Novel Polyclonal Antibody Raised against Tetrodotoxin Using Its Haptenic Antigen Prepared from 4,9-anhydrotetrodotoxin Reacted with 1,2-Ethaneditiol and Further Reacted with Keyhole Limpet Hemocyanin


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Reaction of 4,9-anhydroTTX and Dithiol Reagents
2.2. KLH-TTX Conjugate Used for Antigen
2.3. Development of Antibody against TTX
2.4. Conjugate of Biotin and TTX
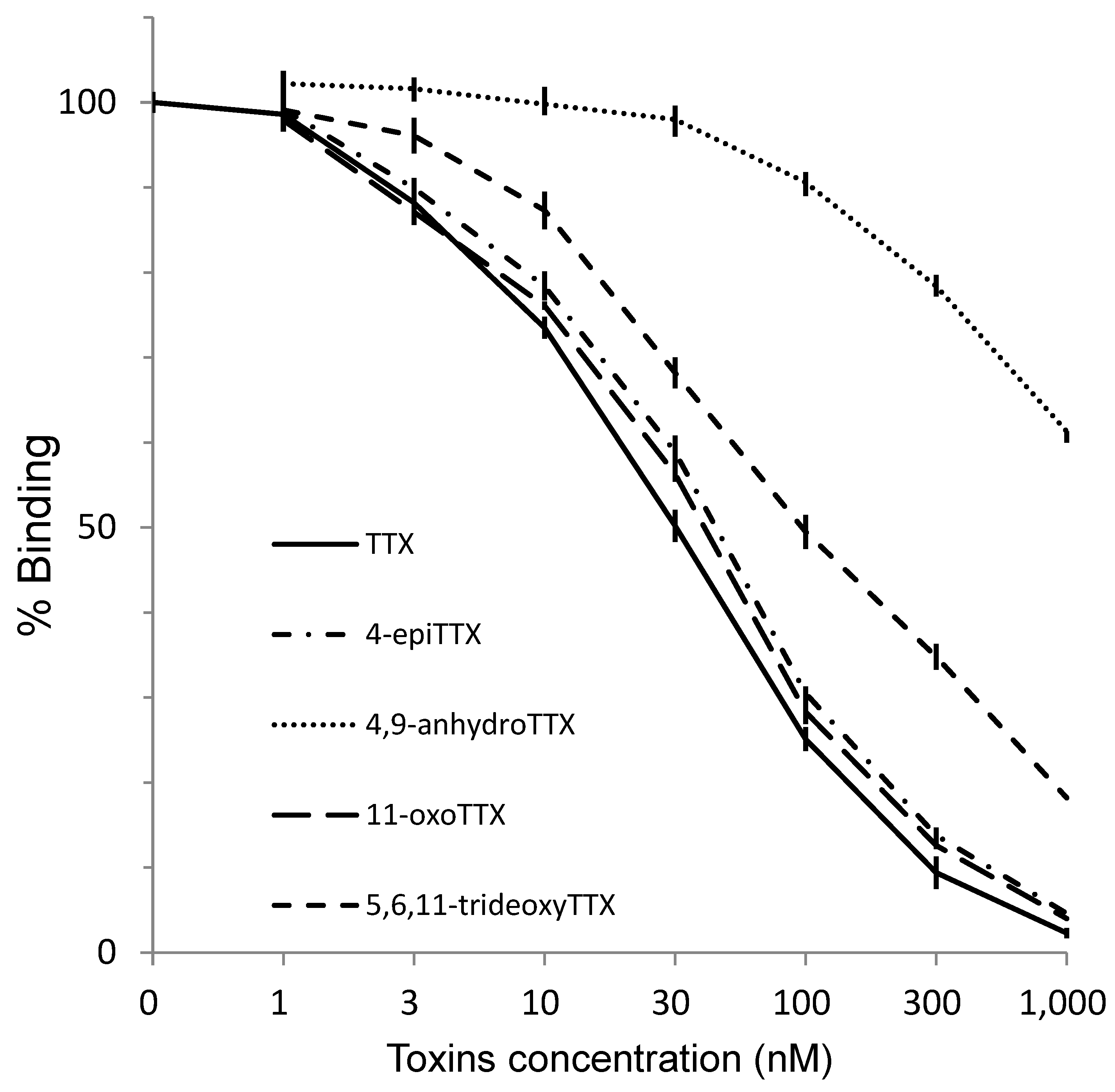
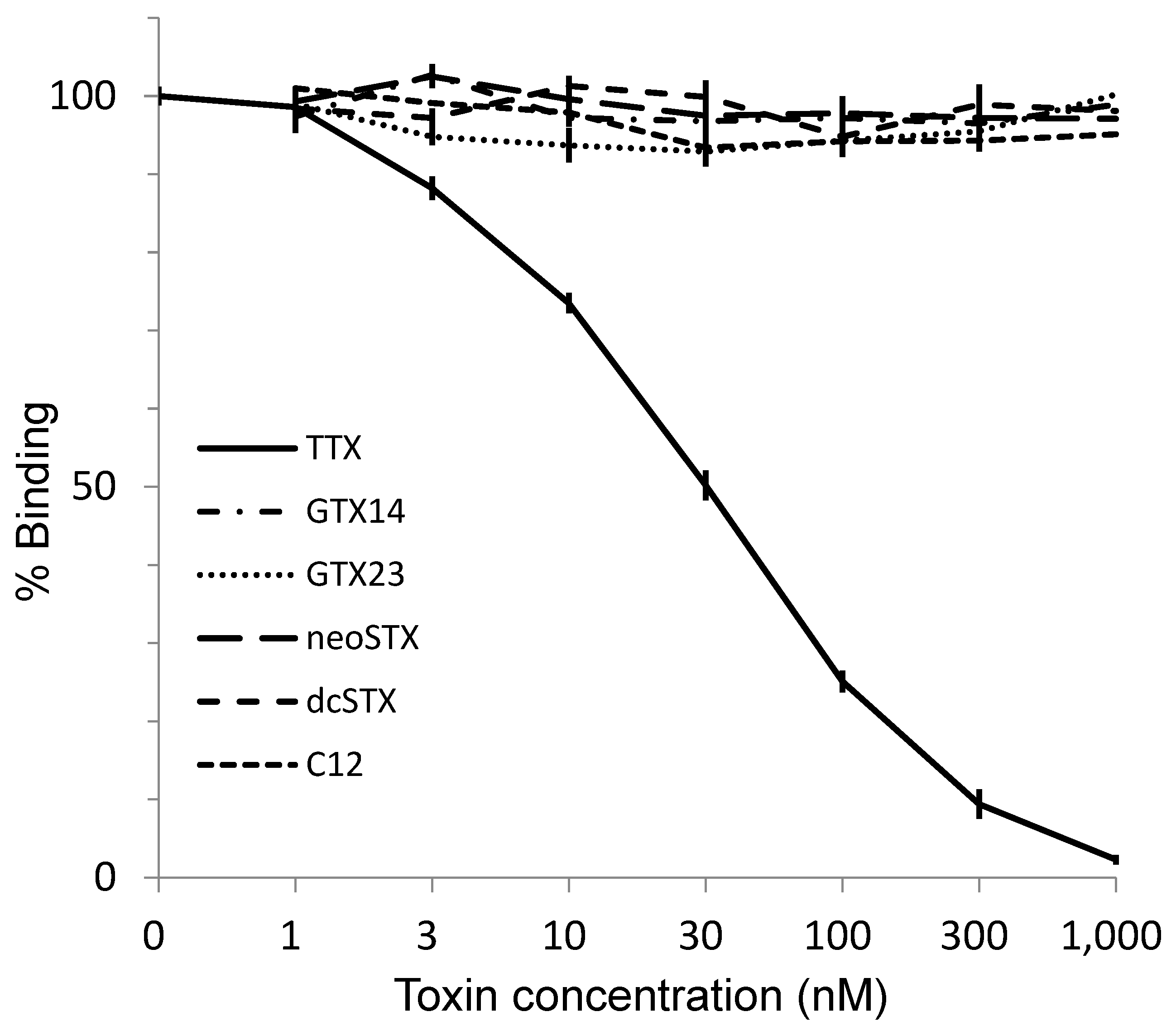
2.5. Reactivity of the Antibody to TTX and Its Analogs and PSTs in ELISA
3. Discussion
4. Conclusions
5. Material and Methods
5.1. Materials
5.2. Preparation of TTX and Its Analogs
5.3. Preparation of the EDT and TTX Adduct
5.4. Analytical Procedures
5.4.1. Confirmation of TTX, 4-epiTTX, and 4,9-anhydroTTX by HPLC-FLD
5.4.2. EDT-TTX Adduct and TTX Coupled to KLH
5.4.3. Confirmation of 5, 6,11-trideoxyTTX and Labeled Toxin by LC-qTOFMS
5.4.4. Inoculation of the Haptenic Antigen to Rabbits
5.4.5. Monitoring the Antibody Activity
5.5. Preparation of Biotin-TTX as a Competitive Labeled Toxin for ELISA
5.6. Preparation of Polyclonal Antibody
5.6.1. Preparation of TTX-coupled Affinity Resin
5.6.2. Purification of Polyclonal Antibody
5.6.3. Direct One Step ELISA for TTX and Its Analogs
5.7. Cross-Reactivity with Paralytic Shellfish Toxins
5.8. Ethics Statement
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Narahashi, T.; Moore, J.W.; Posto, R.N. Tetrodotoxin derivatives: Chemical structure and blockage of nerve membrane conductance. Science 1967, 156, 976–979. [Google Scholar] [CrossRef]
- Kao, C.Y. Actions of nortetrodotoxin on frog muscle and squid axon. Toxicon 1982, 20, 1043–1050. [Google Scholar] [CrossRef]
- Takahashi, D.; Inoko, Y. Unter suchungen über das Fugugift. Cen-tralbl. Meb. Wiss. 1889, 27, 529–530. [Google Scholar]
- Takahashi, D.; Inoko, Y. Chemische unter suchungen über das Fugugift. Cen-Tralbl. Meb. Wiss. 1889, 27, 881–882. [Google Scholar]
- Tsuda, K.; Ikuma, S.; Kawamura, M.; Tachikawa, R.; Sakai, K.; Tamura, C.; Amakasu, O. Tetrodotoxin. VII. On the structures of tetrodotoxin and its derivatives. Chem. Pharm. Bull. Jpn. 1964, 12, 1357–1374. [Google Scholar] [CrossRef]
- Woodward, R.B. The structure of tetrodotoxin. Pure Appl. Chem. 1964, 9, 49–74. [Google Scholar] [CrossRef]
- Goto, T.; Kishi, Y.; Takahashi, S.; Hirata, Y. Tetrodotoxin. Tetrahedron 1965, 21, 2059–2088. [Google Scholar] [CrossRef]
- Mosher, H.S.; Fuhrman, G.J.; Fuhrman, F.A.; Fischer, H.G. Tarichatoxin-tetrodotoxin, a potent neurotoxin. Science 1964, 144, 1100–1110. [Google Scholar] [CrossRef]
- Noguchi, T.; Hashimoto, Y. Isolation of tetrodotoxin from a goby Gobius Criniger. Toxicon 1973, 11, 305–307. [Google Scholar] [CrossRef]
- Kim, Y.H.; Brown, G.B.; Mosher, H.S. Tetrodotoxin: Occurrence in atelopid frogs of Costa Rica. Science 1975, 189, 151–152. [Google Scholar] [CrossRef]
- Noguchi, T.; Arakawa, O. Tetrodotoxin—Distribution and accumulation in aquatic organisms, and cases of human intoxication. Mar. Drugs 2008, 6, 220–242. [Google Scholar] [CrossRef]
- Biessy, L.; Boundy, M.J.; Smith, K.F.; Harwood, D.T.; Hawes, I.; Wood, S.A. Tetrodotoxin in marine bivalves and edible gastropods: A mini-review. Chemosphere 2019. [Google Scholar] [CrossRef]
- Yasumoto, T.; Yasumura, D.; Yotsu, M.; Michishita, T.; Endo, A.; Kotaki, Y. Bacterial production of tetrodotoxin and anhydrotetrodotoxin. Agric. Biol. Chem. 1986, 50, 793–795. [Google Scholar]
- Noguchi, T.; Hwang, D.F.; Arakawa, O.; Sugita, H.; Deguchi, Y.; Shida, Y.; Hashimoto, K. Vibrio alginolyticus, a tetrodotoxin-producing bacterium, in the intestines of the fish Fugu vermicularis vermicularis. Mar. Biol. 1987, 94, 625–630. [Google Scholar] [CrossRef]
- Simidu, U.; Noguchi, T.; Saito, T.; Shida, Y.; Hashimoto, K. Marine bacteria which produce tetrodotoxin. Appl. Environ. Microbiol. 1987, 53, 1714–1715. [Google Scholar] [Green Version]
- Nagashima, Y.; Arakawa, O.; Sato, S. Puffer fish toxin. In Natural History of Toxic Fish and Approaching Mystery; Matsuura, K., Nagashima, Y., Eds.; Hokkaido University Press: Sapporo, Japan, 2015; pp. 33–103. (In Japanese) [Google Scholar]
- Chau, R.; Kalaitzi, J.A.; Neilan, B.A. On the origins and biosynthesis of tetrodotoxin. Aqua. Toxicol. 2011, 104, 61–72. [Google Scholar] [CrossRef]
- Bane, V.; Lehane, M.; Dikshit, M.; O’Riordan, A.; Furey, A. Tetrodotoxin: Chemistry, toxicity, source, distribution and detection. Toxins 2014, 6, 693–755. [Google Scholar] [CrossRef]
- Vlasenko, A.E.; Velansky, P.V.; Chernyshev, A.V.; Kuznetsov, V.G.; Magarlamov, T.Y. Tetrodotoxin and its analogues profile in nemertean species from the Sea of Japan. Toxicon 2018, 156, 48–51. [Google Scholar] [CrossRef]
- Yotsu-Yamashita, M.; Goto, A.; Yasumoto, T. Identification of 4-S-cysteinyltetrodotoxin from the liver of puffer fish, Fugu pardalis, and formation of thiol adducts of tetrodotoxin from 4,9-anhydrotetrodotoxin. Chem. Res. Toxicol. 2005, 18, 865–871. [Google Scholar] [CrossRef]
- Yotsu-Yamashita, M.; Abe, Y.; Kudo, Y.; Ritson-Williams, R.; Paul, V.J.; Konoki, K.; Cho, Y.; Adachi, M.; Imazu, T.; Nishikawa, T.; et al. First identification of 5,11-dideoxytetrodotoxin in marine animals, and characterization of major fragment ions of tetrodotoxin and its analogs by high resolution ESI-MS/MS. Mar. Drugs 2013, 11, 799–2813. [Google Scholar] [CrossRef]
- Jang, J.H.; Lee, J.S.; Yotsu-Yamashita, M. LC/MS analysis of tetrodotoxin and its deoxy analogues in the marine puffer fish Fugu niphobles from the southern coast of Korea, and the brackishwater puffer fishes Tetraodon nigroviridis and Tetraodon biocellatus from Southeast Asia. Mar. Drugs 2010, 8, 1049–1058. [Google Scholar] [CrossRef]
- Puilingi, C.G.; Kudo, Y.; Cho, Y.; Konoki, K.; Yotsu-Yamashita, M. Tetrodotoxin and its analogues in the pufferfish Arothron hispidus and A. nigropunctatus from the Solomon Islands: A comparison of their toxin profiles with the same species from Okinawa, Japan. Toxins 2015, 7, 3436–3454. [Google Scholar] [CrossRef]
- Turner, A.; Fenwick, D.; Powell, A.; Dhanji-Rapkova, M.; Ford, C.; Hatfield, R.; Santos, A.; Martinez-Urtaza, J.; Bean, T.; Baker-Austin, C.; et al. New invasive nemertean species (Cephalothrix Simula) in England with high levels of tetrodotoxin and a microbiome linked to toxin metabolism. Mar. Drugs 2018, 16, 452. [Google Scholar] [CrossRef]
- Watabe, S.; Sato, Y.; Nakaya, M.; Hashimoto, K.; Enomoto, A.; Kaminogawa, S.; Yamauchi, K. Monoclonal antibody raised against tetrodonic acid, a derivative of tetrodotoxin. Toxicon 1989, 27, 265–268. [Google Scholar] [CrossRef]
- Matsumura, K. A monoclonal antibody against tetrodotoxin that reacts to the active group for the toxicity. Eur. J. Pharmacol. 1995, 293, 41–45. [Google Scholar] [CrossRef]
- Matsumura, K. In vivo neutralization of tetrodotoxin by a monoclonal antibody. Toxicon 1995, 33, 1239–1241. [Google Scholar] [CrossRef]
- Kawatsu, K.; Hamano, Y.; Yoda, T.; Terano, Y.; Shibata, T. Rapid and highly sensitive enzyme immunoassay for quantitative determination of tetrodotoxin. Jpn. J. Med. Sci. Biol. 1997, 50, 133–150. [Google Scholar] [CrossRef]
- Kodama, M.; Noguchi, T.; Maruyama, J.; Ogata, T.; Hashimoto, K. Release of tetrodotoxin and paralytic shellfish poison from puffer liver by RNase. J. Biochem. 1983, 93, 243–247. [Google Scholar] [CrossRef]
- Williams, B.L.; Stark, M.R.; Caldwell, R.L. Intra-organismal distribution of tetrodotoxin in two species of blue-ringed octopuses (Hapalochlaena fasciata and H. lunulata). Toxicon 2009, 54, 345–353. [Google Scholar] [CrossRef]
- Mebs, D.; Arakawa, O.; Yotsu-Yamashita, M. Tissue distribution of tetrodotoxin in the red-spotted newt Notophthalmus viridescens. Toxicon 2010, 55, 1353–1357. [Google Scholar] [CrossRef]
- Turner, A.D.; Powell, A.; Schofield, A.; Lees, D.N.; Baker-Austin, C. Detection of the pufferfish toxin tetrodotoxin in European bivalves, England, 2013 to 2014. Eurosurveillance 2015, 20, 21009. [Google Scholar] [CrossRef]
- Vlamis, A.; Katikou, P.; Rodriguez, I.; Rey, V.; Alfonso, A.; Papazachariou, A.; Zacharaki, T.; Botana, A.M.; Botana, L.M. First detection of tetrodotoxin in Greek shellfish by UPLC-MS/MS potentially linked to the presence of the dinoflagellate Prorocentrum minimum. Toxins 2015, 7, 1779–1807. [Google Scholar]
- Mahmud, Y.; Okada, K.; Takatani, T.; Kawatsu, K.; Hamano, Y.; Arakawa, O.; Noguchi, T. Intra-tissue distribution of tetrodotoxin in two marine puffers Takifugu vermicularis and Chelonodon patoca. Toxicon 2003, 41, 13–18. [Google Scholar] [CrossRef]
- Sato, S.; Sakai, R.; Kodama, M. Identification of thioether intermediates in the reductive transformation of gonyautoxins into saxitoxins by thiols. Bioorg. Med. Chem. Lett. 2000, 10, 1787–1789. [Google Scholar] [CrossRef]
- Sato, S.; Takata, Y.; Kondo, S.; Kotoda, A.; Hongo, N.; Kodama, M. Quantitative ELISA kit for paralytic shellfish toxins coupled with sample pretreatment. J. AOAC Int. 2014, 97, 339–344. [Google Scholar] [CrossRef]
- Sato, S.; Kodama, M. Puffer fish toxin. In Shokuhin Eisei Kensa Shishin, Rikagaku-hen; Japanese Society of Food Hygiene, Ed.; Japanese Society of Food Hygiene: Tokyo, Japan, 2015; pp. 813–820. (In Japanese) [Google Scholar]
- Yotsu, M.; Endo, A.; Yasumoto, T. An improved tetrodotoxin analyzer. Agric. Biol. Chem. 1989, 53, 893–895. [Google Scholar]
- Eangoor, P.; Indapurkar, A.S.; Vakkalanka, M.; Yeh, J.S.; Knaack, J.S. Rapid and Sensitive ELISA Screening Assay for Several Paralytic Shellfish Toxins in Human Urine. J. Anal. Toxicol. 2017, 41, 755–759. [Google Scholar] [CrossRef]
- Wu, B.Q.; Yang, L.; Kao, C.Y.; Levinson, S.R.; Yotsu-Yamashita, M.; Yasumoto, T. 11-oxo-tetrodotoxin and a specifically labelled 3H-tetrodotoxin. Toxicon 1996, 34, 407–416. [Google Scholar] [CrossRef]
- Oshima, Y. Postcolumn derivatization liquid chromatographic method for paralytic shellfish toxins. J. AOAC Int. 1995, 78, 528–532. [Google Scholar]







© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sato, S.; Takaishi, S.; Yasumoto, K.; Watabe, S. Novel Polyclonal Antibody Raised against Tetrodotoxin Using Its Haptenic Antigen Prepared from 4,9-anhydrotetrodotoxin Reacted with 1,2-Ethaneditiol and Further Reacted with Keyhole Limpet Hemocyanin. Toxins 2019, 11, 551. https://doi.org/10.3390/toxins11100551
Sato S, Takaishi S, Yasumoto K, Watabe S. Novel Polyclonal Antibody Raised against Tetrodotoxin Using Its Haptenic Antigen Prepared from 4,9-anhydrotetrodotoxin Reacted with 1,2-Ethaneditiol and Further Reacted with Keyhole Limpet Hemocyanin. Toxins. 2019; 11(10):551. https://doi.org/10.3390/toxins11100551
Chicago/Turabian StyleSato, Shigeru, Suzuka Takaishi, Ko Yasumoto, and Shugo Watabe. 2019. "Novel Polyclonal Antibody Raised against Tetrodotoxin Using Its Haptenic Antigen Prepared from 4,9-anhydrotetrodotoxin Reacted with 1,2-Ethaneditiol and Further Reacted with Keyhole Limpet Hemocyanin" Toxins 11, no. 10: 551. https://doi.org/10.3390/toxins11100551