Genomic Characteristics of Invasive Mucinous Adenocarcinomas of the Lung and Potential Therapeutic Targets of B7-H3

 , , and
, , and
Abstract
1. Introduction
2. Results
2.1. Patient Characteristics
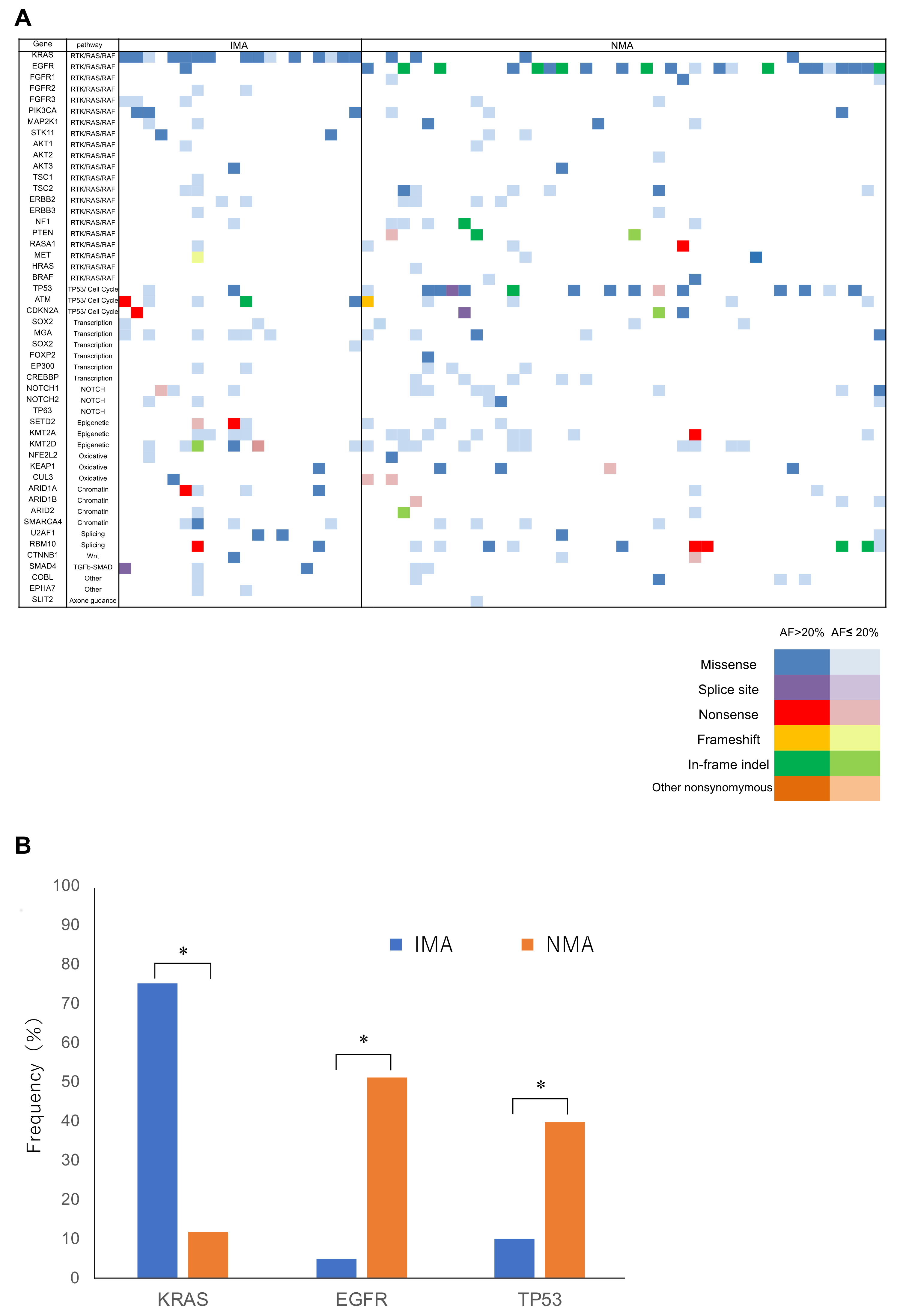
2.2. Panel Sequencing
2.3. In Silico Analysis
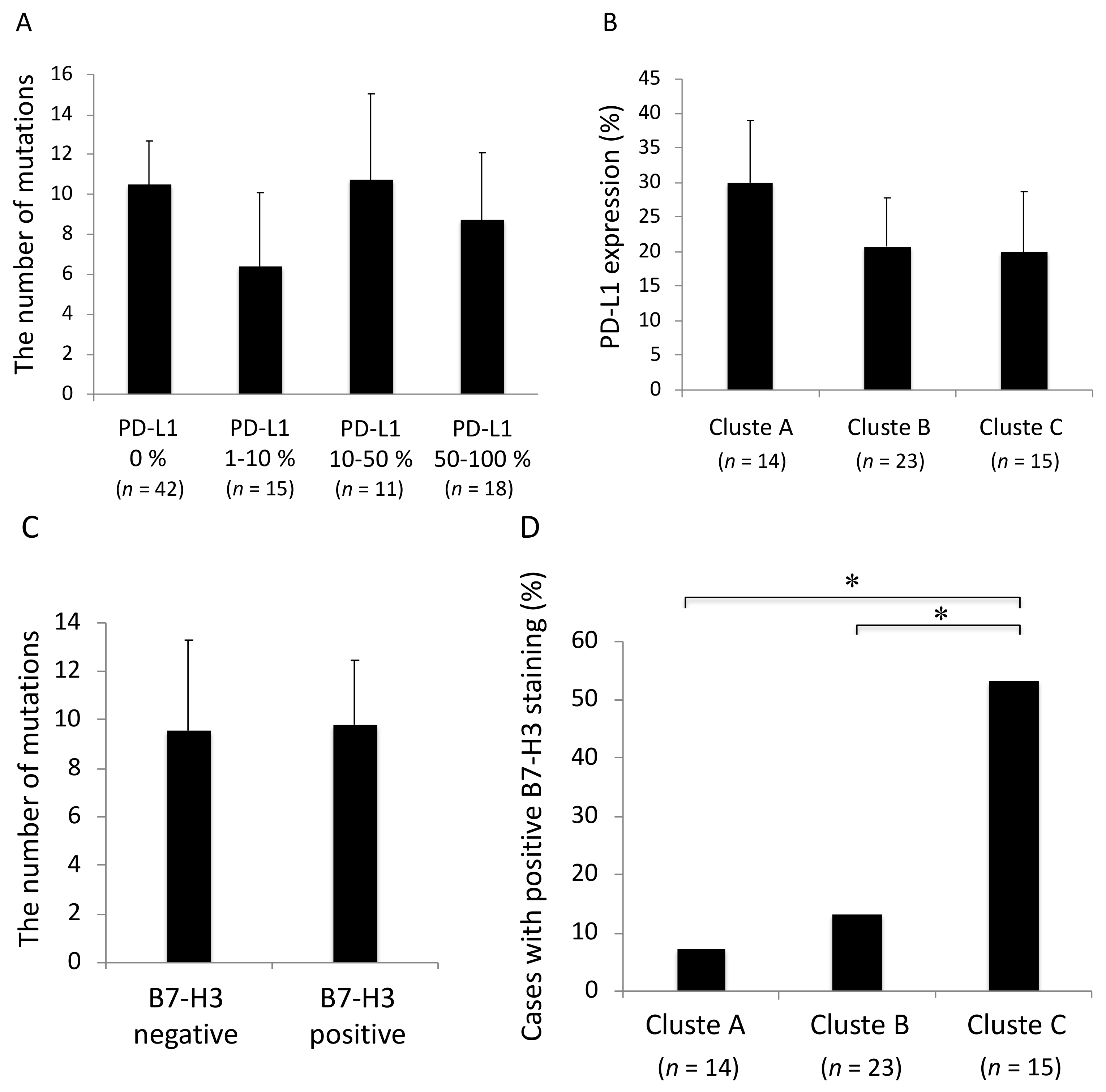
2.4. Correlation between Cluster Classification and Outcome
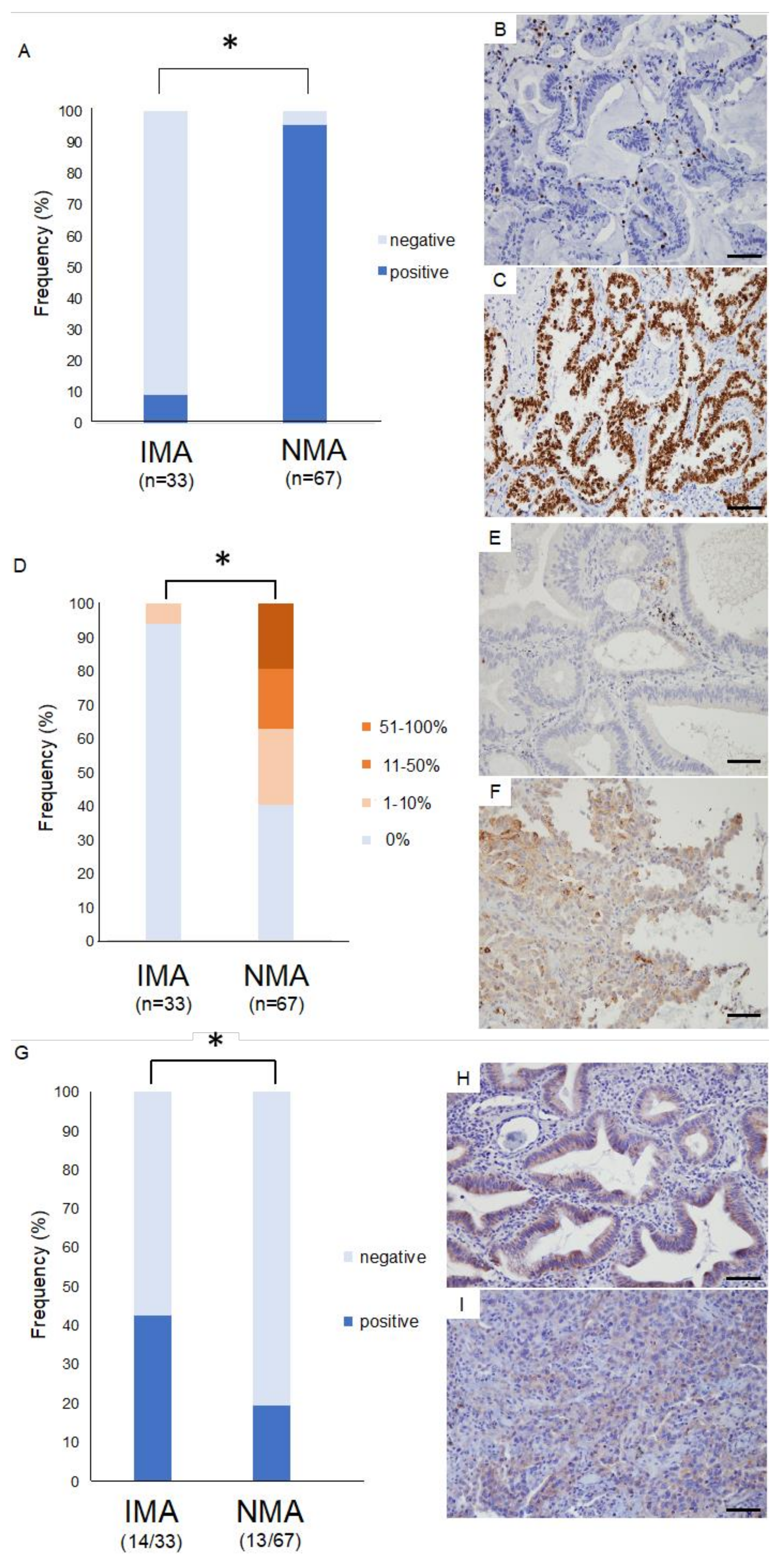
2.5. Immunohistology for Thyroid Transcription Factor 1 (TTF-1)
2.6. Immunohistology for Immunocheckpoint Proteins
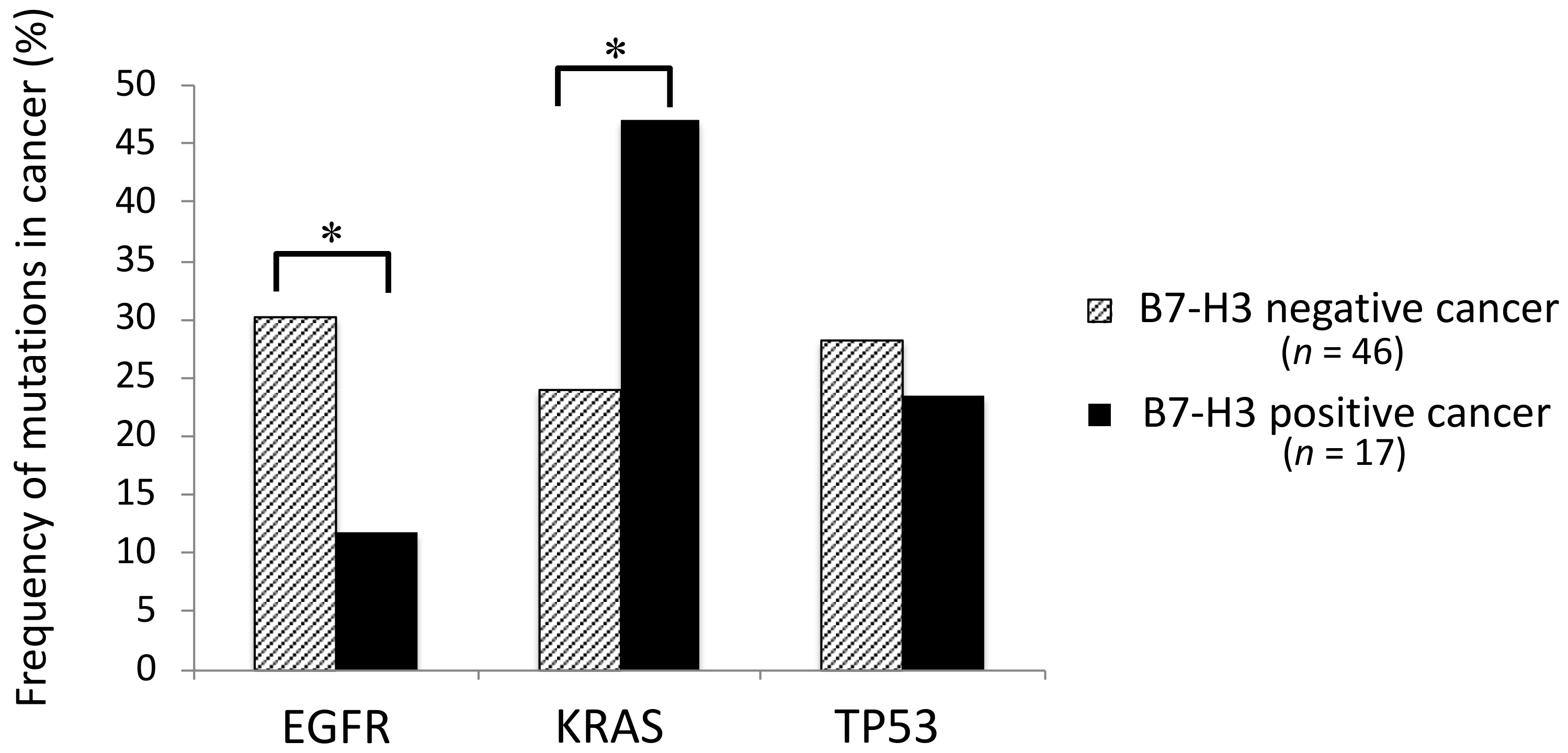
2.7. Association between the Mutation Profiles and Immunocheckpoint Molecules
3. Discussion
4. Methods
4.1. Patients and Sample Preparation
4.2. Targeted Deep Sequencing and Data Analysis
4.3. In Silico Clustering
4.4. Immunohistochemistry
4.5. Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Travis, W.D.; Brambilla, E.; Noguchi, M.; Nicholson, A.G.; Geisinger, K.R.; Yatabe, Y.; Beer, D.G.; Powell, C.A.; Riely, G.J.; Van Schil, P.E.; et al. International association for the study of lung cancer/american thoracic society/european respiratory society international multidisciplinary classification of lung adenocarcinoma. J. Thorac. Oncol. 2011, 6, 244–285. [Google Scholar] [CrossRef] [PubMed]
- Warth, A.; Muley, T.; Meister, M.; Stenzinger, A.; Thomas, M.; Schirmacher, P.; Schnabel, P.A.; Budczies, J.; Hoffmann, H.; Weichert, W. The novel histologic international association for the study of lung cancer/american thoracic society/european respiratory society classification system of lung adenocarcinoma is a stage-independent predictor of survival. J. Clin. Oncol. 2012, 30, 1438–1446. [Google Scholar] [CrossRef] [PubMed]
- Yoshizawa, A.; Motoi, N.; Riely, G.J.; Sima, C.S.; Gerald, W.L.; Kris, M.G.; Park, B.J.; Rusch, V.W.; Travis, W.D. Impact of proposed iaslc/ats/ers classification of lung adenocarcinoma: Prognostic subgroups and implications for further revision of staging based on analysis of 514 stage i cases. Mod. Pathol. 2011, 24, 653–664. [Google Scholar] [CrossRef] [PubMed]
- Hata, A.; Katakami, N.; Fujita, S.; Kaji, R.; Imai, Y.; Takahashi, Y.; Nishimura, T.; Tomii, K.; Ishihara, K. Frequency of EGFR and KRAS mutations in japanese patients with lung adenocarcinoma with features of the mucinous subtype of bronchioloalveolar carcinoma. J. Thorac. Oncol. 2010, 5, 1197–1200. [Google Scholar] [CrossRef] [PubMed]
- Casali, C.; Rossi, G.; Marchioni, A.; Sartori, G.; Maselli, F.; Longo, L.; Tallarico, E.; Morandi, U. A single institution-based retrospective study of surgically treated bronchioloalveolar adenocarcinoma of the lung: Clinicopathologic analysis, molecular features, and possible pitfalls in routine practice. J. Thorac. Oncol. 2010, 5, 830–836. [Google Scholar] [CrossRef] [PubMed]
- Finberg, K.E.; Sequist, L.V.; Joshi, V.A.; Muzikansky, A.; Miller, J.M.; Han, M.; Beheshti, J.; Chirieac, L.R.; Mark, E.J.; Iafrate, A.J. Mucinous differentiation correlates with absence of EGFR mutation and presence of KRAS mutation in lung adenocarcinomas with bronchioloalveolar features. J. Mol. Diagn. 2007, 9, 320–326. [Google Scholar] [CrossRef] [PubMed]
- Kadota, K.; Yeh, Y.C.; D’Angelo, S.P.; Moreira, A.L.; Kuk, D.; Sima, C.S.; Riely, G.J.; Arcila, M.E.; Kris, M.G.; Rusch, V.W.; et al. Associations between mutations and histologic patterns of mucin in lung adenocarcinoma: Invasive mucinous pattern and extracellular mucin are associated with KRAS mutation. Am. J. Surg. Pathol. 2014, 38, 1118–1127. [Google Scholar] [CrossRef] [PubMed]
- Shim, H.S.; Kenudson, M.; Zheng, Z.; Liebers, M.; Cha, Y.J.; Hoang Ho, Q.; Onozato, M.; Phi Le, L.; Heist, R.S.; Iafrate, A.J. Unique genetic and survival characteristics of invasive mucinous adenocarcinoma of the lung. J. Thorac. Oncol. 2015, 10, 1156–1162. [Google Scholar] [CrossRef] [PubMed]
- Cox, A.D.; Fesik, S.W.; Kimmelman, A.C.; Luo, J.; Der, C.J. Drugging the undruggable ras: Mission possible? Nat. Rev. Drug Discov. 2014, 13, 828–851. [Google Scholar] [CrossRef] [PubMed]
- Stephen, A.G.; Esposito, D.; Bagni, R.K.; McCormick, F. Dragging ras back in the ring. Cancer Cell 2014, 25, 272–281. [Google Scholar] [CrossRef] [PubMed]
- Manchado, E.; Weissmueller, S.; Morris, J.P.T.; Chen, C.C.; Wullenkord, R.; Lujambio, A.; de Stanchina, E.; Poirier, J.T.; Gainor, J.F.; Corcoran, R.B.; et al. A combinatorial strategy for treating KRAS-mutant lung cancer. Nature 2016, 534, 647–651. [Google Scholar] [CrossRef] [PubMed]
- Kitai, H.; Ebi, H.; Tomida, S.; Floros, K.V.; Kotani, H.; Adachi, Y.; Oizumi, S.; Nishimura, M.; Faber, A.C.; Yano, S. Epithelial-to-mesenchymal transition defines feedback activation of receptor tyrosine kinase signaling induced by mek inhibition in KRAS-mutant lung cancer. Cancer Discov. 2016, 6, 754–769. [Google Scholar] [CrossRef] [PubMed]
- Ordonez, N.G. Thyroid transcription factor-1 is a marker of lung and thyroid carcinomas. Adv. Anat. Pathol. 2000, 7, 123–127. [Google Scholar] [CrossRef] [PubMed]
- Cha, Y.J.; Shim, H.S. Biology of invasive mucinous adenocarcinoma of the lung. Transl. Lung Cancer Res. 2017, 6, 508–512. [Google Scholar] [CrossRef] [PubMed]
- Hwang, D.H.; Sholl, L.M.; Rojas-Rudilla, V.; Hall, D.L.; Shivdasani, P.; Garcia, E.P.; MacConaill, L.E.; Vivero, M.; Hornick, J.L.; Kuo, F.C.; et al. KRAS and NKX2-1 mutations in invasive mucinous adenocarcinoma of the lung. J. Thorac. Oncol. 2016, 11, 496–503. [Google Scholar] [CrossRef] [PubMed]
- Winslow, M.M.; Dayton, T.L.; Verhaak, R.G.; Kim-Kiselak, C.; Snyder, E.L.; Feldser, D.M.; Hubbard, D.D.; DuPage, M.J.; Whittaker, C.A.; Hoersch, S.; et al. Suppression of lung adenocarcinoma progression by NKX2-1. Nature 2011, 473, 101–104. [Google Scholar] [CrossRef] [PubMed]
- Maeda, Y.; Tsuchiya, T.; Hao, H.; Tompkins, D.H.; Xu, Y.; Mucenski, M.L.; Du, L.; Keiser, A.R.; Fukazawa, T.; Naomoto, Y.; et al. KRAS(g12d) and NKX2-1 haploinsufficiency induce mucinous adenocarcinoma of the lung. J. Clin. Invest. 2012, 122, 4388–4400. [Google Scholar] [CrossRef] [PubMed]
- Snyder, E.L.; Watanabe, H.; Magendantz, M.; Hoersch, S.; Chen, T.A.; Wang, D.G.; Crowley, D.; Whittaker, C.A.; Meyerson, M.; Kimura, S.; et al. NKX2-1 represses a latent gastric differentiation program in lung adenocarcinoma. Mol. Cell 2013, 50, 185–199. [Google Scholar] [CrossRef] [PubMed]
- Guo, M.; Tomoshige, K.; Meister, M.; Muley, T.; Fukazawa, T.; Tsuchiya, T.; Karns, R.; Warth, A.; Fink-Baldauf, I.M.; Nagayasu, T.; et al. Gene signature driving invasive mucinous adenocarcinoma of the lung. EMBO Mol. Med. 2017, 9, 462–481. [Google Scholar] [CrossRef] [PubMed]
- Pardoll, D.M. The blockade of immune checkpoints in cancer immunotherapy. Nat. Rev. Cancer 2012, 12, 252–264. [Google Scholar] [CrossRef] [PubMed]
- Keir, M.E.; Butte, M.J.; Freeman, G.J.; Sharpe, A.H. Pd-1 and its ligands in tolerance and immunity. Annu. Rev. Immunol. 2008, 26, 677–704. [Google Scholar] [CrossRef] [PubMed]
- Brahmer, J.R.; Tykodi, S.S.; Chow, L.Q.; Hwu, W.J.; Topalian, S.L.; Hwu, P.; Drake, C.G.; Camacho, L.H.; Kauh, J.; Odunsi, K.; et al. Safety and activity of anti-pd-l1 antibody in patients with advanced cancer. N. Engl. J. Med. 2012, 366, 2455–2465. [Google Scholar] [CrossRef] [PubMed]
- Topalian, S.L.; Sznol, M.; McDermott, D.F.; Kluger, H.M.; Carvajal, R.D.; Sharfman, W.H.; Brahmer, J.R.; Lawrence, D.P.; Atkins, M.B.; Powderly, J.D.; et al. Survival, durable tumor remission, and long-term safety in patients with advanced melanoma receiving nivolumab. J. Clin. Oncol. 2014, 32, 1020–1030. [Google Scholar] [CrossRef] [PubMed]
- Ni, L.; Dong, C. New b7 family checkpoints in human cancers. Mol. Cancer Ther. 2017, 16, 1203–1211. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Wang, J.; Zhou, J.; Zhan, S.; Huang, Y.; Wang, F.; Zhang, Z.; Zhu, D.; Zhao, H.; Li, D.; et al. B7-h3 combats apoptosis induced by chemotherapy by delivering signals to pancreatic cancer cells. Oncotarget 2017, 8, 74856–74868. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Yang, X.; Wu, Y.; Zhao, K.; Ye, Z.; Zhu, J.; Xu, X.; Zhao, X.; Xing, C. B7-h3 promotes gastric cancer cell migration and invasion. Oncotarget 2017, 8, 71725–71735. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.; Wang, X.; Mo, N.; Zhang, L.; Yuan, X.; Lu, Z. B7-homolog 4 promotes epithelial-mesenchymal transition and invasion of bladder cancer cells via activation of nuclear factor-kappab. Oncol. Res. 2018, 26, 1267–1274. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Liu, L.; Han, S.; Li, Y.; Qian, Q.; Zhang, Q.; Zhang, H.; Yang, Z.; Zhang, Y. B7-h3 is related to tumor progression in ovarian cancer. Oncol. Rep. 2017, 38, 2426–2434. [Google Scholar] [CrossRef] [PubMed]
- Zhou, D.; Zhou, Y.; Li, C.; Yang, L. Silencing of b7-h4 suppresses the tumorigenicity of the mgc-803 human gastric cancer cell line and promotes cell apoptosis via the mitochondrial signaling pathway. Int. J. Oncol. 2018, 52, 1267–1276. [Google Scholar] [CrossRef] [PubMed]
- Castellanos, J.R.; Purvis, I.J.; Labak, C.M.; Guda, M.R.; Tsung, A.J.; Velpula, K.K.; Asuthkar, S. B7-h3 role in the immune landscape of cancer. Am. J. Clin. Exp. Immunol. 2017, 6, 66–75. [Google Scholar] [PubMed]
- Flem-Karlsen, K.; Fodstad, O.; Tan, M.; Nunes-Xavier, C.E. B7-h3 in cancer—Beyond immune regulation. Trends Cancer 2018, 4, 401–404. [Google Scholar] [CrossRef] [PubMed]
- Loo, D.; Alderson, R.F.; Chen, F.Z.; Huang, L.; Zhang, W.; Gorlatov, S.; Burke, S.; Ciccarone, V.; Li, H.; Yang, Y.; et al. Development of an fc-enhanced anti-b7-h3 monoclonal antibody with potent antitumor activity. Clin. Cancer Res. 2012, 18, 3834–3845. [Google Scholar] [CrossRef] [PubMed]
- Inamura, K.; Yokouchi, Y.; Kobayashi, M.; Sakakibara, R.; Ninomiya, H.; Subat, S.; Nagano, H.; Nomura, K.; Okumura, S.; Shibutani, T.; et al. Tumor b7-h3 (cd276) expression and smoking history in relation to lung adenocarcinoma prognosis. Lung Cancer 2017, 103, 44–51. [Google Scholar] [CrossRef] [PubMed]
- Proctor, D.T.; Patel, Z.; Lama, S.; Resch, L.; van Marle, G.; Sutherland, G.R.J.O. Identification of pd-l2, b7–h3 and ctla-4 immune checkpoint proteins in genetic subtypes of meningioma. OncoImmunology 2018, 1–12. [Google Scholar] [CrossRef]
- Zou, W.; Chen, L. Inhibitory b7-family molecules in the tumour microenvironment. Nat. Rev. Immunol. 2008, 8, 467–477. [Google Scholar] [CrossRef] [PubMed]
- Chapoval, A.I.; Ni, J.; Lau, J.S.; Wilcox, R.A.; Flies, D.B.; Liu, D.; Dong, H.; Sica, G.L.; Zhu, G.; Tamada, K.; et al. B7-h3: A costimulatory molecule for t cell activation and ifn-gamma production. Nat. Immunol. 2001, 2, 269–274. [Google Scholar] [CrossRef] [PubMed]
- Hofmeyer, K.A.; Ray, A.; Zang, X. The contrasting role of b7-h3. Proc. Natl. Acad. Sci. USA 2008, 105, 10277–10278. [Google Scholar] [CrossRef] [PubMed]
- Sun, M.; Richards, S.; Prasad, D.V.; Mai, X.M.; Rudensky, A.; Dong, C. Characterization of mouse and human b7-h3 genes. J. Immunol. 2002, 168, 6294–6297. [Google Scholar] [CrossRef] [PubMed]
- Yi, K.H.; Chen, L. Fine tuning the immune response through b7-h3 and b7-h4. Immunol. Rev. 2009, 229, 145–151. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Wang, Y.; Zhao, J.; Gu, M.; Giscombe, R.; Lefvert, A.K.; Wang, X. B7-h3 and b7-h4 expression in non-small-cell lung cancer. Lung Cancer 2006, 53, 143–151. [Google Scholar] [CrossRef] [PubMed]
- Sun, T.W.; Gao, Q.; Qiu, S.J.; Zhou, J.; Wang, X.Y.; Yi, Y.; Shi, J.Y.; Xu, Y.F.; Shi, Y.H.; Song, K.; et al. B7-h3 is expressed in human hepatocellular carcinoma and is associated with tumor aggressiveness and postoperative recurrence. Cancer Immunol. Immunother. 2012, 61, 2171–2182. [Google Scholar] [CrossRef] [PubMed]
- Yamato, I.; Sho, M.; Nomi, T.; Akahori, T.; Shimada, K.; Hotta, K.; Kanehiro, H.; Konishi, N.; Yagita, H.; Nakajima, Y. Clinical importance of b7-h3 expression in human pancreatic cancer. Br. J. Cancer 2009, 101, 1709–1716. [Google Scholar] [CrossRef] [PubMed]
- Gibbs, A.R.; Thunnissen, F.B. Histological typing of lung and pleural tumours: Third edition. J. Clin. Pathol. 2001, 54, 498–499. [Google Scholar] [CrossRef] [PubMed]
- Chansky, K.; Detterbeck, F.C.; Nicholson, A.G.; Rusch, V.W.; Vallieres, E.; Groome, P.; Kennedy, C.; KRASnik, M.; Peake, M.; Shemanski, L.; et al. The iaslc lung cancer staging project: External validation of the revision of the tnm stage groupings in the eighth edition of the tnm classification of lung cancer. J. Thorac. Oncol. 2017, 12, 1109–1121. [Google Scholar] [CrossRef] [PubMed]
- The Cancer Genome Atlas Research Network. Comprehensive molecular profiling of lung adenocarcinoma. Nature 2014, 511, 543–550. [Google Scholar] [CrossRef] [PubMed]
- The Cancer Genome Atlas Research Network. Comprehensive genomic characterization of squamous cell lung cancers. Nature 2012, 489, 519–525. [Google Scholar] [CrossRef] [PubMed]
- Clinical Lung Cancer Genome Project; Network Genomic Medicine. A genomics-based classification of human lung tumors. Sci. Transl. Med. 2013, 5, 209ra153. [Google Scholar]
- Rudin, C.M.; Durinck, S.; Stawiski, E.W.; Poirier, J.T.; Modrusan, Z.; Shames, D.S.; Bergbower, E.A.; Guan, Y.; Shin, J.; Guillory, J.; et al. Comprehensive genomic analysis identifies sox2 as a frequently amplified gene in small-cell lung cancer. Nat. Genet. 2012, 44, 1111–1116. [Google Scholar] [CrossRef] [PubMed]
- Peifer, M.; Fernandez-Cuesta, L.; Sos, M.L.; George, J.; Seidel, D.; Kasper, L.H.; Plenker, D.; Leenders, F.; Sun, R.; Zander, T.; et al. Integrative genome analyses identify key somatic driver mutations of small-cell lung cancer. Nat. Genet. 2012, 44, 1104–1110. [Google Scholar] [CrossRef] [PubMed]
- Imielinski, M.; Berger, A.H.; Hammerman, P.S.; Hernandez, B.; Pugh, T.J.; Hodis, E.; Cho, J.; Suh, J.; Capelletti, M.; Sivachenko, A.; et al. Mapping the hallmarks of lung adenocarcinoma with massively parallel sequencing. Cell 2012, 150, 1107–1120. [Google Scholar] [CrossRef] [PubMed]
- Govindan, R.; Ding, L.; Griffith, M.; Subramanian, J.; Dees, N.D.; Kanchi, K.L.; Maher, C.A.; Fulton, R.; Fulton, L.; Wallis, J.; et al. Genomic landscape of non-small cell lung cancer in smokers and never-smokers. Cell 2012, 150, 1121–1134. [Google Scholar] [CrossRef] [PubMed]
- Catalogue of Somatic Mutations In Cancer. Available online: http://cancer.sanger.ac.uk/cancergenome/projects/cosmic (accessed on 4 May 2014).
- Hirotsu, Y.; Nakagomi, H.; Sakamoto, I.; Amemiya, K.; Oyama, T.; Mochizuki, H.; Omata, M. Multigene panel analysis identified germline mutations of DNA repair genes in breast and ovarian cancer. Mol. Genet. Genomic Med. 2015, 3, 459–466. [Google Scholar] [CrossRef] [PubMed]
- Hirotsu, Y.; Nakagomi, H.; Sakamoto, I.; Amemiya, K.; Mochizuki, H.; Omata, M. Detection of brca1 and brca2 germline mutations in japanese population using next-generation sequencing. Mol. Genet. Genomic Med. 2015, 3, 121–129. [Google Scholar] [CrossRef] [PubMed]
- Goto, T.; Hirotsu, Y.; Amemiya, K.; Nakagomi, T.; Shikata, D.; Yokoyama, Y.; Okimoto, K.; Oyama, T.; Mochizuki, H.; Omata, M. Distribution of circulating tumor DNA in lung cancer: Analysis of the primary lung and bone marrow along with the pulmonary venous and peripheral blood. Oncotarget 2017, 8, 59268–59281. [Google Scholar] [CrossRef] [PubMed]
- Goto, T.; Hirotsu, Y.; Oyama, T.; Amemiya, K.; Omata, M. Analysis of tumor-derived DNA in plasma and bone marrow fluid in lung cancer patients. Med. Oncol. 2016, 33, 29. [Google Scholar] [CrossRef] [PubMed]
- Goto, T.; Hirotsu, Y.; Nakagomi, T.; Shikata, D.; Yokoyama, Y.; Amemiya, K.; Tsutsui, T.; Kakizaki, Y.; Oyama, T.; Mochizuki, H.; et al. Detection of tumor-derived DNA dispersed in the airway improves the diagnostic accuracy of bronchoscopy for lung cancer. Oncotarget 2017, 8, 79404–79413. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Amemiya, K.; Hirotsu, Y.; Goto, T.; Nakagomi, H.; Mochizuki, H.; Oyama, T.; Omata, M. Touch imprint cytology with massively parallel sequencing (tic-seq): A simple and rapid method to snapshot genetic alterations in tumors. Cancer Med. 2016, 5, 3426–3436. [Google Scholar] [CrossRef] [PubMed]
- Iijima, Y.; Hirotsu, Y.; Amemiya, K.; Ooka, Y.; Mochizuki, H.; Oyama, T.; Nakagomi, T.; Uchida, Y.; Kobayashi, Y.; Tsutsui, T.; et al. Very early response of circulating tumour-derived DNA in plasma predicts efficacy of nivolumab treatment in patients with non-small cell lung cancer. Eur. J. Cancer 2017, 86, 349–357. [Google Scholar] [CrossRef] [PubMed]
- Goto, T.; Hirotsu, Y.; Mochizuki, H.; Nakagomi, T.; Oyama, T.; Amemiya, K.; Omata, M. Stepwise addition of genetic changes correlated with histological change from “well-differentiated” to “sarcomatoid” phenotypes: A case report. BMC Cancer 2017, 17, 65. [Google Scholar] [CrossRef] [PubMed]
- Nakagomi, T.; Goto, T.; Hirotsu, Y.; Shikata, D.; Yokoyama, Y.; Higuchi, R.; Amemiya, K.; Okimoto, K.; Oyama, T.; Mochizuki, H.; et al. New therapeutic targets for pulmonary sarcomatoid carcinomas based on their genomic and phylogenetic profiles. Oncotarget 2018, 9, 10635–10649. [Google Scholar] [CrossRef] [PubMed]
- Nakagomi, T.; Goto, T.; Hirotsu, Y.; Shikata, D.; Amemiya, K.; Oyama, T.; Mochizuki, H.; Omata, M. Elucidation of radiation-resistant clones by a serial study of intratumor heterogeneity before and after stereotactic radiotherapy in lung cancer. J. Thorac. Dis. 2017, 9, E598–E604. [Google Scholar] [CrossRef] [PubMed]
- Goto, T.; Hirotsu, Y.; Mochizuki, H.; Nakagomi, T.; Shikata, D.; Yokoyama, Y.; Oyama, T.; Amemiya, K.; Okimoto, K.; Omata, M. Mutational analysis of multiple lung cancers: Discrimination between primary and metastatic lung cancers by genomic profile. Oncotarget 2017, 8, 31133–31143. [Google Scholar] [CrossRef] [PubMed]
- Ahr, A.; Holtrich, U.; Solbach, C.; Scharl, A.; Strebhardt, K.; Karn, T.; Kaufmann, M. Molecular classification of breast cancer patients by gene expression profiling. J. Pathol. 2001, 195, 312–320. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristic | IMA (n = 20) | NMA (n = 43) | p Value |
|---|---|---|---|
| Age | 72.7 ± 7.9 | 67.9 ± 8.3 | 0.98 |
| Sex | |||
| Male | 12 (60%) | 27 (62.8%) | 0.83 |
| Female | 8 (40%) | 16 (37.2%) | |
| Performance Status | |||
| 0 | 17 (85%) | 36 (83.7%) | |
| 1 | 3 (15%) | 7 (16.3%) | 0.90 |
| ≧2 | 0 (0%) | 0 (0%) | |
| Smoking habit | |||
| Never | 10 (50%) | 15 (34.9%) | 0.26 |
| Former/current | 10 (50%) | 28 (65.1%) | |
| Smoking index | 383 ± 107 | 476 ± 86 | 0.26 |
| CT finding | |||
| Solid | 9 (45%) | 20 (46.5%) | |
| Part solid GGN | 11 (55%) | 8 (18.6%) | 0.0002 |
| GGN | 0 (0%) | 15 (34.9%) | |
| Tumor location | |||
| Central | 0 (0%) | 0 (0%) | |
| Middle | 7 (35%) | 8 (18.6%) | 0.16 |
| Peripheral | 13 (65%) | 35 (81.4%) | |
| Surgical procedure received | |||
| Sublobar resection | 1 (5%) | 10 (23.3%) | |
| Lobectomy | 18 (90%) | 30 (69.8%) | 0.15 |
| Pneumonectomy | 1 (5%) | 1 (2.3%) | |
| Other | 0 (0%) | 2 (4.6%) | |
| Tumor size (mm) | 37.2 ± 6.8 | 20.7 ± 1.4 | 1.00 |
| Pathological stage | |||
| I | 17 (85%) | 38 (88.4%) | |
| II | 3 (15%) | 1 (2.3%) | 0.10 |
| III | 0 (0%) | 3 (7.0%) | |
| IV | 0 (0%) | 1 (2.3%) | |
| Pathological lymphatic invasion | |||
| Absent | 18 (90%) | 37 (86.5%) | 0.66 |
| Present | 2 (10%) | 6 (13.5%) | |
| Pathological vessel invasion | |||
| Absent | 17 (85%) | 33 (76.7%) | |
| Microscopically present | 3 (15%) | 10 (23.3%) | 0.44 |
| Macroscopically present | 0 (0%) | 0 (0%) |
| Characteristic | Cluster A | Cluster B | Cluster C | p Value | |
|---|---|---|---|---|---|
| n = 14 | n = 23 | n = 15 | |||
| Sex | 0.003 | ||||
| male | 14 (100%) | 13 (56.5%) | 9 (60%) | ||
| female | 0 (0%) | 10 (43.5%) | 6 (40%) | ||
| Age | 70.4 ± 8.7 | 70.2 ± 7.9 | 70.6 ± 8.0 | 0.952 | |
| Smoking index | 0.008 | ||||
| 0 | 0 | 10 | 5 | ||
| 1–1000 | 7 | 10 | 6 | ||
| 1000< | 7 | 3 | 4 | ||
| Histology | 0.001 | ||||
| NMA | 6 | 20 | 4 | ||
| IMA | 0 | 2 | 8 | ||
| SCC | 4 | 1 | 1 | ||
| other | 4 | 0 | 2 | ||
| Pathological stage | 0.158 | ||||
| 0-IA | 4 | 15 | 8 | ||
| IB | 4 | 6 | 3 | ||
| IIA-IV | 6 | 2 | 4 | ||
| Variables | Hazard ratio (95% CI) | p Value |
|---|---|---|
| Cluster | ||
| cluster A | 1 (Ref.) | |
| cluster B | 0.25 (0.05–0.94) | 0.04 |
| cluster C | 0.18 (0.03–0.78) | 0.02 |
| Pathological stage | ||
| stage 0 or IA | 1 (Ref.) | |
| stage IB | 22.58 (1.80–881.50) | 0.01 |
| stage IIA or more | 36.09 (2.83–1972.69) | 0.003 |
| Male (ref. Female) | 1.25 (0.24–9.21) | 0.75 |
| Age | ||
| –65 | 1 (Ref.) | |
| 66–75 | 1.08 (0.34–7.36) | 0.71 |
| 76– | 1.22 (0.56–8.31) | 0.55 |
| Smoker (ref. non-smoker) | 1.56 (0.61–8.54) | 0.74 |
| Histology | ||
| NMA | 1 (Ref.) | |
| IMA | 0.46 (0.01–7.14) | 0.60 |
| SCC | 6.94 (0.34–163.65) | 0.20 |
| Others | 3.05 (0.05–295.07) | 0.59 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nakagomi, T.; Goto, T.; Hirotsu, Y.; Shikata, D.; Yokoyama, Y.; Higuchi, R.; Otake, S.; Amemiya, K.; Oyama, T.; Mochizuki, H.; et al. Genomic Characteristics of Invasive Mucinous Adenocarcinomas of the Lung and Potential Therapeutic Targets of B7-H3. Cancers 2018, 10, 478. https://doi.org/10.3390/cancers10120478
Nakagomi T, Goto T, Hirotsu Y, Shikata D, Yokoyama Y, Higuchi R, Otake S, Amemiya K, Oyama T, Mochizuki H, et al. Genomic Characteristics of Invasive Mucinous Adenocarcinomas of the Lung and Potential Therapeutic Targets of B7-H3. Cancers. 2018; 10(12):478. https://doi.org/10.3390/cancers10120478
Chicago/Turabian StyleNakagomi, Takahiro, Taichiro Goto, Yosuke Hirotsu, Daichi Shikata, Yujiro Yokoyama, Rumi Higuchi, Sotaro Otake, Kenji Amemiya, Toshio Oyama, Hitoshi Mochizuki, and et al. 2018. "Genomic Characteristics of Invasive Mucinous Adenocarcinomas of the Lung and Potential Therapeutic Targets of B7-H3" Cancers 10, no. 12: 478. https://doi.org/10.3390/cancers10120478
APA StyleNakagomi, T., Goto, T., Hirotsu, Y., Shikata, D., Yokoyama, Y., Higuchi, R., Otake, S., Amemiya, K., Oyama, T., Mochizuki, H., & Omata, M. (2018). Genomic Characteristics of Invasive Mucinous Adenocarcinomas of the Lung and Potential Therapeutic Targets of B7-H3. Cancers, 10(12), 478. https://doi.org/10.3390/cancers10120478