Uniform Orientation of Biotinylated Nanobody as an Affinity Binder for Detection of Bacillus thuringiensis (Bt) Cry1Ac Toxin

Abstract
:1. Introduction
2. Results and Discussion
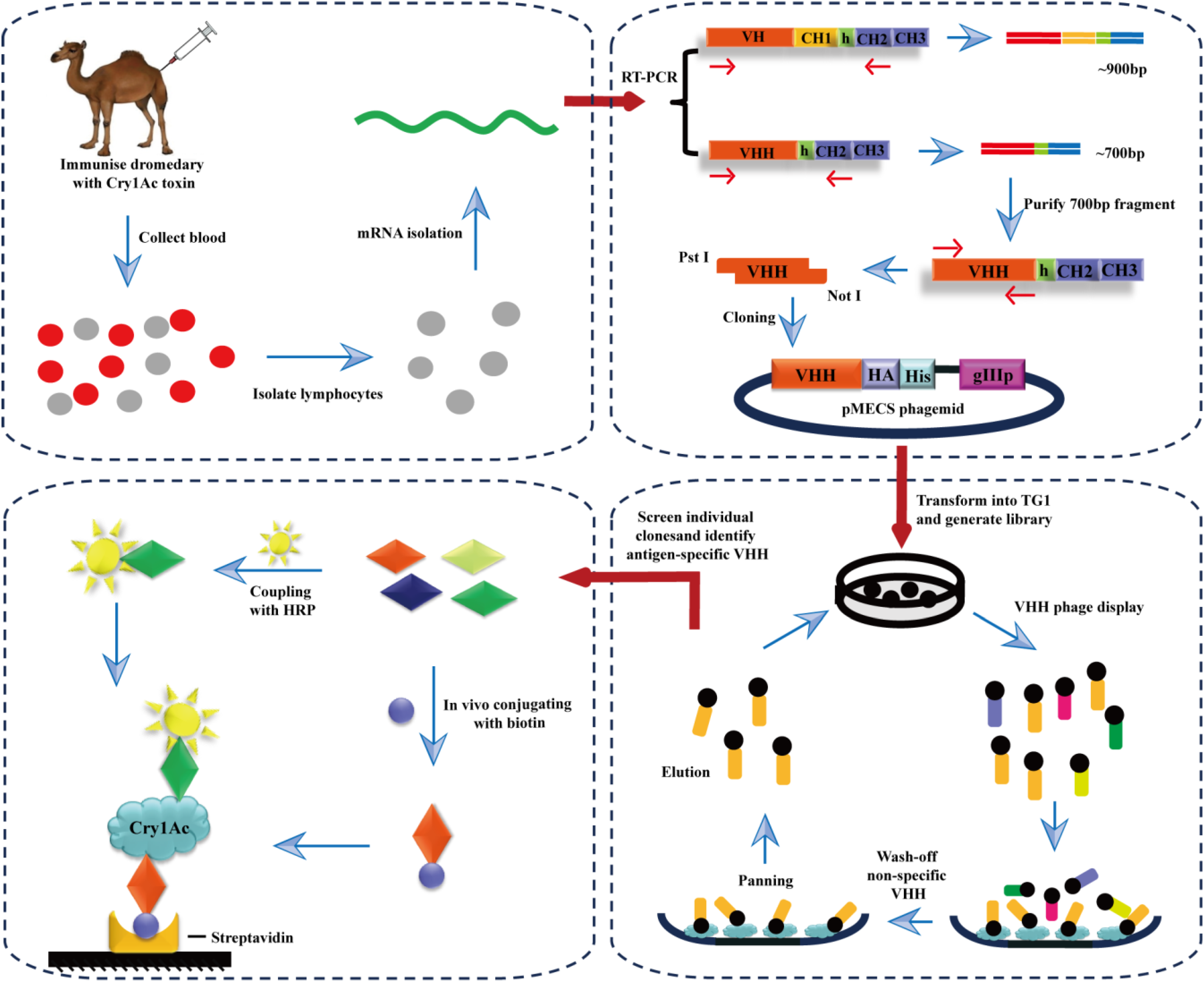
2.1. Immunized VHH Library Construction
2.2. Library Screening and Identification of Cry1Ac-Specific Nanobodies

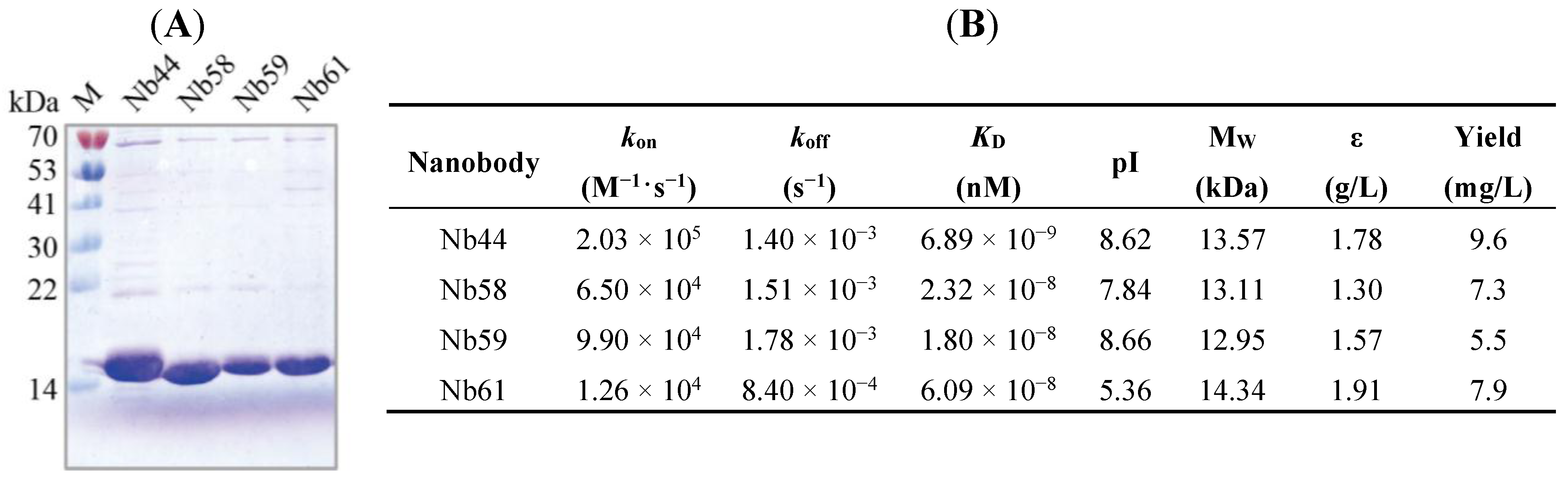
2.3. Nbs Expression, Purification and Affinity Determination

2.4. Nanobody Conjugation to HRP in Vitro and Biotinylation in Vivo
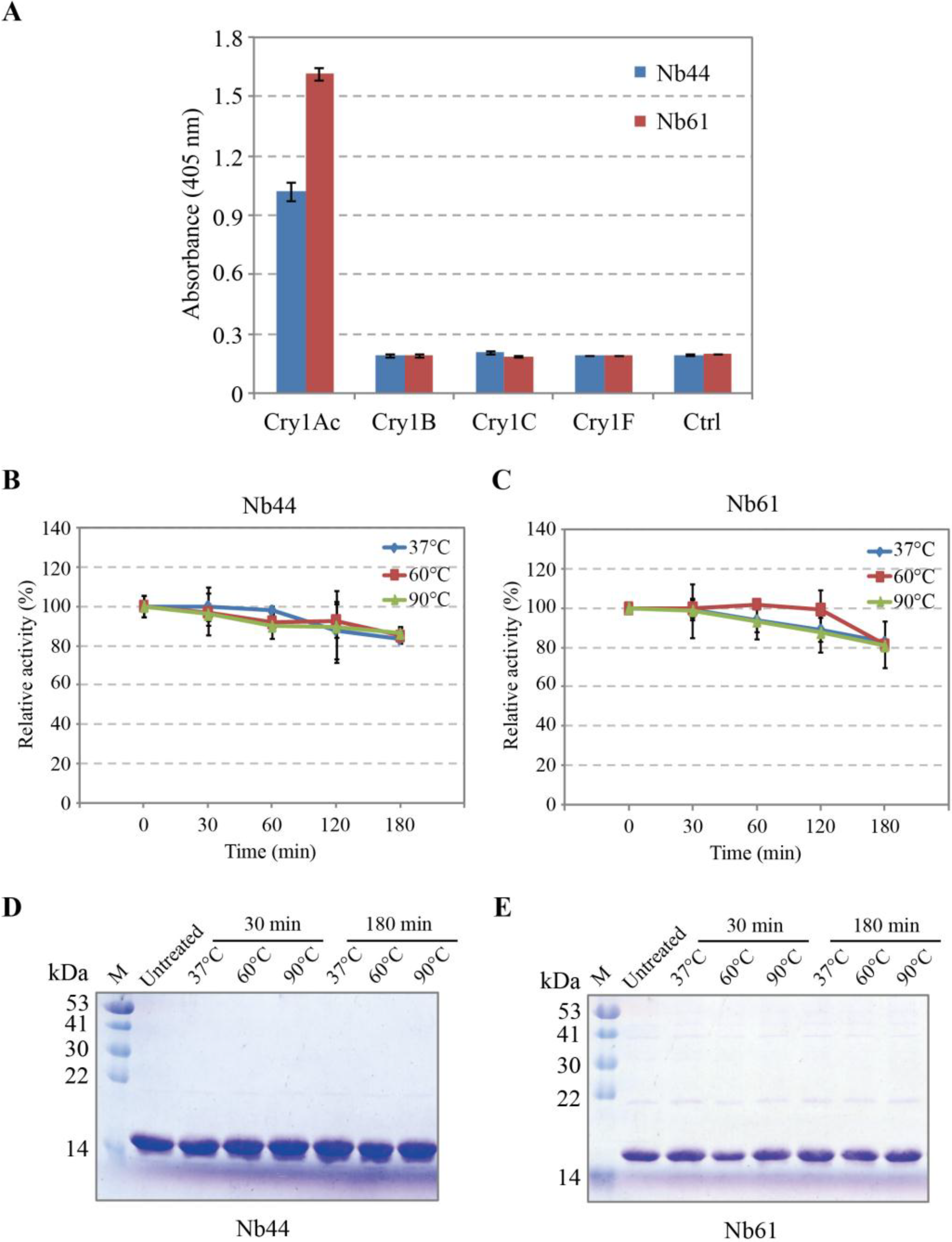
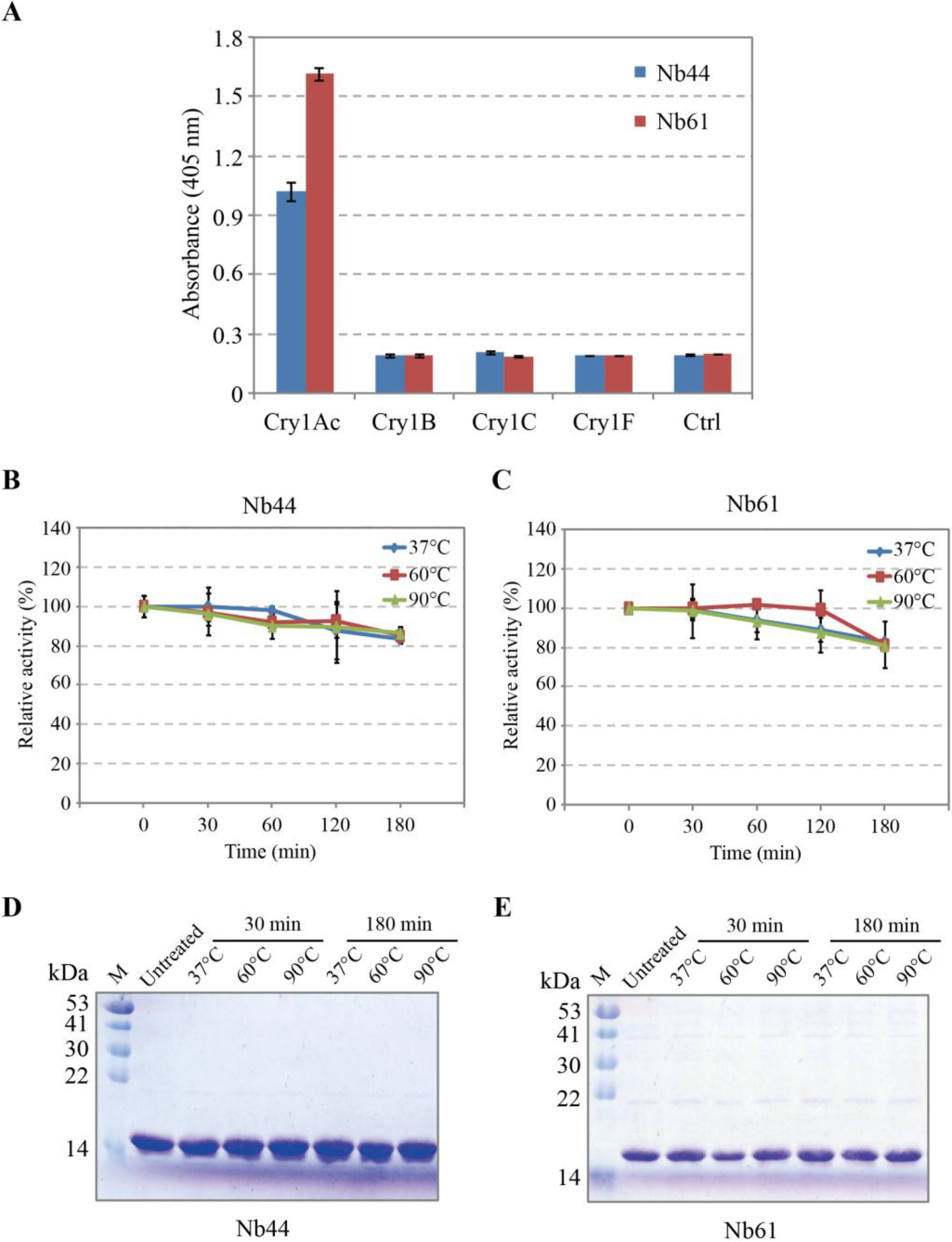
2.5. Specificity and Thermostability Analysis
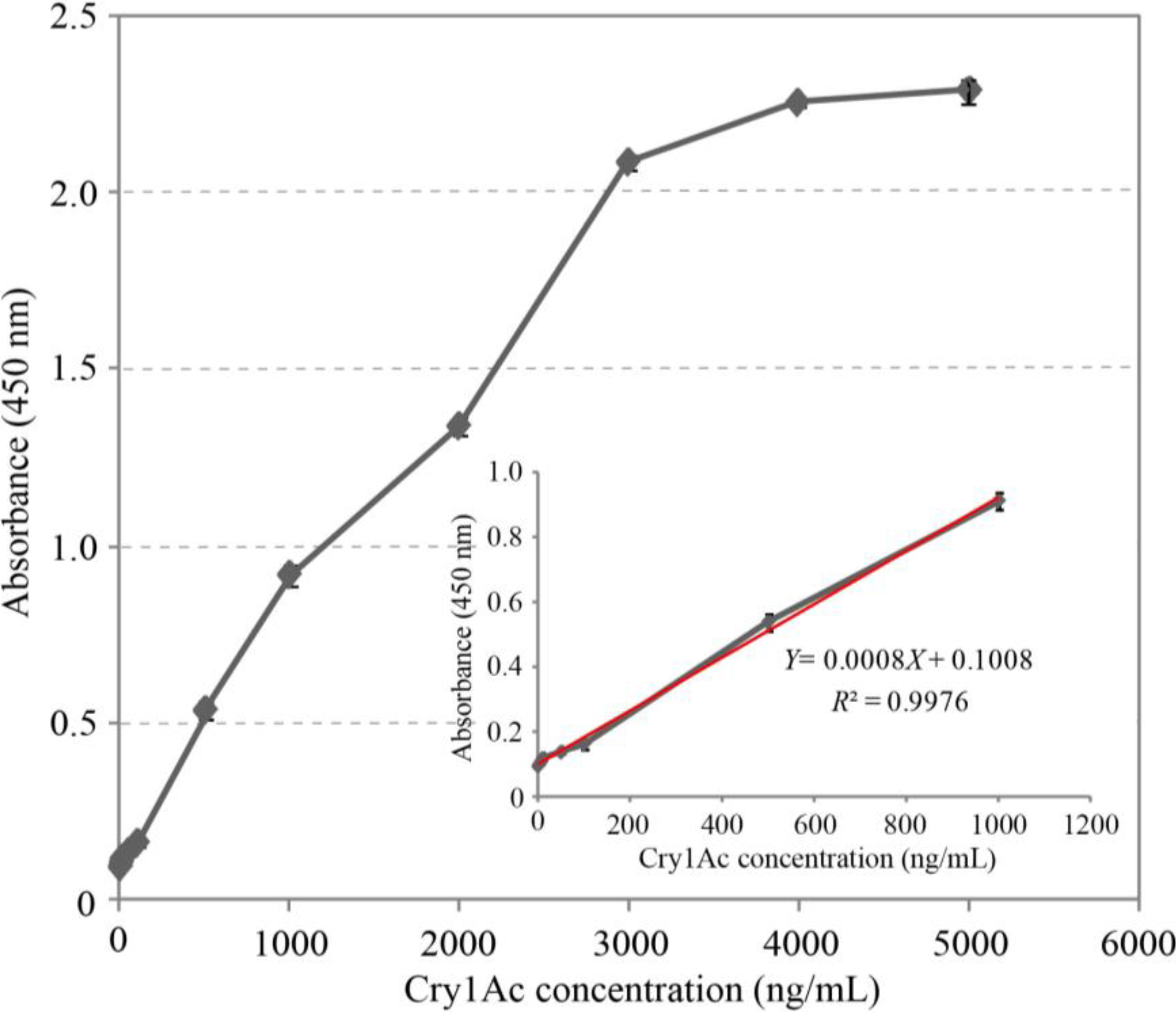
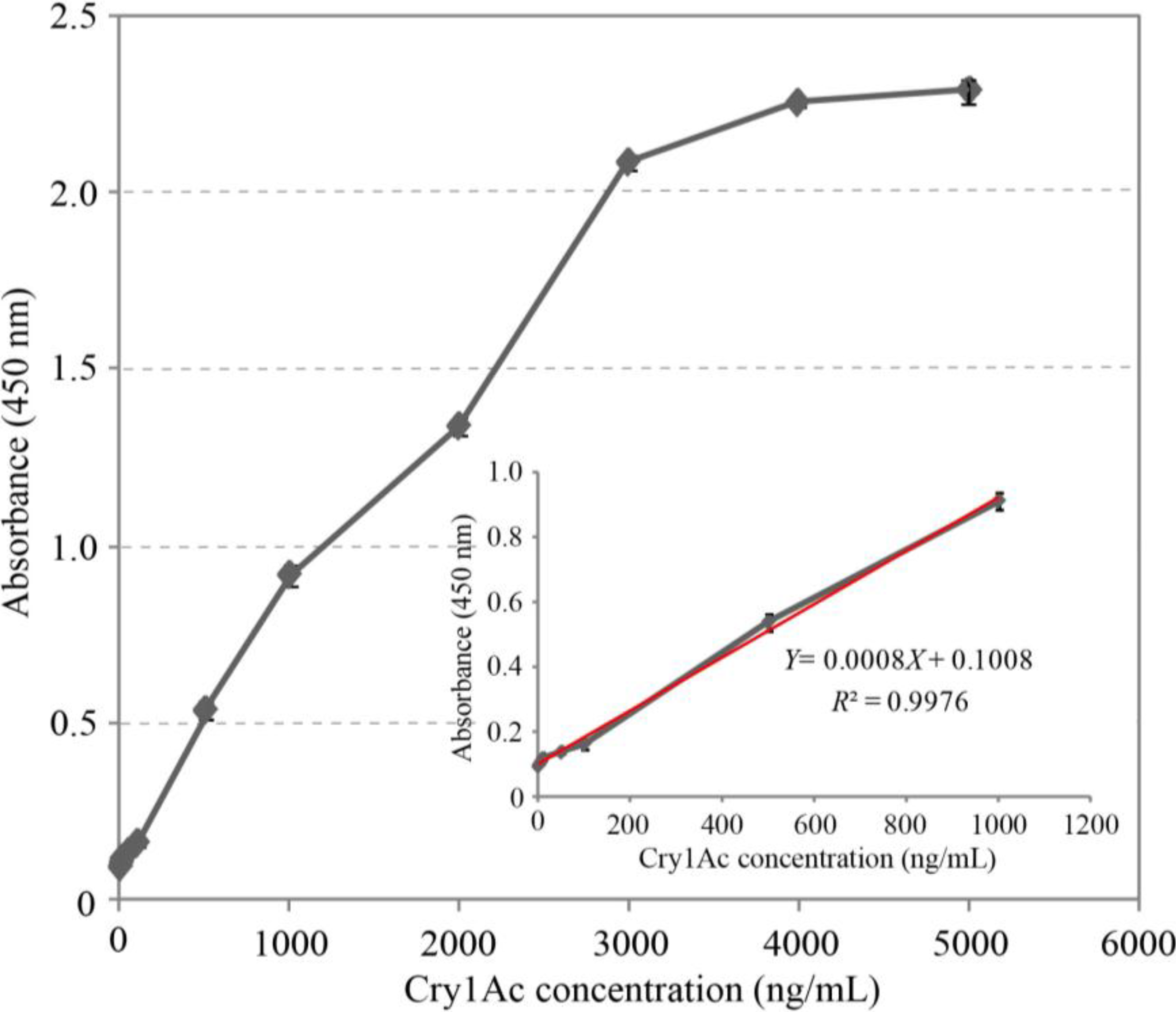
2.6. Cry1Ac Toxin Detection by DAS-ELISA

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Cry1Ac Concentration (ng/mL) | Recovery (%) | RSD (%) | ||
|---|---|---|---|---|---|
| Added | Found | ||||
| Corn | 1 | 50 | 40.66 | 81.32 | 4.32 |
| 2 | 100 | 82.68 | 82.68 | 7.09 | |
| 3 | 250 | 275.02 | 110.00 | 6.83 | |
| 4 | 500 | 424.80 | 86.96 | 4.70 | |
| 5 | 1000 | 1024.40 | 104.24 | 5.12 | |
| Soybean | 1 | 50 | 48.29 | 96.59 | 5.19 |
| 2 | 100 | 106.19 | 106.19 | 7.36 | |
| 3 | 250 | 272.18 | 107.21 | 6.35 | |
| 4 | 500 | 420.80 | 84.16 | 2.52 | |
| 5 | 1000 | 1029.60 | 102.96 | 3.28 | |
2.7. Assessment of the DAS-ELISA by Spiked Samples
3. Experimental Section
3.1. Chemicals and Materials
3.2. Dromedary Immunization
3.3. Library Construction
3.4. Selection of Cry1Ac-Specific Nanobodies from Phage-Display Library
3.5. Periplasmic Extract ELISA (PE-ELISA)
3.6. Expression and Purification of Soluble Nanobodies against Cry1Ac
3.7. Affinity Determination
3.8. HRP Coupling: Preparation of HRP-Conjugated Second Antibody for Detection
3.9. Nanobody Biotinylation: Preparation of Biotinylated First Antibody for Capture
3.10. ELISA for Nanobody Specificity Detection
3.11. Thermostability Analysis
3.12. Cry1Ac Toxin Detection by DAS-ELISA Assay Based on Nbs and Directional System
3.13. Assessment of the DAS-ELISA by Spiked Samples
4. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Clark, B.W.; Phillips, T.A.; Coats, J.R. Environmental fate and effects of Bacillus thuringiensis (Bt) proteins from transgenic crops: A review. J. Agric. Food Chem. 2005, 53, 4643–4653. [Google Scholar] [CrossRef] [PubMed]
- Aronson, A.I.; Shai, Y. Why Bacillus thuringiensis insecticidal toxins are so effective: Unique features of their mode of action. FEMS Microbiol. Lett. 2001, 195, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Soberon, M.; Gill, S.S.; Bravo, A. Signaling versus punching hole: How do Bacillus thuringiensis toxins kill insect midgut cells? Cellular and molecular life sciences. CMLS 2009, 66, 1337–1349. [Google Scholar] [CrossRef] [PubMed]
- Sanahuja, G.; Banakar, R.; Twyman, R.M.; Capell, T.; Christou, P. Bacillus thuringiensis: A century of research, development and commercial applications. Plant Biotechnol. J. 2011, 9, 283–300. [Google Scholar] [CrossRef] [PubMed]
- Tabashnik, B.E.; Fabrick, J.A.; Unnithan, G.C.; Yelich, A.J.; Masson, L.; Zhang, J.; Bravo, A.; Soberon, M. Efficacy of genetically modified Bt toxins alone and in combinations against pink bollworm resistant to Cry1Ac and Cry2Ab. PLoS One 2013, 8. [Google Scholar] [CrossRef]
- Tabashnik, B.E. Evolution of resistance to Bacillus-thuringiensis. Annu. Rev. Entomol. 1994, 39, 47–79. [Google Scholar] [CrossRef]
- Lemaux, P.G. Genetically engineered plants and foods: A scientist’s analysis of the issues (part I). Annu. Rev. Plant Biol. 2008, 59, 771–812. [Google Scholar] [CrossRef] [PubMed]
- Lemaux, P.G. Genetically engineered plants and foods: A scientist’s analysis of the issues (part II). Annu. Rev. Plant Biol. 2009, 60, 511–559. [Google Scholar] [CrossRef] [PubMed]
- James, C.; International Service for the Acquisition of Agri-Biotech Applications. Global Status of Commercialized Biotech/GM Crops, 2009; International Service for the Acquisition of Agri-Biotech Applications. ISAAA: Los Angeles, California, USA. Available online: http://report.agropages.com/ReportDetail-934.htm (accessed on 1 July 2014).
- Kuiper, H.A.; Kleter, G.A.; Noteborn, H.P.J.M.; Kok, E.J. Substantial equivalence—An appropriate paradigm for the safety assessment of genetically modified foods? Toxicology 2002, 181, 427–431. [Google Scholar] [CrossRef] [PubMed]
- Mezzomo, B.P.; Miranda-Vilela, A.L.; de Souza Freire, I.; Barbosa, L.C.; Portilho, F.A.; Lacava, Z.G.; Grisolia, C.K. Withdrawn: Effects of oral administration of Bacillus thuringiensis as spore-crystal strains Cry1Aa, Cry1Ab, Cry1Ac or Cry2Aa on hematologic and genotoxic endpoints of Swiss albino mice. Food Chem. Toxicol. 2012. [Google Scholar] [CrossRef]
- Kamle, S.; Ali, S. Genetically modified crops: Detection strategies and biosafety issues. Gene 2013, 522, 123–132. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.H. Screening DNA chip and event-specific multiplex PCR detection methods for biotech crops. J. Sci. Food Agric. 2014, 94, 2856–2862. [Google Scholar] [CrossRef] [PubMed]
- Fagan, J.; Schoel, B.; Haegert, A.; Moore, J.; Beeby, J. Performance assessment under field conditions of a rapid immunological test for transgenic soybeans. Int. J. Food Sci. Technol. 2001, 36, 357–367. [Google Scholar] [CrossRef]
- Asensio, L.; Gonzalez, I.; Garcia, T.; Martin, R. Determination of food authenticity by enzyme-linked immunosorbent assay (ELISA). Food Control 2008, 19, 1–8. [Google Scholar] [CrossRef]
- Morisset, D.; Demsar, T.; Gruden, K.; Vojvoda, J.; Stebih, D.; Zel, J. Detection of genetically modified organisms-closing the gaps. Nat. Biotechnol. 2009, 27, 700–701. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Xu, C.; Zhang, C.; Liu, Y.; Xie, Y.; Liu, X. Established a new double antibodies sandwich enzyme-linked immunosorbent assay for detecting Bacillus thuringiensis (Bt) Cry1Ab toxin based single-chain variable fragments from a naive mouse phage displayed library. Toxicon 2014, 81, 13–22. [Google Scholar] [CrossRef] [PubMed]
- Hassanzadeh-Ghassabeh, G.; Devoogdt, N.; de Pauw, P.; Vincke, C.; Muyldermans, S. Nanobodies and their potential applications. Nanomedicine (U.K.) 2013, 8, 1013–1026. [Google Scholar] [CrossRef]
- Araste, F.; Ebrahimizadeh, W.; Rasooli, I.; Rajabibazl, M.; Mousavi Gargari, S.L. A novel VHH nanobody against the active site (the ca domain) of tumor-associated, carbonic anhydrase isoform IX and its usefulness for cancer diagnosis. Biotechnol. Lett. 2014, 36, 21–28. [Google Scholar] [CrossRef] [PubMed]
- De Meyer, T.; Muyldermans, S.; Depicker, A. Nanobody-based products as research and diagnostic tools. Trends Biotechnol. 2014, 32, 263–270. [Google Scholar]
- Roovers, R.C.; van Dongen, G.A.; van Bergen en Henegouwen, P.M. Nanobodies in therapeutic applications. Curr. Opin. Mol. Ther. 2007, 9, 327–335. [Google Scholar] [PubMed]
- Ebrahimizadeh, W.; Gargari, S.M.; Rajabibazl, M.; Ardekani, L.S.; Zare, H.; Bakherad, H. Isolation and characterization of protective anti-LPS nanobody against v. Cholerae O1 recognizing Inaba and Ogawa serotypes. Appl. Microbiol. Biot. 2013, 97, 4457–4466. [Google Scholar] [CrossRef]
- Stijlemans, B.; Conrath, K.; Cortez-Retamozo, V.; van Xong, H.; Wyns, L.; Senter, P.; Revets, H.; de Baetselier, P.; Muyldermans, S.; Magez, S. Efficient targeting of conserved cryptic epitopes of infectious agents by single domain antibodies. African trypanosomes as paradigm. J. Biol. Chem. 2004, 279, 1256–1261. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Sun, Y.; Elseviers, J.; Muyldermans, S.; Liu, S.; Wan, Y. A nanobody-based electrochemiluminescent immunosensor for sensitive detection of human procalcitonin. Analyst 2014, 139, 3718–3721. [Google Scholar] [CrossRef] [PubMed]
- Ma, L.; Sun, Y.; Kang, X.; Wan, Y. Development of nanobody-based flow injection chemiluminescence immunoassay for sensitive detection of human prealbumin. Biosens. Bioelectr. 2014, 61, 165–171. [Google Scholar] [CrossRef]
- Tang, J.C.; Szikra, T.; Kozorovitskiy, Y.; Teixiera, M.; Sabatini, B.L.; Roska, B.; Cepko, C.L. A nanobody-based system using fluorescent proteins as scaffolds for cell-specific gene manipulation. Cell 2013, 154, 928–939. [Google Scholar] [CrossRef] [PubMed]
- Ditlev, S.B.; Florea, R.; Nielsen, M.A.; Theander, T.G.; Magez, S.; Boeuf, P.; Salanti, A. Utilizing nanobody technology to target non-immunodominant domains of VAR2CSA. PLoS One 2014, 9. [Google Scholar] [CrossRef]
- Muyldermans, S.; Baral, T.N.; Retamozzo, V.C.; de Baetselier, P.; de Genst, E.; Kinne, J.; Leonhardt, H.; Magez, S.; Nguyen, V.K.; Revets, H.; et al. Camelid immunoglobulins and nanobody technology. Vet. Immunol. Immunopathol. 2009, 128, 178–183. [Google Scholar] [CrossRef]
- Armbruster, D.A.; Pry, T. Limit of blank, limit of detection and limit of quantitation. Clin. Biochem. Rev. Aust. Assoc. Clin. Biochem. 2008, 29 (Suppl. 1), S49–S52. [Google Scholar]
- Vincke, C.; Gutierrez, C.; Wernery, U.; Devoogdt, N.; Hassanzadeh-Ghassabeh, G.; Muyldermans, S. Generation of single domain antibody fragments derived from camelids and generation of manifold constructs. Methods Mol. Biol. 2012, 907, 145–176. [Google Scholar] [PubMed]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, M.; Zhu, M.; Zhang, C.; Liu, X.; Wan, Y. Uniform Orientation of Biotinylated Nanobody as an Affinity Binder for Detection of Bacillus thuringiensis (Bt) Cry1Ac Toxin. Toxins 2014, 6, 3208-3222. https://doi.org/10.3390/toxins6123208
Li M, Zhu M, Zhang C, Liu X, Wan Y. Uniform Orientation of Biotinylated Nanobody as an Affinity Binder for Detection of Bacillus thuringiensis (Bt) Cry1Ac Toxin. Toxins. 2014; 6(12):3208-3222. https://doi.org/10.3390/toxins6123208
Chicago/Turabian StyleLi, Min, Min Zhu, Cunzheng Zhang, Xianjin Liu, and Yakun Wan. 2014. "Uniform Orientation of Biotinylated Nanobody as an Affinity Binder for Detection of Bacillus thuringiensis (Bt) Cry1Ac Toxin" Toxins 6, no. 12: 3208-3222. https://doi.org/10.3390/toxins6123208