RNF128 Promotes Invasion and Metastasis Via the EGFR/MAPK/MMP-2 Pathway in Esophageal Squamous Cell Carcinoma


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
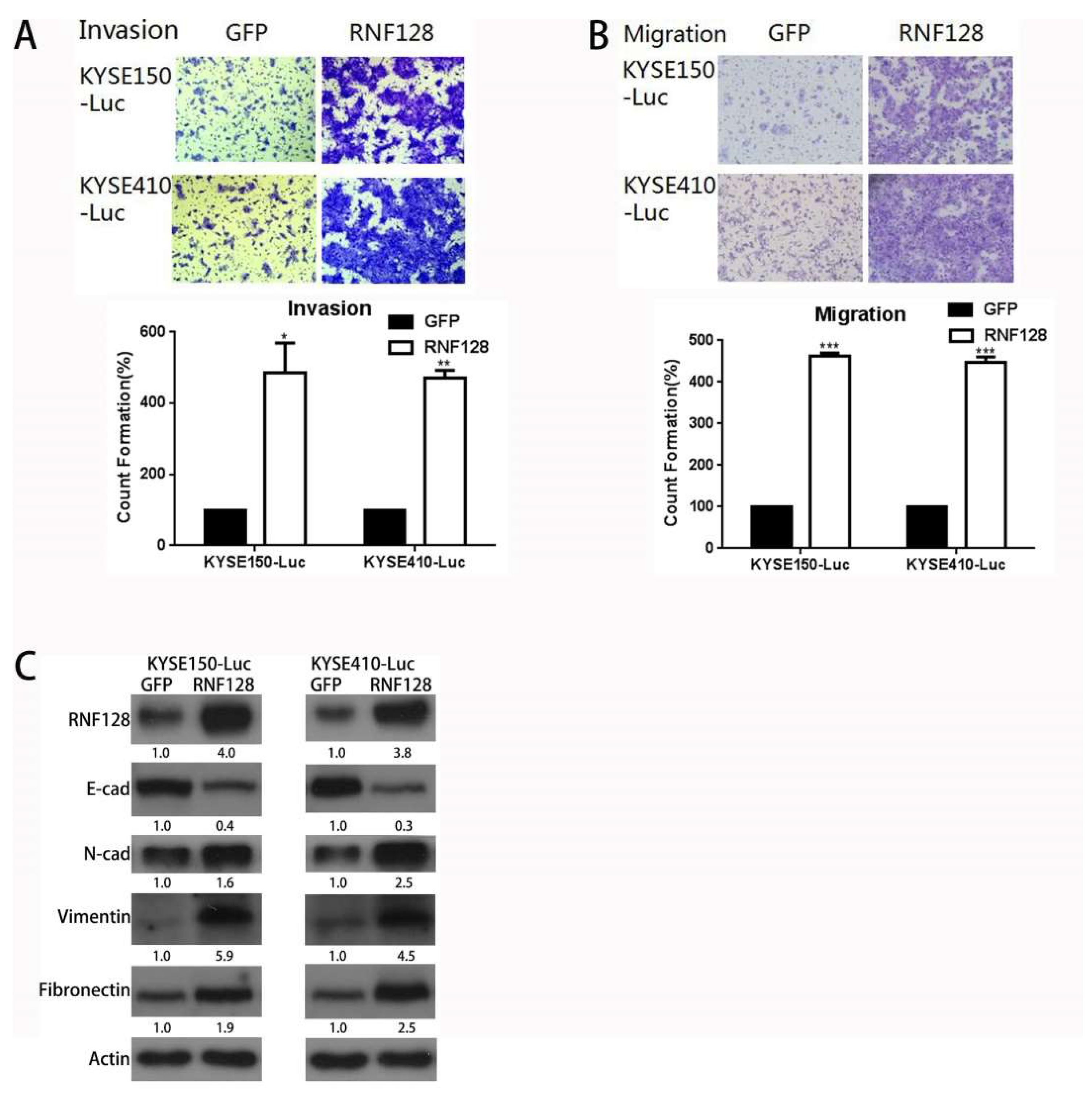
2.1. RNF128 Promoted ESCC Cell Migration and Invasion In Vitro
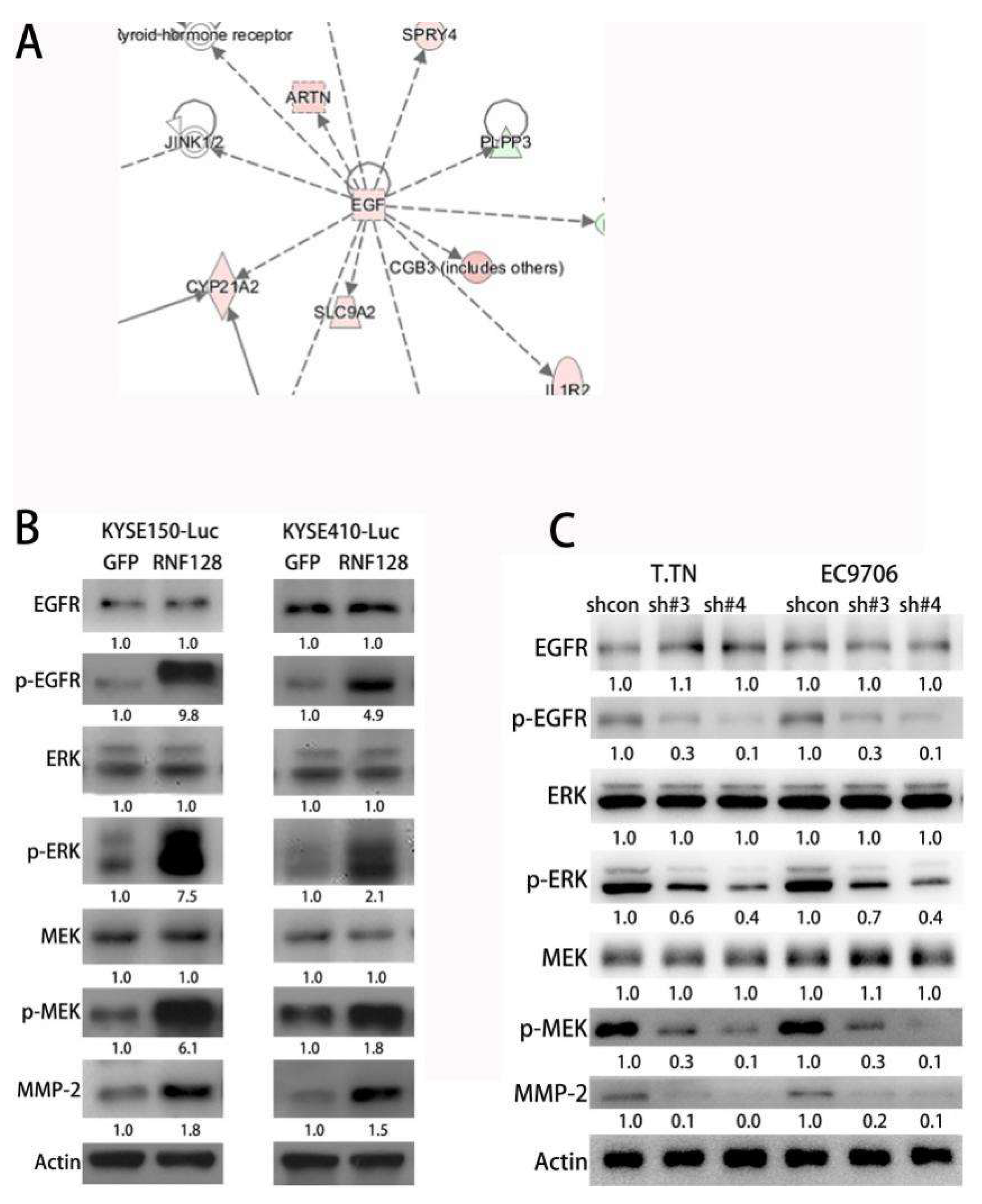
2.2. Transcriptome Sequencing Identified the Activation of EGFR/MAPK/MMP-2 Signaling Induced by RNF128
2.3. RNF128 Drove ESCC Cell Invasion and Metastasis through the EGFR/MAPK/MMP-2 Signaling Pathway
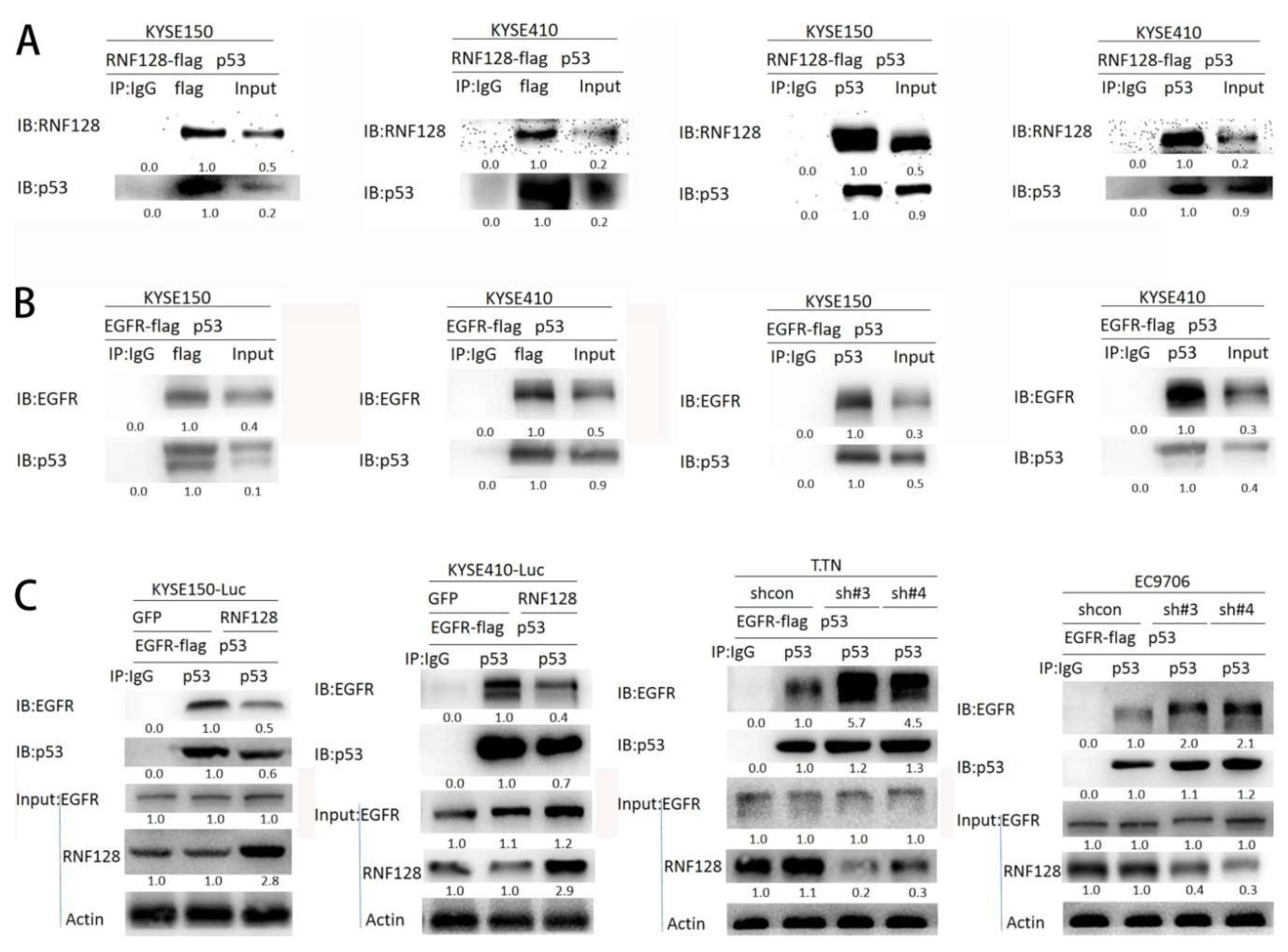
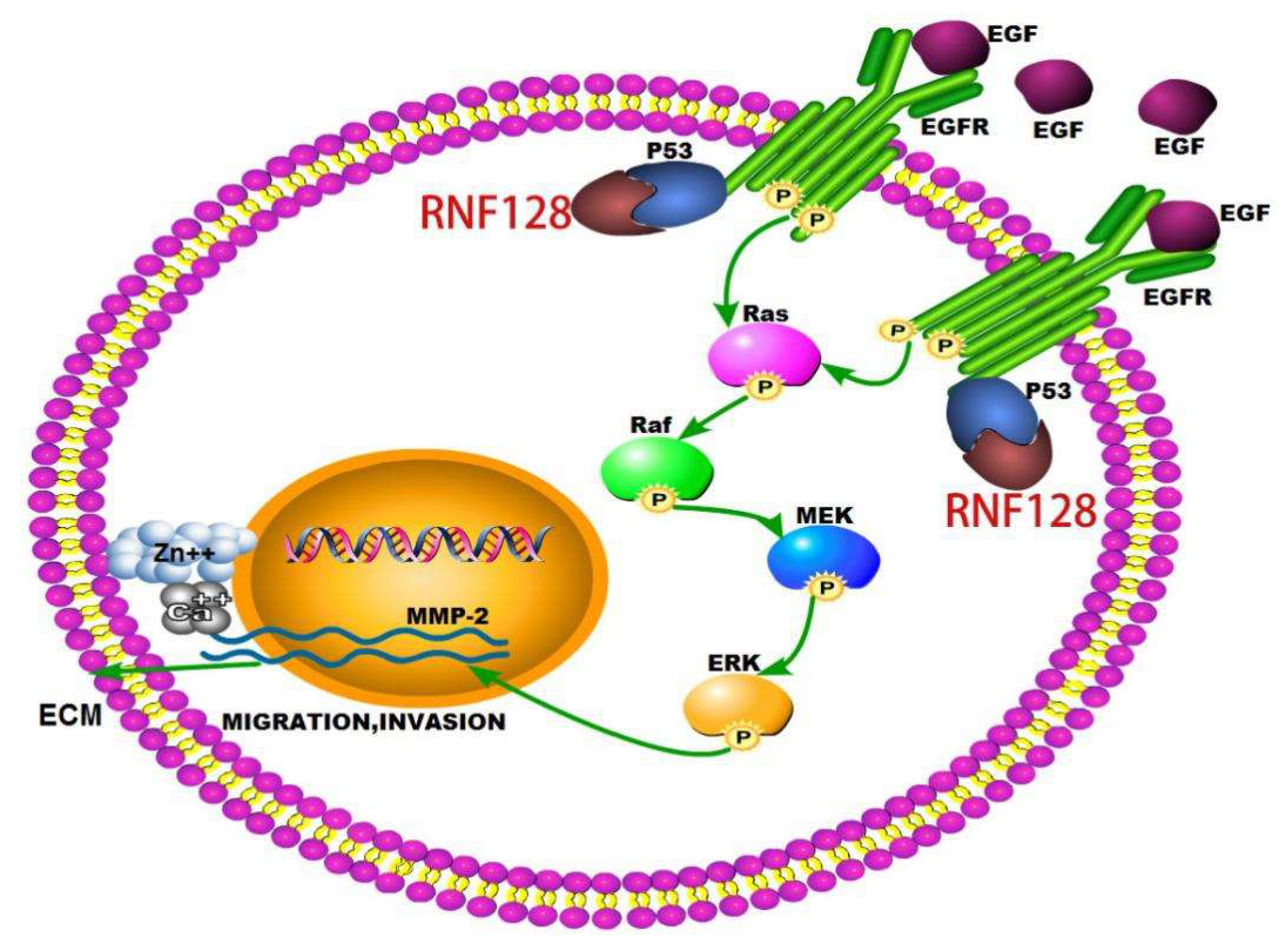
2.4. RNF128 Promoted the Activation of EGFR/MAPK/MMP-2 Pathway by Interacting with p53 and p53 Interacting with EGFR
2.5. RNF128 Promoted Tumor Metastasis In Vivo
3. Discussion
4. Materials and Methods
4.1. Cell Lines and Culture
4.2. Transduction
4.3. Western Blot Analysis
4.4. In Vitro Transwell Assay of Cell Migration and Invasion
4.5. Co-Immunoprecipitation (CoIP) Assay
4.6. In Vivo Tumor Metastatic Assay
4.7. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Global Burden of Disease Cancer Collaboration. The Global Burden of Cancer 2013. JAMA Oncol. 2015, 1, 505–527. [Google Scholar] [CrossRef]
- Torre, L.A.; Bray, F.; Siegel, R.L.; Ferlay, J.; Lortet-Tieulent, J.; Jemal, A. Global cancer statistics, 2012. CA Cancer J. Clin. 2015, 65, 87–108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arnold, M.; Soerjomataram, I.; Ferlay, J.; Forman, D. Global incidence of oesophageal cancer by histological subtype in 2012. Gut 2015, 64, 381–387. [Google Scholar] [CrossRef] [PubMed]
- Enzinger, P.C.; Mayer, R.J. Esophageal cancer. N. Engl. J. Med. 2003, 349, 2241–2252. [Google Scholar] [CrossRef] [PubMed]
- Pennathur, A.; Gibson, M.K.; Jobe, B.A.; Luketich, J.D. Oesophageal carcinoma. Lancet 2013, 381, 400–412. [Google Scholar] [CrossRef] [Green Version]
- Anandasabapathy, N.; Ford, G.S.; Bloom, D.; Holness, C.; Paragas, V.; Seroogy, C.; Skrenta, H.; Hollenhorst, M.; Fathman, C.G.; Soares, L. GRAIL: An E3 ubiquitin ligase that inhibits cytokine gene transcription is expressed in anergic CD4+ T cells. Immunity 2003, 18, 535–547. [Google Scholar] [CrossRef]
- Seroogy, C.M.; Soares, L.; Ranheim, E.A.; Su, L.; Holness, C.; Bloom, D.; Fathman, C.G. The Gene Related to Anergy in Lymphocytes, an E3 Ubiquitin Ligase, Is Necessary for Anergy Induction in CD4 T Cells. J. Immunol. 2004, 173, 79–85. [Google Scholar] [CrossRef] [Green Version]
- Heissmeyer, V.; Macián, F.; Im, S.-H.; Varma, R.; Feske, S.; Venuprasad, K.; Gu, H.; Liu, Y.-C.; Dustin, M.L.; Rao, A. Calcineurin imposes T cell unresponsiveness through targeted proteolysis of signaling proteins. Nat. Immunol. 2004, 5, 255–265. [Google Scholar] [CrossRef]
- Nurieva, R.I.; Zheng, S.; Jin, W.; Chung, Y.; Zhang, Y.; Martinez, G.J.; Reynolds, J.M.; Wang, S.-L.; Lin, X.; Sun, S.-C.; et al. The E3 ubiquitin ligase GRAIL regulates T cell tolerance and regulatory T cell function by mediating T cell receptor-CD3 degradation. Immunity 2010, 32, 670–680. [Google Scholar] [CrossRef]
- Sahoo, A.; Alekseev, A.; Obertas, L.; Nurieva, R. Grail controls Th2 cell development by targeting STAT6 for degradation. Nat. Commun. 2014, 5, 4732. [Google Scholar] [CrossRef]
- Whiting, C.C.; Su, L.L.; Lin, J.T.; Fathman, C.G. GRAIL: A unique mediator of CD4 T-lymphocyte unresponsiveness. FEBS J. 2011, 278, 47–58. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.-C.; Chan, J.Y.-H.; Chiu, Y.-L.; Liu, S.-T.; Lozano, G.; Wang, S.-L.; Ho, C.-L.; Huang, S.-M. Grail as a molecular determinant for the functions of the tumor suppressor p53 in tumorigenesis. Cell Death Differ. 2013, 20, 732–743. [Google Scholar] [CrossRef] [PubMed]
- Nakamichi, S.; Senga, Y.; Inoue, H.; Emi, A.; Matsuki, Y.; Watanabe, E.; Hiramatsu, R.; Ogawa, W.; Kasuga, M. Role of the E3 ubiquitin ligase gene related to anergy in lymphocytes in glucose and lipid metabolism in the liver. J. Mol. Endocrinol. 2009, 42, 161–169. [Google Scholar] [CrossRef] [PubMed]
- MacKenzie, D.A.; Seroogy, C.M. Sustained Expression of GRAIL during Hematopoiesis Results in Dysregulated Differentiation. Acta Haematol. 2009, 122, 230–237. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Xia, D.; Dubois, R.N. The Crosstalk of PTGS2 and EGF Signaling Pathways in Colorectal Cancer. Cancers 2011, 3, 3894–3908. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, F.F.; Deng, J.P.; Liu, X.; Li, W.; Zheng, J.N. HCRP-1 regulates cell migration and invasion via EGFR-ERK mediated up-regulation of MMP-2 with prognostic signifcance in human renal cell carcinoma. Sci. Rep. 2015, 5, 13470. [Google Scholar] [CrossRef] [PubMed]
- Ito, H. Prostaglandin E2 Enhances Pancreatic Cancer Invasiveness through an Ets-1-Dependent Induction of Matrix Metalloproteinase-2. Cancer Res. 2004, 64, 7439–7446. [Google Scholar] [CrossRef]
- Sheikh, M.S.; Carrier, F.; Johnson, A.C.; Ogdon, S.E.; Fornace, A.J. Identification of an additional p53-responsive site in the human epidermal growth factor receptor gene promotor. Oncogene 1997, 15, 1095–1101. [Google Scholar] [CrossRef] [Green Version]
- Nishi, H. p53 Homologue p63 represses Epidermal Growth Factor Receptor Expression. J. Boil. Chem. 2001, 276, 41717–41724. [Google Scholar] [CrossRef]
- Radisky, E.S.; Radisky, D.C. Matrix Metalloproteinase-Induced Epithelial-Mesenchymal Transition in Breast Cancer. J. Mammary Gland. Boil. 2010, 15, 201–212. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Ma, J.; Guo, Q.; Duan, F.; Tang, F.; Zheng, P.; Zhao, Z.; Lu, G. Overexpression of MMP-2 and MMP-9 in esophageal squamous cell carcinoma. Dis. Esophagus 2009, 22, 664–667. [Google Scholar] [CrossRef] [PubMed]
- Zhao, G.; Kang, J.; Jiao, K.; Xu, G.; Yang, L.; Tang, S.; Zhang, H.; Wang, Y.; Nie, Y.; Wu, K.; et al. High Expression of GRP78 Promotes Invasion and Metastases in Patients with Esophageal Squamous Cell Carcinoma. Dig. Dis. Sci. 2015, 60, 2690–2699. [Google Scholar] [CrossRef] [PubMed]
- Ludes-Meyers, J.H.; Subler, M.A.; Shivakumar, C.V.; Munoz, R.M.; Jiang, P.; Bigger, J.E.; Brown, D.R.; Deb, S.P.; Deb, S. Transcriptional activation of the human epidermal growth factor receptor promoter by human p53. Mol. Cell. Biol. 1996, 16, 6009–6019. [Google Scholar] [CrossRef] [PubMed] [Green Version]







© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gao, J.; Wang, Y.; Yang, J.; Zhang, W.; Meng, K.; Sun, Y.; Li, Y.; He, Q.-Y. RNF128 Promotes Invasion and Metastasis Via the EGFR/MAPK/MMP-2 Pathway in Esophageal Squamous Cell Carcinoma. Cancers 2019, 11, 840. https://doi.org/10.3390/cancers11060840
Gao J, Wang Y, Yang J, Zhang W, Meng K, Sun Y, Li Y, He Q-Y. RNF128 Promotes Invasion and Metastasis Via the EGFR/MAPK/MMP-2 Pathway in Esophageal Squamous Cell Carcinoma. Cancers. 2019; 11(6):840. https://doi.org/10.3390/cancers11060840
Chicago/Turabian StyleGao, Jing, Yang Wang, Jie Yang, Weixia Zhang, Kun Meng, Yue Sun, Yangjia Li, and Qing-Yu He. 2019. "RNF128 Promotes Invasion and Metastasis Via the EGFR/MAPK/MMP-2 Pathway in Esophageal Squamous Cell Carcinoma" Cancers 11, no. 6: 840. https://doi.org/10.3390/cancers11060840