Development and Evaluation of Monoclonal Antibodies for the Glucoside of T-2 Toxin (T2-Glc)




Abstract
:1. Introduction
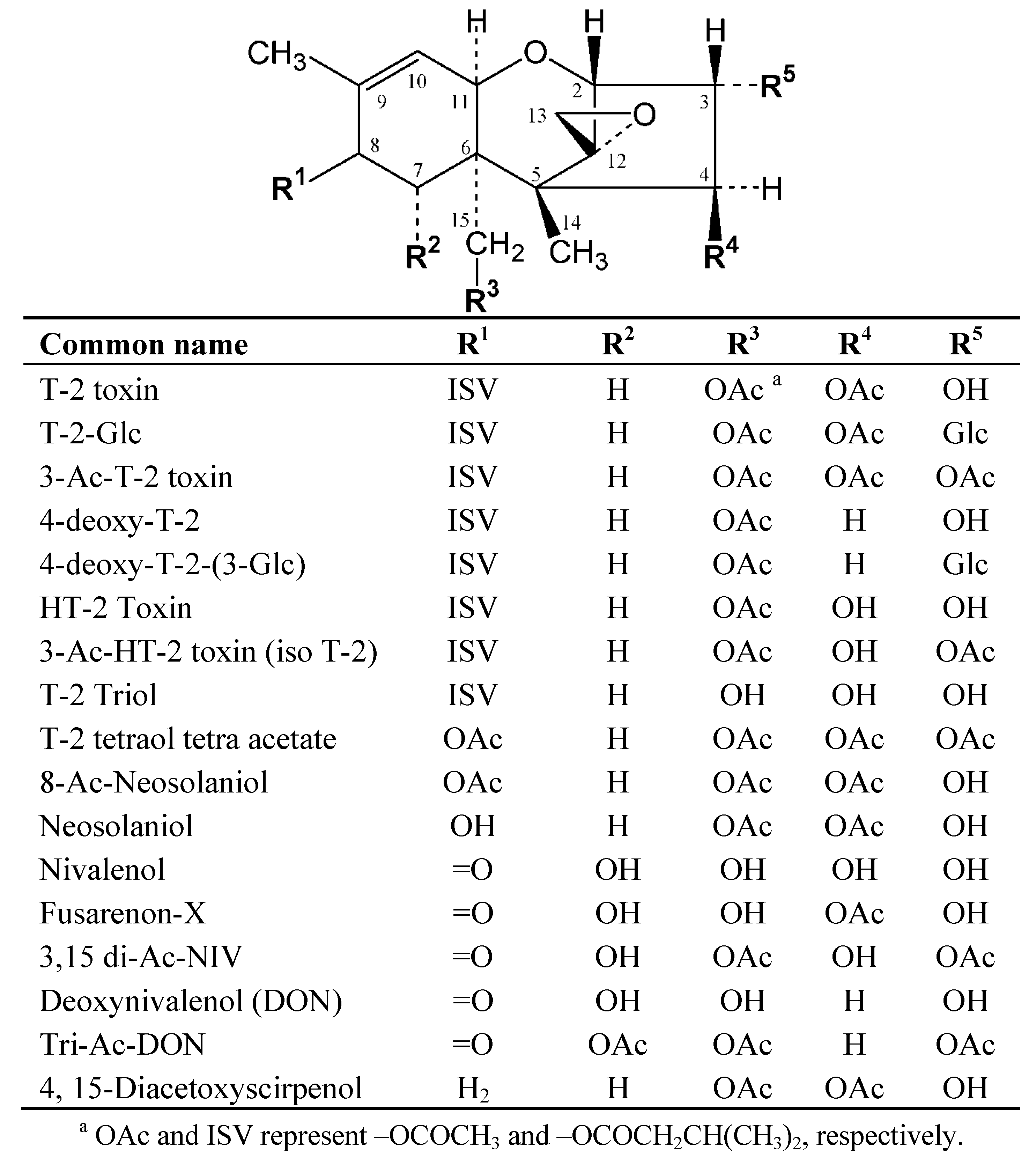
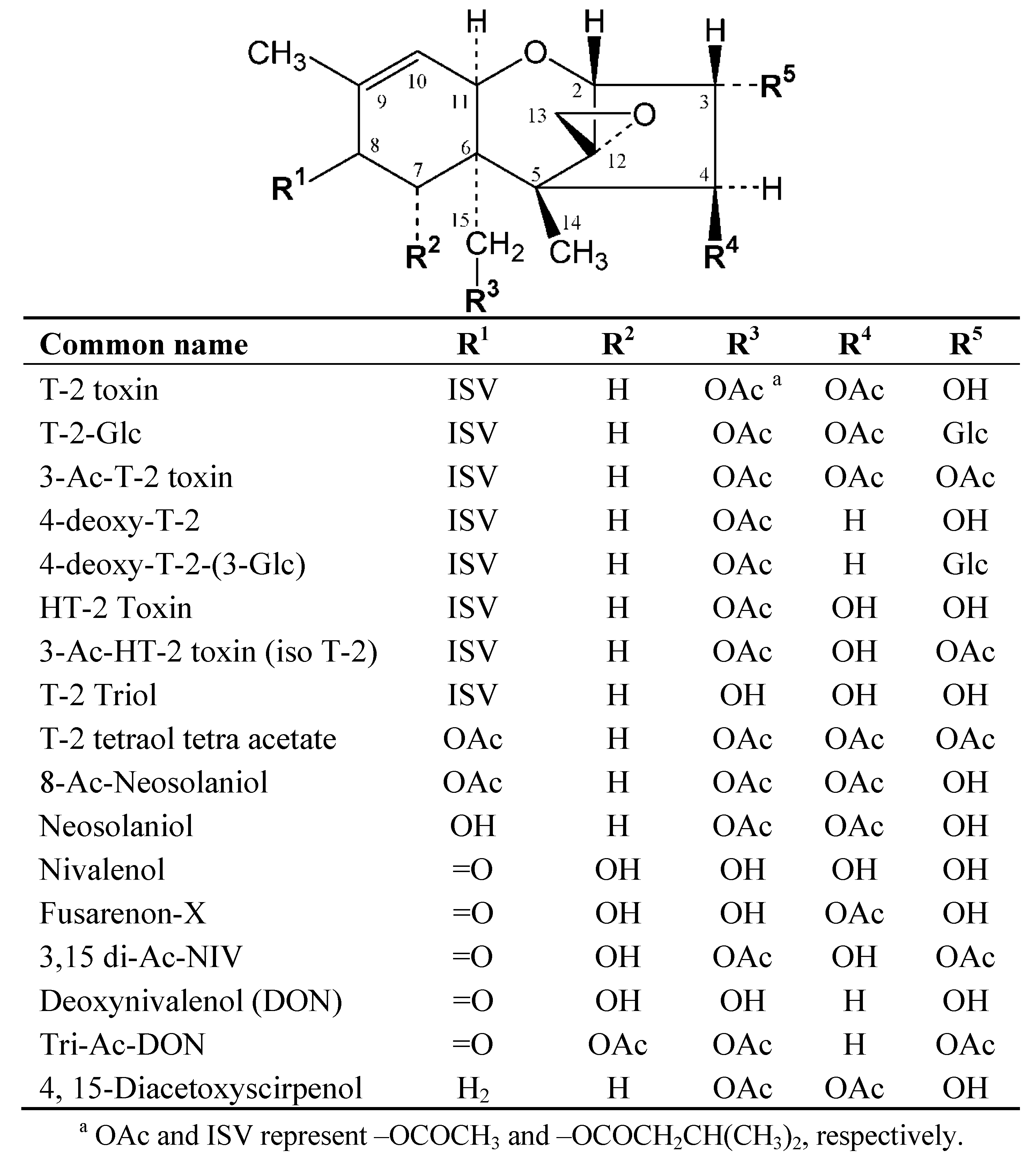
2. Results and Discussion
2.1. Production of Mabs to T2-Glc

{kind=link}
{kind=link}
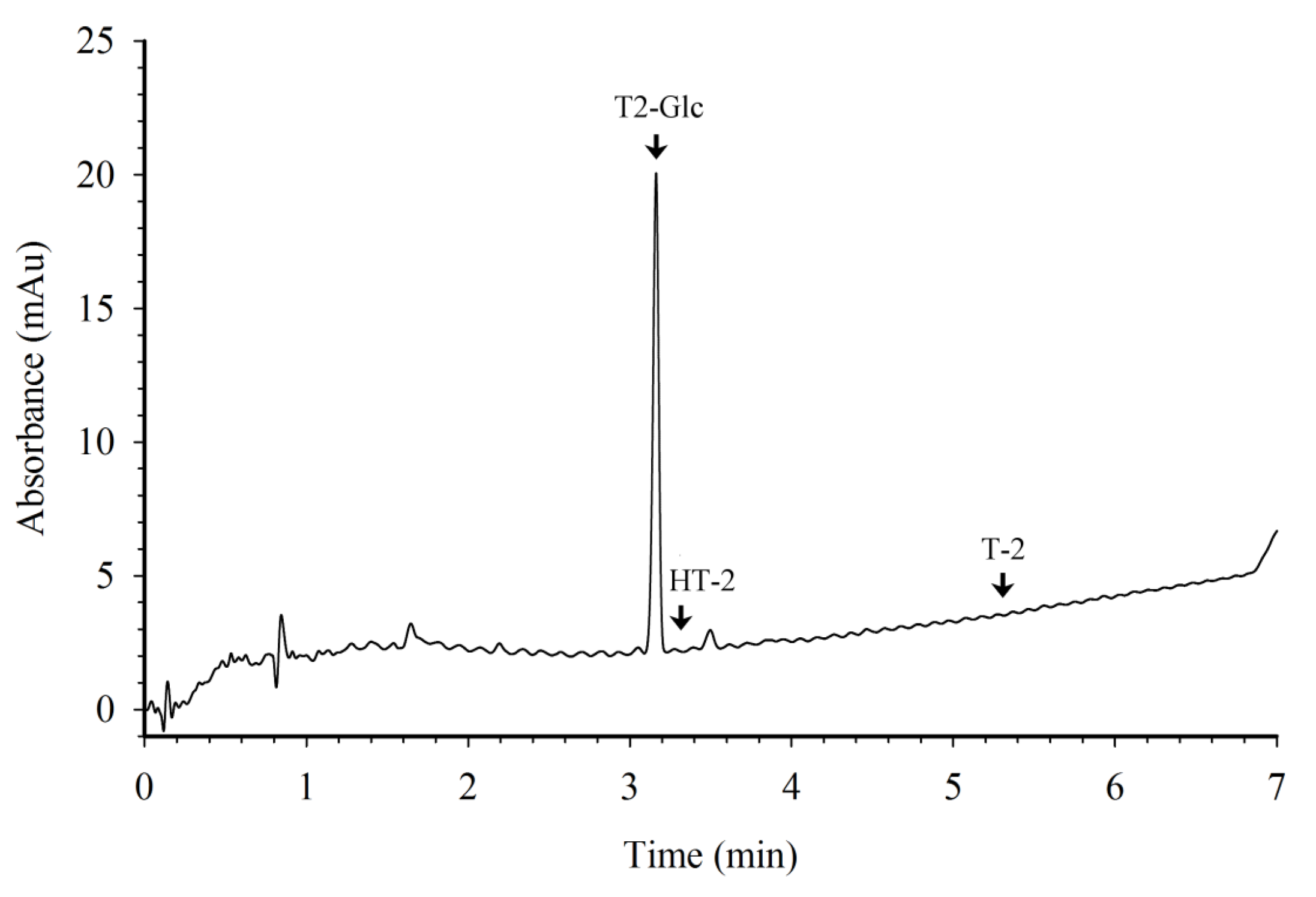{kind=link}
| IC50 (ng/mL) a | Cross-reactivity (%) b | |||||
|---|---|---|---|---|---|---|
| Mab | T2-Glc | T-2 toxin | HT-2 toxin | Mab | T-2 toxin | HT-2 toxin |
| 1-2 | 623 ± 15 | 614 ± 47 | >60000 | 1-2 | 101 ± 8 | <1.0 |
| 1-3 | 8.6 ± 0.8 | 6.3 ± 0.6 | 43.2 ± 7.2 | 1-3 | 135 ± 14 | 19.9 ± 3.3 |
| 1-4 | 13.3 ± 1.7 | 13.5 ± 0.6 | 243 ± 22 | 1-4 | 99.0 ± 5.0 | 5.5 ± 0.5 |
| 2-5 | 13.4 ± 0.6 | 14.0 ± 1.9 | 33.8 ± 2.8 | 2-5 | 96.3 ± 13.1 | 39.8 ± 3.3 |
| 2-11 | 17.6 ± 0.7 | 20.2 ± 1.2 | 562 ± 71 | 2-11 | 86.9 ± 5.5 | 3.1 ± 0.4 |
| 2-13 | 3.5 ± 0.4 | 3.8 ± 0.1 | 271 ± 19 | 2-13 | 91.6 ± 4.1 | 1.3 ± 0.1 |
| 2-16 | 13.6 ± 0.3 | 17.4 ± 1.0 | 321 ± 16 | 2-16 | 78.6 ± 4.9 | 4.3 ± 0.2 |
| 2-17 | 11.3 ± 0.1 | 16.7 ± 1.3 | 118 ± 12 | 2-17 | 67.5 ± 5.5 | 9.5 ± 0.9 |
| 2-21 | 28.2 ± 0.5 | 23.1 ± 1.0 | 255 ± 4 | 2-21 | 122 ± 5 | 11.1 ± 0.2 |
| 2-44 | 7.8 ± 0.4 | 8.5 ± 0.2 | 28.1 ± 5.4 | 2-44 | 91.3 ± 2.6 | 27.7 ± 5.3 |
2.2. Cross-Reactivity of Mab 2-13 with Additional Trichothecenes
| Common name | Cross-Reactivity to Mab 2-13 (%) | a R1 | R2 | R3 | R4 | R5 |
|---|---|---|---|---|---|---|
| T-2-Glc | 100 | ISV | H | OAc | OAc | Glc |
| T-2 toxin | 91.6 ± 4.1 | ISV | H | OAc | OAc | OH |
| 4-deoxy-T-2-(3-Glc) | 2.6 ± 0.1 | ISV | H | OAc | H | Glc |
| 8-Ac-Neosolaniol | 2.5 ± 0.2 | OAc | H | OAc | OAc | OH |
| 3-Ac-T-2 toxin | 2.3 ± 0.1 | ISV | H | OAc | OAc | OAc |
| HT-2 Toxin | 1.3 ± 0.1 | ISV | H | OAc | OH | OH |
| 3-Ac-HT-2 toxin (iso T-2) | 0.1 to 1 | ISV | H | OAc | OH | OAc |
| 4-deoxy-T-2 | 0.1 to 1 | ISV | H | OAc | H | OH |
| Neosolaniol | 0.1 to 1 | OH | H | OAc | OAc | OH |
| T-2 Triol | <0.1 | ISV | H | OH | OH | OH |
| T-2 tetraol tetra acetate | <0.1 | OAc | H | OAc | OAc | OAc |
| Nivalenol (NIV) | <0.1 | =O | OH | OH | OH | OH |
| Fusarenon-X | <0.1 | =O | OH | OH | OAc | OH |
| 3,15 di-Ac-NIV | <0.1 | =O | OH | OAc | OH | OAc |
| Deoxynivalenol (DON) | <0.1 | =O | OH | OH | H | OH |
| Tri-Ac-DON | <0.1 | =O | OAc | OAc | H | OAc |
| 4,15-Diacetoxyscirpenol | <0.1 | H2 | H | OAc | OAc | OH |
2.3. Solvent Tolerance
| Solvent | Solvent concentration a | IC50 (ng/mL) b | Relative response (%) c | N d |
|---|---|---|---|---|
| PBS | 0 | 3.3 ± 0.1 | 100% | 12 |
| Methanol | 5% | 3.3 ± 0.2 | 98% | 3 |
| 10% | 3.7 ± 0.1 | 87% | 3 | |
| 20% | 4.0 ± 0.3 | 81% | 12 | |
| 30% | 4.9 ± 0.5 | 66% | 3 | |
| 50% | 9.2 ± 0.9 | 35% | 3 | |
| Acetonitrile | 5% | 3.7 ± 0.2 | 87% | 3 |
| 10% | 4.4 ± 0.3 | 73% | 6 | |
| 15% | 6.3 ± 0.2 | 52% | 3 | |
| 20% | 7.4 ± 0.2 | 44% | 3 | |
| 30% | 14.3 ± 1.5 | 23% | 3 |
3. Experimental Section
3.1. Reagents
3.2. HPLC with Photodiode Array Detection
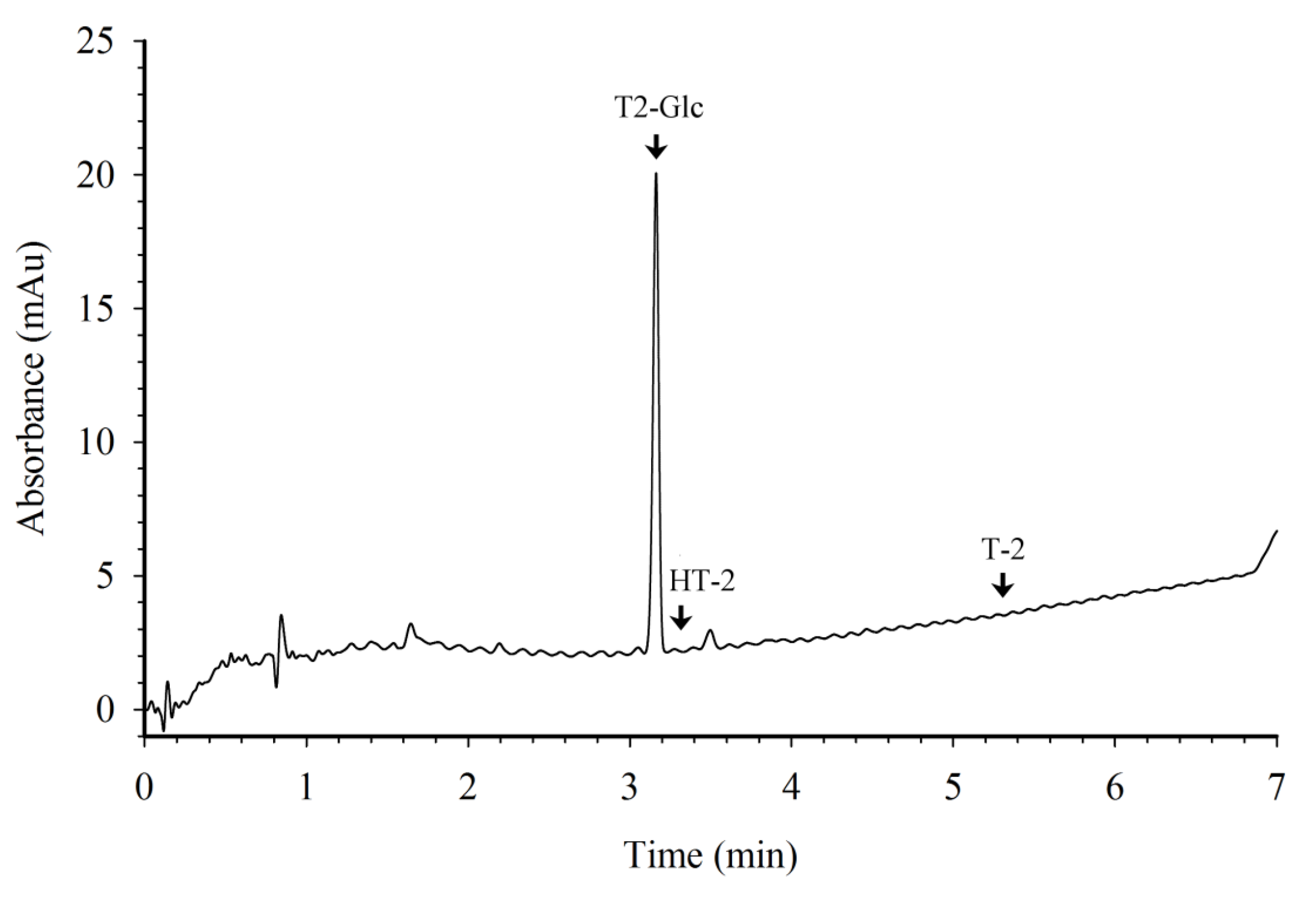
3.3. Preparation and Evaluation of T2-Glc Protein Conjugates
3.4. Immunizations and Screening for T2-Glc Specific Antibodies by CI-ELISA
3.5. Production and Purification of Monoclonal Antibodies
3.6. Cross Reactivity of Mabs and Solvent Effects
4. Conclusions
Acknowledgments
Disclaimer
Conflict of Interest
References
- Ueno, Y. Toxicological features of T-2 toxin and related trichothecenes. Toxicol. Sci. 1984, 4, S124–S132. [Google Scholar] [CrossRef]
- Canady, R.A.; Coker, R.D.; Egan, S.K.; Krska, R.; Olsen, M.; Resnik, S.; Schlatter, J. T-2 and HT-2 Toxins. WHO Food Additive Series: 47 Safety Evaluation of Certain Mycotoxins in Food. Joint FAO/WHO Expert Committee on Food Additives (JECFA), FAO Food and nutrition paper 74. 2001. Available online: http://www.inchem.org/documents/jecfa/jecmono/v47je06.htm (accessed on 31 May 2013).
- Kubena, L.F.; Edrington, T.S.; Harvey, R.B.; Buckley, S.A.; Phillips, T.D.; Rottinghaus, G.E.; Casper, H.H. Individual and combined effects of fumonisin B1 present in Fusarium moniliforme culture material and T-2 toxin or deoxynivalenol in broiler chicks. Poult. Sci. 1997, 76, 1239–1247. [Google Scholar]
- Wyatt, R.D. Biological Effects of Mycotoxins (other than Aflatoxin) on Poultry. In Interactions of Mycotoxins in Animal Production; Naber, E.C., Wallace, H.D., Eds.; National Academy of Sciences: Washington, DC, USA, 1979; pp. 87–95. [Google Scholar]
- European Food Safety Authority (EFSA) Panel on Contaminants in the Food Chain. Scientific opinion on the risks for animal and public health related to the presence of T-2 and HT-2 toxin in food and feed. EFSA J. 2011, 9, 2481–2667.
- Shephard, G.S.; Berthiller, F.; Burdaspal, P.A.; Crews, C.; Jonker, M.A.; Krska, R.; MacDonald, S.; Malone, R.J.; Maragos, C.; Sabino, M.; et al. Developments in mycotoxin analysis: An update for 2010–2011. World Mycotoxin J. 2012, 5, 3–30. [Google Scholar] [CrossRef]
- Li, Y.; Wang, Z.; Beier, R.C.; Shen, J.; de Smet, D.; de Saeger, S.; Zhang, S. T-2 toxin, a trichothecene mycotoxin: Review of toxicity, metabolism, and analytical methods. J. Agric. Food Chem. 2011, 59, 3441–3453. [Google Scholar] [CrossRef]
- Meneely, J.P.; Ricci, F.; van Egmond, H.P.; Elliott, C.T. Current methods of analysis for the determination of trichothecene mycotoxins in food. Trends Anal. Chem. 2011, 30, 192–203. [Google Scholar]
- Pascale, M.; Panzarini, G.; Visconti, A. Determination of HT-2 and T-2 toxins in oats and wheat by ultra-performance liquid chromatography with photodiode array detection. Talanta 2012, 89, 231–236. [Google Scholar] [CrossRef]
- Lippolis, V.; Pascale, M.; Maragos, C.M.; Visconti, A. Improvement of detection sensitivity of T-2 and HT-2 toxins using different fluorescent labeling reagents by high-performance liquid chromatography. Talanta 2008, 74, 1476–1483. [Google Scholar] [CrossRef]
- Rasmussen, P.H.; Nielsen, K.F.; Ghorbani, F.; Spliid, N.H.; Nielsen, G.C.; Jørgensen, L.N. Occurrence of different trichothecenes and deoxynivalenol-3-β-D-glucoside in naturally and artificially contaminated Danish cereal grains and whole maize plants. Mycotoxin Res. 2012, 28, 181–190. [Google Scholar] [CrossRef]
- Chu, F.S.; Grossman, S.; Wei, R.D.; Mirocha, C.J. Production of antibody against T-2 toxin. Appl. Environ. Microbiol. 1979, 37, 104–108. [Google Scholar]
- Pestka, J.J.; Lee, S.C.; Lau, H.P.; Chu, F.S. Enzyme-linked immunosorbent assay for T-2 toxin. J. Am. Oil Chem. Soc. 1981, 58, A940–A944. [Google Scholar] [CrossRef]
- Hunter, K.W.; Brimfield, A.A.; Miller, M.; Finkelman, F.D.; Chu, S.F. Preparation and characterization of monoclonal antibodies to the trichothecene mycotoxin T-2. Appl. Environ. Microbiol. 1985, 49, 168–172. [Google Scholar]
- Ohtani, K.; Kawamura, O.; Kajii, H.; Chiba, J.; Ueno, Y. Development of enzyme-linked immunosorbent assay (ELISA) for T-2 toxin using monoclonal antibodies. Mycotoxins 1985, 22, 31–32. [Google Scholar]
- Zhang, G.S.; Schubring, S.L.; Chu, F.S. Improved method for production of antibodies against T-2 toxin and diacetoxyscirpenol in rabbits. Appl. Environ. Microbiol. 1986, 51, 132–137. [Google Scholar]
- Fan, T.S.; Zhang, G.S.; Chu, F.S. Production and characterization of antibodies against HT-2 toxin and T-2 tetraol tetraacetate. Appl. Environ. Microbiol. 1987, 53, 17–21. [Google Scholar]
- Goodbrand, I.A.; Stimson, W.H.; Smith, I.E. A monoclonal antibody to T-2 toxin. Lett. Appl. Microbiol. 1987, 5, 97–99. [Google Scholar] [CrossRef]
- Gendloff, E.H.; Pestka, J.J.; Dixon, D.E.; Hart, L.P. Production of a monoclonal antibody to T-2 toxin with strong cross-reactivity to T-2 metabolites. Phytopathology 1987, 77, 57–59. [Google Scholar] [CrossRef]
- Wei, R.D.; Swanson, S.; Chu, F.S. Cross-reactivity of antibodies against T-2 with deepoxide T-2 toxin. Mycotoxin Res. 1988, 4, 15–19. [Google Scholar] [CrossRef]
- Fan, T.S.L.; Schubring, S.L.; Wei, R.D.; Chu, F.S. Production and characterization of a monoclonal antibody cross-reactive with most group A trichothecenes. Appl. Environ. Microbiol. 1988, 54, 2959–2963. [Google Scholar]
- Chiba, J.; Kawamura, O.; Kajii, H.; Ohtani, K.; Nagayama, S.; Ueno, Y. A sensitive enzyme-linked immunosorbent assay for detection of T-2 toxin with monoclonal antibodies. Food Addit. Contam. 1988, 5, 629–639. [Google Scholar] [CrossRef]
- Hack, R.; Martlbauer, E.; Terplan, G. A monoclonal antibody-based enzyme immunoassay for the detection of T-2 toxin at picogram levels. Lett. Appl. Microbiol. 1989, 9, 133–135. [Google Scholar] [CrossRef]
- Chanh, T.C.; Hewetson, J.F. Structure/function studies of T-2 mycotoxin with a monoclonal antibody. Immunopharmacology 1991, 21, 83–89. [Google Scholar] [CrossRef]
- Barna-Vetró, I.; Gyöngyösi, A.; Solti, L. Monoclonal antibody-based enzyme-linked immunosorbent assay of Fusarium T-2 and zearalenone toxins in cereals. Appl. Environ. Microbiol. 1994, 60, 729–731. [Google Scholar]
- Molinelli, A.; Grossalber, K.; Fűhrer, M.; Baumgartner, S.; Sulyok, M.; Krska, R. Development of qualitative and semiquantitative immunoassay-based rapid strip tests for the detection of T-2 toxin in wheat and oat. J. Agric. Food Chem. 2008, 56, 2589–2594. [Google Scholar]
- Baumgartner, S.; Fűhrer, M.; Krska, R. Comparison of monoclonal antibody performance characteristics for the detection of two representatives of A- and B-trichothecenes: T-2 toxin and deoxynivalenol. World Mycotoxin J. 2010, 3, 233–238. [Google Scholar] [CrossRef]
- Wang, J.; Duan, S.; Zhang, Y.; Wang, S. Enzyme-linked immunosorbent assay for the determination of T-2 toxin in cereals and feedstuff. Microchim. Acta 2010, 169, 137–144. [Google Scholar]
- Lippolis, V.; Pascale, M.; Valenzano, S.; Pluchinotta, V.; Baumgartner, S.; Krska, R.; Visconti, A. A rapid fluorescence polarization immunoassay for the determination of T-2 and HT-2 toxins in wheat. Anal. Bioanal. Chem. 2011, 401, 2561–2571. [Google Scholar] [CrossRef]
- Meneely, J.P.; Sulyok, M.; Baumgartner, S.; Krska, R.; Elliott, C.T. A rapid optical immunoassay for the screening of T-2 and HT-2 toxin in cereals and maize-based baby food. Talanta 2010, 81, 630–636. [Google Scholar] [CrossRef]
- Wang, Y.; Liu, N.; Ning, B.; Liu, M.; Lv, Z.; Sun, Z.; Peng, Y.; Chen, C.; Li, J.; Gao, Z. Simultaneous and rapid detection of six different mycotoxins using an immunochip. Biosens. Bioelectron. 2012, 34, 44–50. [Google Scholar] [CrossRef]
- De Saeger, S.; van Egmond, H. Special issue: Masked mycotoxins. World Mycotoxin J. 2012, 5, 203–206. [Google Scholar] [CrossRef]
- Zachariasova, M.; Hajslova, J.; Kostelanska, M.; Poustka, J.; Krplova, A.; Cuhra, P.; Hochel, I. Deoxynivalenol and its conjugates in beer: A critical assessment of data obtained by enzyme-linked immunosorbent assay and liquid chromatography coupled to tandem mass spectrometry. Anal. Chim. Acta 2008, 625, 77–86. [Google Scholar] [CrossRef]
- Berthiller, F.; Schuhmacher, R.; Adam, G.; Krska, R. Formation, determination and significance of masked and other conjugated mycotoxins. Anal. Bioanal. Chem. 2009, 395, 1243–1252. [Google Scholar] [CrossRef]
- Tangni, E.K.; Motte, J.C.; Callebaut, A.; Pussemier, L. Cross-reactivity of antibodies in some commercial deoxynivalenol test kits against some fusariotoxins. J. Agric. Food Chem. 2010, 58, 12625–12633. [Google Scholar] [CrossRef]
- Versilovskis, A.; Huybrecht, B.; Tangni, E.K.; Pussemier, L.; de Saeger, S.; Callebaut, A. Cross-reactivity of some commercially available deoxynivalenol (DON) and zearalenone (ZEN) immunoaffinity columns to DON- and ZEN-conjugated forms and metabolites. Food Addit. Contam. Part A 2011, 28, 1687–1693. [Google Scholar]
- Goryacheva, I.Y.; de Saeger, S. Immunochemical detection of masked mycotoxins: A short review. World Mycotoxin J. 2012, 5, 281–287. [Google Scholar] [CrossRef]
- Busman, M.; Poling, S.M.; Maragos, C.M. Observation of T-2 toxin and HT-2 toxin glucosides from Fusarium sporotrichioides by liquid chromatography coupled to tandem mass spectrometry (LC-MS/MS). Toxins 2011, 3, 1554–1568. [Google Scholar] [CrossRef]
- Lattanzio, V.M.T.; Visconti, A.; Haidukowski, M.; Pascale, M. Identification and characterization of new Fusarium masked mycotoxins, T2 and HT2 glycosyl derivatives, in naturally contaminated wheat and oats by liquid chromatography high-resolution mass spectrometry. J. Mass. Spectrom. 2012, 47, 466–475. [Google Scholar] [CrossRef]
- Veprikova, Z.; Vaclavikova, M.; Lacina, O.; Dzuman, Z.; Zachariasova, M.; Hajslova, J. Occurrence of mono- and di-glycosylated conjugates of T-2 and HT-2 toxins in naturally contaminated cereals. World Mycotoxin J. 2012, 5, 231–240. [Google Scholar] [CrossRef]
- Nakagawa, H.; Ohmichi, K.; Sakamoto, S.; Sago, Y.; Kushiro, M.; Nagashima, H.; Yoshida, M.; Nakajima, T. Detection of a new Fusarium masked mycotoxin in wheat grain by high-resolution LC-Orbitrap MS. Food Addit. Contam. Part A 2011, 28, 1447–1456. [Google Scholar] [CrossRef]
- Nakagawa, H.; Sakamoto, S.; Sago, Y.; Kushiro, M.; Nagashima, H. The use of LC-Orbitrap MS for the detection of Fusarium masked mycotoxins: the case of type A trichothecenes. World Mycotoxin J. 2012, 5, 271–280. [Google Scholar] [CrossRef]
- McCormick, S.P.; Price, N.P.J.; Kurtzman, C.P. Glucosylation and other biotransformations of T-2 toxin by yeasts of the Trichomonascus clade. Appl. Environ. Microbiol. 2012, 78, 8694–8702. [Google Scholar] [CrossRef]
- Saba, J.A.; Shen, X.; Jamieson, J.C.; Perreault, H. Investigation of different combinations of derivitization, separation methods and electrospray ionization mass spectrometry for standard oligosaccharides and glycans from ovalbumin. J. Mass Spectrom. 2001, 36, 563–574. [Google Scholar] [CrossRef]
- Poppenberger, B.; Berthiller, F.; Lucyshyn, D.; Sieberer, T.; Schuhmacher, R.; Krska, R.; Kuchler, K.; Glössl, J.; Luschnig, C.; Adam, G. Detoxification of Fusarium mycotoxin deoxynivalenol by a UDP-glucosyltransferase from Arabidopsis thaliana. J. Biol. Chem. 2003, 278, 47905–47914. [Google Scholar] [CrossRef]
- Zachariasova, M.; Vaclavikova, M.; Lacina, O.; Vaclavik, L.; Hajslova, J. Deoxynivalenol oligoglycosides: New “masked” Fusarium toxins occurring in malt, beer, and breadstuff. J. Agric. Food Chem. 2012, 60, 9280–9291. [Google Scholar] [CrossRef]
- Maragos, C.M.; McCormick, S.P. Monoclonal antibodies for the mycotoxins deoxynivalenol and 3-acetyl-deoxynivalenol. Food Agric. Immunol. 2000, 12, 181–192. [Google Scholar] [CrossRef]
- Maragos, C.M.; Busman, M.; Plattner, R.D. Development of monoclonal antibodies for the fusarin mycotoxins. Food Addit. Contam. 2008, 25, 105–114. [Google Scholar] [CrossRef]
- Hoogenraad, N.; Newman, T.; Hoogenraad, J. The effect of pre-injection of mice with pristine on ascites tumour formation and monoclonal antibody production. J. Immunol. Methods 1983, 61, 317–320. [Google Scholar] [CrossRef]
© The authors are employees of the U.S. Department of Agriculture (USDA), and the article is a U.S. Government Work, published by MDPI, Basel, Switzerland, with the permission of USDA. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Maragos, C.M.; Kurtzman, C.; Busman, M.; Price, N.; McCormick, S. Development and Evaluation of Monoclonal Antibodies for the Glucoside of T-2 Toxin (T2-Glc). Toxins 2013, 5, 1299-1313. https://doi.org/10.3390/toxins5071299
Maragos CM, Kurtzman C, Busman M, Price N, McCormick S. Development and Evaluation of Monoclonal Antibodies for the Glucoside of T-2 Toxin (T2-Glc). Toxins. 2013; 5(7):1299-1313. https://doi.org/10.3390/toxins5071299
Chicago/Turabian StyleMaragos, Chris M., Cletus Kurtzman, Mark Busman, Neil Price, and Susan McCormick. 2013. "Development and Evaluation of Monoclonal Antibodies for the Glucoside of T-2 Toxin (T2-Glc)" Toxins 5, no. 7: 1299-1313. https://doi.org/10.3390/toxins5071299
APA StyleMaragos, C. M., Kurtzman, C., Busman, M., Price, N., & McCormick, S. (2013). Development and Evaluation of Monoclonal Antibodies for the Glucoside of T-2 Toxin (T2-Glc). Toxins, 5(7), 1299-1313. https://doi.org/10.3390/toxins5071299