Investigation of Zearalenone Adsorption and Biotransformation by Microorganisms Cultured under Cellular Stress Conditions


 , , ,
, , , 
Abstract
1. Introduction
2. Results
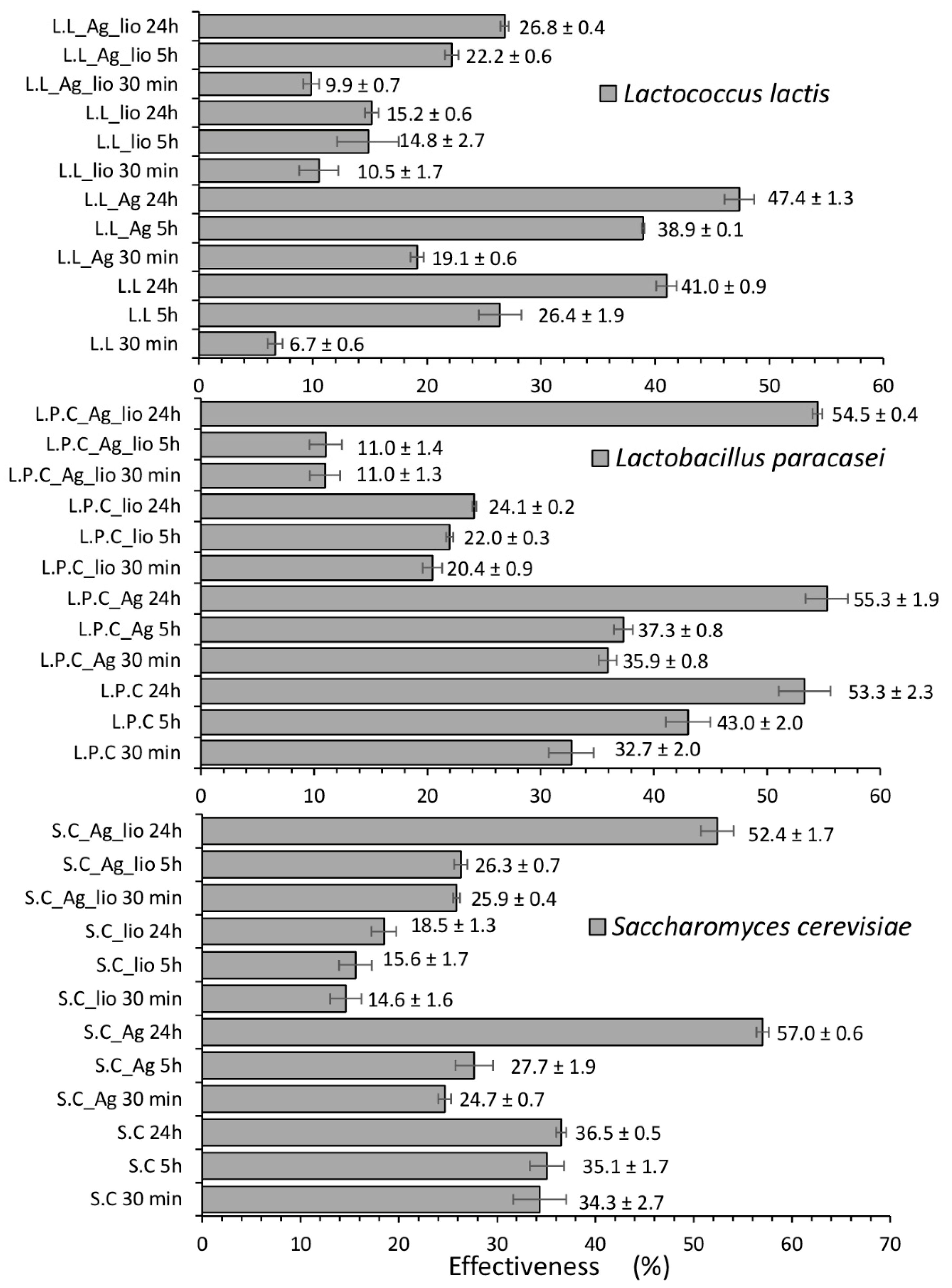
2.1. Influence of Culture Conditions on Zearalenone Neutralization and Metabolization Process
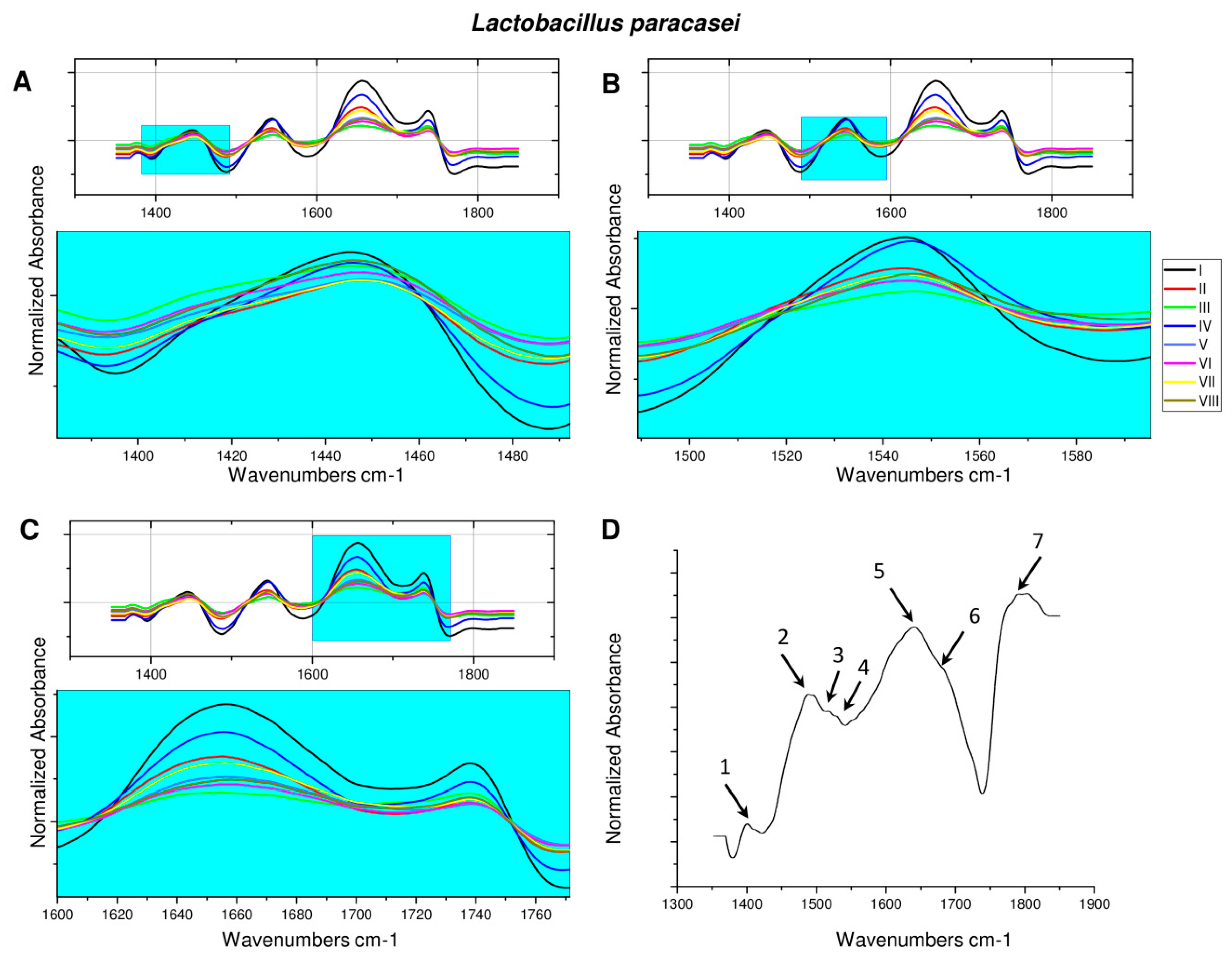
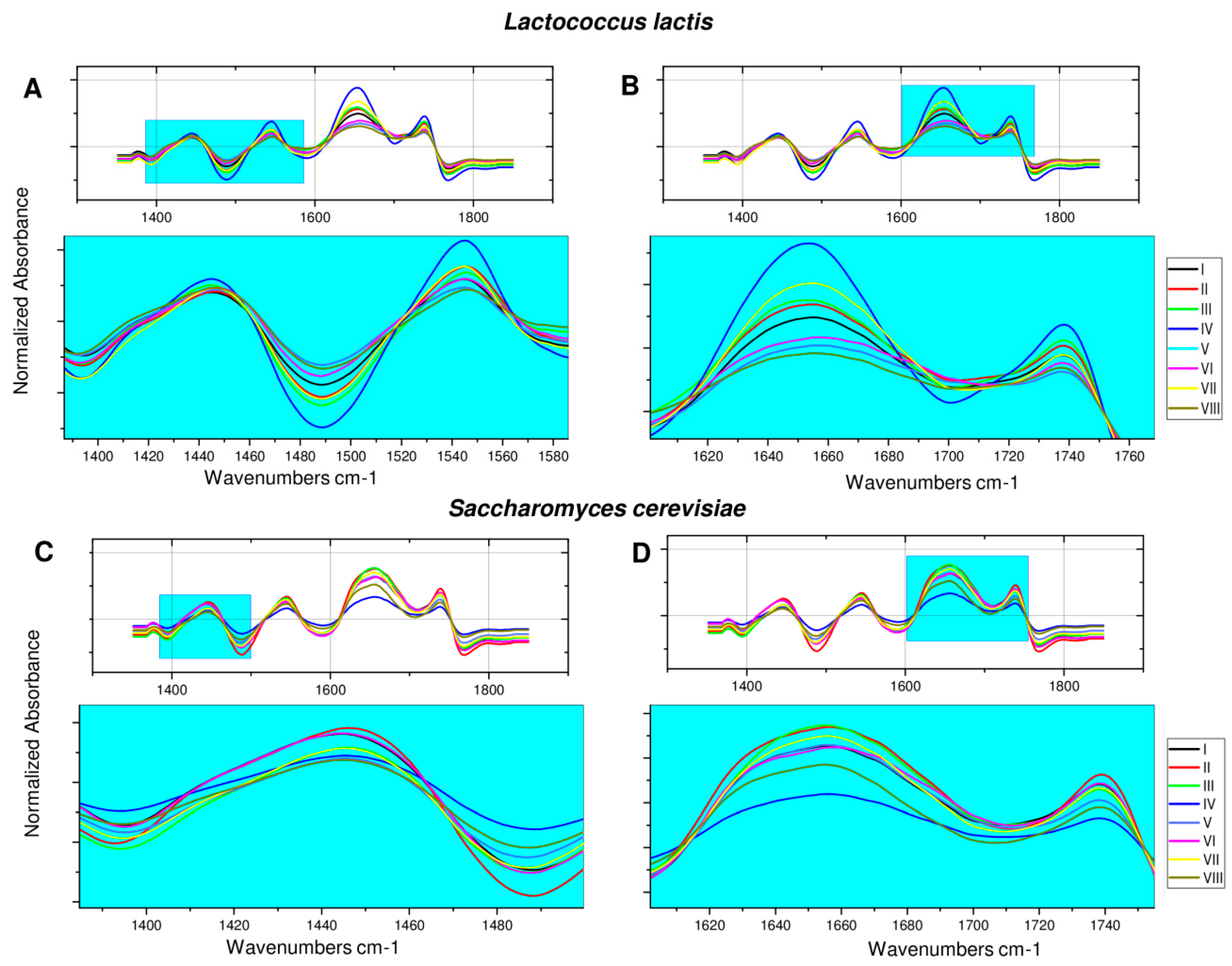
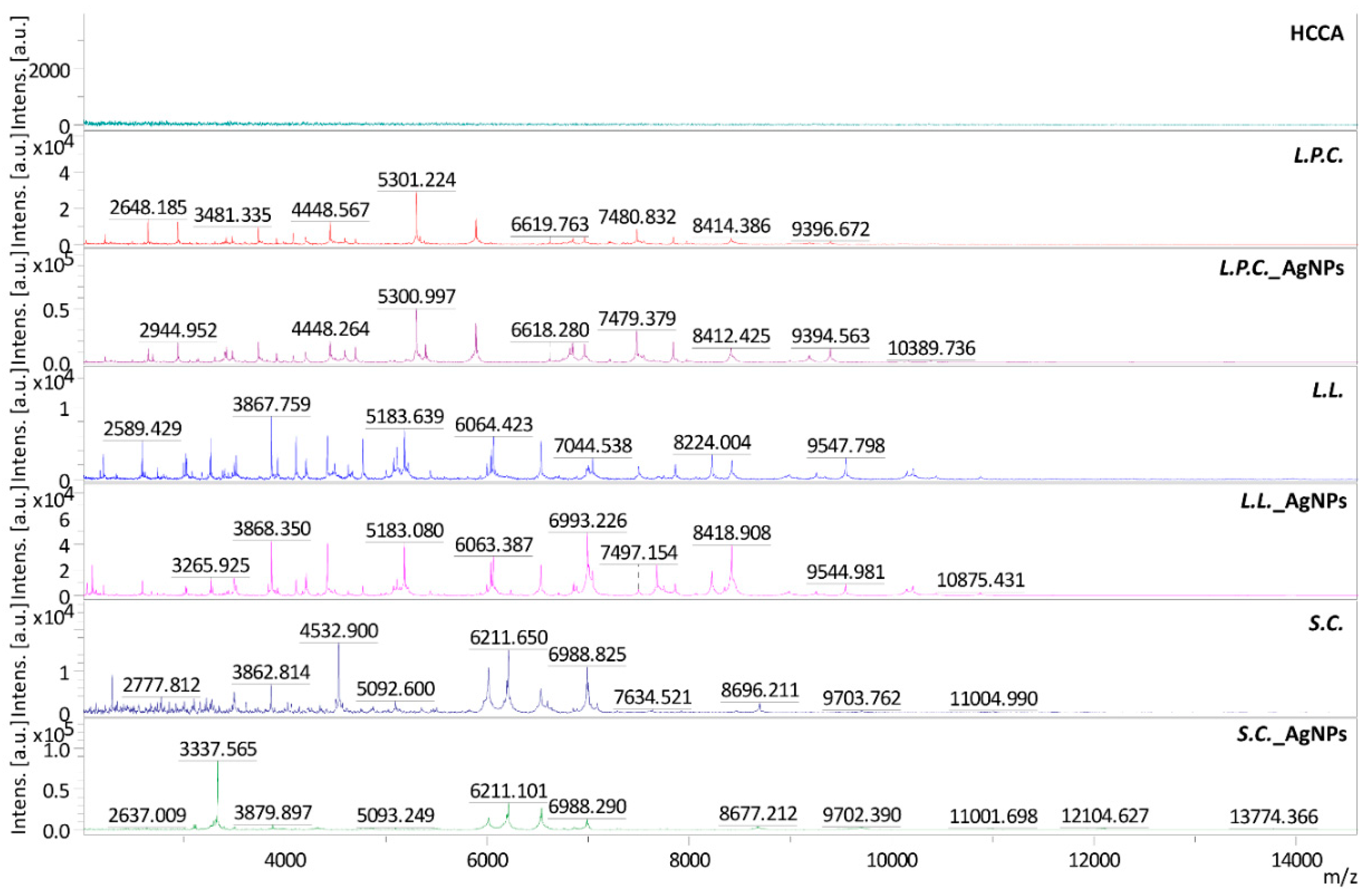
2.2. Fourier Transform Infrared Spectroscopy Analysis of Microorganisms after ZEA Neutralization
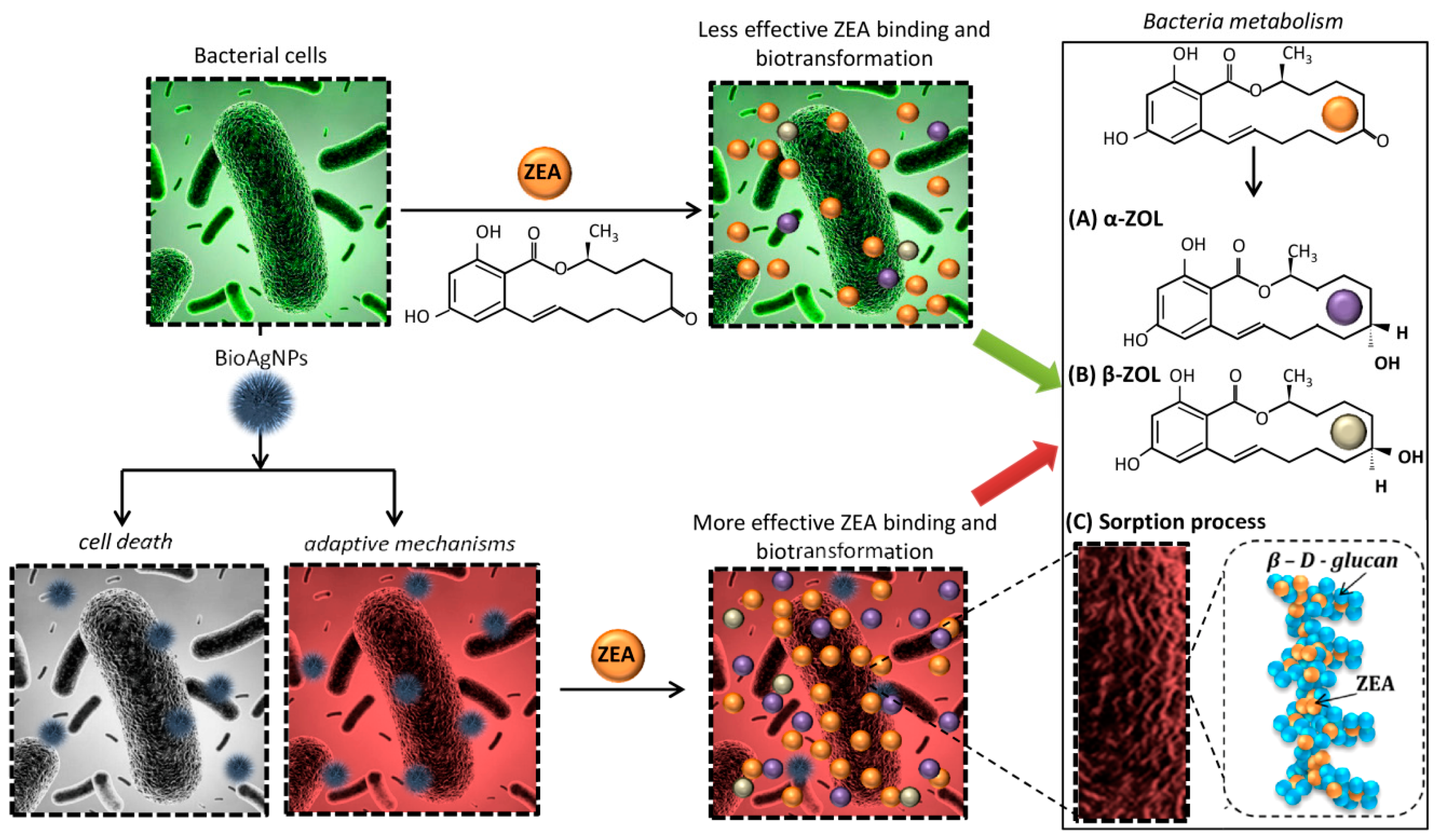
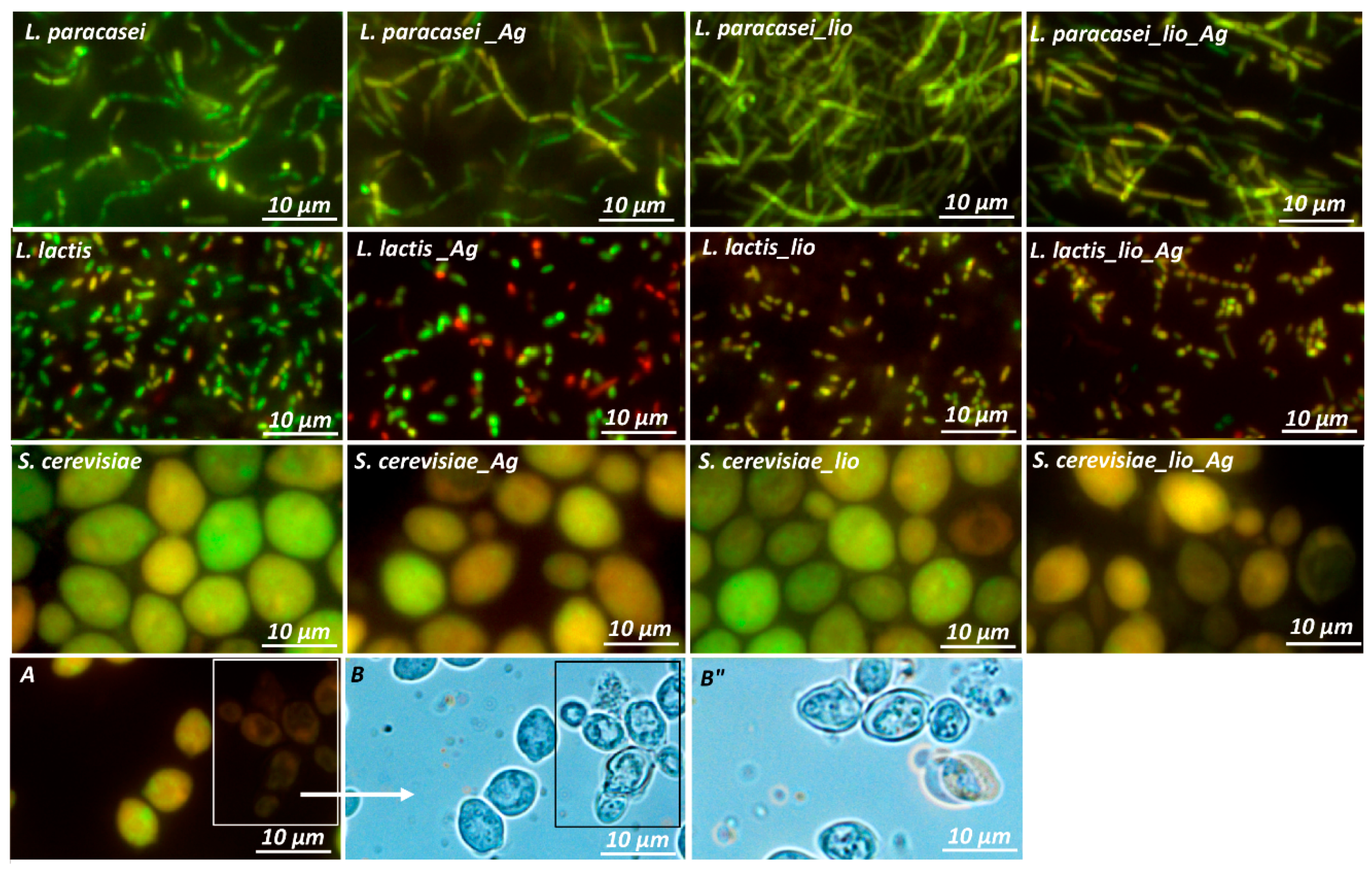
2.3. Changes in Morphology and Metabolism of Cells Cultured in Different Conditions
3. Discussion
4. Materials and Methods
4.1. Chemicals and Reagents
4.2. Microorganisms
4.3. Culturing of Microorganisms under Cellular Stress Conditions
4.4. Zearalenone Neutralization and Metabolization
4.5. High-Performance Liquid Chromatography Analysis
4.6. FT-IR Spectroscopic Analysis
4.7. Microscopic Analysis
4.8. Matrix-Assisted Laser Desorption/Ionization with Mass Spectrometry Analysis
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Appell, M.; Wang, L.C.; Bosma, W.B. Analysis of the photophysical properties of zearalenone using density functional theory. J. Lumin. 2017, 188, 551–557. [Google Scholar] [CrossRef]
- De Boevre, M.; Di Mavungu, J.D.; Landschoot, S.; Audenaert, K.; Eeckhout, M.; Maene, P.; Haesaert, G.; De Saeger, S. Natural occurrence of mycotoxins and their masked forms in food and feed products. World Mycotoxin J. 2012, 5, 207–219. [Google Scholar] [CrossRef]
- Zinedine, A.; Soriano, J.M.; Moltó, J.C.; Mañes, J. Review on the toxicity, occurrence, metabolism, detoxification, regulations and intake of zearalenone: An oestrogenic mycotoxin. Food Chem. Toxicol. 2007, 45, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Bai, X.; Sun, C.; Xu, J.; Liu, D.; Han, Y.; Wu, S.; Luo, X. Detoxification of zearalenone from corn oil by adsorption of functionalized GO systems. Appl. Surf. Sci. 2018, 430, 198–207. [Google Scholar] [CrossRef]
- Zhang, H.; Dong, M.; Yang, Q.; Apaliya, M.T.; Li, J.; Zhang, X. Biodegradation of zearalenone by Saccharomyces cerevisiae: Possible involvement of ZEN responsive proteins of the yeast. J. Proteomics 2016, 143, 416–423. [Google Scholar] [CrossRef] [PubMed]
- Keller, L.; Abrunhosa, L.; Keller, K.; Rosa, C.A.; Cavaglieri, L.; Venâncio, A. Zearalenone and its derivatives α-zearalenol and β-zearalenol decontamination by Saccharomyces cerevisiae strains isolated from bovine forage. Toxins 2015, 7, 3297–3308. [Google Scholar] [CrossRef] [PubMed]
- Rogowska, A.; Pomastowski, P.; Sagandykova, G.; Buszewski, B. Zearalenone and its metabolites: Effect on human health, metabolism and neutralisation methods. Toxicon 2019, 162, 46–56. [Google Scholar] [CrossRef] [PubMed]
- Minervini, F.; Giannoccaro, A.; Cavallini, A.; Visconti, A. Investigations on cellular proliferation induced by zearalenone and its derivatives in relation to the estrogenic parameters. Toxicol. Lett. 2005, 159, 272–283. [Google Scholar] [CrossRef]
- Dellafiora, L.; Ruotolo, R.; Perotti, A.; Cirlini, M.; Galaverna, G.; Cozzini, P.; Buschini, A.; Dall’Asta, C. Molecular insights on xenoestrogenic potential of zearalenone-14-glucoside through a mixed in vitro/in silico approach. Food Chem. Toxicol. 2017, 108, 257–266. [Google Scholar] [CrossRef]
- Bennett, J.W.; Klich, M. Mycotoxins. Clin. Microbiol. Rev. 2003, 16, 497–516. [Google Scholar] [CrossRef]
- Bronowicka-Kłys, D.E.; Lianeri, M.; Jagodziński, P.P. The role and impact of estrogens and xenoestrogen on the development of cervical cancer. Biomed. Pharmacother. 2016, 84, 1945–1953. [Google Scholar] [CrossRef] [PubMed]
- Schoevers, E.J.; Santos, R.R.; Colenbrander, B.; Fink-Gremmels, J.; Roelen, B.A.J. Transgenerational toxicity of Zearalenone in pigs. Reprod. Toxicol. 2012, 34, 110–119. [Google Scholar] [CrossRef] [PubMed]
- Hueza, I.M.; Raspantini, P.C.F.; Raspantini, L.E.R.; Latorre, A.O.; Górniak, S.L. Zearalenone, an estrogenic mycotoxin, is an immunotoxic compound. Toxins 2014, 6, 1080–1095. [Google Scholar] [CrossRef] [PubMed]
- Mokoena, M.P.; Chelule, P.K.; Gqaleni, N. Reduction of fumonisin B1 and zearalenone by lactic acid bacteria in fermented maize meal. J. Food Prot. 2005, 68, 2095–2099. [Google Scholar] [CrossRef] [PubMed]
- Shier, W.T.; Shier, A.C.; Xie, W.; Mirocha, C.J. Structure-activity relationships for human estrogenic activity in zearalenone mycotoxins. Toxicon 2001, 39, 1435–1438. [Google Scholar] [CrossRef]
- Shetty, P.H.; Jespersen, L. Saccharomyces cerevisiae and lactic acid bacteria as potential mycotoxin decontaminating agents. Trends Food Sci. Technol. 2006, 17, 48–55. [Google Scholar] [CrossRef]
- Niderkorn, V.; Morgavi, D.P.; Pujos, E.; Tissandier, A.; Boudra, H. Screening of fermentative bacteria for their ability to bind and biotransform deoxynivalenol, zearalenone and fumonisins in an in vitro simulated corn silage model. Food Addit. Contam. 2007, 24, 406–415. [Google Scholar] [CrossRef]
- El-Nezami, H.; Polychronaki, N.; Salminen, S.; Mykkänen, H. Binding rather than metabolism may explain the interaction of two food-Grade Lactobacillus strains with zearalenone and its derivative ά-zearalenol. Appl. Environ. Microbiol. 2002, 68, 3545–3549. [Google Scholar] [CrossRef]
- Vega, M.F.; Dieguez, S.N.; Riccio, B.; Aranguren, S.; Giordano, A.; Denzoin, L.; Soraci, A.L.; Tapia, M.O.; Ross, R.; Apás, A.; et al. Zearalenone adsorption capacity of lactic acid bacteria isolated from pigs. Braz. J. Microbiol. 2017, 48, 715–723. [Google Scholar] [CrossRef]
- Taheur, F.B.; Fedhila, K.; Chaieb, K.; Kouidhi, B.; Bakhrouf, A.; Abrunhosa, L. Adsorption of aflatoxin B1, zearalenone and ochratoxin A by microorganisms isolated from Kefir grains. Int. J. Food Microbiol. 2017, 251, 1–7. [Google Scholar] [CrossRef]
- Król, A.; Pomastowski, P.; Rafińska, K.; Railean-Plugaru, V.; Walczak, J.; Buszewski, B. Microbiology neutralization of zearalenone using Lactococcus lactis and Bifidobacterium sp. Anal. Bioanal. Chem. 2018, 410, 943–952. [Google Scholar] [CrossRef] [PubMed]
- Ghofrani Tabari, D.; Kermanshahi, H.; Golian, A.; Majidzadeh Heravi, R. In Vitro Binding Potentials of Bentonite, Yeast Cell Wall and Lactic Acid Bacteria for Aflatoxin B1 and Ochratoxin A. Iran. J. Toxicol. 2018, 12, 7–13. [Google Scholar] [CrossRef]
- Slaveykova, V.; Sonntag, B.; Gutiérrez, J.C. Stress and Protists: No life without stress. Eur. J. Protistol. 2016, 55, 39–49. [Google Scholar] [CrossRef] [PubMed]
- Grogan, D.W. Physiology of Prokaryotic Cells. In Cell Physiology Source Book; Elsevier: London, UK, 2012; pp. 891–906. [Google Scholar]
- Boor, K.J. Bacterial Stress Responses: What Doesn’t Kill Them Can Make Them Stronger. PLoS Biol. 2006, 4, e23. [Google Scholar] [CrossRef]
- Andersson, S.G.E. Stress management strategies in single bacterial cells. Proc. Natl. Acad. Sci. USA 2016, 113, 3921–3923. [Google Scholar] [CrossRef] [PubMed]
- Morano, K.A.; Grant, C.M.; Moye-Rowley, W.S.; Van Dyk, T.K.; Ficke, M.B.; Golin, J. The response to heat shock and oxidative stress in Saccharomyces cerevisiae. Genetics 2012, 190, 1157–1195. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Yang, Q.; Li, J.; Gao, H.; Li, P.; Zhou, H. The impact of temperature on microbial diversity and AOA activity in the Tengchong Geothermal Field, China. Sci. Rep. 2015, 5, 17056. [Google Scholar] [CrossRef] [PubMed]
- Hechmi, N.; Bosso, L.; El-Bassi, L.; Scelza, R.; Testa, A.; Jedidi, N.; Rao, M.A. Depletion of pentachlorophenol in soil microcosms with Byssochlamys nivea and Scopulariopsis brumptii as detoxification agents. Chemosphere 2016, 165, 547–554. [Google Scholar] [CrossRef]
- Zhang, L.; Chung, J.; Jiang, Q.; Sun, R.; Zhang, J.; Zhong, Y.; Ren, N. Characteristics of rumen microorganisms involved in anaerobic degradation of cellulose at various pH values. RSC Adv. 2017, 7, 40303–40310. [Google Scholar] [CrossRef]
- Zhang, X.-F.; Shen, W.; Gurunathan, S.; Zhang, X.-F.; Shen, W.; Gurunathan, S. Biologically Synthesized Gold Nanoparticles Ameliorate Cold and Heat Stress-Induced Oxidative Stress in Escherichia coli. Molecules 2016, 21, 731. [Google Scholar] [CrossRef]
- Ševců, A.; El-Temsah, Y.S.; Joner, E.J.; Černík, M. Oxidative stress induced in microorganisms by zero-valent iron nanoparticles. Microbes Environ. 2011, 26, 271–281. [Google Scholar] [CrossRef]
- Saccà, M.L.; Fajardo, C.; Martinez-Gomariz, M.; Costa, G.; Nande, M.; Martin, M. Molecular Stress Responses to Nano-Sized Zero-Valent Iron (nZVI) Particles in the Soil Bacterium Pseudomonas stutzeri. PLoS ONE 2014, 9, e89677. [Google Scholar] [CrossRef]
- Patil, M.P.; Seo, Y.B.; Kim, G.-D. Morphological changes of bacterial cells upon exposure of silver-silver chloride nanoparticles synthesized using Agrimonia pilosa. Microb. Pathog. 2018, 116, 84–90. [Google Scholar] [CrossRef]
- Naumann, D.; Keller, S.; Helm, D.; Schultz, C.; Schrader, B. FT-IR spectroscopy and FT-Raman spectroscopy are powerful analytical tools for the non-invasive characterization of intact microbial cells. J. Mol. Struct. 1995, 347, 399–405. [Google Scholar] [CrossRef]
- Movasaghi, Z.; Rehman, S.; ur Rehman, D.I. Fourier Transform Infrared (FTIR) Spectroscopy of Biological Tissues. Appl. Spectrosc. Rev. 2008, 43, 134–179. [Google Scholar] [CrossRef]
- Bejaoui, H.; Mathieu, F.; Taillandier, P.; Lebrihi, A. Ochratoxin A removal in synthetic and natural grape juices by selected oenological Saccharomyces strains. J. Appl. Microbiol. 2004, 97, 1038–1044. [Google Scholar] [CrossRef]
- Wang, L.; Wang, Z.; Yuan, Y.; Cai, R.; Niu, C.; Yue, T. Identification of Key Factors Involved in the Biosorption of Patulin by Inactivated Lactic Acid Bacteria (LAB) Cells. PLoS ONE 2015, 10, e0143431. [Google Scholar] [CrossRef]
- Berner, D.; Viernstein, H. Effect of protective agents on the viability of Lactococcus lactis subjected to freeze-thawing and freeze-drying. Sci. Pharm. 2006, 74, 137–149. [Google Scholar] [CrossRef]
- Bircher, L.; Geirnaert, A.; Hammes, F.; Lacroix, C.; Schwab, C. Effect of cryopreservation and lyophilization on viability and growth of strict anaerobic human gut microbes. Microb. Biotechnol. 2018, 11, 721. [Google Scholar] [CrossRef]
- Wang, J.Q.; Yang, F.; Yang, P.L.; Liu, J.; Lv, Z.H. Microbial reduction of zearalenone by a new isolated Lysinibacillus sp. ZJ-2016-1. World Mycotoxin J. 2018, 11, 571–578. [Google Scholar] [CrossRef]
- Dalié, D.K.D.; Deschamps, A.M.; Richard-Forget, F. Lactic acid bacteria–Potential for control of mould growth and mycotoxins: A review. Food Control 2010, 21, 370–380. [Google Scholar] [CrossRef]
- Bleuven, C.; Landry, C.R. Molecular and cellular bases of adaptation to a changing environment in microorganisms. Proc. R. Soc. B Biol. Sci. 2016, 283, 20161458. [Google Scholar] [CrossRef]
- El-Sharkaway, S.H.; Selim, M.I.; Afifi, M.S.; Halaweish, F.T. Microbial transformation of zearalenone to a zearalenone sulfate. Appl. Environ. Microbiol. 1991, 57, 549–552. [Google Scholar]
- Böswald, C.; Engelhardt, G.; Vogel, H.; Wallnöfer, P.R. Metabolism of the Fusarium mycotoxins zearalenone and deoxynivalenol by yeast strains of technological relevance. Nat. Toxins 1995, 3, 138–144. [Google Scholar] [CrossRef]
- Dalai, S.; Pakrashi, S.; Chakravarty, S.; Hussain, S.; Chandrasekaran, N.; Mukherjee, A. Studies on interfacial interactions of TiO2 nanoparticles with bacterial cells under light and dark conditions. Bull. Mater. Sci. 2014, 37, 371–381. [Google Scholar] [CrossRef]
- Nishitani, K.; Tominaga, R. Endo-xyloglucan transferase, a novel class of glycosyltransferase that catalyzes transfer of a segment of xyloglucan molecule to another xyloglucan molecule. J. Biol. Chem. 1992, 267, 21058–21064. [Google Scholar]
- Colicelli, J. Human RAS Superfamily Proteins and Related GTPases. Sci. STKE 2004, 250, re13. [Google Scholar] [CrossRef]
- Siezen, R.J.; Starrenburg, M.J.C.; Boekhorst, J.; Renckens, B.; Molenaar, D.; van Hylckama Vlieg, J.E.T. Genome-Scale Genotype-Phenotype Matching of Two Lactococcus lactis Isolates from Plants Identifies Mechanisms of Adaptation to the Plant Niche. Appl. Environ. Microbiol. 2008, 74, 424–436. [Google Scholar] [CrossRef]
- Milanowski, M.; Pomastowski, P.; Railean-Plugaru, V.; Rafińska, K.; Ligor, T.; Buszewski, B. Biosorption of silver cations onto Lactococcus lactis and Lactobacillus casei isolated from dairy products. PLoS ONE 2017, 12, e0174521. [Google Scholar] [CrossRef]
- Rogowska, A.; Pomastowski, P.; Złoch, M.; Railean-Plugaru, V.; Król, A.; Rafińska, K.; Szultka-Młyńska, M.; Buszewski, B. The influence of different pH on the electrophoretic behaviour of Saccharomyces cerevisiae modified by calcium ions. Sci. Rep. 2018, 8, 7261. [Google Scholar] [CrossRef]
- Railean-Plugaru, V.; Pomastowski, P.; Meller, K.; Złoch, M.; Rafinska, K.; Buszewski, B. Lactococcus lactis as a safe and inexpensive source of bioactive silver composites. Appl. Microbiol. Biotechnol. 2017, 101, 7141–7153. [Google Scholar]
- Sprynskyy, M.; Gadzała-Kopciuch, R.; Nowak, K.; Buszewski, B. Removal of zearalenone toxin from synthetics gastric and body fluids using talc and diatomite: A batch kinetic study. Colloids Surf. B Biointerfaces 2012, 94, 7–14. [Google Scholar] [CrossRef]
- Bosso, L.; Lacatena, F.; Cristinzio, G.; Cea, M.; Diez, M.C.; Rubilar, O. Biosorption of pentachlorophenol by Anthracophyllum discolor in the form of live fungal pellets. New Biotechnol. 2015, 32, 21–25. [Google Scholar] [CrossRef]
- Daneshvar, E.; Vazirzadeh, A.; Niazi, A.; Sillanpää, M.; Bhatnagar, A. A comparative study of methylene blue biosorption using different modified brown, red and green macroalgae–Effect of pretreatment. Chem. Eng. J. 2017, 307, 435–446. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| CZEA ± SD [μg/mL] | ||||
|---|---|---|---|---|
| t = 0 min | 2.09 ± 0.02 | |||
| Lactococcus lactis | ||||
| Variant | L.L. | L.L._Ag | L.L._lio | L.L._lio_Ag |
| t = 30 min | 1.95 ± 0.01 | 1.69 ± 0.01 | 1.87 ± 0.04 | 1.88 ± 0.01 |
| t = 5 h | 1.54 ± 0.04 | 1.27 ± 0.00 | 1.78 ± 0.06 | 1.63 ± 0.01 |
| t = 24 h | 1.23 ± 0.02 | 1.10 ± 0.03 | 1.77 ± 0.01 | 1.53 ± 0.01 |
| Lactobacillus paracasei | ||||
| Variant | L.P.C. | L.P.C._Ag | L.P.C._lio | L.P.C._lio_Ag |
| t = 30 min | 1.40 ± 0.04 | 1.34 ± 0.02 | 1.66 ± 0.02 | 1.86 ± 0.03 |
| t = 5 h | 1.19 ± 0.04 | 1.31 ± 0.02 | 1.63 ± 0.01 | 1.86 ± 0.03 |
| t = 24 h | 0.97 ± 0.05 | 0.93 ± 0.04 | 1.59 ± 0.00 | 0.95 ± 0.01 |
| Saccharomyces cerevisiae | ||||
| Variant | S.C. | S.C._Ag | S.C._lio | S.C._lio_Ag |
| t = 30 min | 1.37 ± 0.06 | 1.57 ± 0.01 | 1.78 ± 0.03 | 1.55 ± 0.01 |
| t = 5 h | 1.35 ± 0.04 | 1.51 ± 0.04 | 1.76 ± 0.03 | 1.54 ± 0.01 |
| t = 24 h | 1.32 ± 0.01 | 0.90 ± 0.01 | 1.70 ± 0.03 | 1.00 ± 0.04 |
| C α-ZOL ± SD [µg/mL] | C β-ZOL ± SD [µg/mL] | |||||||
|---|---|---|---|---|---|---|---|---|
| Lactococcus lactis | ||||||||
| Variant | L.L. | L.L._Ag | L.L._lio | L.L._lio_Ag | L.L. | L.L._Ag | L.L._lio | L.L._lio_Ag |
| t = 30 min | - | - | - | - | - | - | - | - |
| t = 5 h | - | 0.107 ± 0.004 | - | - | - | 1.097 ± 0.041 | - | - |
| t = 24 h | 0.048 ± 0.002 | 0.262 ± 0.006 | - | 0.066 ± 0.002 | - | 1.464 ± 0.020 | - | 0.356 ± 0.019 |
| Lactobacillus paracasei | ||||||||
| Variant | L.P.C. | L.P.C._Ag | L.P.C._lio | L.P.C._lio_Ag | L.P.C. | L.P.C._Ag | L.P.C._lio | L.P.C._lio_Ag |
| t = 30 min | - | - | - | - | - | - | - | - |
| t = 5 h | - | - | - | - | - | - | - | - |
| t = 24 h | - | 0.142 ± 0.004 | - | 0.190 ± 0.008 | - | 0.699 ± 0.054 | - | 0.310 ± 0.024 |
| Saccharomyces cerevisiae | ||||||||
| Variant | S.C. | S.C._Ag | S.C._lio | S.C._lio_Ag | S.C. | S.C._Ag | S.C._lio | S.C._lio_Ag |
| t = 30 min | - | - | - | - | - | - | - | - |
| t = 5 h | - | - | - | - | 0.034 ± 0.000 | 0.012 ± 0.002 | - | - |
| t = 24 h | - | 0.152 ± 0.002 | - | 0.127 ± 0.004 | 0.102 ± 0.017 | 0.857 ± 0.013 | - | 0.423 ± 0.013 |
| m/z | L. paracasei | m/z | L. lactis | m/z | S. cerevisiae |
|---|---|---|---|---|---|
| 2696 | + | 2044 | + | 2293 | − |
| 3145 | + | 2098 | + | 3337 | + |
| 3825 | + | 2332 | − | 3863 | − |
| 5393 | + | 3084 | − | 3880 | + |
| 6750 | + | 3840 | + | 4324 | + |
| 7347 | − | 6234 | + | 4534 | − |
| 7647 | + | 6857 | + | 5352 | − |
| 8997 | + | 6993 | + | 7091 | − |
| 10168 | + | 7678 | + | 7635 | − |
| 10390 | + | 8348 | + | 8677 | + |
| 8696 | − | ||||
| 11604 | − | ||||
| 12105 | + | ||||
| 13774 | + | ||||
| 14639 | + |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rogowska, A.; Pomastowski, P.; Walczak, J.; Railean-Plugaru, V.; Rudnicka, J.; Buszewski, B. Investigation of Zearalenone Adsorption and Biotransformation by Microorganisms Cultured under Cellular Stress Conditions. Toxins 2019, 11, 463. https://doi.org/10.3390/toxins11080463
Rogowska A, Pomastowski P, Walczak J, Railean-Plugaru V, Rudnicka J, Buszewski B. Investigation of Zearalenone Adsorption and Biotransformation by Microorganisms Cultured under Cellular Stress Conditions. Toxins. 2019; 11(8):463. https://doi.org/10.3390/toxins11080463
Chicago/Turabian StyleRogowska, Agnieszka, Paweł Pomastowski, Justyna Walczak, Viorica Railean-Plugaru, Joanna Rudnicka, and Bogusław Buszewski. 2019. "Investigation of Zearalenone Adsorption and Biotransformation by Microorganisms Cultured under Cellular Stress Conditions" Toxins 11, no. 8: 463. https://doi.org/10.3390/toxins11080463
APA StyleRogowska, A., Pomastowski, P., Walczak, J., Railean-Plugaru, V., Rudnicka, J., & Buszewski, B. (2019). Investigation of Zearalenone Adsorption and Biotransformation by Microorganisms Cultured under Cellular Stress Conditions. Toxins, 11(8), 463. https://doi.org/10.3390/toxins11080463