Human Sinusoidal Subendothelial Cells Regulate Homing and Invasion of Circulating Metastatic Prostate Cancer Cells to Bone Marrow

 ,
,
Abstract
:1. Introduction
2. Results
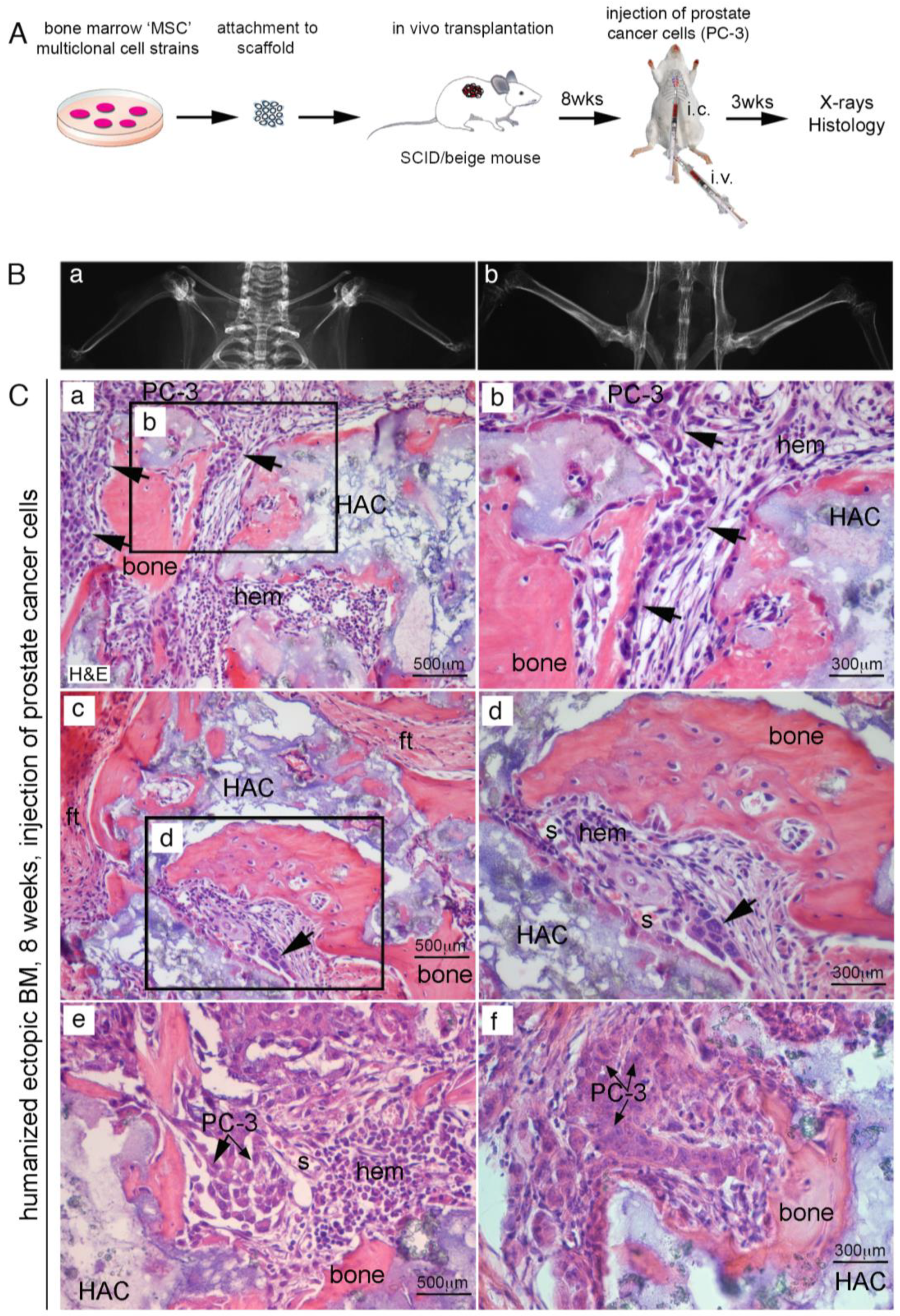
2.1. Homing of Endogenous Hematopoietic Murine Cancer Cells to Human Extraskeletal HME
2.2. Human PC Cells Do Not Efficiently Colonize Adult Mouse Bone/BM
2.3. Efficient Homing of Human PC Cells to Perisinusoidal Niches of Mice-Implanted Heterotopic Bone/BM
2.4. CD146+ hBMSCs Are Sufficient to Establish Human PC-3 Cells Extravasation
3. Discussion
4. Materials and Methods
4.1. Cell Cultures and Reagents
4.2. FACS Analysis
4.3. Immunodeficient Mice
4.4. Lenti-Viral Vectors
4.5. In Vivo Transplantation and In Vivo Experimental Bone Metastases
4.6. Recovery of Tumor Cells from Murine BM
4.7. Immunohistochemistry and Immunolocalization Studies
4.8. CD146 Expression in Tumor Cells
4.9. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Keller, E.T.; Zhang, J.; Cooper, C.R.; Smith, P.C.; McCauley, L.K.; Pient, K.J.; Taichman, R.S. Prostate carcinoma skeletal metastases: Cross-talk between tumor and bone. Cancer Metastasis Rev. 2001, 20, 333–349. [Google Scholar] [CrossRef] [PubMed]
- Ewing, J. Neoplastic Diseases; W.B. Saunders: Philadelphia, PA, USA, 1928. [Google Scholar]
- Paget, S. The distribution of secondary growths in cancer of the breast. Cancer Metastasis Rev. 1989, 8, 98–101. [Google Scholar] [CrossRef]
- Colmone, A.; Amorim, M.; Pontier, A.L.; Wang, S.; Jablonski, E.; Sipkins, D.A. Leukemic cells create bone marrow niches that disrupt the behavior of normal hematopoietic progenitor cells. Science 2008, 322, 1861–1865. [Google Scholar] [CrossRef] [PubMed]
- Sipkins, D.A.; Wei, X.; Wu, J.W.; Runnels, J.M.; Côté, D.; Means, T.K.; Luster, A.D.; Scadden, D.T.; Lin, C.P. In vivo imaging of specialized bone marrow endothelial microdomains for tumour engraftment. Nature 2005, 435, 969–973. [Google Scholar] [CrossRef] [PubMed]
- Kucia, M.; Reca, R.; Miekus, K.; Wanzeck, J.; Wojakowski, W.; Janowska-Wieczorek, A.; Ratajczak, J.; Ratajczak, M.Z. Trafficking of normal stem cells and metastasis of cancer stem cells involve similar mechanisms: Pivotal role of the SDF-1-CXCR4 axis. Stem Cells 2005, 23, 879–894. [Google Scholar] [CrossRef] [PubMed]
- Sacchetti, B.; Funari, A.; Michienzi, S.; Di Cesare, S.; Piersanti, S.; Saggio, I.; Tagliafico, E.; Ferrari, S.; Robey, P.G.; Riminucci, M.; et al. Self-renewing osteoprogenitors in bone marrow sinusoids can organize a hematopoietic microenvironment. Cell 2007, 131, 324–336. [Google Scholar] [CrossRef] [PubMed]
- Yonou, H.; Yokose, T.; Kamijo, T.; Kanomata, N.; Hasebe, T.; Nagai, K.; Hatano, T.; Ogawa, Y.; Ochiai, A. Establishment of a novel species- and tissue-specific metastasis model of human prostate cancer in humanized non-obese diabetic/severe combined immunodeficient mice engrafted with human adult lung and bone. Cancer Res. 2001, 61, 2177–2182. [Google Scholar]
- Nemeth, J.A.; Harb, J.F.; Barroso, U.; He, Z.; Grignon, D.J.; Cher, M.L. Severe combined immunodeficient-hu model of human prostate cancer metastasis to human bone. Cancer Res. 1999, 59, 1987–1993. [Google Scholar]
- Feeley, B.T.; Krenek, L.; Liu, N.; Hsu, W.K.; Gamradt, S.C.; Schwarz, E.M.; Huard, J.; Lieberman, J.R. Overexpression of noggin inhibits BMP-mediated growth of osteolytic prostate cancer lesions. Bone 2006, 38, 154–166. [Google Scholar] [CrossRef]
- Sacchetti, B.; Funari, A.; Remoli, C.; Giannicola, G.; Kogler, G.; Liedtke, S.; Cossu, G.; Serafini, M.; Sampaolesi, M.; Tagliafico, E.; et al. No identical “mesenchymal stem cells” at different times and sites: Committed progenitors of distinct origin and differentiation potential are incorporated as adventitial cells in microvessels. Stem Cell Rep. 2016, 14, 897–913. [Google Scholar] [CrossRef]
- Custer, R.P.; Bosma, G.C.; Bosma, M.J. Severe combined immunodeficiency (SCID) in the mouse. Pathology, reconstitution, neoplasms. Am. J. Pathol. 1985, 120, 464–477. [Google Scholar]
- Singh, A.S.; Figg, W.D. In vivo models of prostate cancer metastasis to bone. J. Urol. 2005, 174, 820–826. [Google Scholar] [CrossRef]
- Karnoub, A.E.; Dash, A.B.; Vo, A.P.; Sullivan, A.; Brooks, M.W.; Bell, G.W.; Richardson, A.L.; Polyak, K.; Tubo, R.; Weinberg, R.A. Mesenchymal stem cells within tumour stroma promote breast cancer metastasis. Nature 2007, 449, 557–563. [Google Scholar] [CrossRef]
- Hall, B.; Dembinski, J.; Sasser, A.K.; Studeny, M.; Andreeff, M.; Marini, F. Mesenchymal stem cells in cancer: Tumor-associated fibroblasts and cell-based delivery vehicles. Int. J. Hematol. 2007, 86, 8–16. [Google Scholar] [CrossRef]
- Cross, N.A.; Papageorgiou, M.; Eaton, C.L. Bone marrow stromal cells promote growth and survival of prostate cancer cells. Biochem. Soc. Trans. 2007, 35, 698–700. [Google Scholar] [CrossRef]
- Correa, D.; Somoza, R.A.; Lin, P.; Schiemann, W.P.; Caplan, A.I. Mesenchymal stem cells regulate melanoma cancer cells extravasation to bone and liver at their perivascular niche. Int. J. Cancer 2016, 138, 417–427. [Google Scholar] [CrossRef]
- Shiozawa, Y.; Eber, M.R.; Berry, J.E.; Taichman, R.S. Bone marrow as a metastatic niche for disseminated tumor cells from solid tumors. BoneKEy Rep. 2015, 20, 689. [Google Scholar] [CrossRef]
- Yang, K.Q.; Liu, Y.; Huang, Q.H.; Mo, N.; Zhang, Q.Y.; Meng, Q.G.; Cheng, J.W. Bone marrow-derived mesenchymal stem cells induced by inflammatory cytokines produce angiogenetic factors and promote prostate cancer growth. BMC Cancer 2017, 17, 878. [Google Scholar] [CrossRef]
- Rossnagl, S.; Ghura, H.; Groth, C.; Altrock, E.; Jakob, F.; Schott, S.; Wimberger, P.; Link, T.; Kuhlmann, J.D.; Stenzl, A.; et al. A Subpopulation of Stromal Cells Controls Cancer Cell Homing to the Bone Marrow. Cancer Res. 2018, 78, 129–142. [Google Scholar] [CrossRef]
- Jinnah, A.H.; Zacks, B.C.; Gwam, C.U.; Kerr, B.A. Emerging and Established Models of Bone Metastasis. Cancers 2018, 10, 176. [Google Scholar] [CrossRef]
- De Wever, O.; Mareel, M. Role of tissue stroma in cancer cell invasion. J. Pathol. 2003, 200, 429–447. [Google Scholar] [CrossRef]
- Gleave, M.E.; Hsieh, J.T.; Von Eschenbach, A.C.; Chung, L.W. Prostate and bone fibroblasts induce human prostate cancer growth in vivo: Implications for bidirectional tumor-stromal cell interaction in prostate carcinoma growth and metastasis. J. Urol. 1992, 147, 1151–1159. [Google Scholar] [CrossRef]
- Wu, G.J.; Wu, M.W.; Wangm, C.; Liu, Y. Enforced expression of METCAM/MUC18 increases tumorigenesis of human prostate cancer LNCaP cells in nude mice. J. Urol. 2011, 185, 1504–1512. [Google Scholar] [CrossRef]
- Zoni, E.; Astrologo, L.; Ng, C.K.Y.; Piscuoglio, S.; Melsen, J.; Grosjean, J.; Klima, I.; Chen, L.; Snaar-Jagalska, E.B.; Flanagan, K.; et al. Therapeutic Targeting of CD146/MCAM Reduces Bone Metastasis in Prostate Cancer. Mol. Cancer Res. 2019, 17, 1049–1062. [Google Scholar] [CrossRef] [Green Version]
- Serafini, M.; Sacchetti, B.; Pievani, A.; Redaelli, D.; Remoli, C.; Biondi, A.; Riminucci, M.; Bianco, P. Establishment of bone marrow and hematopoietic niches in vivo by reversion of chondrocyte differentiation of human bone marrow stromal cells. Stem Cell Res. 2014, 12, 659–672. [Google Scholar] [CrossRef] [Green Version]
- Wu, G.J.; Peng, Q.; Fu, P.; Wang, S.W.; Chiang, C.F.; Dillehay, D.L.; Wu, M.W. Ectopical expression of human MUC18 increases metastasis of human prostate cancer cells. Gene 2004, 327, 201–213. [Google Scholar] [CrossRef]
- Ridge, S.M.; Sullivan, F.J.; Glynn, S.A. Mesenchymal stem cells: Key players in cancer progression. Mol. Cancer 2017, 16, 31. [Google Scholar] [CrossRef]
- Kuznetsov, S.A.; Krebsbach, P.H.; Satomura, K.; Kerr, J.; Riminucc, M.; Benayahu, D.; Robey, P.G. Single-colony derived strains of human marrow stromal fibroblasts form bone after transplantation in vivo. J. Bone Min. Res. 1997, 12, 1335–1347. [Google Scholar] [CrossRef]
- Piersanti, S.; Sacchetti, B.; Funari, A.; Di Cesare, S.; Bonci, D.; Cherubini, G.; Peschle, C.; Riminucci, M.; Bianco, P.; Saggio, I. Lentiviral transduction of human postnatal skeletal (stromal, mesenchymal) stem cells: In vivo transplantation and gene silencing. Calcif. Tissue Int. 2006, 78, 372–384. [Google Scholar] [CrossRef]
- Krebsbach, P.H.; Kuznetsov, S.A.; Satomura, K.; Emmons, R.V.; Rowe, D.W.; Robey, P.G. Bone formation in vivo: Comparison of osteogenesis by transplanted mouse and human marrow stromal fibroblasts. Transplantation 1997, 63, 1059–1069. [Google Scholar] [CrossRef]
- Persichini, T.; Funari, A.; Colasanti, M.; Sacchetti, B. Clonogenic, myogenic progenitors expressing MCAM/CD146 are incorporated as adventitial reticular cells in the microvascular compartment of human post-natal skeletal muscle. PLoS ONE 2017, 12, e0188844. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cell Line | Route | Transplants | Transplants with Metastases | Metastases in Mouse Bones |
|---|---|---|---|---|
| PC-3 | i.c. | 12 | 7 | - |
| PC-3 | i.v. | 8 | 4 | - |
| RWPE-1 | i.c. | 8 | 0 | - |
| RWPE-1 | i.v. | 12 | 0 | - |
| Antigen | Type | Cat# | Distributor |
|---|---|---|---|
| human Mitochondria | MM | MAB-1273 | Chemicon |
| human CD146/MCAM | MM | NCL-CD146 | Novocastra |
| human CD34 | MM | NCL-END | Novocastra |
| murine CD34 | RaM | ab8158 | Abcam |
| CD3 | RP | A0452 | Dako |
| GFP | RP | ab6556 | Abcam |
| GFP | RP | A6455 | Invitrogen |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Funari, A.; Alimandi, M.; Pierelli, L.; Pino, V.; Gentileschi, S.; Sacchetti, B. Human Sinusoidal Subendothelial Cells Regulate Homing and Invasion of Circulating Metastatic Prostate Cancer Cells to Bone Marrow. Cancers 2019, 11, 763. https://doi.org/10.3390/cancers11060763
Funari A, Alimandi M, Pierelli L, Pino V, Gentileschi S, Sacchetti B. Human Sinusoidal Subendothelial Cells Regulate Homing and Invasion of Circulating Metastatic Prostate Cancer Cells to Bone Marrow. Cancers. 2019; 11(6):763. https://doi.org/10.3390/cancers11060763
Chicago/Turabian StyleFunari, Alessia, Maurizio Alimandi, Luca Pierelli, Valentina Pino, Stefano Gentileschi, and Benedetto Sacchetti. 2019. "Human Sinusoidal Subendothelial Cells Regulate Homing and Invasion of Circulating Metastatic Prostate Cancer Cells to Bone Marrow" Cancers 11, no. 6: 763. https://doi.org/10.3390/cancers11060763