c-Myc Acts as a Competing Endogenous RNA to Sponge miR-34a, in the Upregulation of CD44, in Urothelial Carcinoma



Abstract
:1. Introduction
2. Results
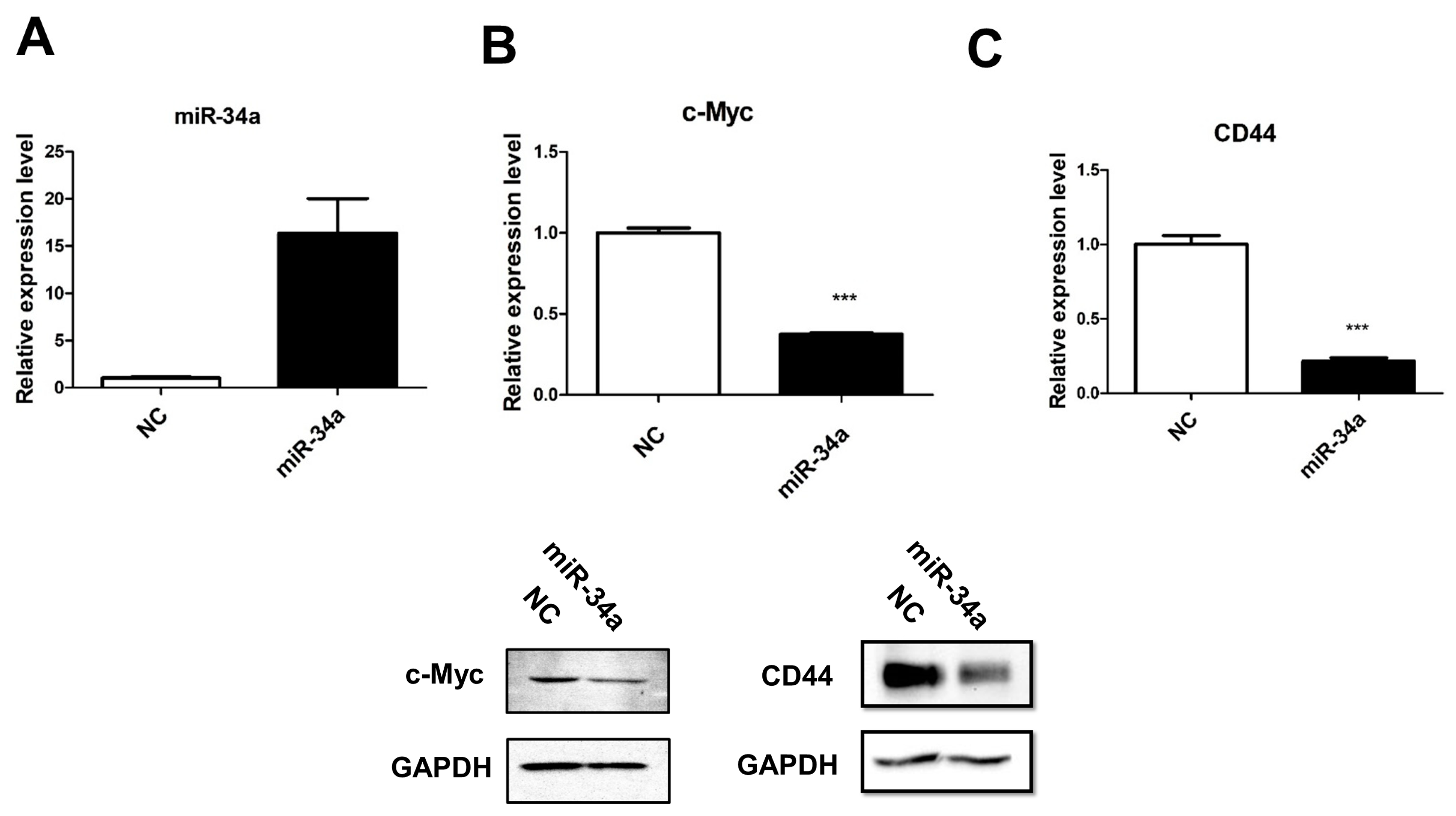
2.1. miR-34a Is Epigenetically Silenced in the UMUC3 Cell Line
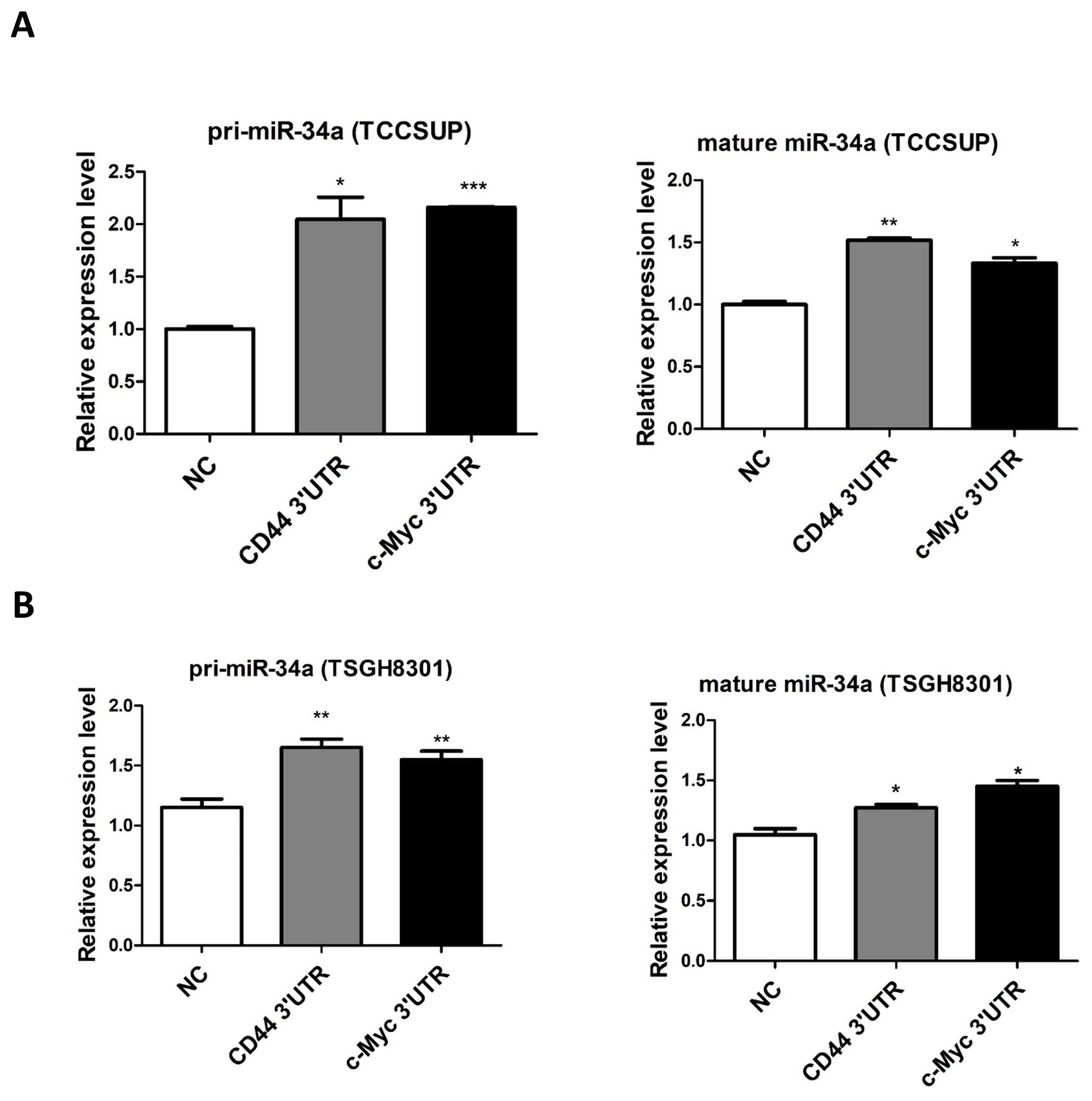
2.2. c-Myc Acts as a ceRNA of CD44 in Urothelial Carcinoma
2.3. Dicer Knockdown Abolishes ceRNA Effect in UC
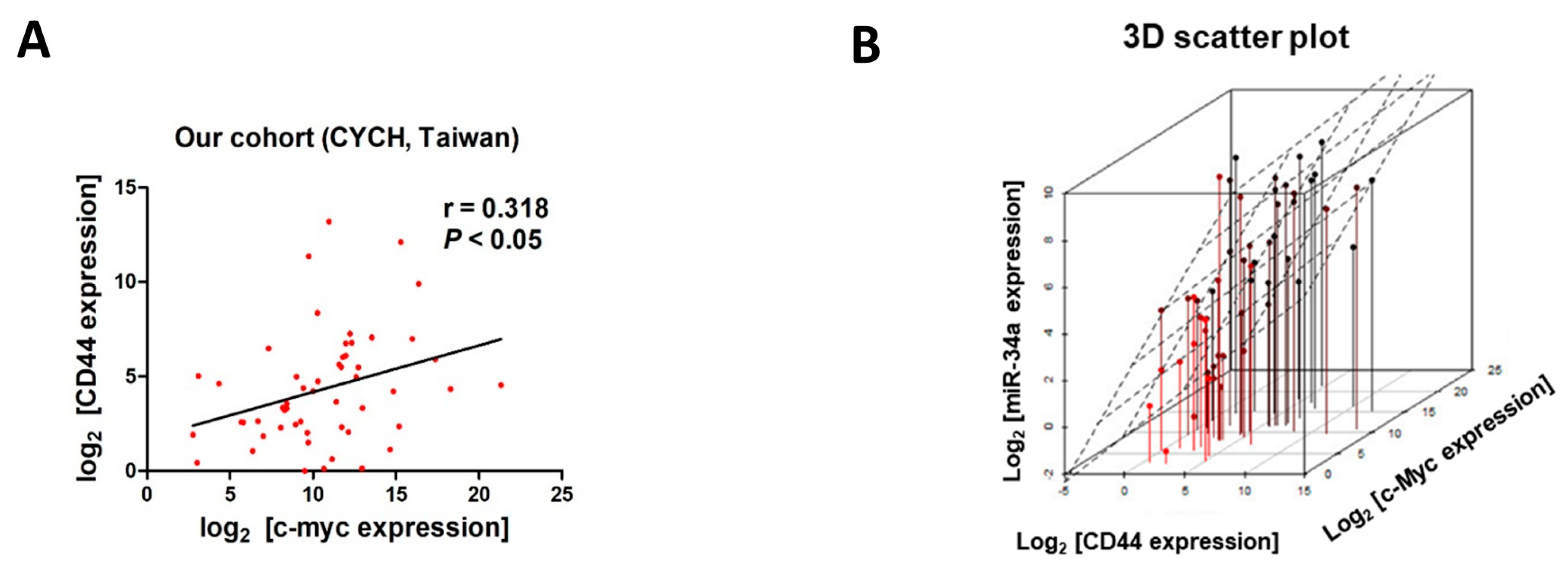
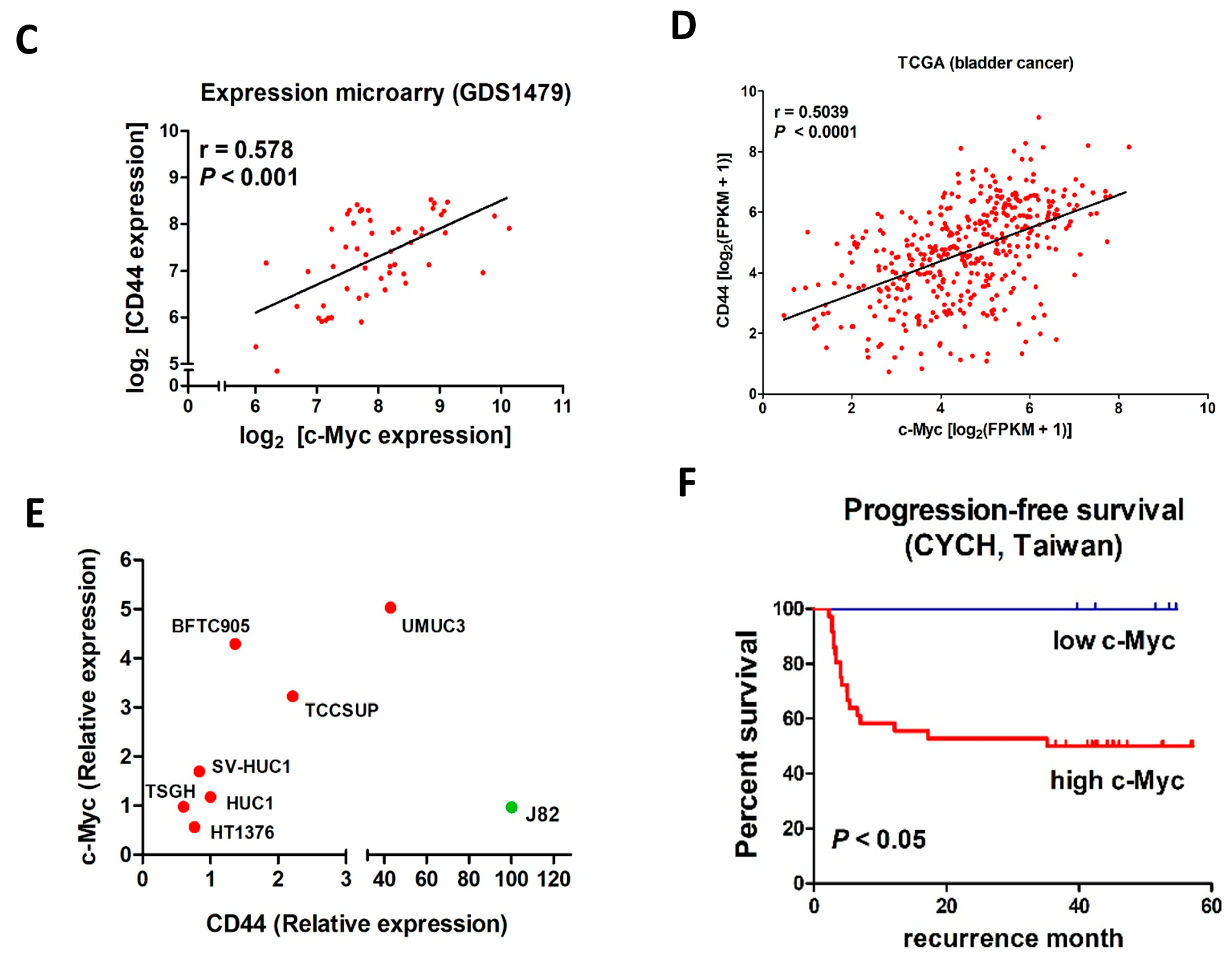
2.4. Expression of c-Myc Correlates with Expression of CD44, in Samples from UC patients and Cell Lines
2.5. Dominant-Negative p53 Mutation Downregulates miR-34a in UC
2.6. miR-34a Suppresses Tumorigenicity and Drug Resistance in UC
3. Discussion
4. Materials and Methods
4.1. Cell Cultures and Patient Samples
4.2. Plasmid Constructs and shRNA Knockdown
4.3. RNA Expression
4.4. DNA Methylation Analysis
4.5. Cell Invasion Assay
4.6. Cell Viability Assay
4.7. Soft Agar Assay
4.8. Western Blotting
4.9. Bioinformatics Analysis
4.10. In Vivo Tumorigenicity Assay
4.11. Statistical Calculation
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Green, D.A.; Rink, M.; Xylinas, E.; Matin, S.F.; Stenzl, A.; Roupret, M.; Karakiewicz, P.I.; Scherr, D.S.; Shariat, S.F. Urothelial carcinoma of the bladder and the upper tract: Disparate twins. J. Urol. 2013, 189, 1214–1221. [Google Scholar] [CrossRef]
- Tan, L.B.; Chang, L.L.; Cheng, K.I.; Huang, C.H.; Kwan, A.L. Transitional cell carcinomas of the renal pelvis and the ureter: Comparative demographic characteristics, pathological grade and stage and 5-year survival in a Taiwanese population. BJU Int. 2009, 103, 312–316. [Google Scholar] [CrossRef]
- Nguyen, D.P.; Thalmann, G.N. Contemporary update on neoadjuvant therapy for bladder cancer. Nat. Rev. Urol. 2017, 14, 348–358. [Google Scholar] [CrossRef]
- Chan, K.S.; Espinosa, I.; Chao, M.; Wong, D.; Ailles, L.; Diehn, M.; Gill, H.; Presti, J., Jr.; Chang, H.Y.; van de Rijn, M.; et al. Identification, molecular characterization, clinical prognosis, and therapeutic targeting of human bladder tumor-initiating cells. Proc. Natl. Acad. Sci. USA 2009, 106, 14016–14021. [Google Scholar] [CrossRef] [Green Version]
- Wu, C.T.; Lin, W.Y.; Chang, Y.H.; Chen, W.C.; Chen, M.F. Impact of CD44 expression on radiation response for bladder cancer. J. Cancer 2017, 8, 1137–1144. [Google Scholar] [CrossRef] [Green Version]
- Xu, H.; Tian, Y.; Yuan, X.; Wu, H.; Liu, Q.; Pestell, R.G.; Wu, K. The role of CD44 in epithelial-mesenchymal transition and cancer development. Onco Targets Ther. 2015, 8, 3783–3792. [Google Scholar] [CrossRef]
- van der Horst, G.; Bos, L.; van der Pluijm, G. Epithelial plasticity, cancer stem cells, and the tumor-supportive stroma in bladder carcinoma. Mol. Cancer Res. 2012, 10, 995–1009. [Google Scholar] [CrossRef]
- Ponta, H.; Sherman, L.; Herrlich, P.A. CD44: From adhesion molecules to signalling regulators. Nat. Rev. Mol. Cell Biol. 2003, 4, 33–45. [Google Scholar] [CrossRef]
- Jones, P.A.; Issa, J.P.; Baylin, S. Targeting the cancer epigenome for therapy. Nat. Rev. Genet. 2016, 17, 630–641. [Google Scholar] [CrossRef]
- Bartel, D.P. MicroRNAs: Genomics, biogenesis, mechanism, and function. Cell 2004, 116, 281–297. [Google Scholar] [CrossRef]
- Poliseno, L.; Salmena, L.; Zhang, J.; Carver, B.; Haveman, W.J.; Pandolfi, P.P. A coding-independent function of gene and pseudogene mRNAs regulates tumour biology. Nature 2010, 465, 1033–1038. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baranwal, S.; Alahari, S.K. miRNA control of tumor cell invasion and metastasis. Int. J. Cancer 2010, 126, 1283–1290. [Google Scholar] [CrossRef] [PubMed]
- Song, C.; Lu, P.; Sun, G.; Yang, L.; Wang, Z.; Wang, Z. miR-34a sensitizes lung cancer cells to cisplatin via p53/miR-34a/MYCN axis. Biochem. Biophys. Res. Commun. 2017, 482, 22–27. [Google Scholar] [CrossRef] [PubMed]
- Christoffersen, N.R.; Shalgi, R.; Frankel, L.B.; Leucci, E.; Lees, M.; Klausen, M.; Pilpel, Y.; Nielsen, F.C.; Oren, M.; Lund, A.H. p53-independent upregulation of miR-34a during oncogene-induced senescence represses MYC. Cell Death Differ. 2010, 17, 236–245. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Kelnar, K.; Liu, B.; Chen, X.; Calhoun-Davis, T.; Li, H.; Patrawala, L.; Yan, H.; Jeter, C.; Honorio, S.; et al. The microRNA miR-34a inhibits prostate cancer stem cells and metastasis by directly repressing CD44. Nat. Med. 2011, 17, 211–215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, H.; Yu, G.; Shi, R.; Lang, B.; Chen, X.; Xia, D.; Xiao, H.; Guo, X.; Guan, W.; Ye, Z.; et al. Cisplatin-induced epigenetic activation of miR-34a sensitizes bladder cancer cells to chemotherapy. Mol. Cancer 2014, 13, 8. [Google Scholar] [CrossRef] [PubMed]
- Yu, G.; Yao, W.; Xiao, W.; Li, H.; Xu, H.; Lang, B. MicroRNA-34a functions as an anti-metastatic microRNA and suppresses angiogenesis in bladder cancer by directly targeting CD44. J. Exp. Clin. Cancer Res. 2014, 33, 779. [Google Scholar] [CrossRef] [PubMed]
- Yu, G.; Li, H.; Wang, J.; Gumireddy, K.; Li, A.; Yao, W.; Tang, K.; Xiao, W.; Hu, J.; Xiao, H.; et al. miRNA-34a suppresses cell proliferation and metastasis by targeting CD44 in human renal carcinoma cells. J. Urol. 2014, 192, 1229–1237. [Google Scholar] [CrossRef]
- Lodygin, D.; Tarasov, V.; Epanchintsev, A.; Berking, C.; Knyazeva, T.; Korner, H.; Knyazev, P.; Diebold, J.; Hermeking, H. Inactivation of miR-34a by aberrant CpG methylation in multiple types of cancer. Cell Cycle 2008, 7, 2591–2600. [Google Scholar] [CrossRef] [Green Version]
- Suzuki, H.; Maruyama, R.; Yamamoto, E.; Kai, M. DNA methylation and microRNA dysregulation in cancer. Mol. Oncol. 2012, 6, 567–578. [Google Scholar] [CrossRef] [Green Version]
- Zarringhalam, K.; Tay, Y.; Kulkarni, P.; Bester, A.C.; Pandolfi, P.P.; Kulkarni, R.V. Identification of competing endogenous RNAs of the tumor suppressor gene PTEN: A probabilistic approach. Sci. Rep. 2017, 7, 7755. [Google Scholar] [CrossRef] [PubMed]
- Yamamura, S.; Saini, S.; Majid, S.; Hirata, H.; Ueno, K.; Deng, G.; Dahiya, R. MicroRNA-34a modulates c-Myc transcriptional complexes to suppress malignancy in human prostate cancer cells. PLoS ONE 2012, 7, e29722. [Google Scholar] [CrossRef] [PubMed]
- Mahdy, E.; Pan, Y.; Wang, N.; Malmstrom, P.U.; Ekman, P.; Bergerheim, U. Chromosome 8 numerical aberration and C-MYC copy number gain in bladder cancer are linked to stage and grade. Anticancer Res. 2001, 21, 3167–3173. [Google Scholar] [PubMed]
- Zaharieva, B.; Simon, R.; Ruiz, C.; Oeggerli, M.; Mihatsch, M.J.; Gasser, T.; Sauter, G.; Toncheva, D. High-throughput tissue microarray analysis of CMYC amplificationin urinary bladder cancer. Int. J. Cancer 2005, 117, 952–956. [Google Scholar] [CrossRef] [PubMed]
- Carthew, R.W.; Sontheimer, E.J. Origins and Mechanisms of miRNAs and siRNAs. Cell 2009, 136, 642–655. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, C.Y.; Loven, J.; Rahl, P.B.; Paranal, R.M.; Burge, C.B.; Bradner, J.E.; Lee, T.I.; Young, R.A. Transcriptional amplification in tumor cells with elevated c-Myc. Cell 2012, 151, 56–67. [Google Scholar] [CrossRef] [PubMed]
- Raver-Shapira, N.; Marciano, E.; Meiri, E.; Spector, Y.; Rosenfeld, N.; Moskovits, N.; Bentwich, Z.; Oren, M. Transcriptional activation of miR-34a contributes to p53-mediated apoptosis. Mol. Cell 2007, 26, 731–743. [Google Scholar] [CrossRef]
- Navarro, F.; Lieberman, J. miR-34 and p53: New Insights into a Complex Functional Relationship. PLoS ONE 2015, 10, e0132767. [Google Scholar] [CrossRef]
- Chang, F.L.; Ling, Y.F.; Lai, M.D. Exogenous mutant p53 DNA enhanced cisplatin-induced apoptosis in TSGH-8301 human bladder cancer cells. Anticancer Res. 2000, 20, 329–336. [Google Scholar]
- Chen, A.H.; Qin, Y.E.; Tang, W.F.; Tao, J.; Song, H.M.; Zuo, M. MiR-34a and miR-206 act as novel prognostic and therapy biomarkers in cervical cancer. Cancer Cell Int. 2017, 17, 63. [Google Scholar] [CrossRef] [Green Version]
- Slabakova, E.; Culig, Z.; Remsik, J.; Soucek, K. Alternative mechanisms of miR-34a regulation in cancer. Cell Death Dis. 2017, 8, e3100. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Li, T.; Han, G.; Li, Y.; Shi, L.H.; Li, H. Expression and role of miR-34a in bladder cancer. Indian J. Biochem. Biophys. 2013, 50, 87–92. [Google Scholar] [PubMed]
- Kwon, H.; Song, K.; Han, C.; Zhang, J.; Lu, L.; Chen, W.; Wu, T. Epigenetic Silencing of miRNA-34a in Human Cholangiocarcinoma via EZH2 and DNA Methylation: Impact on Regulation of Notch Pathway. Am. J. Pathol. 2017, 187, 2288–2299. [Google Scholar] [CrossRef] [PubMed]
- Siemens, H.; Neumann, J.; Jackstadt, R.; Mansmann, U.; Horst, D.; Kirchner, T.; Hermeking, H. Detection of miR-34a promoter methylation in combination with elevated expression of c-Met and beta-catenin predicts distant metastasis of colon cancer. Clin. Cancer Res. 2013, 19, 710–720. [Google Scholar] [CrossRef] [PubMed]
- Salmena, L.; Poliseno, L.; Tay, Y.; Kats, L.; Pandolfi, P.P. A ceRNA hypothesis: The Rosetta Stone of a hidden RNA language? Cell 2011, 146, 353–358. [Google Scholar] [CrossRef] [PubMed]
- Cheng, F.H.C.; Lin, H.Y.; Hwang, T.W.; Chen, Y.C.; Huang, R.L.; Chang, C.B.; Yang, W.; Lin, R.I.; Lin, C.W.; Chen, G.C.W.; et al. E2F6 functions as a competing endogenous RNA, and transcriptional repressor, to promote ovarian cancer stemness. Cancer Sci. 2018. [Google Scholar] [CrossRef]
- Katsushima, K.; Natsume, A.; Ohka, F.; Shinjo, K.; Hatanaka, A.; Ichimura, N.; Sato, S.; Takahashi, S.; Kimura, H.; Totoki, Y.; et al. Targeting the Notch-regulated non-coding RNA TUG1 for glioma treatment. Nat. Commun. 2016, 7, 13616. [Google Scholar] [CrossRef]
- Yang, R.; Xing, L.; Zheng, X.; Sun, Y.; Wang, X.; Chen, J. The circRNA circAGFG1 acts as a sponge of miR-195-5p to promote triple-negative breast cancer progression through regulating CCNE1 expression. Mol. Cancer 2019, 18, 4. [Google Scholar] [CrossRef]
- Yang, F.; Shen, Y.; Zhang, W.; Jin, J.; Huang, D.; Fang, H.; Ji, W.; Shi, Y.; Tang, L.; Chen, W.; et al. An androgen receptor negatively induced long non-coding RNA ARNILA binding to miR-204 promotes the invasion and metastasis of triple-negative breast cancer. Cell Death Differ. 2018, 25, 2209–2220. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Zeng, C.; Hu, J.; Pan, Y.; Shan, Y.; Liu, B.; Jia, L. Long non-coding RNA-SNHG7 acts as a target of miR-34a to increase GALNT7 level and regulate PI3K/Akt/mTOR pathway in colorectal cancer progression. J. Hematol. Oncol. 2018, 11, 89. [Google Scholar] [CrossRef]
- Huang, X.; Xie, X.; Liu, P.; Yang, L.; Chen, B.; Song, C.; Tang, H.; Xie, X. Adam12 and lnc015192 act as ceRNAs in breast cancer by regulating miR-34a. Oncogene 2018, 37, 6316–6326. [Google Scholar] [CrossRef] [PubMed]
- Zviran, A.; Mor, N.; Rais, Y.; Gingold, H.; Peles, S.; Chomsky, E.; Viukov, S.; Buenrostro, J.D.; Scognamiglio, R.; Weinberger, L.; et al. Deterministic Somatic Cell Reprogramming Involves Continuous Transcriptional Changes Governed by Myc and Epigenetic-Driven Modules. Cell Stem Cell 2019, 24, 328–341. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, G.J. Emerging roles of Myc in stem cell biology and novel tumor therapies. J. Exp. Clin. Cancer Res. 2018, 37, 173. [Google Scholar] [CrossRef] [PubMed]
- van Schaijik, B.; Davis, P.F.; Wickremesekera, A.C.; Tan, S.T.; Itinteang, T. Subcellular localisation of the stem cell markers OCT4, SOX2, NANOG, KLF4 and c-MYC in cancer: A review. J. Clin. Pathol. 2018, 71, 88–91. [Google Scholar] [CrossRef]
- Muller, P.A.; Vousden, K.H. Mutant p53 in cancer: New functions and therapeutic opportunities. Cancer Cell 2014, 25, 304–317. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Vida, A.; Lerner, S.P.; Bellmunt, J. The Cancer Genome Atlas Project in Bladder Cancer. Cancer Treat. Res. 2018, 175, 259–271. [Google Scholar] [CrossRef]
- Rokavec, M.; Li, H.; Jiang, L.; Hermeking, H. The p53/miR-34 axis in development and disease. J. Mol. Cell Biol. 2014, 6, 214–230. [Google Scholar] [CrossRef] [Green Version]
- Menges, C.W.; Kadariya, Y.; Altomare, D.; Talarchek, J.; Neumann-Domer, E.; Wu, Y.; Xiao, G.H.; Shapiro, I.M.; Kolev, V.N.; Pachter, J.A.; et al. Tumor suppressor alterations cooperate to drive aggressive mesotheliomas with enriched cancer stem cells via a p53-miR-34a-c-Met axis. Cancer Res. 2014, 74, 1261–1271. [Google Scholar] [CrossRef]
- Siemens, H.; Jackstadt, R.; Kaller, M.; Hermeking, H. Repression of c-Kit by p53 is mediated by miR-34 and is associated with reduced chemoresistance, migration and stemness. Oncotarget 2013, 4, 1399–1415. [Google Scholar] [CrossRef]
- Lai, X.; Wolkenhauer, O.; Vera, J. Understanding microRNA-mediated gene regulatory networks through mathematical modelling. Nucleic Acids Res. 2016, 44, 6019–6035. [Google Scholar] [CrossRef] [Green Version]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| miR-34a | c-Myc | CD44 | |
|---|---|---|---|
| Gender | |||
| Male | 130.2 ± 173.6 1 | 8.4 ± 4.1 | 457.3 ± 1.6 |
| Female | 31.1 ± 67.5 | 6.1 ± 1.8 | 23.8 ± 42.6 |
| Location | |||
| Bladder | 106.9 ± 157.8 | 1.7 ± 5.5 | 512.5 ± 1.7 |
| Upper Tract | 105.1 ± 169.1 | 1.6 ± 6.2 | 33.8 ± 44.2 |
| Histological Grade | |||
| Low Grade | 143.7 ± 198.1 | 2.3 ± 7.6 | 108.1 ± 270.1 |
| High Grade | 100.2 ± 149.8 | 1.7 ± 5.5 | 413.5 ± 1.6 |
| Pathological Stage | |||
| Stage T1 | 90.9 ± 149.2 | 1.7 ± 5.9 | 68.9 ± 179.1 |
| ≥Stage T2 | 132.2 ± 173.5 | 1.2 ± 5.3 | 680.5 ± 2.1 |
| Relapse | |||
| Yes | 108.6 ± 153.1 | 1.4 ± 3.4 | 566.1 ± 1.8 |
| No | 102.7 ± 174.4 | 1.5 ± 5.8 | 17.9 ± 22.9 |
| Clinical Parameter | Patient Samples (n = 55) |
|---|---|
| Age * | 70.94 ± 11.8 4 |
| Median | 74 |
| Range | 40–90 |
| Gender * | |
| Male | 41 |
| Female | 13 |
| Location 1,* | |
| Bladder | 36 |
| Upper Tract | 18 |
| Histological Grade 2 | |
| Low Grade | 12 |
| High Grade | 43 |
| Pathological Stage | |
| Stage T1 | 30 |
| ≥ Stage T2 | 25 |
| Relapse * | |
| Yes | 33 |
| No | 21 |
| Treatment | |
| TURBT | 34 |
| Non-TURBT 3 | 21 |
| Primer Name | Primer Sequence (5′ to 3′) | Annealing TEMP (°C) | Product Size (bp) |
|---|---|---|---|
| COBRA | |||
| miR-34a-BS-F | TTTTTTTTTTTAGGTGGAGGAG | 60 | 270 |
| miR-34a-BS-R | ATACAAACTTCCAAACCTCTCC | ||
| Bisulphite Pyrosequeincing | |||
| miR-34a-Pyro-F | TAGGTGGGGGTTAGGTAG | 64 | 87 |
| miR-34a-Pyro-R 1 | agctggacatcacctcccacaacgCAAACTCCCACCCCTCCC | ||
| miR-34a-SEQ | GGTAGGGAGTATGAAG | ||
| RT-PCR | |||
| pri-miR-34a RT-F | TGTGATTAACCCCGTCTTGCA | 60 | 101 |
| pri-miR-34a RT-R | GCAGATTCTTGAGCCAGATTGC | ||
| CD44-RT-F | TCCCAGTATGACACATATTGC | 60 | 129 |
| CD44-RT-R | CACCTTCTTCGACTGTTGAC | ||
| c-Myc-RT-F | CTCGGATTCTCTGCTCTCTCCTCG | 60 | 177 |
| c-Myc-RT-R | TCTGACCTTTTGCCAGGAGCCT | ||
| Dicer-RT-F | TTAACCTTTTGGTGTTTGATGAGTGT | 60 | 98 |
| Dicer-RT-R | GCGAGGACATGATGGACAATT | ||
| GAPDH RT-F | CCCCTTCATTGACCTCAACTACAT | 60 | 135 |
| GAPDH RT-R | CGCTCCTGGAAGATGGTGA | ||
| Plasmid construction | |||
| miR-34a construct | |||
| miR-34a-BamHI F | CGCGGATCCGTAGAGATGGAGTCTTGCTAGTTGC | 64 | 514 |
| miR-34a-HindIII R | CCCAAGCTTTTCTCCCTACGTGCAAACTTCT | ||
| 3′UTR construct | |||
| CD44-XhoI F | CCTCGAGTGATCGTTCCAGTTCCCACTTG | 66 | 263 |
| CD44-PstI R | CCCCTGCAGGGGGGTCTGTTGAAGAT | ||
| c-Myc-XhoI F | CCTCGAGCTTGAGACTGAAAGATTTAGCCAT | 66 | 369 |
| c-Myc-PstI R | CCCCTGCAGTAAGATTTGGCTCAATGATATATTTG | ||
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, P.-C.; Yu, C.-C.; Huang, W.-Y.; Huang, W.-H.; Chuang, Y.-M.; Lin, R.-I.; Lin, J.M.J.; Lin, H.-Y.; Jou, Y.-C.; Shen, C.-H.; et al. c-Myc Acts as a Competing Endogenous RNA to Sponge miR-34a, in the Upregulation of CD44, in Urothelial Carcinoma. Cancers 2019, 11, 1457. https://doi.org/10.3390/cancers11101457
Chen P-C, Yu C-C, Huang W-Y, Huang W-H, Chuang Y-M, Lin R-I, Lin JMJ, Lin H-Y, Jou Y-C, Shen C-H, et al. c-Myc Acts as a Competing Endogenous RNA to Sponge miR-34a, in the Upregulation of CD44, in Urothelial Carcinoma. Cancers. 2019; 11(10):1457. https://doi.org/10.3390/cancers11101457
Chicago/Turabian StyleChen, Pie-Che, Chih-Chia Yu, Wen-Yu Huang, Wan-Hong Huang, Yu-Ming Chuang, Ru-Inn Lin, Jora M. J. Lin, Hon-Yi Lin, Yeong-Chin Jou, Cheng-Huang Shen, and et al. 2019. "c-Myc Acts as a Competing Endogenous RNA to Sponge miR-34a, in the Upregulation of CD44, in Urothelial Carcinoma" Cancers 11, no. 10: 1457. https://doi.org/10.3390/cancers11101457
APA StyleChen, P.-C., Yu, C.-C., Huang, W.-Y., Huang, W.-H., Chuang, Y.-M., Lin, R.-I., Lin, J. M. J., Lin, H.-Y., Jou, Y.-C., Shen, C.-H., & Chan, M. W. Y. (2019). c-Myc Acts as a Competing Endogenous RNA to Sponge miR-34a, in the Upregulation of CD44, in Urothelial Carcinoma. Cancers, 11(10), 1457. https://doi.org/10.3390/cancers11101457