Rationally Designed Synthetic Haptens to Generate Anti-Ciguatoxin Monoclonal Antibodies, and Development of a Practical Sandwich ELISA to Detect Ciguatoxins



Abstract
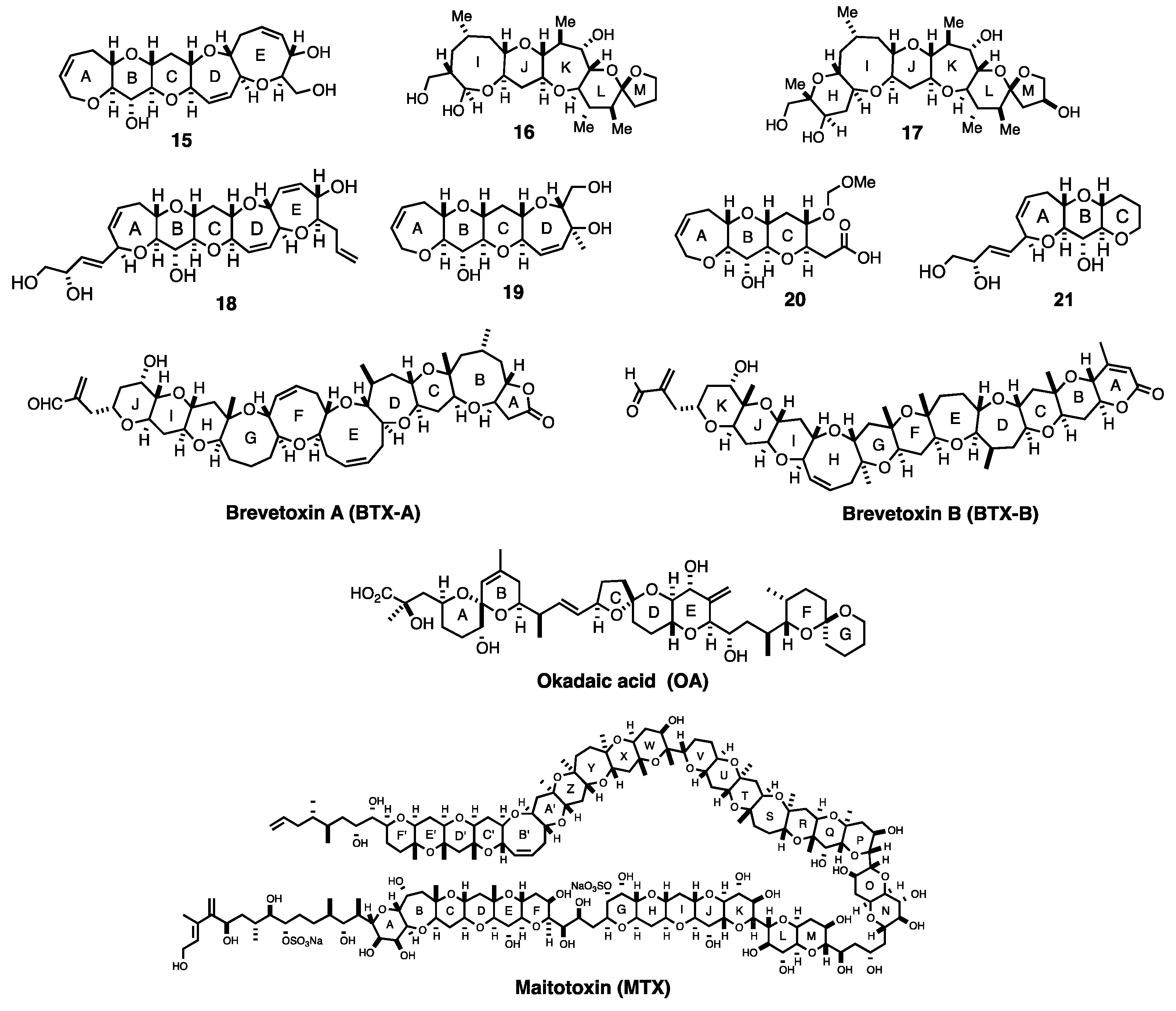
:1. Introduction
2. Generation of Anti-CTX MAbs
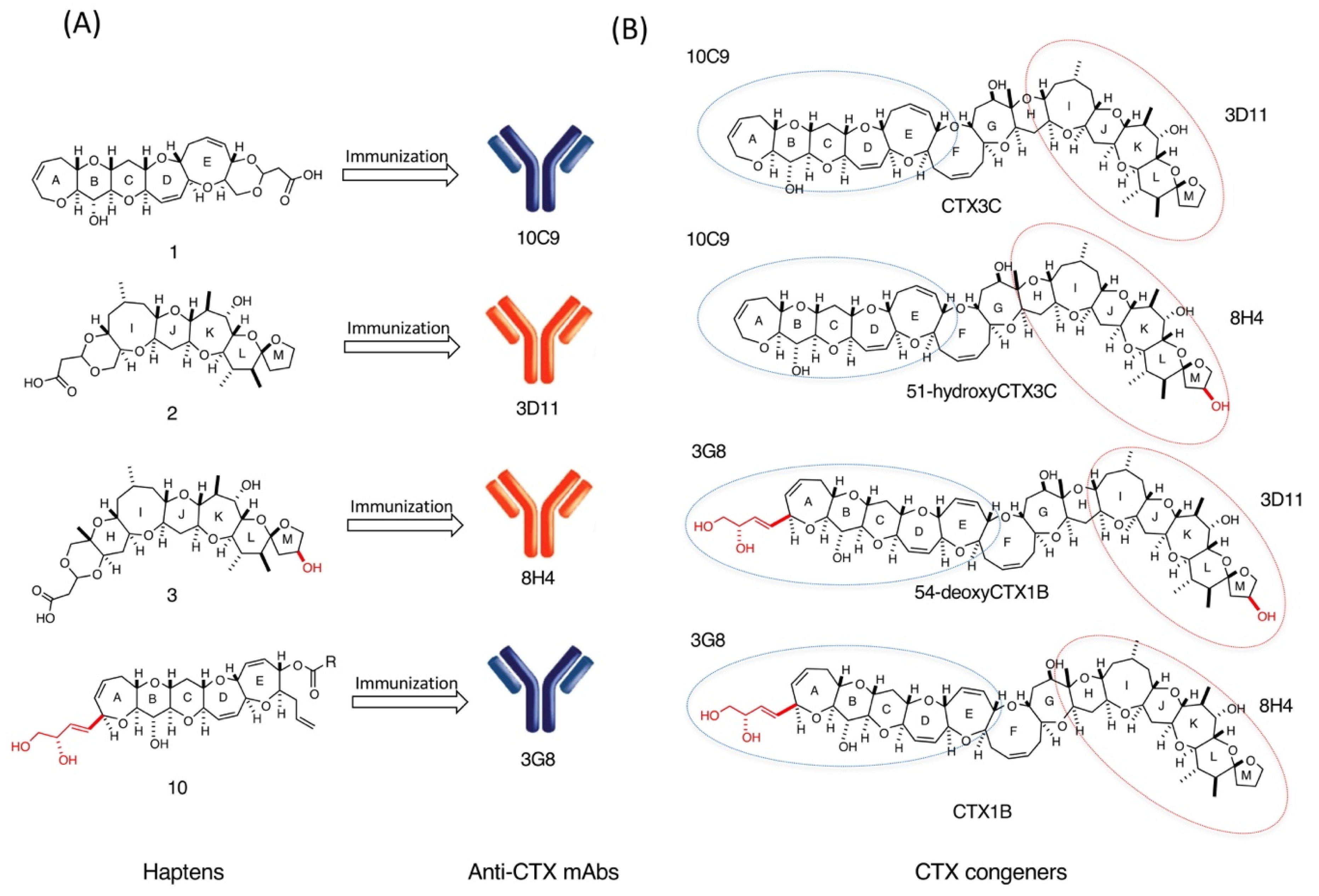
2.1. Design of Haptens and Preparation of Protein-Conjugates
2.2. Production and Affinity Measurement of Anti-CTX mAbs
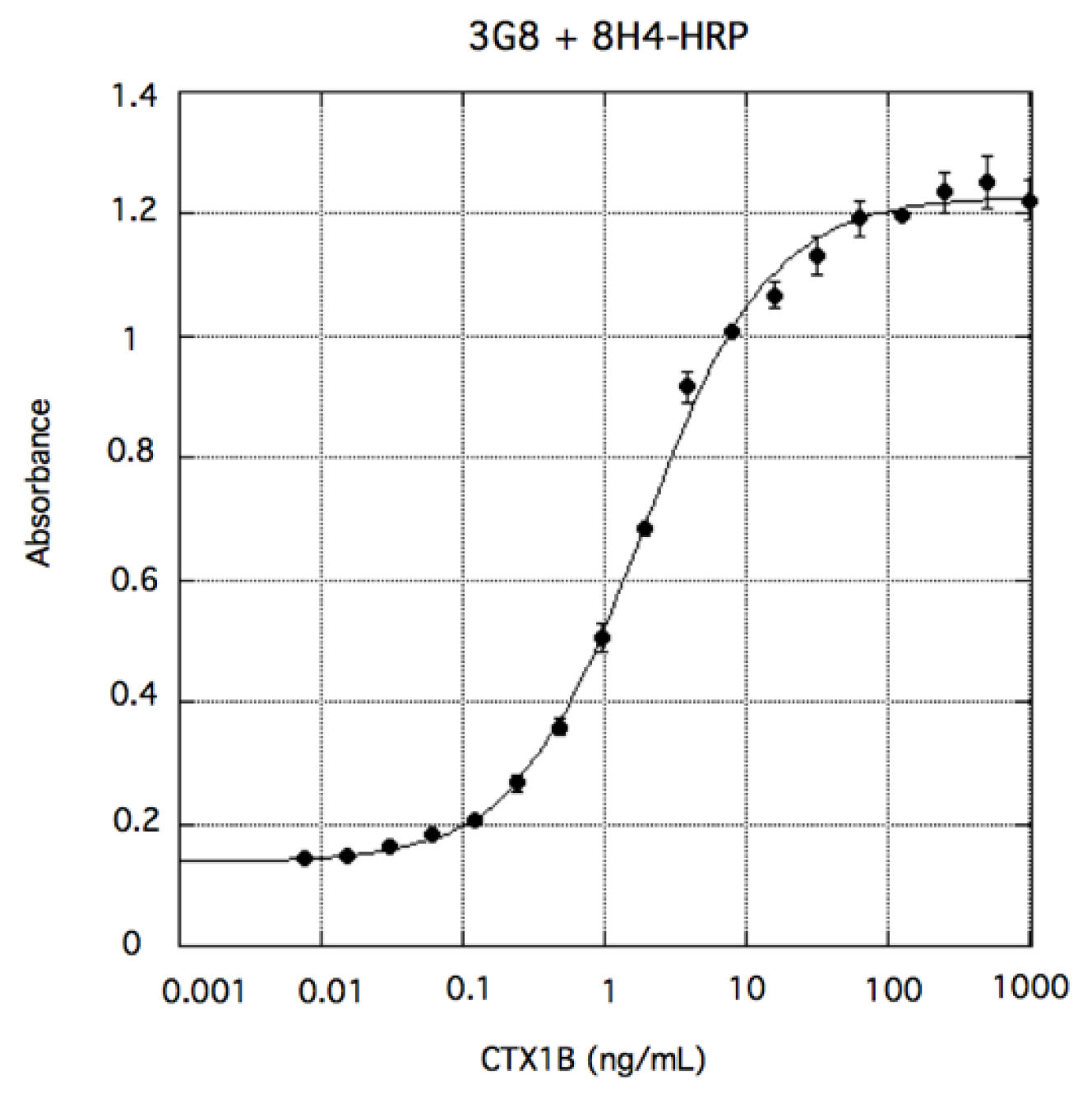
3. Sandwich ELISA Detection of CTXs
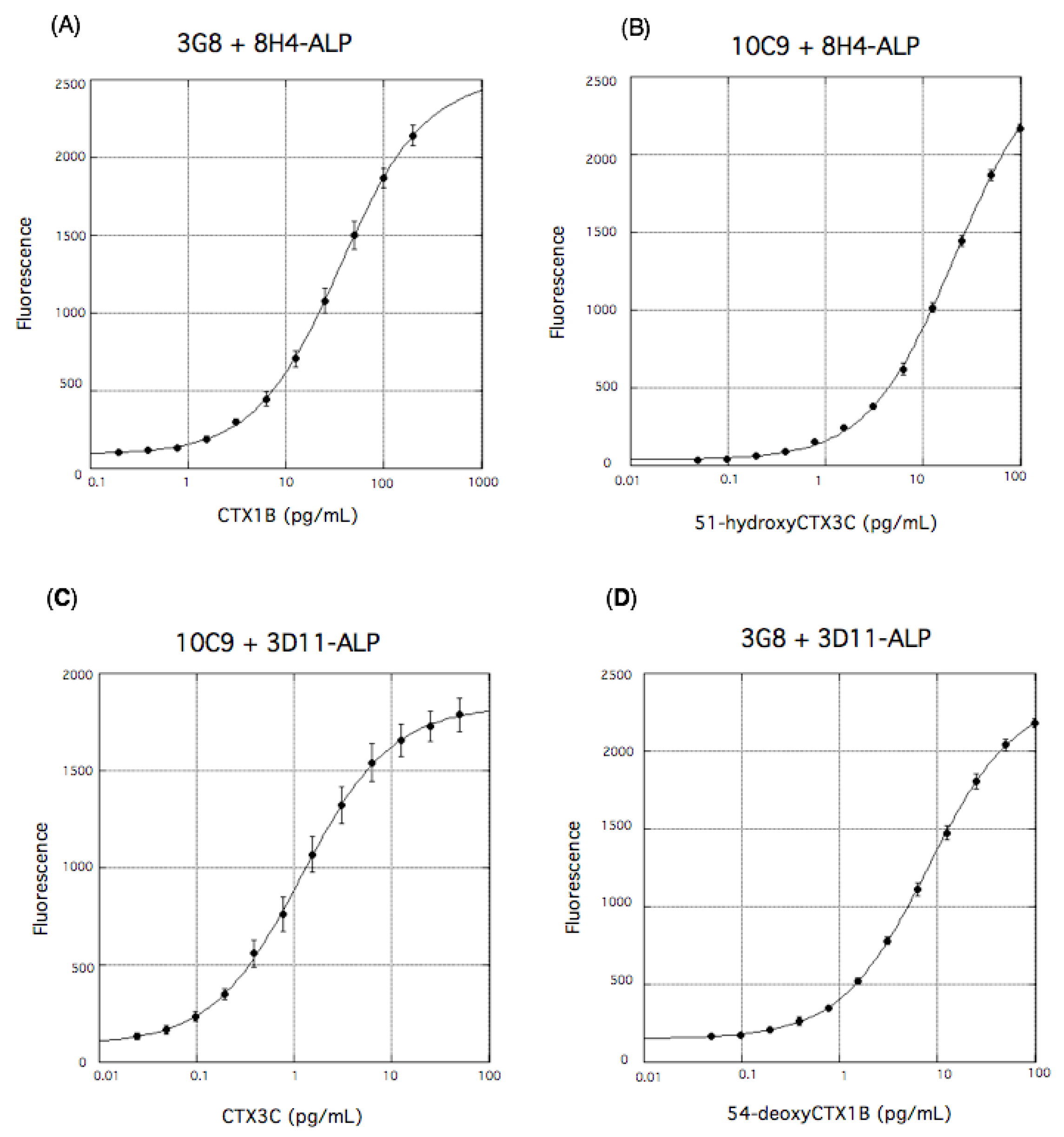
4. Highly Sensitive Detection of CTXs
4.1. Fluorescent Sandwich ELISA Detection of CTXs
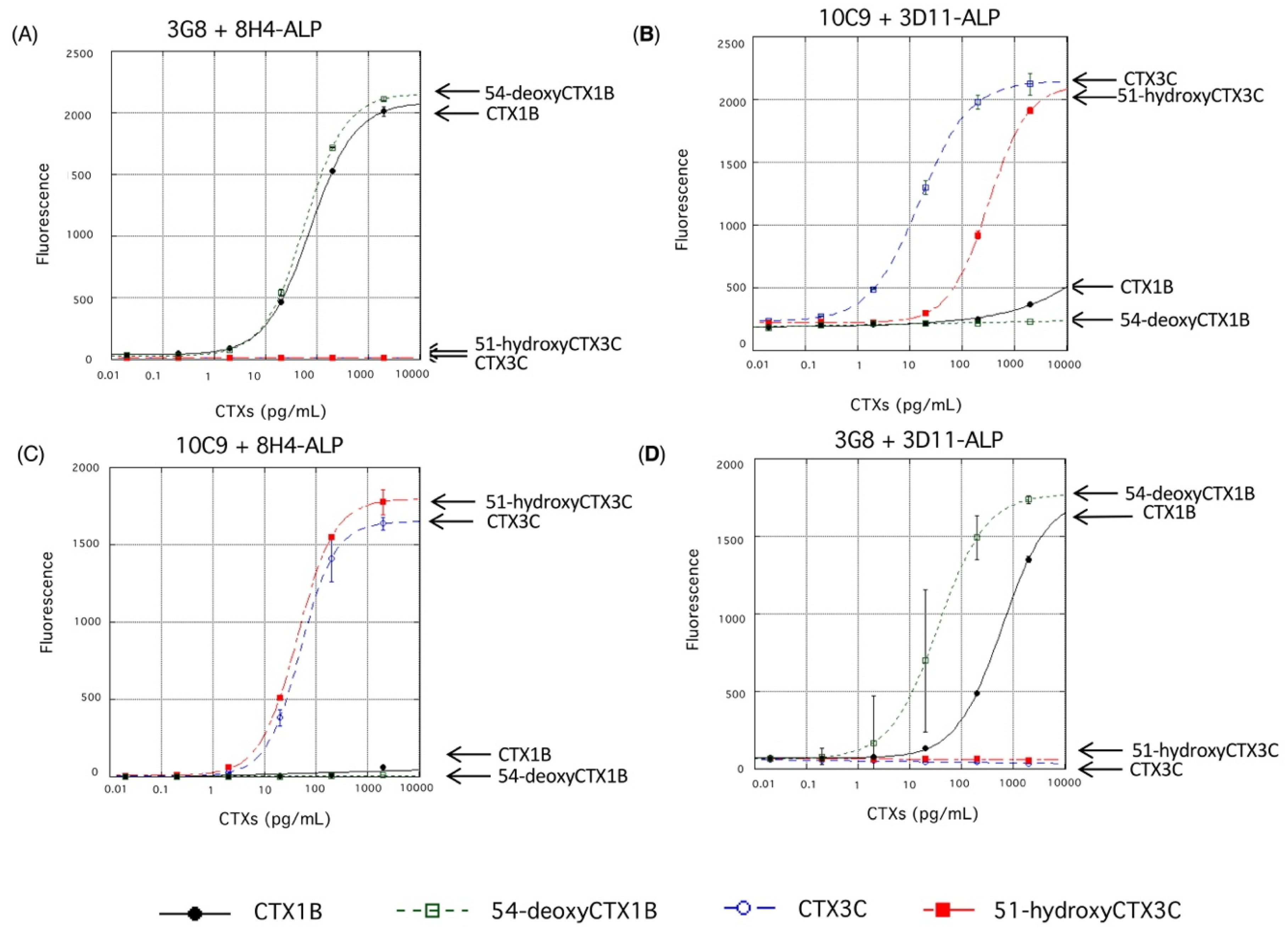
4.2. Cross-Reactivity of the Fluorescent Sandwich ELISA
4.3. Single ELISA Analysis of Four CTX Congeners
4.4. Detection of CTX1B from Fish Flesh and Evaluation of the Matrix Effects at Each Extraction/Purification Step
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Yasumoto, T.; Murata, M. Marine toxins. Chem. Rev. 1993, 93, 1897–1909. [Google Scholar] [CrossRef]
- Yasumoto, T. The chemistry and biological function of natural marine toxins. Chem. Rec. 2001, 1, 228–242. [Google Scholar] [CrossRef] [PubMed]
- Scheuer, P.J. Tetrahedron perspective number 2. Ciguatera and its off-shoots-chance encounters en route to a molecular structures. Tetrahedron 1994, 50, 3–18. [Google Scholar] [CrossRef]
- Lewis, R.J. The changing face of ciguatera. Toxicon 2001, 39, 97–106. [Google Scholar] [CrossRef]
- Dickey, R.W.; Plakas, S.M. Ciguatera: A public health perspective. Toxicon 2010, 56, 123–136. [Google Scholar] [CrossRef] [PubMed]
- Bagnis, R. Ciguatera in French Polynesian islands: Of coral, fish and men | La ciguatera dans les îles de Polynésie française: Des coraux, des poissons et des hommes. Bull. Soc. Pathol. Exot. 1992, 85, 412–414. [Google Scholar] [PubMed]
- Murata, M.; Yasumoto, T. The structure elucidation and biological activities of high molecular weight algal toxins: Maitotoxin, prymnesins and zooxanthellatoxins. Nat. Prod. Rep. 2000, 17, 293–314. [Google Scholar] [CrossRef] [PubMed]
- Murata, M.; Legrand, A.M.; Ishibashi, Y.; Yasumoto, T. Structures of Ciguatoxin and Its Congener. J. Am. Chem. Soc. 1989, 111, 8929–8931. [Google Scholar] [CrossRef]
- Murata, M.; Legrand, A.M.; Ishibashi, Y.; Fukui, M.; Yasumoto, T. Structures and Configurations of Ciguatoxin from the Moray Eel Gymnothorax javanicus and Its Likely Precursor from the Dinoflagellate Gambierdiscus toxicus. J. Am. Chem. Soc. 1990, 112, 4380–4386. [Google Scholar] [CrossRef]
- Satake, M.; Murata, M.; Yasumoto, T. The structure of CTX3C, a ciguatoxin congener isolated from cultured Gambierdiscus toxicus. Tetrahedron Lett. 1993, 34, 1975–1978. [Google Scholar] [CrossRef]
- Satake, M.; Fukui, M.; Legrand, A.-M.; Cruchet, P.; Yasumoto, T. Isolation and structures of new ciguatoxin analogs, 2,3-dihydroxyCTX3C and 51-hydroxyCTX3C, accumulated in tropical reef fish. Tetrahedron Lett. 1998, 39, 1197–1198. [Google Scholar] [CrossRef]
- Yasumoto, T.; Igarashi, T.; Legrand, A.-M.; Cruchet, P.; Chinain, M.; Fujita, T.; Naoki, H. Structural elucidation of ciguatoxin congeners by fast-atom bombardment tandem mass spectroscopy. J. Am. Chem. Soc. 2000, 122, 4988–4989. [Google Scholar] [CrossRef]
- Otero, P.; Pérez, S.; Alfonso, A.; Vale, C.; Rodríguez, P.; Gouveia, N.N.; Gouveia, N.; Delgado, J.; Vale, P.; Hirama, M.; et al. First toxin profile of ciguateric fish in Madeira Arquipelago (Europe). Anal. Chem. 2010, 82, 6032–6039. [Google Scholar] [CrossRef] [PubMed]
- Silva, M.; Rodriguez, I.; Barreiro, A.; Kaufmann, M.; Neto, A.I.; Hassouani, M.; Sabour, B.; Alfonso, A.; Botana, L.M.; Vasconcelos, V. First report of ciguatoxins in two starfish species: Ophidiaster ophidianus and Marthasterias glacialis. Toxins (Basel) 2015, 7, 3740–3757. [Google Scholar] [CrossRef] [PubMed]
- Lewis, R.J.; Sellin, M.; Poli, M.A.; Norton, R.S.; MacLeod, J.K.; Sheil, M.M. Purification and characterization of ciguatoxins from moray eel (Lycodontis javanicus, Muraenidae). Toxicon 1991, 29, 1115–1127. [Google Scholar] [CrossRef]
- Dechraoui, M.-Y.; Naar, J.; Pauillac, S.; Legrand, A.-M. Ciguatoxins and brevetoxins, neurotoxic polyether compounds active on sodium channels. Toxicon 1999, 37, 125–143. [Google Scholar] [CrossRef]
- Lewis, R.J. Structure of Caribbean ciguatoxin isolated from Caranx latus. J. Am. Chem. Soc. 1998, 120, 5914–5920. [Google Scholar] [CrossRef]
- Manger, R.L.; Leja, L.S.; Lee, S.Y.; Hungerford, J.M.; Hokama, Y.; Dickey, R.W.; Granade, H.R.; Lewis, R.; Yasumoto, T.; Wekell, M.M. Detection of sodium channel toxins: Directed cytotoxicity assays of purified ciguatoxins, brevetoxins, saxitoxins, and seafood extracts. J. AOAC Int. 1995, 78, 521–527. [Google Scholar]
- Poli, M.A.; Lewis, R.J.; Dickey, R.W.; Musser, S.M.; Buckner, C.A.; Carpenter, L.G. Identification of Caribbean ciguatoxins as the cause of an outbreak of fish poisoning among U.S. soldiers in Haiti. Toxicon 1997, 35, 733–741. [Google Scholar] [CrossRef]
- Hardison, D.R.; Holland, W.C.; McCall, J.R.; Bourdelais, A.J.; Baden, D.G.; Darius, H.T.; Chinain, M.; Tester, P.A.; Shea, D.; Quintana, H.A.F.; et al. Fluorescent receptor binding assay for detecting ciguatoxins in fish. PLoS ONE 2016, 11, e0153348. [Google Scholar] [CrossRef]
- Yasumoto, T.; Fukui, M.; Sasaki, K.; Sugiyama, K. Determinations of marine toxins in foods. J. AOAC Int. 1995, 78, 574–582. [Google Scholar] [PubMed]
- Lewis, R.J.; Jones, A. Characterization of ciguatoxins and ciguatoxin congeners present in ciguateric fish by gradient reverse-phase high-performance liquid chromatography/mass spectrometry. Toxicon 1997, 35, 159–168. [Google Scholar] [CrossRef]
- Lewis, R.J.; Jones, A.; Vernoux, J.-P. HPLC/tandem electrospray mass spectrometry for the determination of Sub-ppb levels of pacific and caribbean ciguatoxins in crude extracts of fish. Anal. Chem. 1999, 71, 247–250. [Google Scholar] [CrossRef] [PubMed]
- Lewis, R.J.; Yang, A.; Jones, A. Rapid extraction combined with LC-tandem mass spectrometry (CREM-LC/MS/MS) for the determination of ciguatoxins in ciguateric fish flesh. Toxicon 2009, 54, 62–66. [Google Scholar] [CrossRef] [PubMed]
- Yogi, K.; Oshiro, N.; Inafuku, Y.; Hirama, M.; Yasumoto, T. Detailed LC-MS/MS analysis of ciguatoxins revealing distinct regional and species characteristics in fish and causative alga from the pacific. Anal. Chem. 2011, 83, 8886–8891. [Google Scholar] [CrossRef] [PubMed]
- Yogi, K.; Oshiro, N.; Matsuda, S.; Sakugawa, S.; Matsuo, T.; Yasumoto, T. Toxin profiles in fish implicated in ciguatera fish poisoning in Amami and Kakeroma Islands, Kagoshima prefecture, Japan. J. Food Hyg. Soc. Jpn. 2013, 54, 385–391. [Google Scholar] [CrossRef]
- Hamilton, B.; Hurbungs, M.; Vernoux, J.-P.; Jones, A.; Lewis, R.J. Isolation and characterisation of Indian Ocean ciguatoxin. Toxicon 2002, 40, 685–693. [Google Scholar] [CrossRef]
- Hamilton, B.; Hurbungs, M.; Jones, A.; Lewis, R.J. Multiple ciguatoxins present in Indian Ocean reef fish. Toxicon 2002, 40, 1347–1353. [Google Scholar] [CrossRef]
- Diogène, J.; Reverté, L.; Rambla-Alegre, M.; Del Rió, V.; De La Iglesia, P.; Campàs, M.; Palacios, O.; Flores, C.; Caixach, J.; Ralijaona, C.; et al. Identification of ciguatoxins in a shark involved in a fatal food poisoning in the Indian Ocean. Sci. Rep. 2017, 7, 8240. [Google Scholar] [CrossRef]
- Hokama, Y.; Banner, A.H.H.; Boylan, D.B.B. A radioimmunoassay for the detection of ciguatoxin. Toxicon 1977, 15, 317–325. [Google Scholar] [CrossRef]
- Hokama, Y.; Hong, T.W.P.; Isobe, M.; Ichikawa, Y.; Yasumoto, T. Cross-reactivity of highly purified okadaic acid (OA), synthetic, spiroketal east sphere of OA and ciguatoxin. J. Clin. Lab. Anal. 1992, 6, 54–58. [Google Scholar] [CrossRef] [PubMed]
- Hokama, Y.; Takenaka, W.E.; Nishimura, K.L.; Ebesu, J.S.M.; Bourke, R.; Sullivan, P.K. A Simple Membrane Immunobead Assay for Detecting Ciguatoxin and Related Polyethers from Human Ciguatera Intoxication and Natural Reef Fishes. J. AOAC Int. 1998, 81, 727–735. [Google Scholar] [PubMed]
- Hokama, Y. Immunological studies using monoclonal antibodies for detection of low dalton marine toxins. Food Addit. Contam. 1993, 10, 83–95. [Google Scholar] [CrossRef] [PubMed]
- Lehane, L.; Lewis, R.J. Ciguatera: Recent advances but the risk remains. Int. J. Food Microbiol. 2000, 61, 91–125. [Google Scholar] [CrossRef]
- Wong, C.-K.; Hung, P.; Lee, K.L.H.; Kam, K.-M. Study of an outbreak of ciguatera fish poisoning in Hong Kong. Toxicon 2005, 46, 563–571. [Google Scholar] [CrossRef]
- Hirama, M.; Oishi, T.; Uehara, H.; Inoue, M.; Maruyama, M.; Oguri, H.; Satake, M. Total synthesis of ciguatoxin CTX3C. Science 2001, 294, 1904–1907. [Google Scholar] [CrossRef]
- Inoue, M.; Miyazaki, K.; Uehara, H.; Maruyama, M.; Hirama, M. First- and second-generation total synthesis of ciguatoxin CTX3C. Proc. Natl. Acad. Sci. USA 2004, 101, 12013–12018. [Google Scholar] [CrossRef] [Green Version]
- Inoue, M.; Miyazaki, K.; Ishihara, Y.; Tatami, A.; Ohnuma, Y.; Kawada, Y.; Komano, K.; Yamashita, S.; Lee, N.; Hirama, M. Total synthesis of ciguatoxin and 51-hydroxyCTX3C. J. Am. Chem. Soc. 2006, 128, 9352–9354. [Google Scholar] [CrossRef]
- Yamashita, S.; Ishihara, Y.; Morita, H.; Uchiyama, J.; Takeuchi, K.; Inoue, M.; Hirama, M. Stereoselective 6-exo radical cyclization using cis-Vinyl sulfoxide: Practical total synthesis of CTX3C. J. Nat. Prod. 2011, 74, 357–364. [Google Scholar] [CrossRef]
- Yamashita, S.; Takeuchi, K.; Koyama, T.; Inoue, M.; Hayashi, Y.; Hirama, M. Practical route to the left wing of CTX1B and total syntheses of CTX1B and 54-deoxyCTX1B. Chem.—A Eur. J. 2015, 21, 2621–2628. [Google Scholar] [CrossRef]
- Oguri, H.; Tanaka, S.-I.; Hishiyama, S.; Oishi, T.; Hirama, M.; Tsumuraya, T.; Tomioka, Y.; Mizugaki, M. Designed hapten aimed at anti-ciguatoxin monoclonal antibody: Synthesis, immunization and discrimination of the C2 configuration. Synthesis (Stuttgart) 1999, 1999, 1431–1436. [Google Scholar] [CrossRef]
- Nagumo, Y.; Oguri, H.; Shindo, Y.; Sasaki, S.-Y.S.; Oishi, T.; Hirama, M.; Tomioka, Y.; Mizugaki, M.; Tsumuraya, T. Concise Synthesis of Ciguatoxin ABC-Ring Fragments and Surface Plasmon Resonance Study of the Interaction of their BSA Conjugates with Monoclonal Antibodies. Bioorg. Med. Chem. Lett. 2001, 11, 2037–2040. [Google Scholar] [CrossRef]
- Nagumo, Y.; Oguri, H.; Tsumoto, K.; Shindo, Y.; Hirama, M.; Tsumuraya, T.; Fujii, I.; Tomioka, Y.; Mizugaki, M.; Kumagai, I. Phage-display selection of antibodies to the left end of CTX3C using synthetic fragments. J. Immunol. Methods 2004, 289, 137–146. [Google Scholar] [CrossRef] [PubMed]
- Oguri, H.; Hirama, M.; Tsumuraya, T.; Fujii, I.; Maruyama, M.; Uehara, H.; Nagumo, Y. Synthesis-based approach toward direct sandwich immunoassay for ciguatoxin CTX3C. J. Am. Chem. Soc. 2003, 125, 7608–7612. [Google Scholar] [CrossRef] [PubMed]
- Tsumuraya, T.; Fujii, I.; Inoue, M.; Tatami, A.; Miyazaki, K.; Hirama, M. Production of monoclonal antibodies for sandwich immunoassay detection of ciguatoxin 51-hydroxyCTX3C. Toxicon 2006, 48, 287–294. [Google Scholar] [CrossRef]
- Tsumuraya, T.; Fujii, I.; Hirama, M. Production of monoclonal antibodies for sandwich immunoassay detection of Pacific ciguatoxins. Toxicon 2010, 56, 797–803. [Google Scholar] [CrossRef] [PubMed]
- Tsumuraya, T.; Takeuchi, K.; Yamashita, S.; Fujii, I.; Hirama, M. Development of a monoclonal antibody against the left wing of ciguatoxin CTX1B: Thiol strategy and detection using a sandwich ELISA. Toxicon 2012, 60, 348–357. [Google Scholar] [CrossRef]
- Tsumuraya, T.; Fujii, I.; Hirama, M. Preparation of anti-ciguatoxin monoclonal antibodies using synthetic haptens: Sandwich ELISA detection of ciguatoxins. J. AOAC Int. 2014, 97, 373–379. [Google Scholar] [CrossRef]
- Tsumuraya, T.; Sato, T.; Hirama, M.; Fujii, I. Highly Sensitive and Practical Fluorescent Sandwich ELISA for Ciguatoxins. Anal. Chem. 2018, 90, 7318–7324. [Google Scholar] [CrossRef]
- Arevalo, J.H.; Taussig, M.J.; Wilson, I.A. Molecular basis of crossreactivity and the limits of antibody–antigen complementarity. Nature 1993, 365, 859–863. [Google Scholar] [CrossRef]
- Tanaka, F.; Kinoshita, K.; Tanimura, R.; Fujii, I. Relaxing Substrate Specificity in Antibody-Catalyzed Reactions: Enantioselective Hydrolysis of N -Cbz-Amino Acid Esters. J. Am. Chem. Soc. 1996, 118, 2332–2339. [Google Scholar] [CrossRef]
- Romesberg, F.E.; Spiller, B.; Schultz, P.G.; Stevens, R.C. Immunological origins of binding and catalysis in a Diels-Alderase antibody. Science 1998, 279, 1929–1933. [Google Scholar] [CrossRef] [PubMed]
- Kurihara, S.; Tsumuraya, T.; Suzuki, K.; Kuroda, M.; Liu, L.; Takaoka, Y.; Fujii, I. Antibody-catalyzed removal of the p-nitrobenzyl ester protecting group: The molecular basis of broad substrate specificity. Chem. Eur. J. 2000, 6, 1656–1662. [Google Scholar] [CrossRef]
- Tsumuraya, T.; Takazawa, N.; Tsunakawa, A.; Fleck, R.; Masamune, S. Catalytic antibodies induced by a zwitterionic hapten. Chem. Eur. J. 2001, 7, 3748–3755. [Google Scholar] [CrossRef]
- Xu, J.; Deng, Q.; Chen, J.; Houk, K.N.; Bartek, J.; Hilvert, D.; Wilson, I.A. Evolution of shape complementarity and catalytic efficiency from a primordial antibody template. Science 1999, 286, 2345–2348. [Google Scholar] [CrossRef] [PubMed]
- Miyamoto, N.; Yoshimura, M.; Okubo, Y.; Suzuki-Nagata, K.; Tsumuraya, T.; Ito, N.; Fujii, I. Structural basis of the broad substrate tolerance of the antibody 7B9-catalyzed hydrolysis of p-nitrobenzyl esters. Bioorganic Med. Chem. 2018, 26, 1412–1417. [Google Scholar] [CrossRef] [PubMed]
- Friguet, B.; Chaffotte, A.F.; Djavadi-Ohaniance, L.; Goldberg, M.E. Measurements of the true affinity constant in solution of antigen-antibody complexes by enzyme-linked immunosorbent assay. J. Immunol. Methods 1985, 77, 305–319. [Google Scholar] [CrossRef]
- Suga, H.; Ersoy, O.; Tsumuraya, T.; Lee, J.; Sinskey, A.J.; Masamune, S.; Tsumuraya, T.; Sinskey, A.J.; Masamune, S. Esterolytic Antibodies Induced to Haptens with a 1,2-Amino Alcohol Functionality. J. Am. Chem. Soc. 1994, 116, 487–494. [Google Scholar] [CrossRef]
- Shimizu, Y.; Chou, H.N.; Bando, H.; Van Duyne, G.; Clardy, J. Structure of brevetoxin A (GB-1 toxin), the most potent toxin in the Florida red tide organism Gymnodinium breve (Ptychodiscus brevis). J. Am. Chem. Soc. 1986, 108, 514–515. [Google Scholar] [CrossRef]
- Pawlak, J.; Tempesta, M.S.; Golik, J.; Zagorski, M.G.; Lee, M.S.; Nakanishi, K.; Iwashita, T.; Gross, M.L.; Tomer, K.B. Structure of brevetoxin A as constructed from NMR and mass spectral data. J. Am. Chem. Soc. 1987, 109, 1144–1150. [Google Scholar] [CrossRef]
- Lin, Y.-Y.; Risk, M.; Ray, S.M.; Van Engen, D.; Clardy, J.; Golik, J.; James, J.C.; Nakanishi, K. Isolation and structure of brevetoxin B from the “Red Tide” dinoflagellate Ptychodiscus brevis (Gymnodinium breve). J. Am. Chem. Soc. 1981, 103, 6773–6775. [Google Scholar] [CrossRef]
- Lee, M.S.; Repeta, D.J.; Nakanishi, K.; Zagorski, M.G. Biosynthetic origins and assignments of carbon 13 NMR peaks of brevetoxin B. J. Am. Chem. Soc. 1986, 108, 7855–7856. [Google Scholar] [CrossRef] [PubMed]
- Tachibana, K.; Scheuer, P.J.; Tsukitani, Y.; Kikuchi, H.; Van Engen, D.; Clardy, J.; Gopichand, Y.; Schmitz, F.J. Okadaic acid, a cytotoxic polyether from two marine sponges of the genus Halichondria. J. Am. Chem. Soc. 1981, 103, 2469–2471. [Google Scholar] [CrossRef]
- Murata, M.; Iwashita, T.; Yokoyama, A.; Sasaki, M.; Yasumoto, T. Partial structures of maitotoxin, the most potent marine toxin from the dinoflagellate Gambierdiscus toxicus. J. Am. Chem. Soc. 1992, 114, 6594–6596. [Google Scholar] [CrossRef]
- Ui, M.; Tanaka, Y.; Tsumuraya, T.; Fujii, I.; Inoue, M.; Hirama, M.; Tsumoto, K. How protein recognizes ladder-like polycyclic ethers: Interactions between ciguatoxin (CTX3C) fragments and its specific antibody 10C9. J. Biol. Chem. 2008, 283, 19440–19447. [Google Scholar] [CrossRef] [PubMed]
- Food and Drug Administration. Fish and Fishery Products Hazards and Controls Guidance, 4th ed.; Diane Publishing Co.: Philadelphia, PA, USA, 2011. [Google Scholar]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| mAb | Kd (nM) | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| 15 | 16 | 17 | 18 | CTX1B | CTX3C | 51-hydroxy-CTX3C | BTX-A | BTX-B | OA | MTX | |
| 10C9 | 0.8 | NI | ND | ND | ND | 2.8 | ND | NI | NI | NI | NI |
| 3D11 | NI | 8.6 | ND | ND | ND | 122 | ND | 43000 | NI | NI | NI |
| 8H4 | ND | ND | 48 | ND | 20.4 | 3200 | 13.6 | NI | NI | NI | NI |
| 3G8 | ND | ND | ND | 1.5 | 15 | NI | ND | NI | NI | NI | ND |
| Methods | Fluorescent Sandwich Elisa | LC-MS/MS | Receptor Binding Assay (RBA) | Cell-Based Assay (CBA-N2a) |
|---|---|---|---|---|
| LOD | 0.09–0.16 pg/mL (18–32 pg/kg) 1 | 20 pg/mL (4 ng/kg) 1 | 0.075 ppb (75 ng/kg) | 0.002 ppb (2 ng/kg) |
| Specificity | high | high | none | none |
| Sample preparation methods | Extraction + solid phase extraction cartridges (Florisil + PSA) | Extraction + solid phase extraction cartridges (Florisil + PSA) | Extraction + solid phase extraction cartridge (C18) | Extraction + solid phase extraction cartridge (C18) |
| Sample-to-answer time | 2.5 h | 30 min | <3 h | 2.5 days |
| References | [49] | [25,26] | [16,19,20] | [18,20] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tsumuraya, T.; Hirama, M. Rationally Designed Synthetic Haptens to Generate Anti-Ciguatoxin Monoclonal Antibodies, and Development of a Practical Sandwich ELISA to Detect Ciguatoxins. Toxins 2019, 11, 533. https://doi.org/10.3390/toxins11090533
Tsumuraya T, Hirama M. Rationally Designed Synthetic Haptens to Generate Anti-Ciguatoxin Monoclonal Antibodies, and Development of a Practical Sandwich ELISA to Detect Ciguatoxins. Toxins. 2019; 11(9):533. https://doi.org/10.3390/toxins11090533
Chicago/Turabian StyleTsumuraya, Takeshi, and Masahiro Hirama. 2019. "Rationally Designed Synthetic Haptens to Generate Anti-Ciguatoxin Monoclonal Antibodies, and Development of a Practical Sandwich ELISA to Detect Ciguatoxins" Toxins 11, no. 9: 533. https://doi.org/10.3390/toxins11090533
APA StyleTsumuraya, T., & Hirama, M. (2019). Rationally Designed Synthetic Haptens to Generate Anti-Ciguatoxin Monoclonal Antibodies, and Development of a Practical Sandwich ELISA to Detect Ciguatoxins. Toxins, 11(9), 533. https://doi.org/10.3390/toxins11090533