Functional Linkage of RKIP to the Epithelial to Mesenchymal Transition and Autophagy during the Development of Prostate Cancer

 ,
,  ,
, 
Abstract
:1. Introduction
2. Results
2.1. Expression Datasets and Gene Annotations
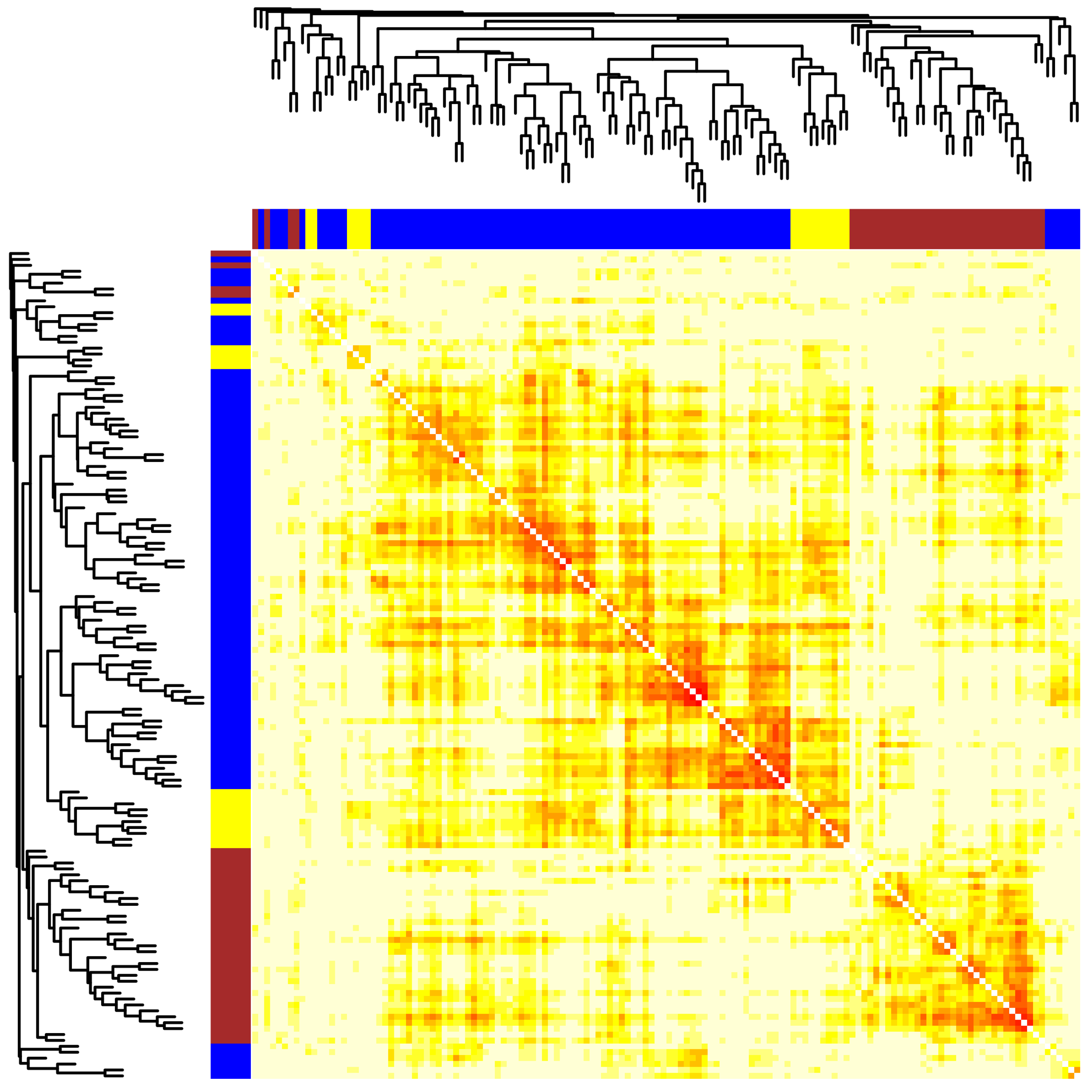
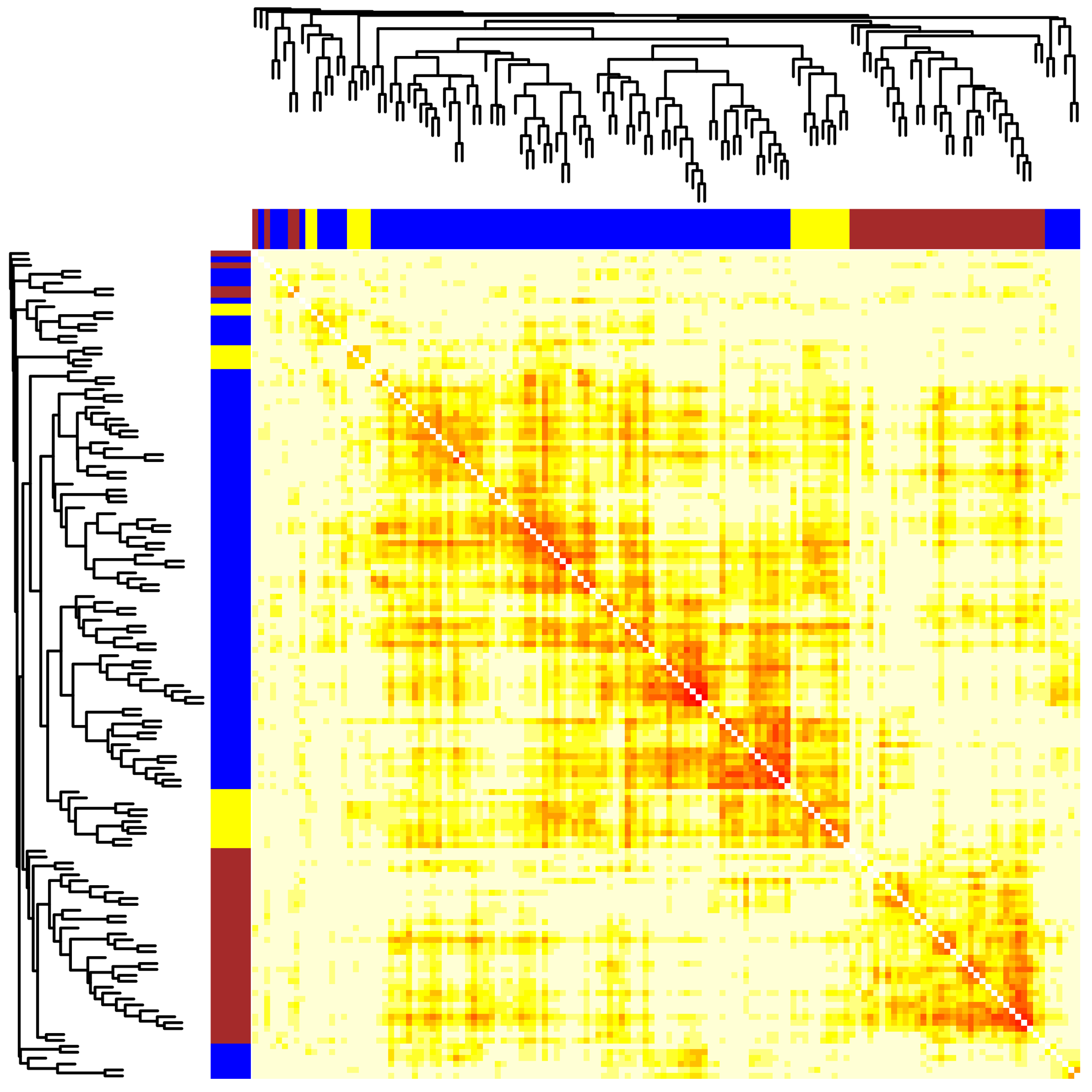
2.2. Module Detection of Interconnected Genes
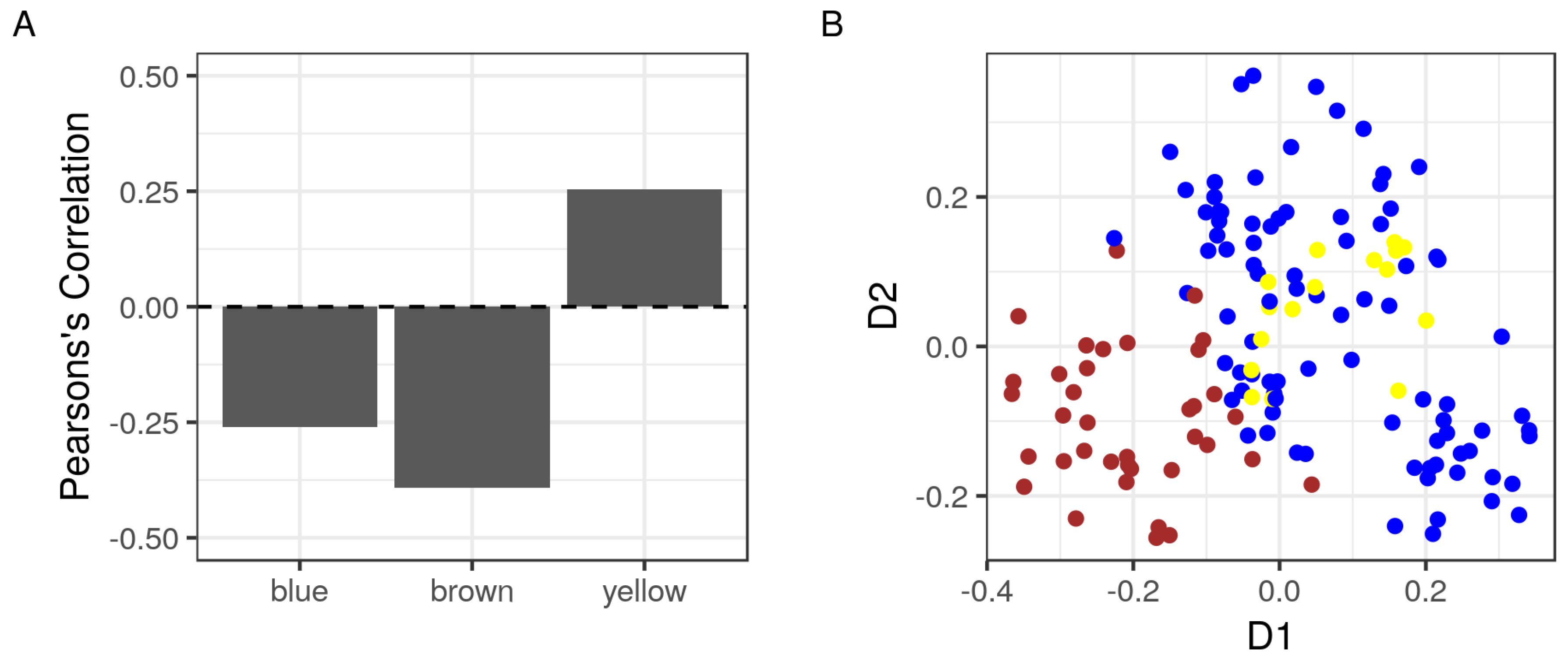
2.3. The Correlation of Modules with the Sample Phenotype
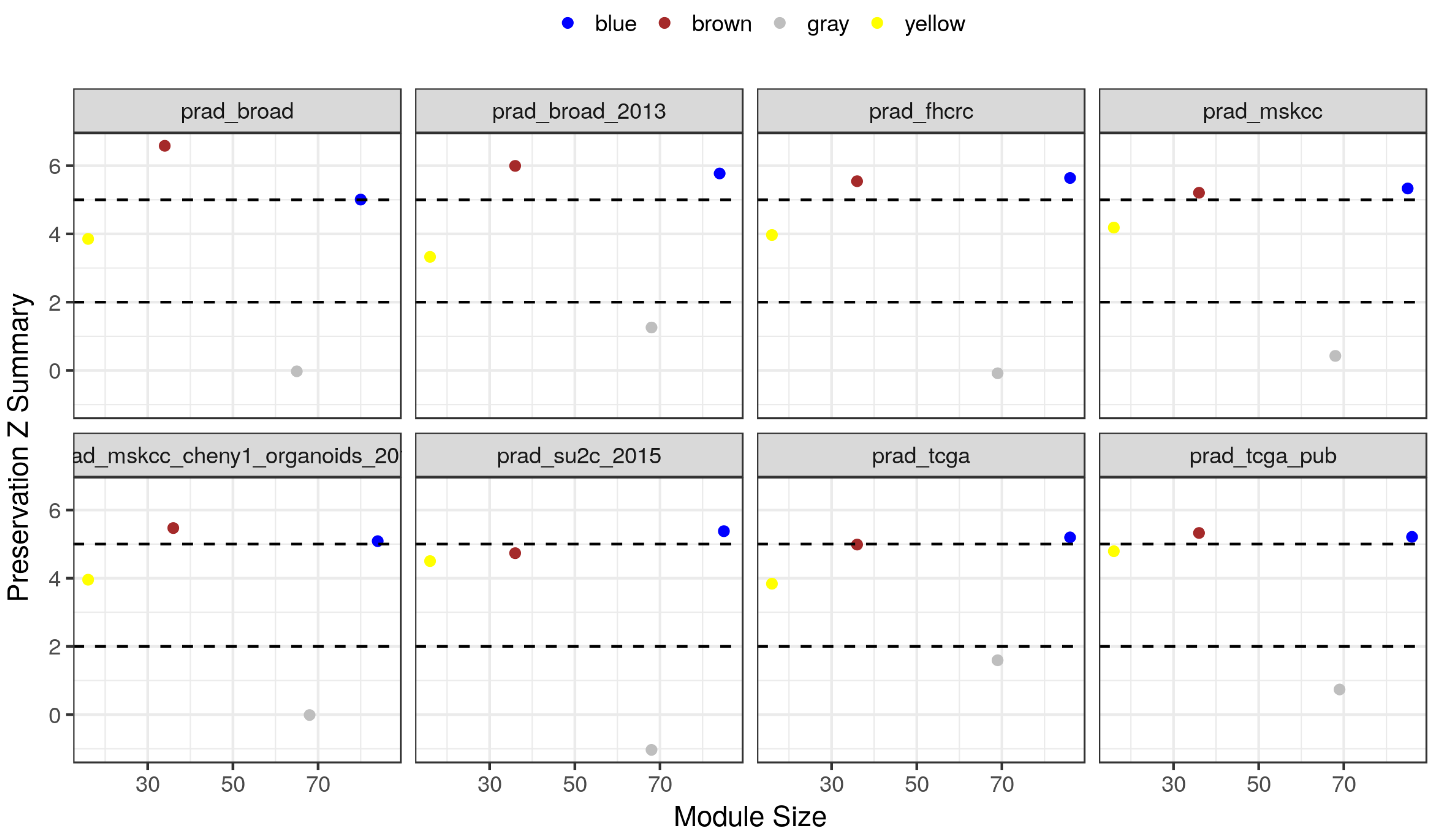
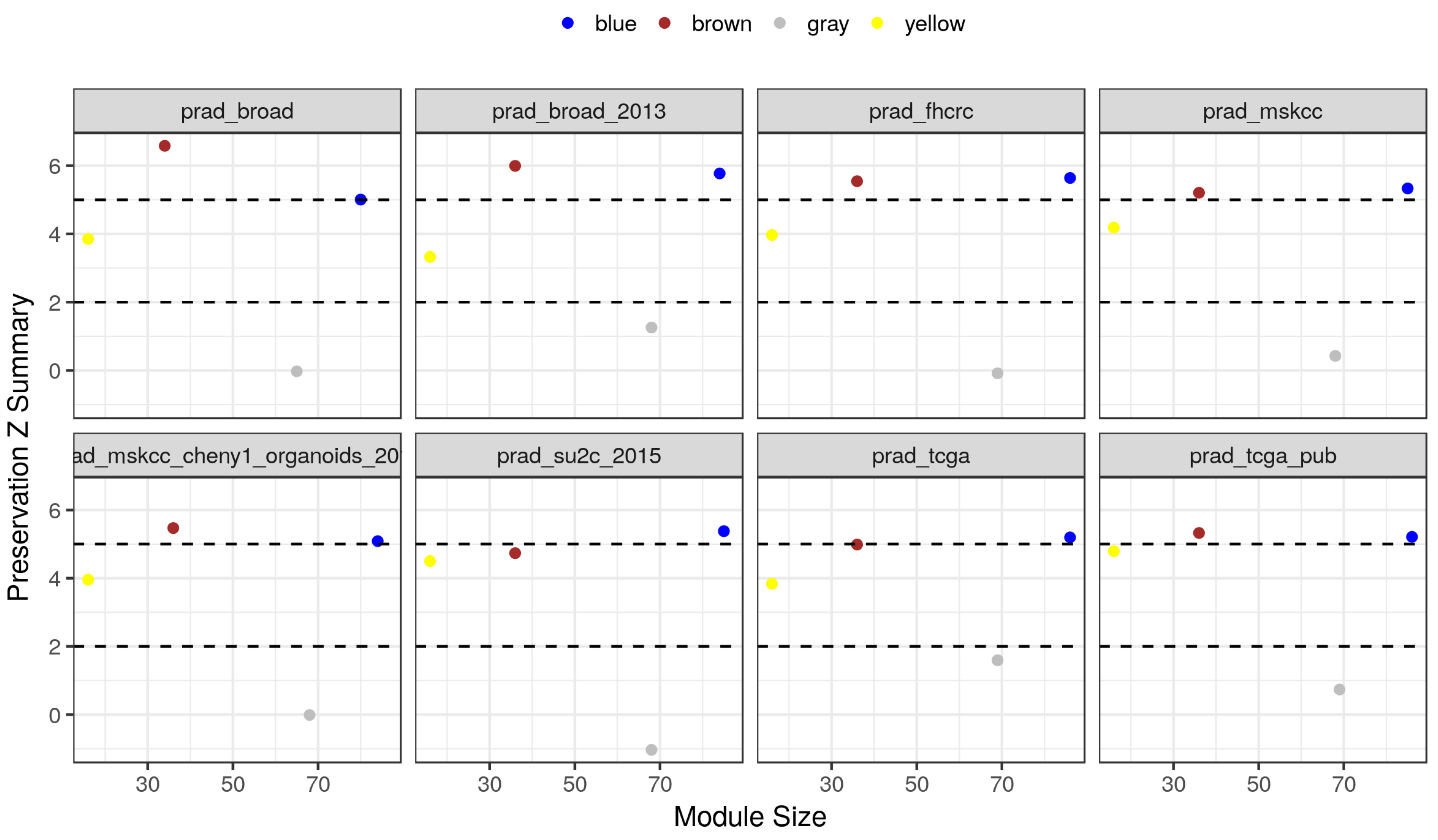
2.4. Module Preservation in Independent Datasets
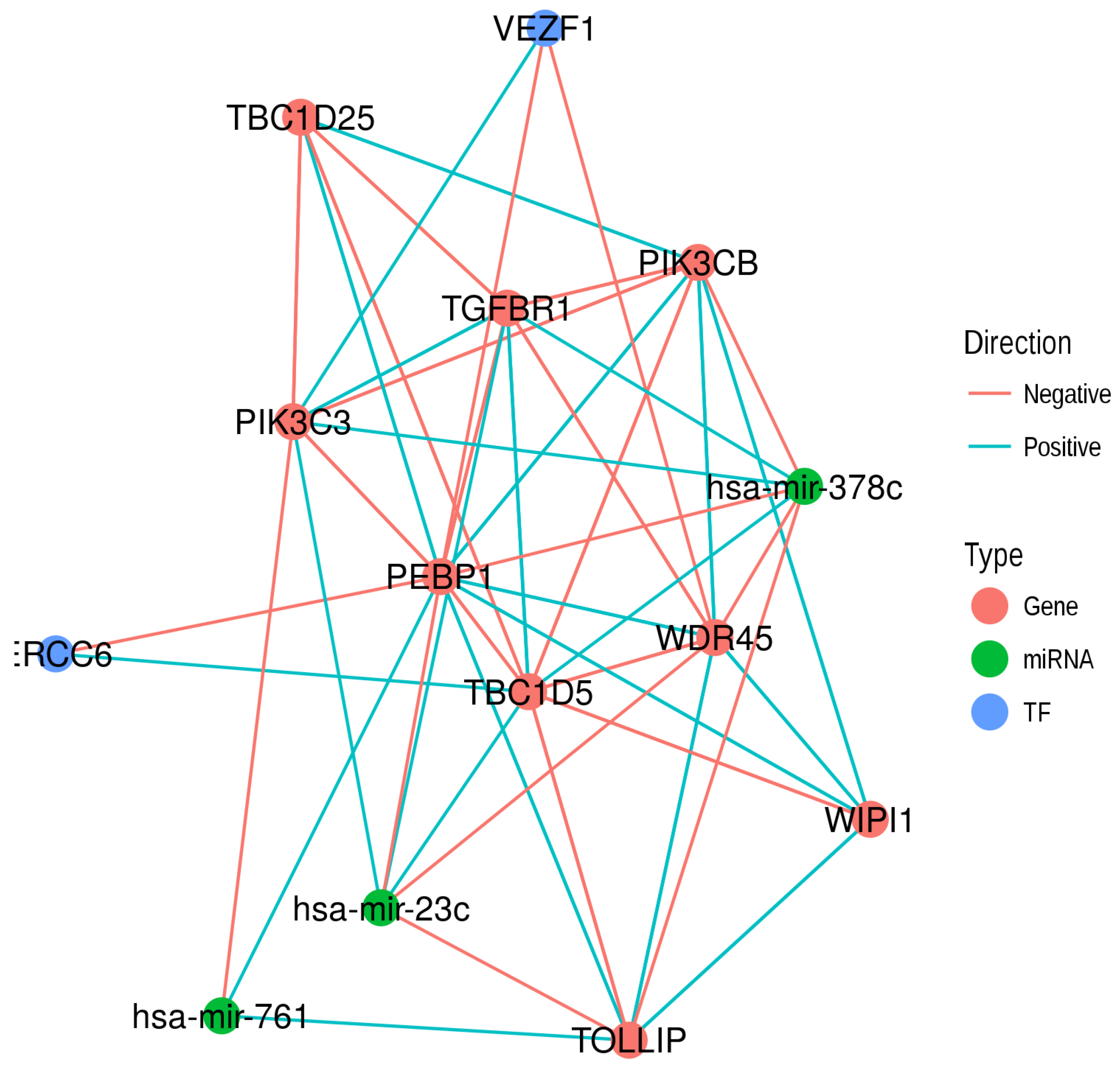
2.5. The Potential Interactions of RKIP/PEBP1 with Autophagy and EMT Gene Products
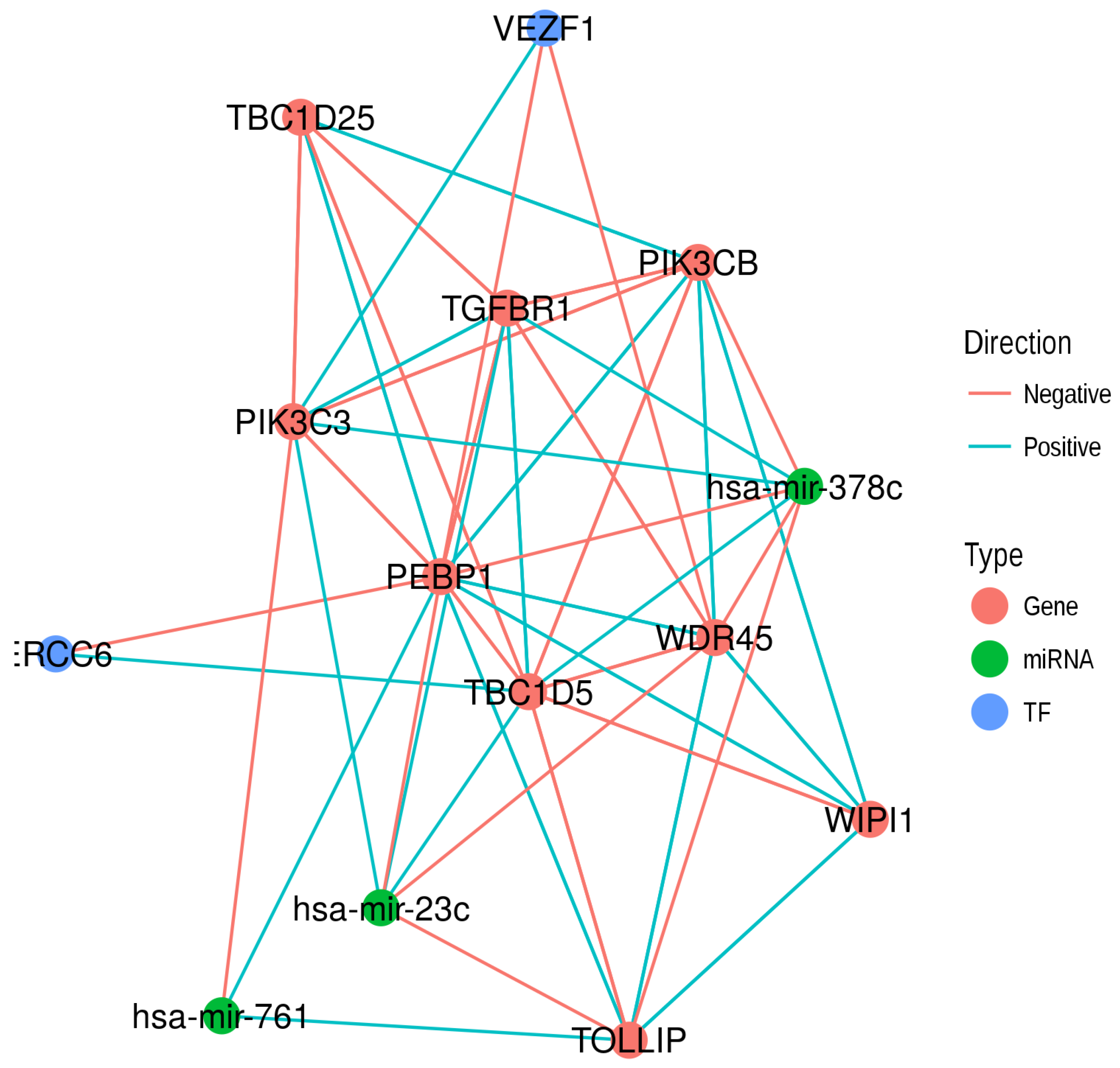
2.6. Common Regulators of RKIP/PEBP1 and Its Interacting Partners
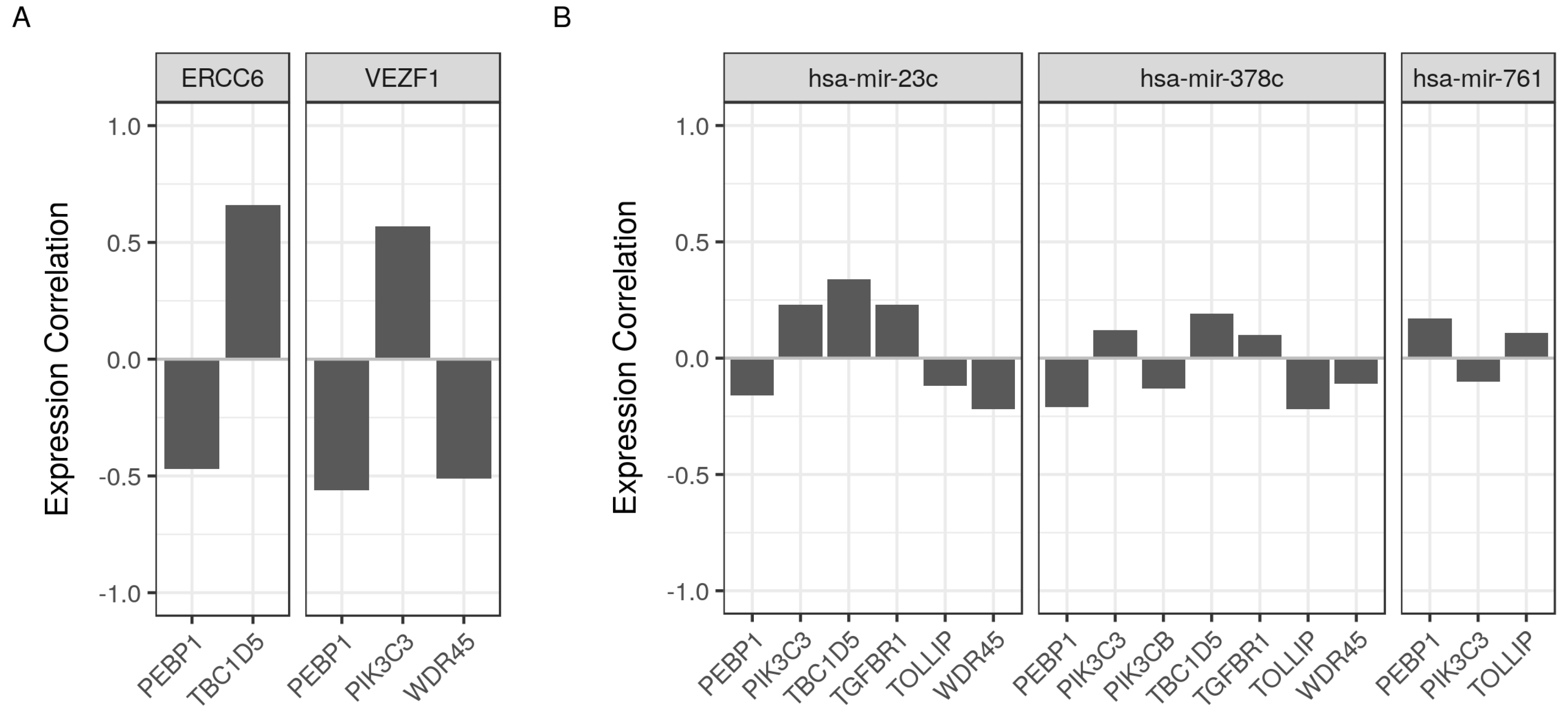
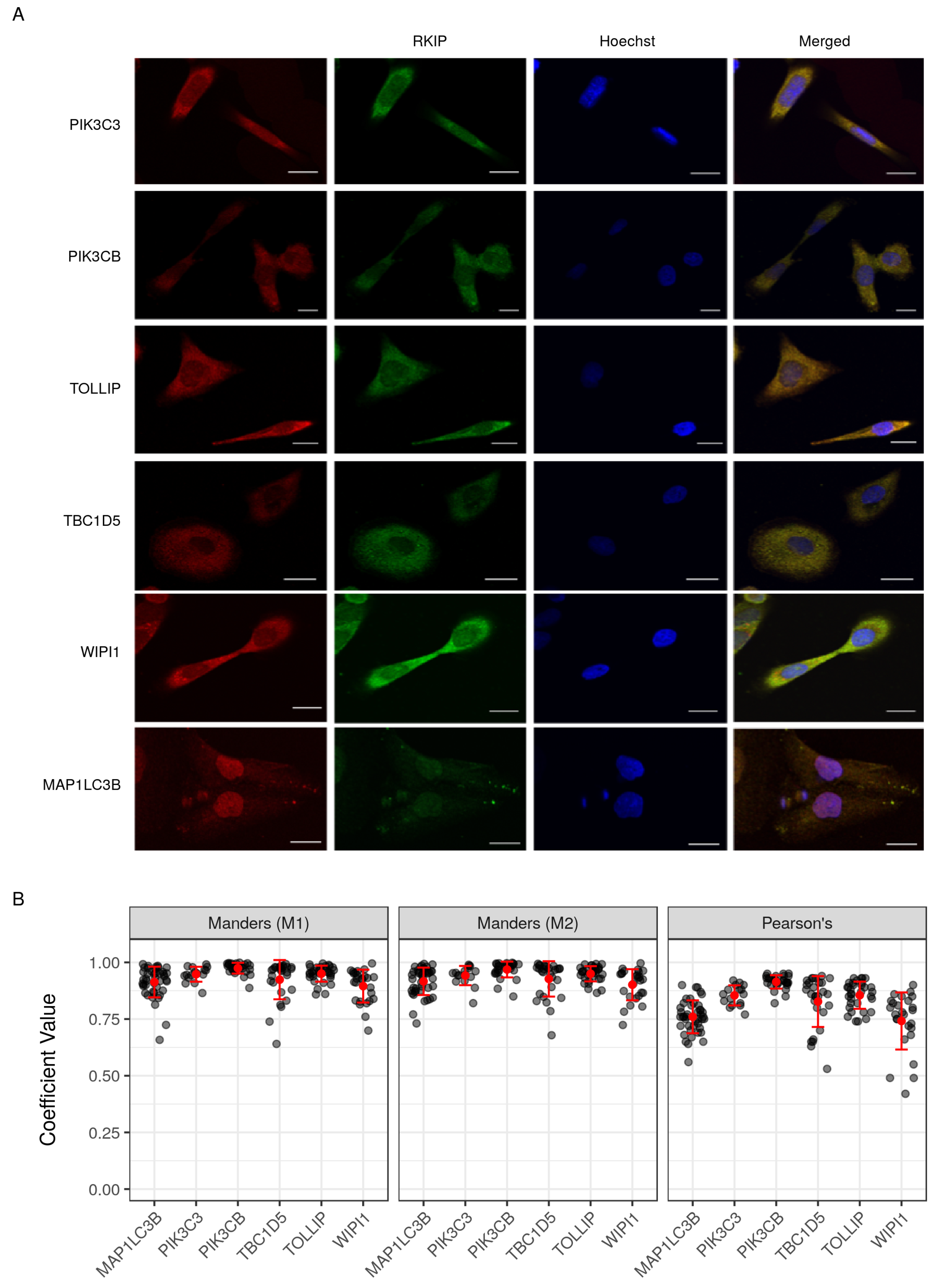
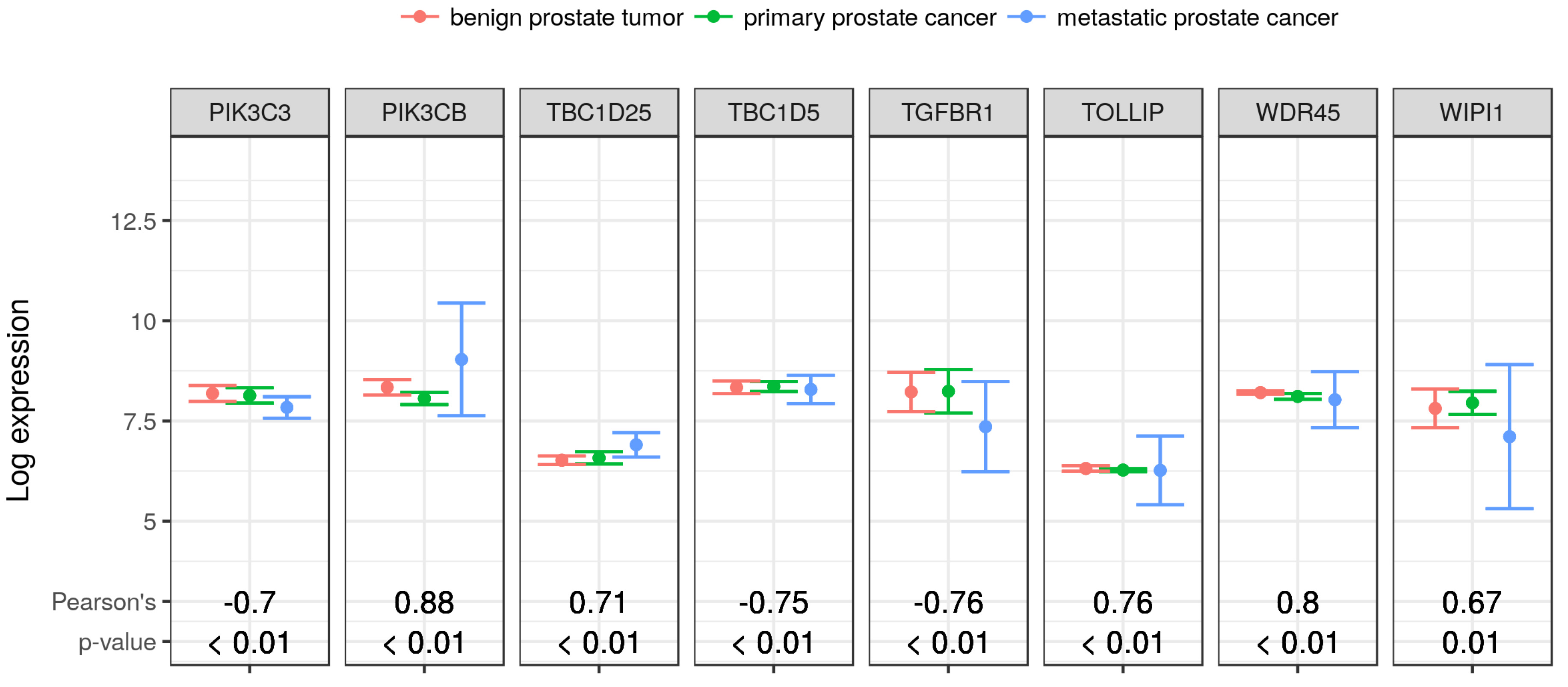
2.7. Validation of Selected Gene Product Correlations with RKIP/PEBP1
3. Discussion
4. Materials and Methods
4.1. Data and Annotation Sources
4.2. Weighted-Gene Co-Expression Network Analysis
4.3. Protein–Protein Interactions
4.4. Transcription Regulators Analysis
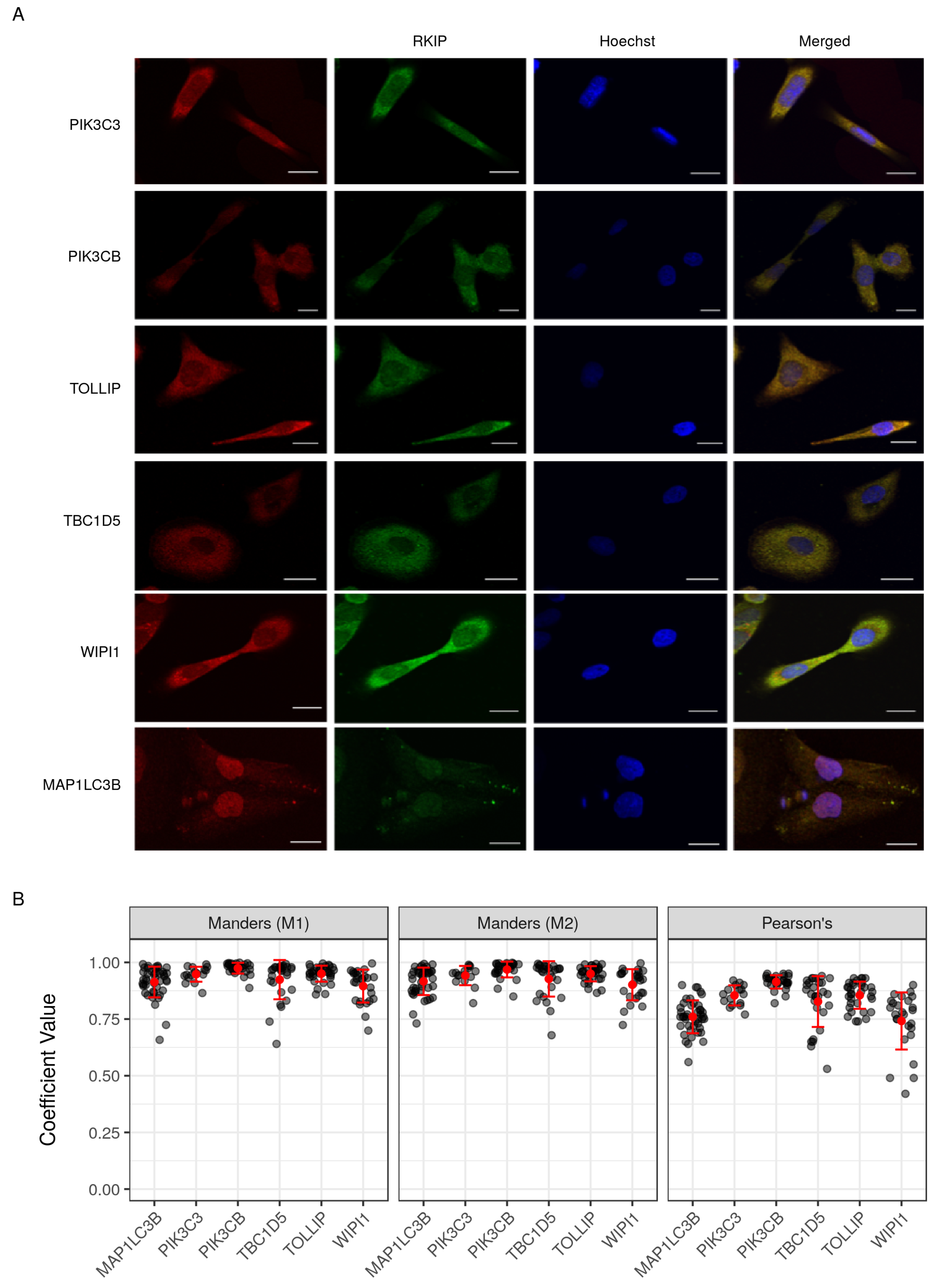
4.5. Cell Culture and Immunocytochemistry
4.6. Co-Localization Image Analysis
4.7. Software Environment and Reproducibility
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| RKIP | Raf Kinase Inhibitor Protein |
| PEBP1 | Phosphatidylethanolamine Binding Protein 1 |
| PEB | Phosphatidylethanolamine Binding |
| EMT | Epithelial to Mesenchymal Transition |
| WGCNA | Weighted-Gene Co-expression Network Analysis |
| GO | Gene Ontology |
| TOM | Topological Overlap Matrix |
| PC | Principal Component |
| ROI | Region of Interest |
| RNA-Seq | RNA Sequencing |
| ChIP-Seq | Chromatin Immunopreciptation followed by DNA Sequencing |
| NCI | National Cancer Institute |
Appendix A. A Note on Reproducing the Analysis
Appendix A.1. Setting up the Docker Environment
- $ docker pull bcmslab/rkip
- $ docker run -it bcmslab/rkip bash
Appendix A.2. Obtaining the Source Code
- ’get_data.R’ This script downloads several datasets from different sources in preparation of the analysis
- ’analysis.R’ This script loads the required libraries, downloads the data and runs all the steps of the analysis described in the manuscript
- ’figures/’ A sub-folder with a separate file for each graph in the manuscript.
- ’tables/’ A sub-folder with a separate file for each table in the manuscript.
- $ git clone https://github.com/BCMSLab/rkip
Appendix A.3. Running the Analysis
- $ cd rkip/analysis/
- $ make
Appendix A.4. Details of the R Environment
References
- Beach, S.; Tang, H.; Park, S.; Dhillon, A.S.; Keller, E.T.; Kolch, W.; Yeung, K.C. Snail is a repressor of RKIP transcription in metastatic prostate cancer cells. Oncogene 2008, 27, 2243–2248. [Google Scholar] [CrossRef] [PubMed]
- Farooqi, A.A.; Li, Y.; Sarkar, F.H. The biological complexity of RKIP signaling in human cancers. Exp. Mol. Med. 2015, 47, e185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Noh, H.S.; Hah, Y.S.; Zada, S.; Ha, J.H.; Sim, G.; Hwang, J.S.; Lai, T.H.; Nguyen, H.Q.; Park, J.Y.; Kim, H.J.; et al. PEBP1, a RAF kinase inhibitory protein, negatively regulates starvation-induced autophagy by direct interaction with LC3. Autophagy 2016, 12, 2183–2196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, B.; Horvath, S. A General Framework for Weighted Gene Co-Expression Network Analysis. Stat. Appl. Genet. Mol. Biol. 2005, 4, 17. [Google Scholar] [CrossRef] [PubMed]
- Horvath, S.; Dong, J. Geometric interpretation of gene coexpression network analysis. PLoS Comput. Biol. 2008, 4. [Google Scholar] [CrossRef] [PubMed]
- Langfelder, P.; Luo, R.; Oldham, M.C.; Horvath, S. Is my network module preserved and reproducible? PLoS Comput. Biol. 2011. [Google Scholar] [CrossRef] [PubMed]
- Varambally, S.; Yu, J.; Laxman, B.; Rhodes, D.R.; Mehra, R.; Tomlins, S.A.; Shah, R.B.; Chandran, U.; Monzon, F.A.; Becich, M.J.; et al. Integrative genomic and proteomic analysis of prostate cancer reveals signatures of metastatic progression. Cancer Cell 2005, 8, 393–406. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baca, S.C.; Prandi, D.; Lawrence, M.S.; Mosquera, J.M.; Romanel, A.; Drier, Y.; Park, K.; Kitabayashi, N.; MacDonald, T.Y.; Ghandi, M.; et al. Punctuated evolution of prostate cancer genomes. Cell 2013, 153, 666–677. [Google Scholar] [CrossRef] [PubMed]
- Barbieri, C.E.; Baca, S.C.; Lawrence, M.S.; Demichelis, F.; Blattner, M.; Theurillat, J.P.; White, T.A.; Stojanov, P.; Van Allen, E.; Stransky, N.; et al. Exome sequencing identifies recurrent SPOP, FOXA1 and MED12 mutations in prostate cancer. Nat. Genet. 2012, 44, 685. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Coleman, I.; Morrissey, C.; Zhang, X.; True, L.D.; Gulati, R.; Etzioni, R.; Bolouri, H.; Montgomery, B.; White, T.; et al. Substantial interindividual and limited intraindividual genomic diversity among tumors from men with metastatic prostate cancer. Nat. Med. 2016, 22, 369. [Google Scholar] [CrossRef] [PubMed]
- Gao, P.; Postiglione, M.P.; Krieger, T.G.; Hernandez, L.; Wang, C.; Han, Z.; Streicher, C.; Papusheva, E.; Insolera, R.; Chugh, K.; et al. Deterministic progenitor behavior and unitary production of neurons in the neocortex. Cell 2014, 159, 775–788. [Google Scholar] [CrossRef] [PubMed]
- Taylor, B.S.; Schultz, N.; Hieronymus, H.; Gopalan, A.; Xiao, Y.; Carver, B.S.; Arora, V.K.; Kaushik, P.; Cerami, E.; Reva, B.; et al. Integrative Genomic Profiling of Human Prostate Cancer. Cancer Cell 2010, 18, 11–22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robinson, D.; Van Allen, E.M.; Wu, Y.M.; Schultz, N.; Lonigro, R.J.; Mosquera, J.M.; Montgomery, B.; Taplin, M.E.; Pritchard, C.C.; Attard, G.; et al. Integrative clinical genomics of advanced prostate cancer. Cell 2015, 161, 1215–1228. [Google Scholar] [CrossRef] [PubMed]
- Cancer Genome Atlas Research Network. The Molecular Taxonomy of Primary Prostate Cancer. Cell 2015, 163, 1011–1025. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lebovitz, C.B.; Robertson, A.G.; Goya, R.; Jones, S.J.; Morin, R.D.; Marra, M.A.; Gorski, S.M. Cross-cancer profiling of molecular alterations within the human autophagy interaction network. Autophagy 2015, 11, 1668–1687. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, L.; Zhang, W.; Zou, F.; Mei, L.; Wu, G.; Teng, Y. KLHL21, a novel gene that contributes to the progression of hepatocellular carcinoma. BMC Cancer 2016, 16, 815. [Google Scholar] [CrossRef] [PubMed]
- Jiang, X.; Bao, Y.; Liu, H.; Kou, X.; Zhang, Z.; Sun, F.; Qian, Z.; Lin, Z.; Li, X.; Liu, X.; Jiang, L.; Yang, Y. VPS34 stimulation of p62 phosphorylation for cancer progression. Oncogene 2017, 36, 6850–6862. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khalil, B.D.; Hsueh, C.; Cao, Y.; Saab, W.F.; Wang, Y.; Condeelis, J.S.; Bresnick, A.R.; Backer, J.M. GPCR signaling mediates tumor metastasis via PI3Kβ. Cancer Res. 2016, 76, 2944–2953. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, W.; Salto-Tellez, M.; Palanisamy, N.; Ganesan, K.; Hou, Q.; Tan, L.K.; Sii, L.H.; Ito, K.; Tan, B.; Wu, J.; et al. Targets of genome copy number reduction in primary breast cancers identified by integrative genomics. Genes Chromosom. Cancer 2007, 46, 288–301. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Clarke, P.A.; Davis, R.; Mumuni, S.; Kwabi-Addo, B. Sex steroid-induced DNA methylation changes and inflammation response in prostate cancer. Cytokine 2016, 86, 110–118. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, T.; Yazbeck, C.; Maalouly, G.; Baz, M.; Haddad, F.; Sabbagh, C.; Chahine, G. TGFBR1*6A polymorphism in Sporadic and Familial colorectal carcinoma: A case-control study and systematic literature review. J. Gastrointest. Cancer 2014, 45, 441–447. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.W.; He, C.Y.; Sun, L.P.; Xu, Q.; Xing, C.Z.; Yuan, Y. The DNA repair gene ERCC6 rs1917799 polymorphism is associated with gastric cancer risk in Chinese. Asian Pac. J. Cancer Prev. 2013, 14, 6103–6108. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Z.; Zhang, G.; Li, W. Elevated expression of ERCC6 confers resistance to 5-Fluorouracil and is associated with poor patient survival in colorectal cancer. DNA Cell Biol. 2017, 36. [Google Scholar] [CrossRef] [PubMed]
- Avissar-Whiting, M.; Koestler, D.C.; Houseman, E.A.; Christensen, B.C.; Kelsey, K.T.; Marsit, C.J. Polycomb group genes are targets of aberrant DNA methylation in renal cell carcinoma. Epigenetics 2011, 6, 703–709. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yin, R.; Guo, L.; Gu, J.; Li, C.; Zhang, W. Over expressing miR-19b-1 suppress breast cancer growth by inhibiting tumor microenvironment induced angiogenesis. Int. J. Biochem. Cell Biol. 2018, 97, 43–51. [Google Scholar] [CrossRef] [PubMed]
- Kuang, X.; Wei, C.; Zhang, T.; Yang, Z.; Chi, J.; Wang, L. miR-378 inhibits cell growth and enhances apoptosis in human myelodysplastic syndromes. Int. J. Oncol. 2016, 49, 1921–1930. [Google Scholar] [CrossRef] [PubMed]
- Guo, G.C.; Wang, J.X.; Han, M.L.; Zhang, L.P.; Li, L. microRNA-761 induces aggressive phenotypes in triple-negative breast cancer cells by repressing TRIM29 expression. Cell. Oncol. 2017, 40, 157–166. [Google Scholar] [CrossRef] [PubMed]
- Guo, W.; Wang, H.; Yang, Y.; Guo, S.; Zhang, W.; Liu, Y.; Yi, X.; Ma, J.; Zhao, T.; Liu, L.; et al. Down-regulated miR-23a contributes to the metastasis of cutaneous melanoma by promoting autophagy. Theranostics 2017, 7, 2231–2249. [Google Scholar] [CrossRef] [PubMed]
- Hatzl, S.; Geiger, O.; Kuepper, M.K.; Caraffini, V.; Seime, T.; Furlan, T.; Nussbaumer, E.; Wieser, R.; Pichler, M.; Scheideler, M.; et al. Increased expression of miR-23a mediates a loss of expression in the RAF kinase inhibitor protein RKIP. Cancer Res. 2016, 76, 3644–3654. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klionsky, D.J.; Emr, S.D. Autophagy as a regulated pathway of cellular degradation. Science 2000, 290, 1717–1721. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.; He, Y.; Qiu, X.; Yang, W.; Liu, W.; Li, X.; Li, Y.; Shen, H.M.; Wang, R.; Yue, Z.; Zhao, Y. Targeting the potent Beclin 1-UVRAG coiled-coil interaction with designed peptides enhances autophagy and endolysosomal trafficking. Proc. Natl. Acad. Sci. USA 2018, 115, E5669–E5678. [Google Scholar] [CrossRef] [PubMed]
- Vincent-Salomon, A.; Thiery, J.P. Host microenvironment in breast cancer development: epithelial-mesenchymal transition in breast cancer development. Breast Cancer Res. 2003, 5, 101–106. [Google Scholar] [CrossRef] [PubMed]
- Slabáková, E.; Pernicová, Z.; Slavíčková, E.; Staršíchová, A.; Kozubík, A.; Souček, K. TGF-β1-induced EMT of non-transformed prostate hyperplasia cells is characterized by early induction of SNAI2/Slug. Prostate 2011, 71, 1332–1343. [Google Scholar] [CrossRef] [PubMed]
- Shiota, M.; Zardan, A.; Takeuchi, A.; Kumano, M.; Beraldi, E.; Naito, S.; Zoubeidi, A.; Gleave, M.E. Clusterin mediates TGF-β-induced epithelial-mesenchymal transition and metastasis via Twist1 in prostate cancer cells. Cancer Res. 2012. [Google Scholar] [CrossRef] [PubMed]
- Stuart, J.M.; Segal, E.; Koller, D.; Kim, S.K. A gene-coexpression network for global discovery of conserved genetic modules. Science 2003, 302, 249–255. [Google Scholar] [CrossRef] [PubMed]
- Carter, S.L.; Brechbühler, C.M.; Griffin, M.; Bond, A.T. Gene co-expression network topology provides a framework for molecular characterization of cellular state. Bioinformatics 2004, 20, 2242–2250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, A.; Datta, S.; Smith, G.; Campbell, P.; Bentley, R.; McKenzie, H.; Bender, D.; Harris, A.; Goodwin, T.; Parish, J.; et al. Oxford Dictionary of Biochemistry and Molecular Biology; Oxford University Press: Oxford, UK, 1997. [Google Scholar]
- Timmerman, L.A.; Grego-Bessa, J.; Raya, A.; Bertrán, E.; Pérez-Pomares, J.M.; Díez, J.; Aranda, S.; Palomo, S.; McCormick, F.; Izpisúa-Belmonte, J.C.; et al. Notch promotes epithelial-mesenchymal transition during cardiac development and oncogenic transformation. Genes Dev. 2004, 18, 99–115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Langfelder, P.; Horvath, S. WGCNA: An R package for weighted correlation network analysis. BMC Bioinform. 2008, 9, 559. [Google Scholar] [CrossRef] [PubMed]
- Szklarczyk, D.; Morris, J.H.; Cook, H.; Kuhn, M.; Wyder, S.; Simonovic, M.; Santos, A.; Doncheva, N.T.; Roth, A.; Bork, P.; et al. The STRING database in 2017: Quality-controlled protein-protein association networks, made broadly accessible. Nucleic Acids Res. 2017, 45, D362–D368. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.; Ortiz, J.A.; Taing, L.; Meyer, C.A.; Lee, B.; Zhang, Y.; Shin, H.; Wong, S.S.; Ma, J.; Lei, Y.; et al. Cistrome: An integrative platform for transcriptional regulation studies. Genome Biol. 2011, 12, R83. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, M.; Nguyen, H.; Lai, T.; Kim, D. MiRCancerdb: A database for correlation analysis between microRNA and gene expression in cancer. BMC Res. Notes 2018, 11, 103. [Google Scholar] [CrossRef] [PubMed]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schmid, B.; Tinevez, J.Y.; White, D.J.; Hartenstein, V.; Eliceiri, K.; Tomancak, P.; Cardona, A. Fiji: An open-source platform for biological-image analysis. Nature Methods 2012, 9, 676–682. [Google Scholar] [CrossRef] [PubMed]
- R Development Core Team. R: A Language and Environment for Statistical Computing; the R Foundation for Statistical Computing: Vienna, Austria, 2011. [Google Scholar]
- Huber, W.; Carey, V.J.; Gentleman, R.; Anders, S.; Carlson, M.; Carvalho, B.S.; Bravo, H.C.; Davis, S.; Gatto, L.; Girke, T.; et al. Orchestrating high-throughput genomic analysis with Bioconductor. Nat. Methods 2015, 12, 115–121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Merkel, D. Docker: Lightweight Linux containers for consistent development and deployment. Linux J. 2014, 2014, 2. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Study ID | Samples | Genes | Reference |
|---|---|---|---|
| prad.broad.2013 | 7 | 150 | [8] |
| prad.broad | 20 | 143 | [9] |
| prad.fhcrc | 171 | 149 | [10] |
| prad.mskcc.cheny1.organoids.2014 | 10 | 148 | [11] |
| prad.mskcc | 150 | 151 | [12] |
| prad.su2c.2015 | 118 | 152 | [13] |
| prad.tcga.pub | 333 | 152 | [14] |
| prad.tcga | 498 | 152 | [14] |
| Module | Autophagy | Epithelial to Mesenchymal Transition | Phosphatidylethanolamine Binding |
|---|---|---|---|
| blue | ABL1, ANXA7, ARSB, BNIP1, VPS51, CLTC, DAP, FOXO1, HMGB1, IFI16, NPC1, S100A8, S100A9, STK11, TMBIM6, TP53, UVRAG, SRPX, BECN1, USP10, ULK2, PLEKHM1, TECPR2, HDAC6, OPTN, RNF41, RGS19, ATG7, TM9SF1, WDR45, PARK7, VPS13A, VPS39, ULK3, PTPN22, TMEM208, NRBF2, RAB39A, FNBP1L, WIPI1, MAP1S, DRAM1, SUPT20H, VPS11, TIGAR, VPS18, PHF23, MAP1LC3B, VMP1, C19orf12, ATG10, EVA1A, WDR24, ATG4C, TRIM5, LRSAM1, RAB39B, LRRK2, DRAM2, SMCR8 | BMP2, BMP7, FGFR2, FOXF2, HNRNPAB, RBPJ, LOXL2, S100A4, SNAI2, TGFB1, TGFB2, TGFBR3, WNT5A, DLG5, NOG, DDX17, LEF1, EPB41L5, FAM83D, LOXL3, RFLNB | ANXA11, MFGE8, PLTP, PEMT, CD300A, MAP1LC3A |
| brown | CTSD, RAB8A, TBC1D25, PIK3C3, PIK3CB, RAB1A, VCP, TFEB, ULK1, SQSTM1, HAP1, ATG5, NAPSA, RUBCN, TBC1D5, SIRT2, ATG4B, TECPR1, CHMP2B, VPS41, TRIM17, TOLLIP, ZKSCAN3, CHMP4B, RAB12, C9orf72 | AMELX, CTNNB1, HGF, HIF1A, SNAI1, SOX9, TGFBR1, HMGA2 | NF1, PEBP1, ESYT2 |
| yellow | ITGB4, PGC, USP13, TMEM59, RB1CC1, GABARAPL2, CLEC16A, UBQLN2, SH3GLB1, WDR41, VTI1A | GSK3B, NOTCH1, WNT11, CUL7, WNT4 |
| Family | Protein | Name | Main Function |
|---|---|---|---|
| WD Repeat Domain | WDR45 | WD Repeat Domain 45 | Frequently mutated in lung adenocarcinomas [15]. |
| WIPI1 | WD Repeat Domain, Phosphoinositide Interacting 1 | High expression is associated with survival in hepatocellular carcinoma patients [16]. | |
| PI3K | PIK3C3 | Phosphatidylinositol 3-Kinase Catalytic Subunit Type 3 | Promote cancer growth through p62 [17]. |
| PIK3CB | Phosphatidylinositol-4, 5-Bisphosphate 3-Kinase Catalytic Subunit Beta | Mediates cancer metastasis [18]. | |
| TBC | TBC1D5 | TBC1 Domain Family Member 5 | Reduced copy number in breast cancer [19]. |
| TBC1D25 | TBC1 Domain Family Member 25 | ||
| Other | TOLLIP | Toll Interacting Protein | Hypermethylated in response to sex hormones in prostate cancer cells [20]. |
| TGFBR1 | Transforming Growth Factor Beta Receptor 1 | Multiple polymorphisms are associated with cancer development [21]. |
| Factor | Name | Function |
|---|---|---|
| ERCC6 | ERCC Excision Repair 6, Chromatin Remodeling Factor | A DNA-binding protein that is important in transcription-coupled excision repair. Several polymorphisms the gene coding region were associated with susceptibility to development of cancer and chemoresistancy [22,23]. |
| VEZF1 | Vascular Endothelial Zinc Finger 1 | A transcriptional regulatory protein that is involved in angiogenesis. Contribute to the epigenetic aberrations and the associated tumorigenesis [24,25]. |
| hsa-miR-378c | Close relative (hsa-miR-378a) | Inhibits cell growth and enhances apoptosis in cancer [26]. |
| hsa-miR-761 | Enhances cancer growth, migration and invasion [27]. | |
| hsa-miR-23c | Close relative (hsa-miR-23a) | Associated with autophagy, loss of RKIP/PEBP1 and multiple tumors [28,29]. |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ahmed, M.; Lai, T.H.; Zada, S.; Hwang, J.S.; Pham, T.M.; Yun, M.; Kim, D.R. Functional Linkage of RKIP to the Epithelial to Mesenchymal Transition and Autophagy during the Development of Prostate Cancer. Cancers 2018, 10, 273. https://doi.org/10.3390/cancers10080273
Ahmed M, Lai TH, Zada S, Hwang JS, Pham TM, Yun M, Kim DR. Functional Linkage of RKIP to the Epithelial to Mesenchymal Transition and Autophagy during the Development of Prostate Cancer. Cancers. 2018; 10(8):273. https://doi.org/10.3390/cancers10080273
Chicago/Turabian StyleAhmed, Mahmoud, Trang Huyen Lai, Sahib Zada, Jin Seok Hwang, Trang Minh Pham, Miyong Yun, and Deok Ryong Kim. 2018. "Functional Linkage of RKIP to the Epithelial to Mesenchymal Transition and Autophagy during the Development of Prostate Cancer" Cancers 10, no. 8: 273. https://doi.org/10.3390/cancers10080273