Suppressive Effects of Bee Venom-Derived Phospholipase A2 on Mechanical Allodynia in a Rat Model of Neuropathic Pain


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
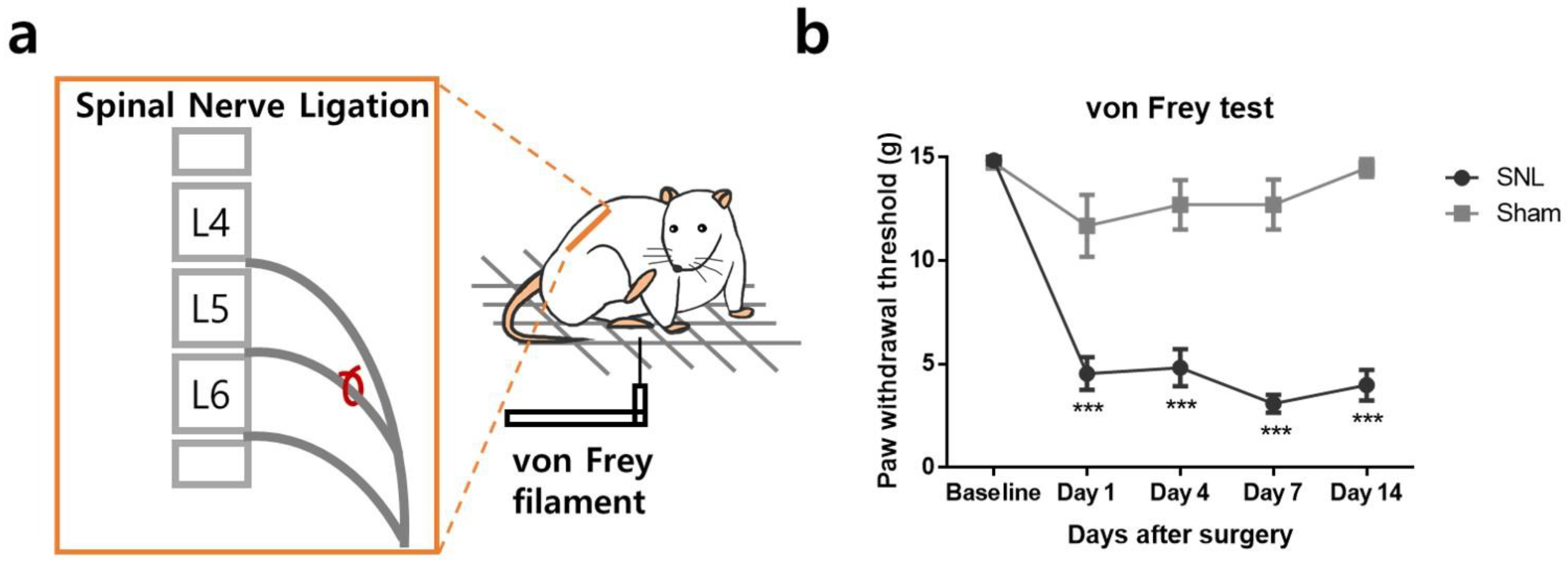
2.1. L5 Spinal Nerve Ligation Induces Mechanical Allodynia in the Affected Hind Paw
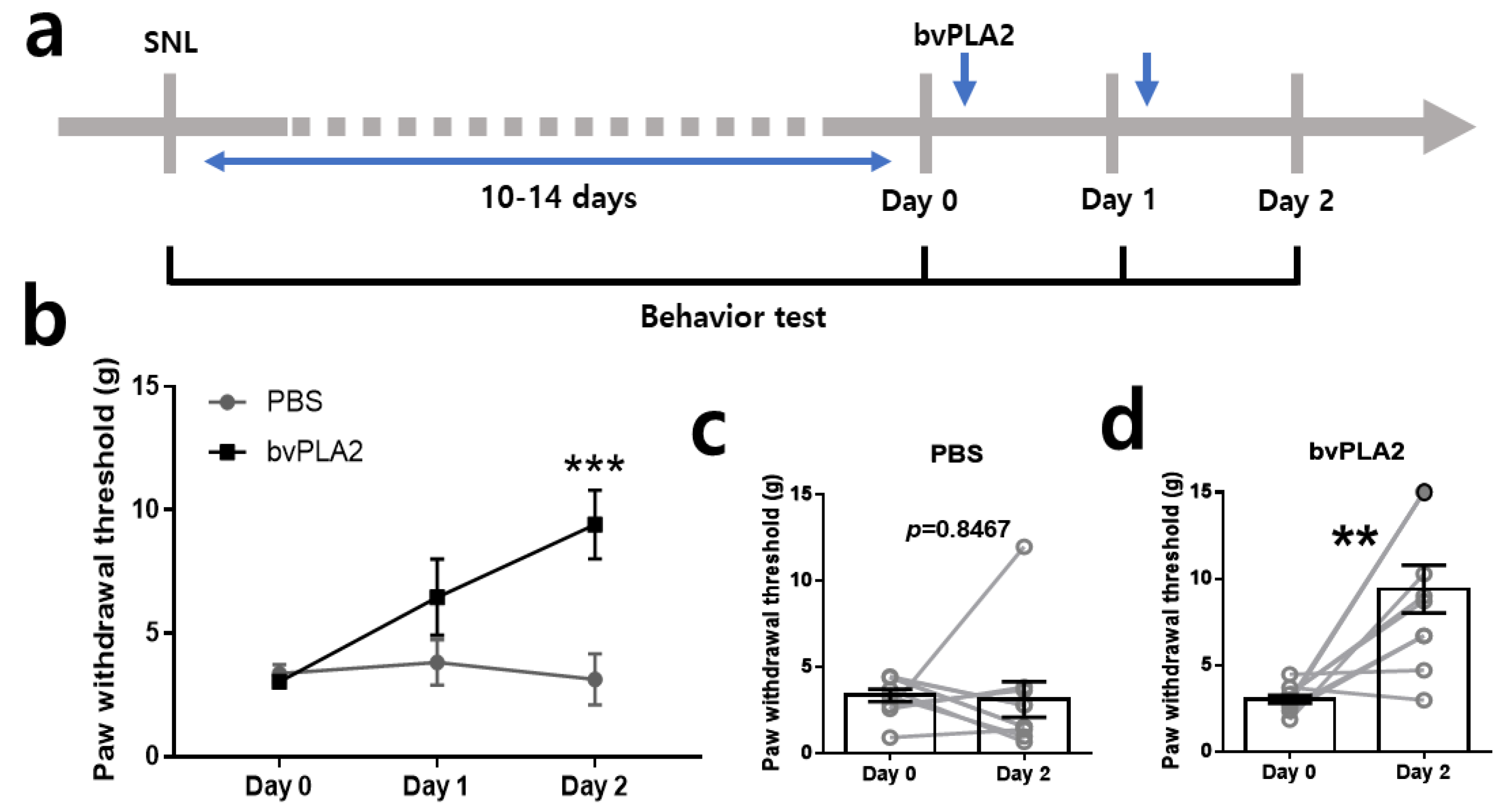
2.2. Repetitive bvPLA2 Treatments Markedly Attenuate Mechanical Allodynia in SNL Rats
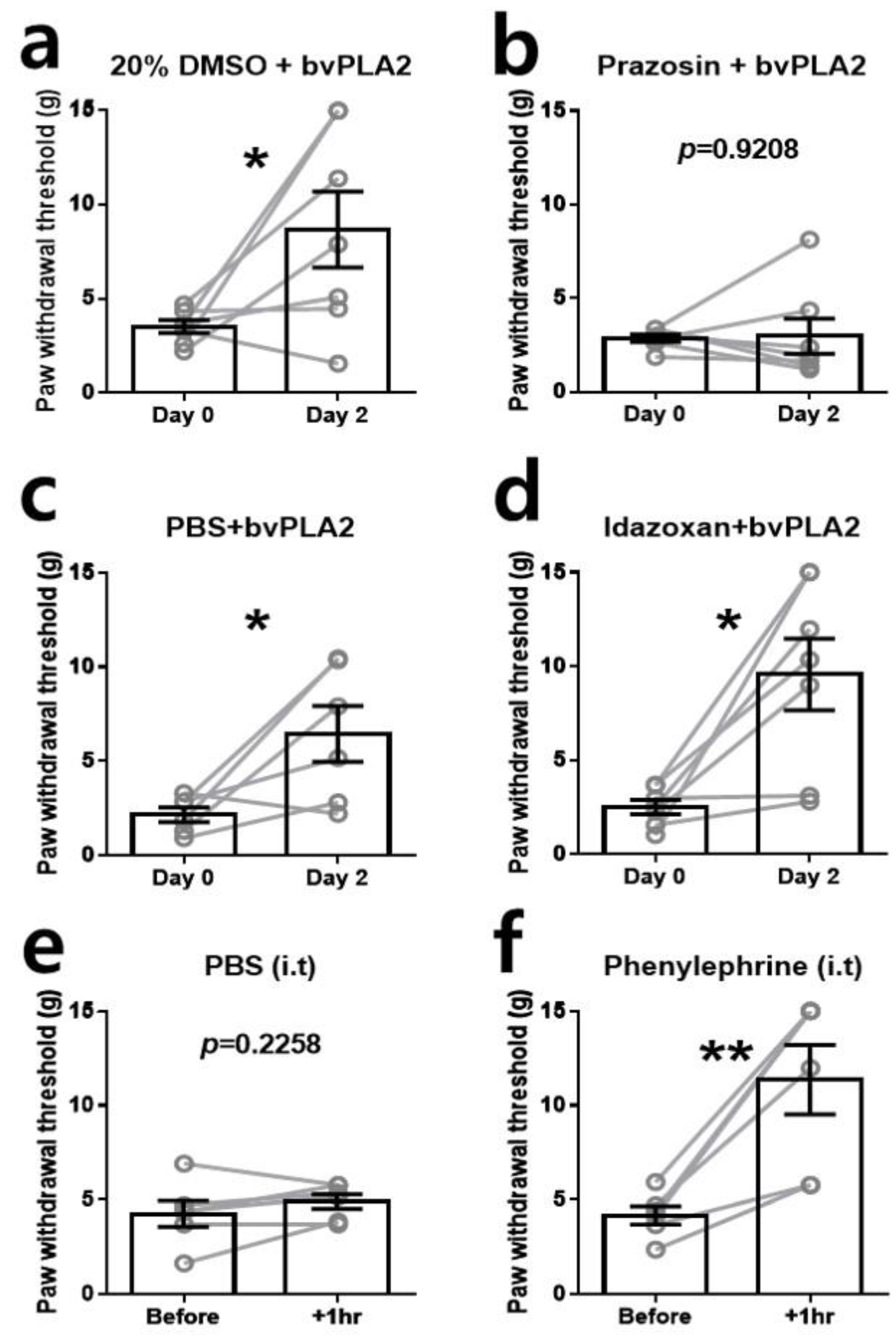
2.3. α1-Adrenergic Receptor (AR) Is Involved in the Antiallodynic Effects of bvPLA2
2.4. A Single Injection of α1-AR Agonist Ameliorates Mechanical Allodynia
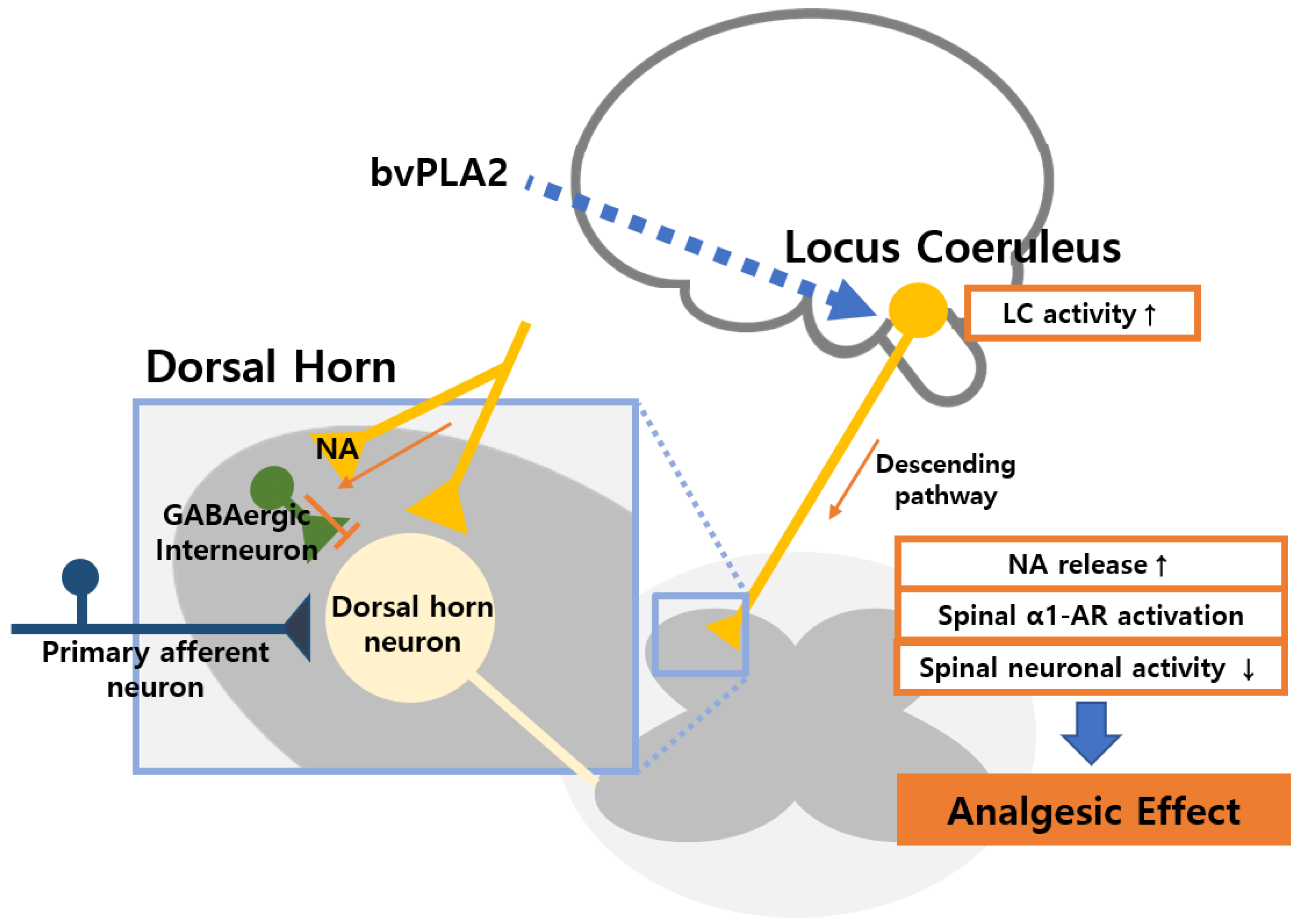
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Spinal Nerve Ligation
4.3. Behavioral Test
4.4. bvPLA2 Administration
4.5. Adrenergic Antagonist Administration
4.6. Adrenergic Agonist Administration
4.7. Statistical Analysis
Author Contributions
Funding
Conflicts of Interest
References
- Colloca, L.; Ludman, T.; Bouhassira, D.; Baron, R.; Dickenson, A.H.; Yarnitsky, D.; Freeman, R.; Truini, A.; Attal, N.; Finnerup, N.B.; et al. Neuropathic pain. Nat. Rev. Dis. Primers 2017, 3, 17002. [Google Scholar] [CrossRef] [PubMed]
- Kessler, T.L. Treatments for neuropathic pain. Lung Cancer 2018, 15, 05. [Google Scholar]
- Hwang, D.-S.; Kim, S.K.; Bae, H. Therapeutic Effects of Bee Venom on Immunological and Neurological Diseases. Toxins 2015, 7, 2413–2421. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.W.; Kwon, Y.B.; Ham, T.W.; Roh, D.H.; Yoon, S.Y.; Lee, H.J.; Han, H.J.; Yang, I.S.; Beitz, A.J.; Lee, J.H. Acupoint stimulation using bee venom attenuates formalin-induced pain behavior and spinal cord fos expression in rats. J. Vet. Med. Sci. 2003, 65, 349–355. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.; Jeon, C.; Lee, J.H. Suppressive Effects of Bee Venom Acupuncture on Paclitaxel-Induced Neuropathic Pain in Rats: Mediation by Spinal alpha(2)-Adrenergic Receptor. Toxins 2017, 9, 351. [Google Scholar] [CrossRef]
- Kang, S.Y.; Roh, D.H.; Yoon, S.Y.; Moon, J.Y.; Kim, H.W.; Lee, H.J.; Beitz, A.J.; Lee, J.H. Repetitive treatment with diluted bee venom reduces neuropathic pain via potentiation of locus coeruleus noradrenergic neuronal activity and modulation of spinal NR1 phosphorylation in rats. J. Pain Off. J. Am. Pain Soc. 2012, 13, 155–166. [Google Scholar] [CrossRef]
- Sobotka, A.K.; Franklin, R.M.; Adkinson, N.F., Jr.; Valentine, M.; Baer, H.; Lichtenstein, L.M. Allergy to insect stings. II. Phospholipase A: the major allergen in honeybee venom. J. Allergy Clin. Immunol. 1976, 57, 29–40. [Google Scholar] [CrossRef]
- Jung, K.H.; Baek, H.; Shin, D.; Lee, G.; Park, S.; Lee, S.; Choi, D.; Kim, W.; Bae, H. Protective Effects of Intratracheally-Administered Bee Venom Phospholipase A2 on Ovalbumin-Induced Allergic Asthma in Mice. Toxins 2016, 8, 269. [Google Scholar] [CrossRef]
- Park, S.; Baek, H.; Jung, K.H.; Lee, G.; Lee, H.; Kang, G.H.; Lee, G.; Bae, H. Bee venom phospholipase A2 suppresses allergic airway inflammation in an ovalbumin-induced asthma model through the induction of regulatory T cells. Immun. Inflamm. Dis. 2015, 3, 386–397. [Google Scholar] [CrossRef]
- Jung, K.H.; Baek, H.; Kang, M.; Kim, N.; Lee, S.Y.; Bae, H. Bee Venom Phospholipase A2 Ameliorates House Dust Mite Extract Induced Atopic Dermatitis Like Skin Lesions in Mice. Toxins 2017, 9, 68. [Google Scholar] [CrossRef]
- Lee, G.; Bae, H. Bee Venom Phospholipase A2: Yesterday’s Enemy Becomes Today’s Friend. Toxins 2016, 8, 48. [Google Scholar] [CrossRef]
- Hossen, M.S.; Shapla, U.M.; Gan, S.H.; Khalil, M.I. Impact of Bee Venom Enzymes on Diseases and Immune Responses. Molecules 2016, 22, 25. [Google Scholar] [CrossRef]
- Zambelli, V.O.; Picolo, G.; Fernandes, C.A.H.; Fontes, M.R.M.; Cury, Y. Secreted Phospholipases A(2) from Animal Venoms in Pain and Analgesia. Toxins 2017, 9, 406. [Google Scholar] [CrossRef]
- Li, D.; Kim, W.; Shin, D.; Jung, Y.; Bae, H.; Kim, S.K. Preventive Effects of Bee Venom Derived Phospholipase A(2) on Oxaliplatin-Induced Neuropathic Pain in Mice. Toxins 2016, 8, 27. [Google Scholar] [CrossRef]
- Li, D.; Lee, Y.; Kim, W.; Lee, K.; Bae, H.; Kim, S.K. Analgesic Effects of Bee Venom Derived Phospholipase A(2) in a Mouse Model of Oxaliplatin-Induced Neuropathic Pain. Toxins 2015, 7, 2422–2434. [Google Scholar] [CrossRef]
- Ossipov, M.H.; Morimura, K.; Porreca, F. Descending pain modulation and chronification of pain. Curr. Opin. Support. Palliat. Care 2014, 8, 143. [Google Scholar]
- Bannister, K.; Dickenson, A.H. What do monoamines do in pain modulation? Curr. Opin. Support. Palliat. Care 2016, 10, 143–148. [Google Scholar] [CrossRef]
- Han, Y.; Smith, M.T. Pathobiology of cancer chemotherapy-induced peripheral neuropathy (CIPN). Front. Pharmacol. 2013, 4, 156. [Google Scholar] [CrossRef]
- Jaggi, A.S.; Jain, V.; Singh, N. Animal models of neuropathic pain. Fundam. Clin. Pharmacol. 2011, 25, 1–28. [Google Scholar] [CrossRef]
- Chung, G.; Kim, C.Y.; Yun, Y.-C.; Yoon, S.H.; Kim, M.-H.; Kim, Y.K.; Kim, S.J. Upregulation of prefrontal metabotropic glutamate receptor 5 mediates neuropathic pain and negative mood symptoms after spinal nerve injury in rats. Sci. Rep. 2017, 7, 9743. [Google Scholar] [CrossRef]
- Kim, S.H.; Chung, J.M. An experimental model for peripheral neuropathy produced by segmental spinal nerve ligation in the rat. Pain 1992, 50, 355–363. [Google Scholar] [CrossRef]
- Urban, M.O.; Ren, K.; Park, K.T.; Campbell, B.; Anker, N.; Stearns, B.; Aiyar, J.; Belley, M.; Cohen, C.; Bristow, L. Comparison of the Antinociceptive Profiles of Gabapentin and 3-Methylgabapentin in Rat Models of Acute and Persistent Pain: Implications for Mechanism of Action. J. Pharmacol. Exp. Ther. 2005, 313, 1209–1216. [Google Scholar] [CrossRef]
- Sullivan, N.R.; Leventhal, L.; Harrison, J.; Smith, V.A.; Cummons, T.A.; Spangler, T.B.; Sun, S.-C.; Lu, P.; Uveges, A.J.; Strassle, B.W.; et al. Pharmacological Characterization of the Muscarinic Agonist (3R,4R)-3-(3-Hexylsulfanyl-pyrazin-2-yloxy)-1-aza-bicyclo[2.2.1]heptane (WAY-132983) in in Vitro and in Vivo Models of Chronic Pain. J. Pharmacol. Exp. Ther. 2007, 322, 1294–1304. [Google Scholar] [CrossRef]
- Kim, W.; Kim, M.J.; Go, D.; Min, B.I.; Na, H.S.; Kim, S.K. Combined Effects of Bee Venom Acupuncture and Morphine on Oxaliplatin-Induced Neuropathic Pain in Mice. Toxins 2016, 8, 33. [Google Scholar] [CrossRef]
- Huh, J.E.; Seo, B.K.; Lee, J.W.; Park, Y.C.; Baek, Y.H. Analgesic Effects of Diluted Bee Venom Acupuncture Mediated by delta-Opioid and alpha2-Adrenergic Receptors in Osteoarthritic Rats. Altern. Ther. Health Med. 2018, 24, 28–35. [Google Scholar]
- Kim, H.W.; Kwon, Y.B.; Han, H.J.; Yang, I.S.; Beitz, A.J.; Lee, J.H. Antinociceptive mechanisms associated with diluted bee venom acupuncture (apipuncture) in the rat formalin test: involvement of descending adrenergic and serotonergic pathways. Pharmacol. Res. 2005, 51, 183–188. [Google Scholar] [CrossRef]
- Lim, B.S.; Moon, H.J.; Li, D.X.; Gil, M.; Min, J.K.; Lee, G.; Bae, H.; Kim, S.K.; Min, B.I. Effect of bee venom acupuncture on oxaliplatin-induced cold allodynia in rats. Evid.-Based Complementary Altern. Med. 2013, 2013, 369324. [Google Scholar] [CrossRef]
- Yeo, J.H.; Yoon, S.Y.; Kwon, S.K.; Kim, S.J.; Lee, J.H.; Beitz, A.J.; Roh, D.H. Repetitive Acupuncture Point Treatment with Diluted Bee Venom Relieves Mechanical Allodynia and Restores Intraepidermal Nerve Fiber Loss in Oxaliplatin-Induced Neuropathic Mice. J. Pain Off. J. Am. Pain Soc. 2016, 17, 298–309. [Google Scholar] [CrossRef]
- Stone, L.S.; Vulchanova, L.; Riedl, M.S.; Wang, J.; Williams, F.G.; Wilcox, G.L.; Elde, R. Effects of peripheral nerve injury on alpha-2A and alpha-2C adrenergic receptor immunoreactivity in the rat spinal cord. Neuroscience 1999, 93, 1399–1407. [Google Scholar] [CrossRef]
- Xie, J.; Ho Lee, Y.; Wang, C.; Mo Chung, J.; Chung, K. Differential expression of alpha1-adrenoceptor subtype mRNAs in the dorsal root ganglion after spinal nerve ligation. Brain Res. Mol. Brain Res. 2001, 93, 164–172. [Google Scholar] [CrossRef]
- Finch, P.M.; Drummond, E.S.; Dawson, L.F.; Phillips, J.K.; Drummond, P.D. Up-regulation of cutaneous alpha1 -adrenoceptors in complex regional pain syndrome type I. Pain Med. 2014, 15, 1945–1956. [Google Scholar] [CrossRef]
- Di Cesare Mannelli, L.; Micheli, L.; Crocetti, L.; Giovannoni, M.P.; Vergelli, C.; Ghelardini, C. alpha2 Adrenoceptor: a Target for Neuropathic Pain Treatment. Mini Rev. Med. Chem. 2017, 17, 95–107. [Google Scholar] [CrossRef]
- Kingery, W.S.; Agashe, G.S.; Guo, T.Z.; Sawamura, S.; Davies, M.F.; Clark, J.D.; Kobilka, B.K.; Maze, M. Isoflurane and nociception: spinal alpha2A adrenoceptors mediate antinociception while supraspinal alpha1 adrenoceptors mediate pronociception. Anesthesiology 2002, 96, 367–374. [Google Scholar] [CrossRef]
- Dogrul, A.; Coskun, I.; Uzbay, T. The contribution of alpha-1 and alpha-2 adrenoceptors in peripheral imidazoline and adrenoceptor agonist-induced nociception. Anesth. Analg. 2006, 103, 471–477. [Google Scholar] [CrossRef]
- Harasawa, I.; Honda, K.; Tanoue, A.; Shinoura, H.; Ishida, Y.; Okamura, H.; Murao, N.; Tsujimoto, G.; Higa, K.; Kamiya, H.-o. Responses to noxious stimuli in mice lacking α1d-adrenergic receptors. Neuroreport 2003, 14, 1857–1860. [Google Scholar] [CrossRef]
- Tasker, R.; Connell, B.; Yole, M. Systemic injections of alpha-1 adrenergic agonists produce antinociception in the formalin test. Pain 1992, 49, 383. [Google Scholar] [CrossRef]
- Baba, H.; Goldstein, P.A.; Okamoto, M.; Kohno, T.; Ataka, T.; Yoshimura, M.; Shimoji, K. Norepinephrine facilitates inhibitory transmission in substantia gelatinosa of adult rat spinal cord (part 2): effects on somatodendritic sites of GABAergic neurons. Anesthesiology 2000, 92, 485–492. [Google Scholar] [CrossRef]
- Yuan, W.X.; Chen, S.R.; Chen, H.; Pan, H.L. Stimulation of alpha(1)-adrenoceptors reduces glutamatergic synaptic input from primary afferents through GABA(A) receptors and T-type Ca(2+) channels. Neuroscience 2009, 158, 1616–1624. [Google Scholar] [CrossRef]
- Sun, G.Y.; Shelat, P.B.; Jensen, M.B.; He, Y.; Sun, A.Y.; Simonyi, A. Phospholipases A2 and inflammatory responses in the central nervous system. Neuromol. Med. 2010, 12, 133–148. [Google Scholar] [CrossRef]
- Kim, H.; Lee, H.; Lee, G.; Jang, H.; Kim, S.-S.; Yoon, H.; Kang, G.-H.; Hwang, D.-S.; Kim, S.K.; Chung, H.-S. Phospholipase A2 inhibits cisplatin-induced acute kidney injury by modulating regulatory T cells by the CD206 mannose receptor. Kidney Int. 2015, 88, 550–559. [Google Scholar] [CrossRef]
- Chung, E.S.; Lee, G.; Lee, C.; Ye, M.; Chung, H.-S.; Kim, H.; Sung-joo, S.B.; Hwang, D.-S.; Bae, H. Bee venom phospholipase A2, a novel Foxp3+ regulatory T cell inducer, protects dopaminergic neurons by modulating neuroinflammatory responses in a mouse model of Parkinson’s disease. J. Immunol. 2015, 195, 4853–4860. [Google Scholar] [CrossRef]
- Austin, P.J.; Kim, C.F.; Perera, C.J.; Moalem-Taylor, G. Regulatory T cells attenuate neuropathic pain following peripheral nerve injury and experimental autoimmune neuritis. Pain 2012, 153, 1916–1931. [Google Scholar] [CrossRef]
- Lees, J.G.; Duffy, S.S.; Perera, C.J.; Moalem-Taylor, G. Depletion of Foxp3+ regulatory T cells increases severity of mechanical allodynia and significantly alters systemic cytokine levels following peripheral nerve injury. Cytokine 2015, 71, 207–214. [Google Scholar] [CrossRef]
- Dixon, W.J. Staircase bioassay: the up-and-down method. Neurosci. Biobehav. Rev. 1991, 15, 47–50. [Google Scholar] [CrossRef]
- Chaplan, S.R.; Bach, F.W.; Pogrel, J.W.; Chung, J.M.; Yaksh, T.L. Quantitative assessment of tactile allodynia in the rat paw. J. Neurosci. Methods 1994, 53, 55–63. [Google Scholar] [CrossRef]
- Ye, M.; Chung, H.S.; Lee, C.; Yoon, M.S.; Yu, A.R.; Kim, J.S.; Hwang, D.S.; Shim, I.; Bae, H. Neuroprotective effects of bee venom phospholipase A2 in the 3xTg AD mouse model of Alzheimer’s disease. J. Neuroinflamm. 2016, 13, 10. [Google Scholar] [CrossRef]
- Ye, M.; Chung, H.-S.; Lee, C.; Hyun Song, J.; Shim, I.; Kim, Y.-S.; Bae, H. Bee venom phospholipase A2 ameliorates motor dysfunction and modulates microglia activation in Parkinson’s disease alpha-synuclein transgenic mice. Exp. Mol. Med. 2016, 48, e244. [Google Scholar] [CrossRef]




© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Woo, S.; Chung, G.; Bae, H.; Kim, S.K. Suppressive Effects of Bee Venom-Derived Phospholipase A2 on Mechanical Allodynia in a Rat Model of Neuropathic Pain. Toxins 2019, 11, 477. https://doi.org/10.3390/toxins11080477
Woo S, Chung G, Bae H, Kim SK. Suppressive Effects of Bee Venom-Derived Phospholipase A2 on Mechanical Allodynia in a Rat Model of Neuropathic Pain. Toxins. 2019; 11(8):477. https://doi.org/10.3390/toxins11080477
Chicago/Turabian StyleWoo, Seunghui, Geehoon Chung, Hyunsu Bae, and Sun Kwang Kim. 2019. "Suppressive Effects of Bee Venom-Derived Phospholipase A2 on Mechanical Allodynia in a Rat Model of Neuropathic Pain" Toxins 11, no. 8: 477. https://doi.org/10.3390/toxins11080477
APA StyleWoo, S., Chung, G., Bae, H., & Kim, S. K. (2019). Suppressive Effects of Bee Venom-Derived Phospholipase A2 on Mechanical Allodynia in a Rat Model of Neuropathic Pain. Toxins, 11(8), 477. https://doi.org/10.3390/toxins11080477