Loss of One or Two PATZ1 Alleles Has a Critical Role in the Progression of Thyroid Carcinomas Induced by the RET/PTC1 Oncogene

 ,
,  ,
, 

Abstract
:1. Introduction
2. Results
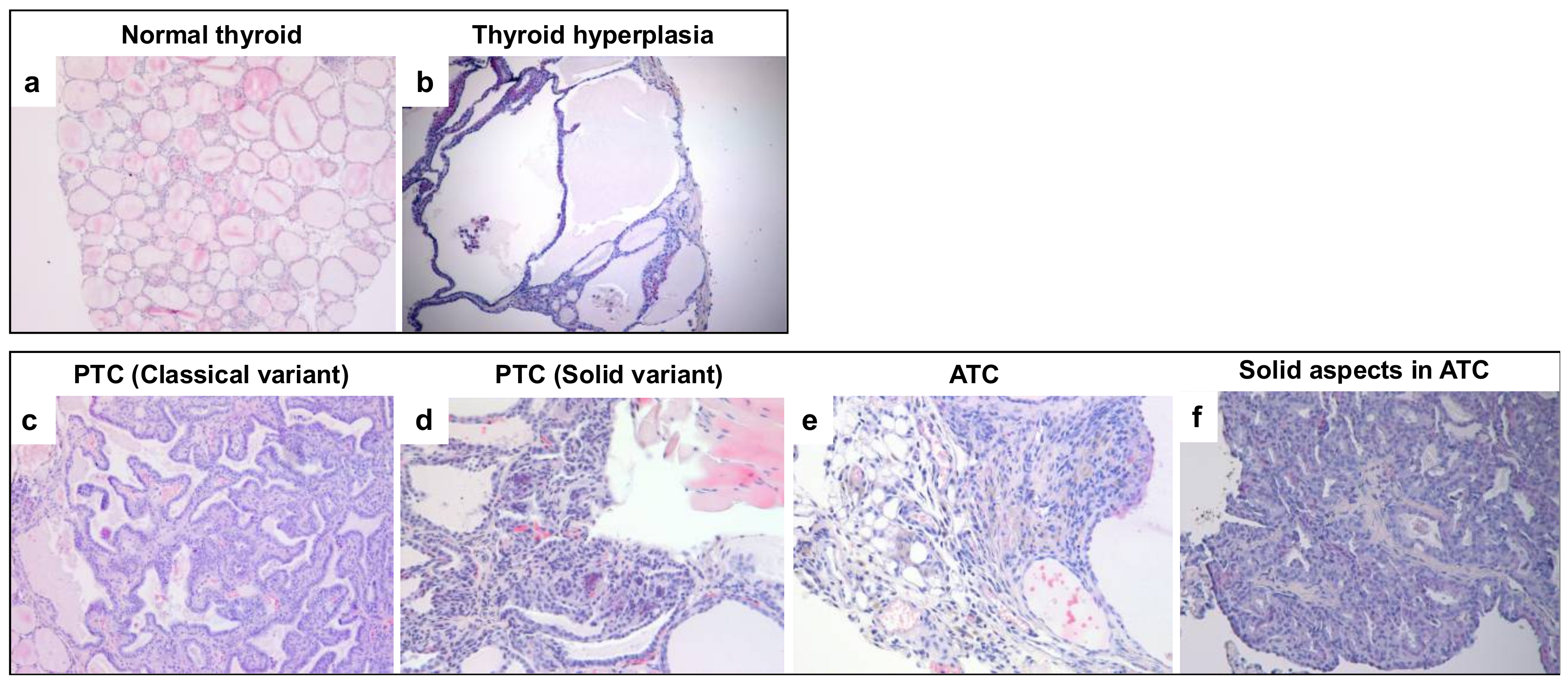
2.1. RET/PTC1 Transgenic Mice Show Higher Incidence of Thyroid Carcinomas in Absence of PATZ1
2.2. Loss of One or Two Patz1 Alleles Enhances Thyroid Tumor Aggressiveness in RET/PTC1 Mice
2.3. Patz1-Null Mutation Enhances Proliferation of Thyroid Cancer Cells in RET/PTC1TG Mice
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Genotyping
4.3. Histopathology and Immunohistochemistry
4.4. Statistical Analysis
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Rahib, L.; Smith, B.D.; Aizenberg, R.; Rosenzweig, A.B.; Fleshman, J.M.; Matrisian, L.M. Projecting Cancer Incidence and Deaths to 2030: The Unexpected Burden of Thyroid, Liver, and Pancreas Cancers in the United States. Cancer Res. 2014, 74, 2913–2921. [Google Scholar] [CrossRef]
- Lloyd, R.V.; Osamura, R.Y.; Klöppel, G.; Rosai, J. (Eds.) WHO Classification of Tumours of Endocrine Organs, 4th ed.; IARC: Lyon, France, 2004; pp. 66–104. ISBN 978-92-832-4493-6. [Google Scholar]
- Molinaro, E.; Romei, C.; Biagini, A.; Sabini, E.; Agate, L.; Mazzeo, S.; Materazzi, G.; Sellari-Franceschini, S.; Ribechini, A.; Torregrossa, L.; et al. Anaplastic thyroid carcinoma: From clinicopathology to genetics and advanced therapies. Nat. Rev. Endocrinol. 2017, 13, 644–660. [Google Scholar] [CrossRef] [PubMed]
- Nikiforov, Y.E. RET/PTC rearrangement in thyroid tumors. Endocr. Pathol. 2002, 13, 3–16. [Google Scholar] [CrossRef] [PubMed]
- Grieco, M.; Santoro, M.; Berlingieri, M.T.; Melillo, R.M.; Donghi, R.; Bongarzone, I.; Pierotti, M.A.; Della Porta, G.; Fusco, A.; Vecchio, G. PTC is a novel rearranged form of the ret proto-oncogene and is frequently detected in vivo in human thyroid papillary carcinomas. Cell 1990, 60, 557–563. [Google Scholar] [CrossRef]
- Santoro, M.; Dathan, N.A.; Berlingieri, M.T.; Bongarzone, I.; Paulin, C.; Grieco, M.; Pierotti, M.A.; Vecchio, G.; Fusco, A. Molecular characterization of RET/PTC3; a novel rearranged version of the RET proto-oncogene in a human thyroid papillary carcinoma. Oncogene 1994, 9, 509–516. [Google Scholar] [PubMed]
- Romei, C.; Ciampi, R.; Elisei, R. A comprehensive overview of the role of the RET proto-oncogene in thyroid carcinoma. Nat. Rev. Endocrinol. 2016, 12, 192–202. [Google Scholar] [CrossRef]
- Cyniak-Magierska, A.; Brzeziańska, E.; Januszkiewicz-Caulier, J.; Jarzab, B.; Lewiński, A. Prevalence of RAS point mutations in papillary thyroid carcinoma; a novel mutation at codon 31 of K-RAS. Exp. Clin. Endocrinol. Diabetes 2007, 115, 594–599. [Google Scholar] [CrossRef] [PubMed]
- Fedele, M.; Crescenzi, E.; Cerchia, L. The POZ/BTB and AT-Hook Containing Zinc Finger 1 (PATZ1) Transcription Regulator: Physiological Functions and Disease Involvement. Int. J. Mol. Sci. 2017, 18, 2524. [Google Scholar] [CrossRef]
- Chiappetta, G.; Valentino, T.; Vitiello, M.; Pasquinelli, R.; Monaco, M.; Palma, G.; Sepe, R.; Luciano, A.; Pallante, P.; Palmieri, D.; et al. PATZ1 acts as a tumor suppressor in thyroid cancer via targeting p53-dependent genes involved in EMT and cell migration. Oncotarget 2015, 6, 5310–5323. [Google Scholar] [CrossRef] [PubMed]
- Iesato, A.; Nakamura, T.; Izumi, H.; Uehara, T.; Ito, K.I. PATZ1 knockdown enhances malignant phenotype in thyroid epithelial follicular cells and thyroid cancer cells. Oncotarget 2017, 8, 82754–82772. [Google Scholar] [CrossRef] [PubMed]
- Santoro, M.; Chiappetta, G.; Cerrato, A.; Salvatore, D.; Zhang, L.; Manzo, G.; Picone, A.; Portella, G.; Santelli, G.; Vecchio, G.; et al. Development of thyroid papillary carcinomas secondary to tissue-specific expression of the RET/PTC1 oncogene in transgenic mice. Oncogene 1996, 12, 1821–1826. [Google Scholar] [PubMed]
- Jhiang, S.M.; Sagartz, J.E.; Tong, Q.; Parker-Thornburg, J.; Capen, C.C.; Cho, J.Y.; Xing, S.; Ledent, C. Targeted expression of the ret/PTC1 oncogene induces papillary thyroid carcinomas. Endocrinology 1996, 137, 375–378. [Google Scholar] [CrossRef] [PubMed]
- Powell, D.J., Jr.; Russell, J.; Nibu, K.; Li, G.; Rhee, E.; Liao, M.; Goldstein, M.; Keane, W.M.; Santoro, M.; Fusco, A.; et al. The RET/PTC3 oncogene: Metastatic solid-type papillary carcinomas in murine thyroids. Cancer Res. 1998, 58, 5523–5528. [Google Scholar] [PubMed]
- La Perle, K.M.; Jhiang, S.M.; Capen, C.C. Loss of p53 promotes anaplasia and local invasion in ret/PTC1-induced thyroid carcinomas. Am. J. Pathol. 2000, 157, 671–677. [Google Scholar] [CrossRef]
- Powell, D.J., Jr.; Russell, J.P.; Li, G.; Kuo, B.A.; Fidanza, V.; Huebner, K.; Rothstein, J.L. Altered gene expression in immunogenic poorly differentiated thyroid carcinomas from RET/PTC3p53-/- mice. Oncogene 2001, 20, 3235–3246. [Google Scholar] [CrossRef] [PubMed]
- Giordano, T.J.; Kuick, R.; Thomas, D.G.; Misek, D.E.; Vinco, M.; Sanders, D.; Zhu, Z.; Ciampi, R.; Roh, M.; Shedden, K.; et al. Molecular classification of papillary thyroid carcinoma: Distinct BRAF, RAS, and RET/PTC mutation-specific gene expression profiles discovered by DNA microarray analysis. Oncogene 2005, 24, 6646–6656. [Google Scholar] [CrossRef] [PubMed]
- Leone, V.; Ferraro, A.; Schepis, F.; Federico, A.; Sepe, R.; Arra, C.; Langella, C.; Palma, G.; De Lorenzo, C.; Troncone, G.; et al. The cl2/dro1/ccdc80 null mice develop thyroid and ovarian neoplasias. Cancer Lett. 2015, 357, 535–541. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Antico Arciuch, V.G.; Russo, M.A.; Dima, M.; Kang, K.S.; Dasrath, F.; Liao, X.H.; Refetoff, S.; Montagna, C.; Di Cristofano, A. Thyrocyte-specific inactivation of p53 and Pten results in anaplastic thyroid carcinomas faithfully recapitulating human tumors. Oncotarget 2011, 2, 1109–1126. [Google Scholar] [CrossRef] [PubMed]
- Charles, R.P.; Silva, J.; Iezza, G.; Phillips, W.A.; McMahon, M. Activating BRAF and PIK3CA mutations cooperate to promote anaplastic thyroid carcinogenesis. Mol. Cancer Res. 2014, 12, 979–986. [Google Scholar] [CrossRef] [PubMed]
- Valentino, T.; Palmieri, D.; Vitiello, M.; Simeone, A.; Palma, G.; Arra, C.; Chieffi, P.; Chiariotti, L.; Fusco, A.; Fedele, M. Embryonic defects and growth alteration in mice with homozygous disruption of the Patz1 gene. J. Cell Physiol. 2013, 228, 646–653. [Google Scholar] [CrossRef] [PubMed]
- Fedele, M.; Cerchia, L.; Chiappetta, G. The Tumor Suppressive Role of PATZ1 in Thyroid Cancer: A Matter of Epithelial-Mesenchymal Transition. Chemother. Open Access 2016, 5, 198. [Google Scholar] [CrossRef]
- Vitiello, M.; Valentino, T.; De Menna, M.; Crescenzi, E.; Francesca, P.; Rea, D.; Arra, C.; Fusco, A.; De Vita, G.; Cerchia, L.; et al. PATZ1 is a target of miR-29b that is induced by Ha-Ras oncogene in rat thyroid cells. Sci. Rep. 2016, 6, 25268. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-Gonzalez, R.; Bologna-Molina, R.; Carreon-Burciaga, R.G.; Gómezpalacio-Gastelum, M.; Molina-Frechero, N.; Salazar-Rodríguez, S. Papillary thyroid carcinoma: Differential diagnosis and prognostic values of its different variants: Review of the literature. ISRN Oncol. 2011, 2011, 915925. [Google Scholar] [CrossRef] [PubMed]
- Nikiforov, Y.E.; Erickson, L.A.; Nikiforova, M.N.; Caudill, C.M.; Lloyd, R.V. Solid variant of papillary thyroid carcinoma: Incidence, clinical-pathologic characteristics, molecular analysis, and biologic behavior. Am. J. Surg. Pathol. 2001, 25, 1478–1484. [Google Scholar] [CrossRef] [PubMed]
- Ohashi, R.; Kawahara, K.; Namimatsu, S.; Igarashi, T.; Sakatani, T.; Sugitani, I.; Naito, Z. Clinicopathological significance of a solid component in papillary thyroid carcinoma. Histopathology 2017, 70, 775–781. [Google Scholar] [CrossRef] [PubMed]
- Ohashi, R.; Murase, Y.; Matsubara, M.; Watarai, Y.; Igarashi, T.; Sugitani, I.; Naito, Z. Fine needle aspiration cytology of the papillary thyroid carcinoma with a solid component: A cytological and clinical correlation. Diag. Cytopathol. 2017, 45, 391–398. [Google Scholar] [CrossRef] [PubMed]
- Ohashi, R.; Kawahara, K.; Namimatsu, S.; Okamura, R.; Igarashi, T.; Sugitani, I.; Naito, Z. Expression of MRP1 and ABCG2 is associated with adverse clinical outcomes of papillary thyroid carcinoma with a solid component. Hum. Pathol. 2017, 67, 11–17. [Google Scholar] [CrossRef]
- Yun, J.Y.; Kim, Y.A.; Choe, J.Y.; Min, H.; Lee, K.S.; Jung, Y.; Oh, S.; Kim, J.E. Expression of cancer stem cell markers is more frequent in anaplastic thyroid carcinoma compared to papillary thyroid carcinoma and is related to adverse clinical outcome. J. Clin. Pathol. 2014, 67, 25–133. [Google Scholar] [CrossRef] [PubMed]
- Hardy, R.G.; Vicente-Dueñas, C.; González-Herrero, I.; Anderson, C.; Flores, T.; Hughes, S.; Tselepis, C.; Ross, J.A.; Sánchez-García, I. Snail family transcription factors are implicated in thyroid carcinogenesis. Am. J. Pathol. 2007, 171, 1037–1046. [Google Scholar] [CrossRef] [PubMed]
- Salerno, P.; Garcia-Rostan, G.; Piccinin, S.; Bencivenga, T.C.; Di Maro, G.; Doglioni, C.; Basolo, F.; Maestro, R.; Fusco, A.; Santoro, M.; et al. TWIST1 plays a pleiotropic role in determining the anaplastic thyroid cancer phenotype. J. Clin. Endocrinol. Metab. 2011, 96, E772–E781. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buehler, D.; Hardin, H.; Shan, W.; Montemayor-Garcia, C.; Rush, P.S.; Asioli, S.; Chen, H.; Lloyd, R.V. Expression of epithelial-mesenchymal transition regulators SNAI2 and TWIST1 in thyroid carcinomas. Mod. Pathol. 2013, 26, 54–61. [Google Scholar] [CrossRef] [PubMed]
- Montemayor-Garcia, C.; Hardin, H.; Guo, Z.; Larrain, C.; Buehler, D.; Asioli, S.; Chen, H.; Lloyd, R.V. The role of epithelial mesenchymal transition markers in thyroid carcinoma progression. Endocr. Pathol. 2013, 24, 206–212. [Google Scholar] [CrossRef] [PubMed]
- McFadden, D.G.; Vernon, A.; Santiago, P.M.; Martinez-McFaline, R.; Bhutkar, A.; Crowley, D.M.; McMahon, M.; Sadow, P.M.; Jacks, T. p53 constrains progression to anaplastic thyroid carcinoma in a Braf-mutant mouse model of papillary thyroid cancer. Proc. Natl. Acad Sci. USA 2014, 111, E1600–E1609. [Google Scholar] [CrossRef] [PubMed]
- Knauf, J.A.; Ma, X.; Smith, E.P.; Zhang, L.; Mitsutake, N.; Liao, X.H.; Refetoff, S.; Nikiforov, Y.E.; Fagin, J.A. Targeted expression of BRAFV600E in thyroid cells of transgenic mice results in papillary thyroid cancers that undergo dedifferentiation. Cancer Res. 2005, 65, 4238–4245. [Google Scholar] [CrossRef] [PubMed]
- Rusinek, D.; Swierniak, M.; Chmielik, E.; Kowal, M.; Kowalska, M.; Cyplinska, R.; Czarniecka, A.; Piglowski, W.; Korfanty, J.; Chekan, M.; et al. BRAFV600E-Associated Gene Expression Profile: Early Changes in the Transcriptome, Based on a Transgenic Mouse Model of Papillary Thyroid Carcinoma. PLoS ONE 2015, 10, e0143688. [Google Scholar] [CrossRef] [PubMed]
- Kimura, E.T.; Nikiforova, M.N.; Zhu, Z.; Knauf, J.A.; Nikiforov, Y.E.; Fagin, J.A. High prevalence of BRAF mutations in thyroid cancer: Genetic evidence for constitutive activation of the RET/PTC-RAS-BRAF signaling pathway in papillary thyroid carcinoma. Cancer Res. 2003, 63, 1454–1457. [Google Scholar] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Pathological Lesion | RET/PTC1TG;Patz1+/+ (n = 22) | RET/PTC1TG;Patz1+/− (n = 24) | RET/PTC1TG;Patz1−/− (n = 6) |
|---|---|---|---|
| Thyroid carcinoma | 12 (54%) | 14 (58%) | 6 (100%) |
| Hyperplasia/goiter | 7 (32%) | 7 (29%) | 0 |
| Normal thyroid | 3 (14%) | 3 (12.5%) | 0 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Monaco, M.; Palma, G.; Vitiello, M.; Capiluongo, A.; D’Andrea, B.; Vuttariello, E.; Luciano, A.; Cerchia, L.; Chiappetta, G.; Arra, C.; et al. Loss of One or Two PATZ1 Alleles Has a Critical Role in the Progression of Thyroid Carcinomas Induced by the RET/PTC1 Oncogene. Cancers 2018, 10, 92. https://doi.org/10.3390/cancers10040092
Monaco M, Palma G, Vitiello M, Capiluongo A, D’Andrea B, Vuttariello E, Luciano A, Cerchia L, Chiappetta G, Arra C, et al. Loss of One or Two PATZ1 Alleles Has a Critical Role in the Progression of Thyroid Carcinomas Induced by the RET/PTC1 Oncogene. Cancers. 2018; 10(4):92. https://doi.org/10.3390/cancers10040092
Chicago/Turabian StyleMonaco, Mario, Giuseppe Palma, Michela Vitiello, Anna Capiluongo, Barbara D’Andrea, Emilia Vuttariello, Antonio Luciano, Laura Cerchia, Gennaro Chiappetta, Claudio Arra, and et al. 2018. "Loss of One or Two PATZ1 Alleles Has a Critical Role in the Progression of Thyroid Carcinomas Induced by the RET/PTC1 Oncogene" Cancers 10, no. 4: 92. https://doi.org/10.3390/cancers10040092
APA StyleMonaco, M., Palma, G., Vitiello, M., Capiluongo, A., D’Andrea, B., Vuttariello, E., Luciano, A., Cerchia, L., Chiappetta, G., Arra, C., Fusco, A., & Fedele, M. (2018). Loss of One or Two PATZ1 Alleles Has a Critical Role in the Progression of Thyroid Carcinomas Induced by the RET/PTC1 Oncogene. Cancers, 10(4), 92. https://doi.org/10.3390/cancers10040092