
- Cordes, N.; Plasswilm, L. Cell line and schedule-dependent cytotoxicity of paclitaxel (Taxol): Role of the solvent Cremophor EL/ethanol. Anticancer Res. 1998, 18, 1851–18577. [Google Scholar] [PubMed]
- Lin, S.F.; Gao, S.P.; Price, D.L.; Li, S.; Chou, T.C.; Singh, P.; Huang, Y.Y.; Fong, Y.; Wong, R.J. Synergy of a herpes oncolytic virus and paclitaxel for anaplastic thyroid cancer. Clin. Cancer Res. 2008, 14, 1519–1528. [Google Scholar] [CrossRef] [Green Version]
- Khabar, K.S. Hallmarks of cancer and AU-rich elements. Wiley Interdiscip Rev. RNA 2017, 8, e1368. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Guo, Y.; Chu, H.; Guan, Y.; Bi, J.; Wang, B. Multiple functions of the RNA-binding protein HuR in cancer progression, treatment responses and prognosis. Int. J. Mol. Sci. 2013, 14, 10015–10041. [Google Scholar] [CrossRef] [Green Version]








© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hossain, E.; Habiba, U.; Yanagawa-Matsuda, A.; Alam, A.; Ahmed, I.; Towfik Alam, M.; Yasuda, M.; Higashino, F. Advantages of Using Paclitaxel in Combination with Oncolytic Adenovirus Utilizing RNA Destabilization Mechanism. Cancers 2020, 12, 1210. https://doi.org/10.3390/cancers12051210
Hossain E, Habiba U, Yanagawa-Matsuda A, Alam A, Ahmed I, Towfik Alam M, Yasuda M, Higashino F. Advantages of Using Paclitaxel in Combination with Oncolytic Adenovirus Utilizing RNA Destabilization Mechanism. Cancers. 2020; 12(5):1210. https://doi.org/10.3390/cancers12051210
Chicago/Turabian StyleHossain, Elora, Umma Habiba, Aya Yanagawa-Matsuda, Arefin Alam, Ishraque Ahmed, Mohammad Towfik Alam, Motoaki Yasuda, and Fumihiro Higashino. 2020. "Advantages of Using Paclitaxel in Combination with Oncolytic Adenovirus Utilizing RNA Destabilization Mechanism" Cancers 12, no. 5: 1210. https://doi.org/10.3390/cancers12051210