Lactobacillus bulgaricus or Lactobacillus rhamnosus Suppresses NF-κB Signaling Pathway and Protects against AFB1-Induced Hepatitis: A Novel Potential Preventive Strategy for Aflatoxicosis?

Abstract
:1. Introduction
- (1)
- Do lactic acid bacteria inhibit TNF-α, IL-1, IL-6, and IL-8 during inflammatory injury of the liver caused by AFB1?
- (2)
- Do lactic acid bacteria activate the NF-κB signaling pathway during inflammatory injury of the liver caused by AFB1?
2. Results
2.1. Limited Effects of L. bulgaricus or L. rhamnosus on Body Weights and Liver Index
2.2. Histopathological Observations of the Liver
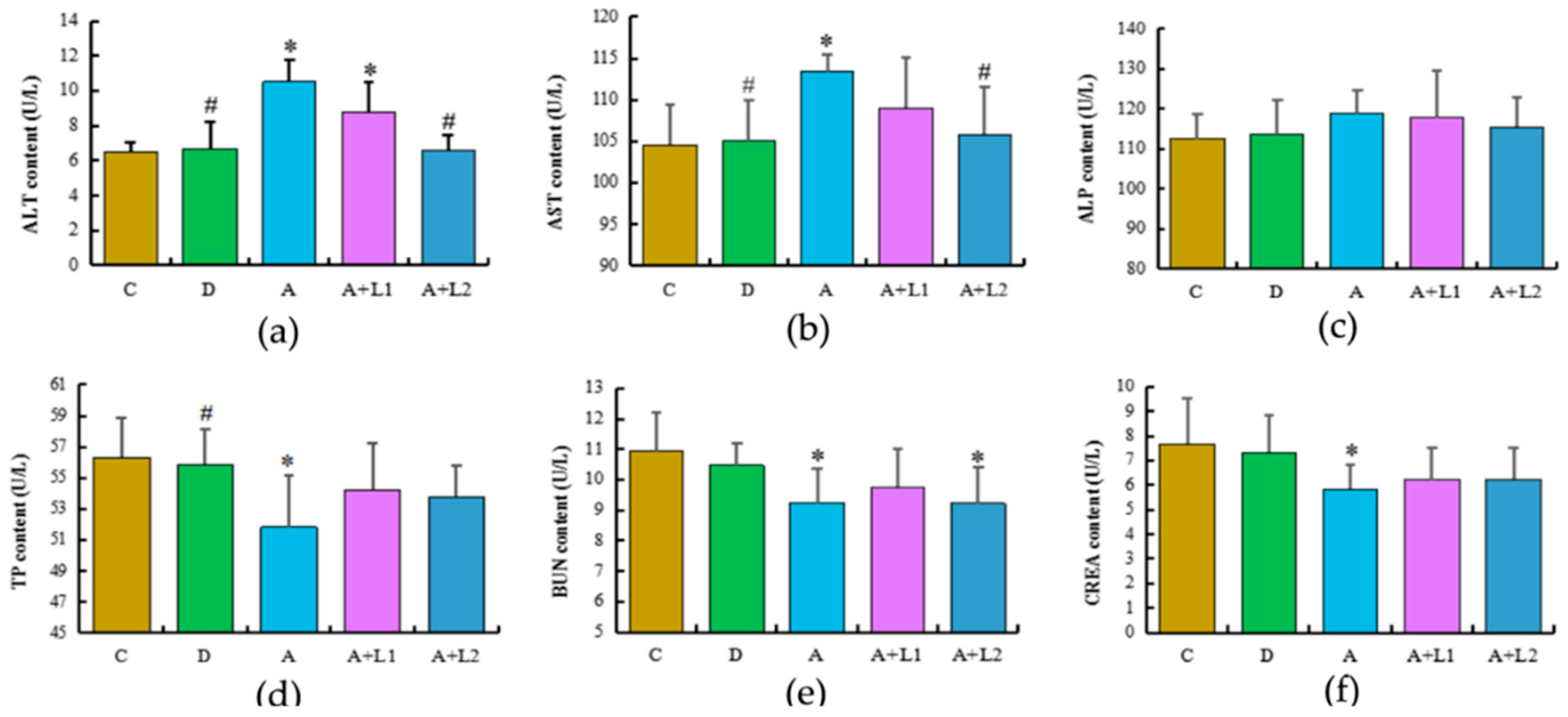
2.3. L. bulgaricus or L. rhamnosus Alleviated AFB1-Induced Hepatic and Renal Dysfunction in Mice
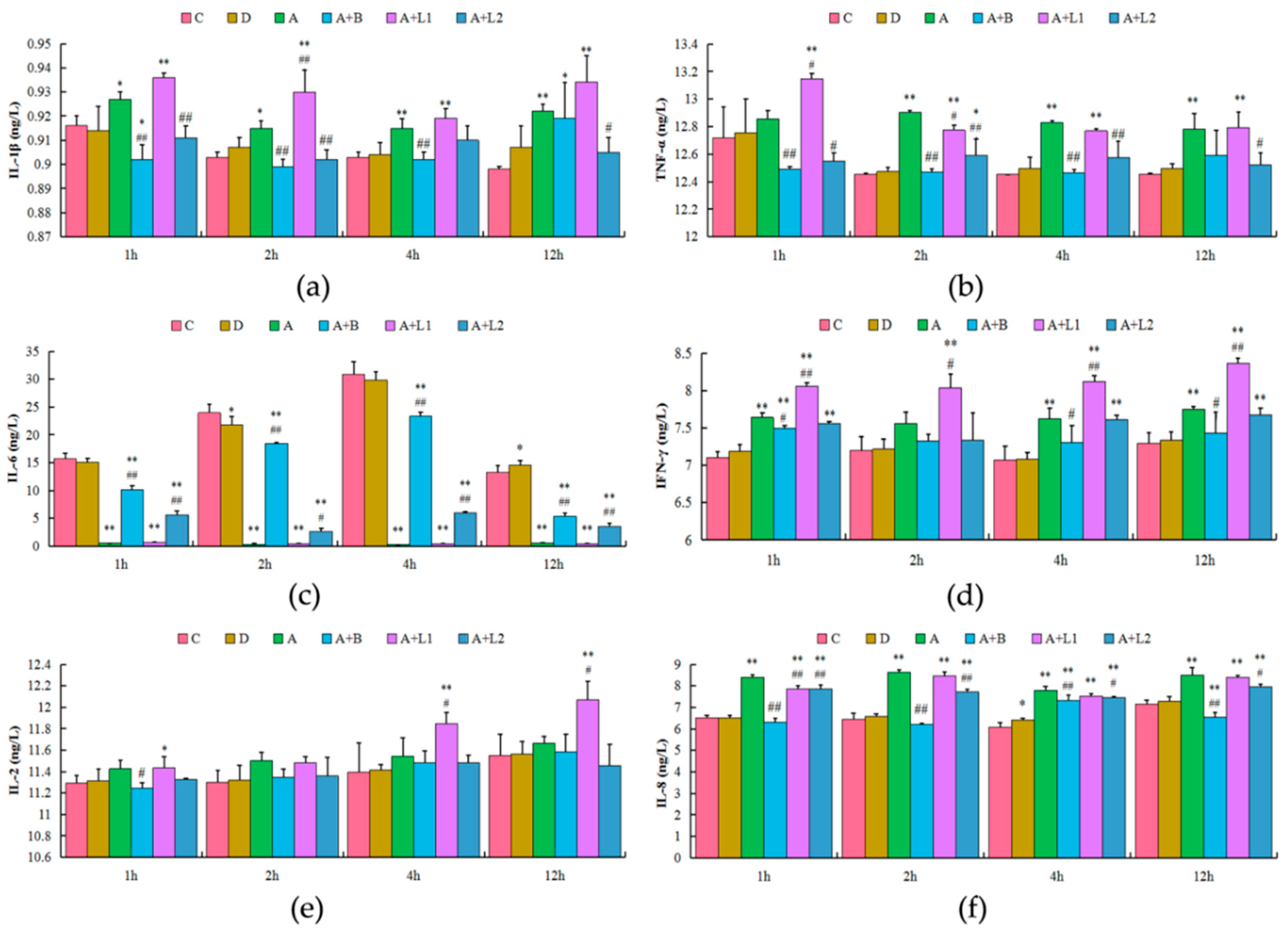
2.4. L. bulgaricus or L. rhamnosus Reduced AFB1-Induced Inflammatory Response in Mice
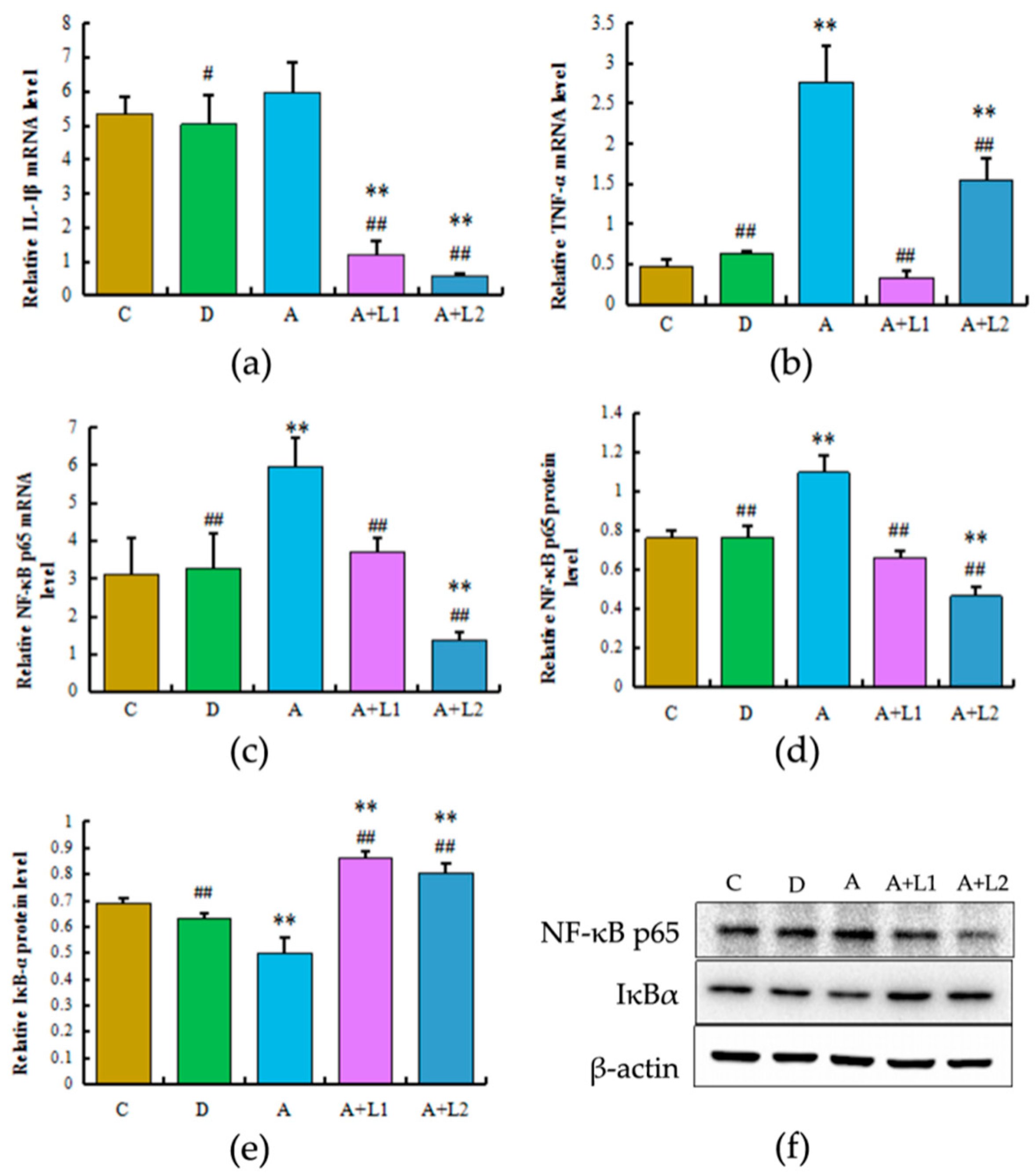
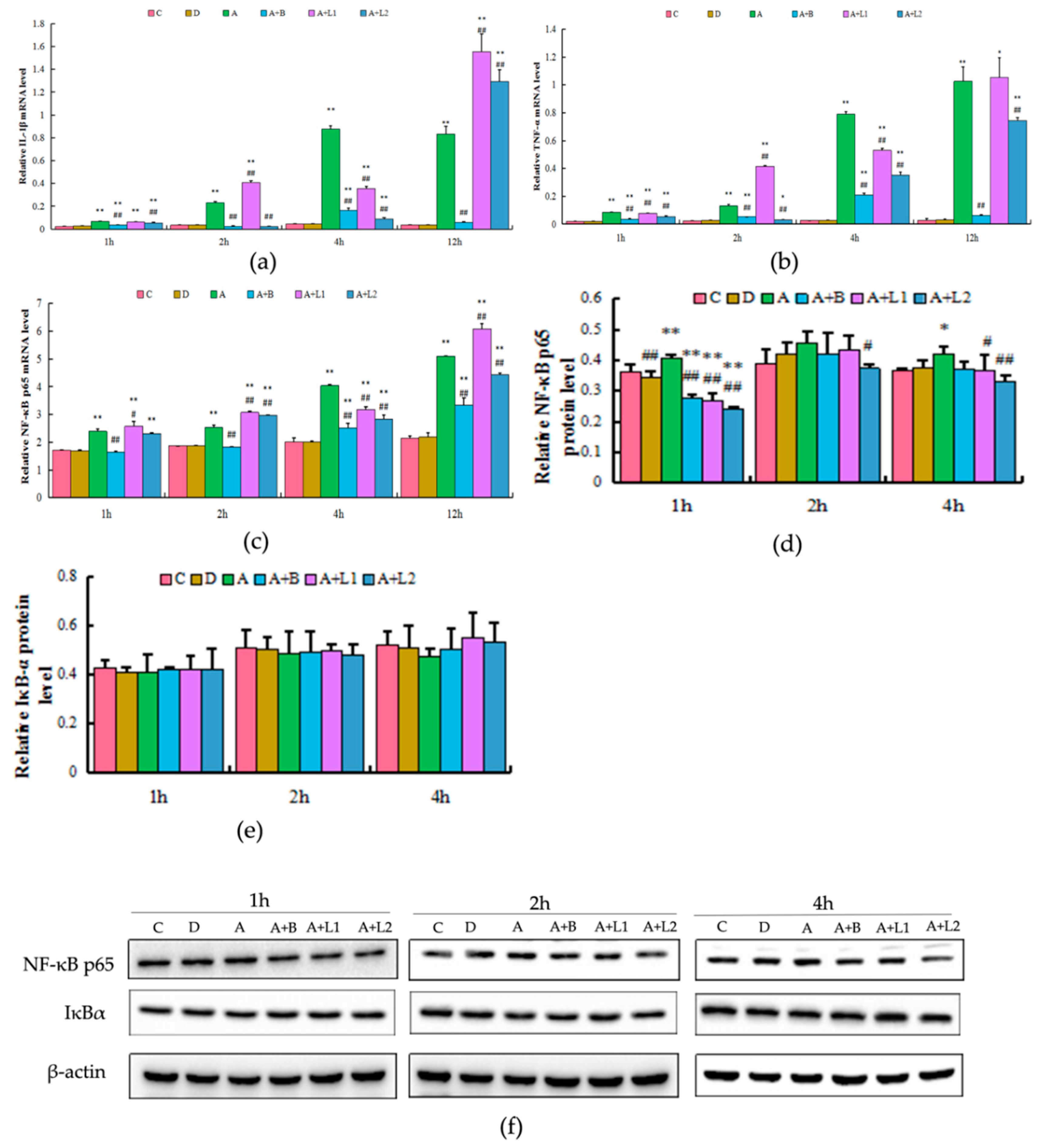
2.5. L. bulgaricus or L. rhamnosus Effectively Downregulated the Expression of Hepatic Inflammatory Cytokines and NF-κB p65 Activation in AFB1-Induced Mice
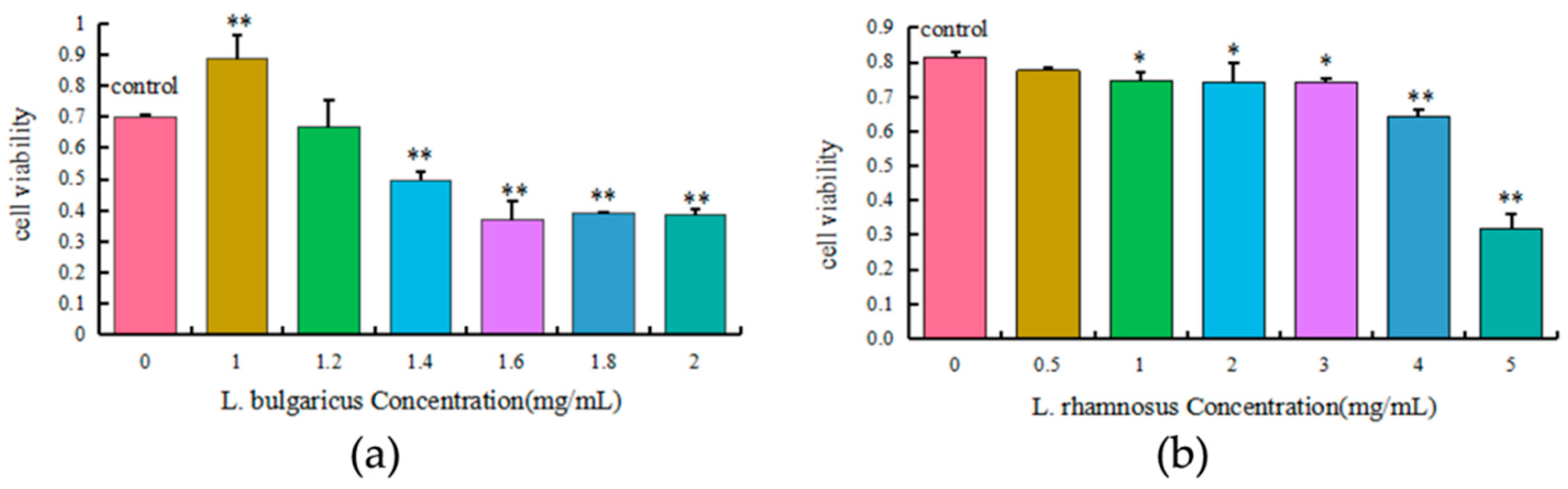
2.6. Cytotoxicity of L. bulgaricus and L. rhamnosus to AML12 Cells
2.7. L. bulgaricus and L. rhamnosus Exerted Varied Influences on the AFB1-Induced Inflammatory Response of AML12 Cells
2.8. L. bulgaricus or L. rhamnosus Attenuated Upregulation of Inflammatory Cytokines and NF-κB p65 Activation in AFB1-Induced AML12 Cells
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Ethics
5.2. Animal Experiments
5.3. Sample Collection
5.4. Histopathological Examination
5.5. Cell Culture
5.6. Biochemical Analysis
5.7. Determination of Parameters Associated with Inflammatory Cytokines in Liver Tissues
5.8. Real-Time RT-PCR Analysis
5.9. Western Blot Analysis
5.10. Statistical Analysis
Author Contributions
Funding
Conflicts of Interest
References
- Williams, J.H.; Phillips, T.D.; Jolly, P.E.; Stiles, J.K.; Jolly, C.M.; Aggarwal, D. Human aflatoxicosis in developing countries: A review of toxicology, exposure, potential health consequences and interventions. Am. J. Clin. Nutr. 2004, 80, 1106–1122. [Google Scholar] [CrossRef] [PubMed]
- Parkin, D.M.; Sitas, F.; Parker, D.M.; Ferlay, J.; Cook, A. Cancer in Africa: Epidemiology: Epidemiology and prevention. IARC Sci. Publ. 2003, 153, 1–41. [Google Scholar] [CrossRef]
- Atanda, S.A. Fungi and mycotoxins in stored foods. Afr. J. Microbiol. Res. 2011, 5, 4373–4382. [Google Scholar] [CrossRef]
- Newberne, P.M.; Butler, W.H. Acute and chronic effects of aflatoxin on the liver of domestic and laboratory animals A: A review. Cancer Res. 1969, 29, 236–250. [Google Scholar] [PubMed]
- Mohammadi, A.; Mehrzad, J.; Mahmoudi, M.; Schneider, M. Environmentally Relevant Level of Aflatoxin B1 Dysregulates Human Dendritic Cells Through Signaling on Key Toll-Like Receptors. Int. J. Toxicol. 2014, 33, 175–186. [Google Scholar] [CrossRef] [PubMed]
- Chaytor, A.C.; See, M.T.; Hansen, J.A.; de Souza, A.L.; Middleton, T.F.; Kim, S.W. Effects of chronic exposure of diets with reduced concentrations of aflatoxin and deoxynivalenol on growth and immune status of pigs. J. Anim. Sci. 2011, 89, 124–135. [Google Scholar] [CrossRef] [PubMed]
- Bishayee, A. The role of inflammation and liver cancer. Adv. Exp. Med. Biol. 2014, 81, 401. [Google Scholar] [CrossRef]
- Singh, K.B.; Maurya, B.K.; Trigun, S.K. Activation of oxidative stress and inflammatory factors could account for histopathological progression of aflatoxin-B1 induced hepatocarcinogenesis in rat. Mol. Cell Biochem. 2015, 401, 185–196. [Google Scholar] [CrossRef] [PubMed]
- Moore, R.J.; Owens, D.M.; Stamp, G.; Arnott, C.; Burke, F.; East, N.; Holdsworth, H.; Turner, L.; Rollins, B.; Pasparkis, M. Mice deficient in tumor necrosis factor-alpha are resistant to skin carcinogenesis. Nat. Med. 1999, 5, 828–831. [Google Scholar] [CrossRef] [PubMed]
- Pierron, A.; Alassane-Kpembi, I.; Oswald, I.P. Impact of mycotoxin on immune response and consequences for pig health. Anim. Nutr. 2016, 2, 63–68. [Google Scholar] [CrossRef] [PubMed]
- Xu, R.X.; Liu, R.Y.; Wu, C.M.; Zhao, Y.S.; Li, Y.; Yao, Y.Q.; Xu, Y. DNA damage-induced NF-κB activation in human glioblastoma cells promotes miR-181b expression and cell proliferation. Cell Physiol. Biochem. 2015, 35, 913. [Google Scholar] [CrossRef] [PubMed]
- Hu, Z.; Song, B.; Xu, L.; Zhong, Y.; Peng, F.; Yang, C.; Zhou, J.; Su, Y.; Chen, S.; He, Y.; et al. Aqueous synthesized quantum dots interfere with the NF-κB pathway and confer anti-tumor, anti-viral and anti-inflammatory effects. Biomaterials 2016, 108, 187–196. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Wang, J.; Wang, X.; Zhou, J.; Wang, M.; Ma, H.; Xiao, S. Increased βTrCP are associated with imiquimod-induced psoriasis-like skin inflammation in mice via NF-κB signaling pathway. Gene 2016, 592, 164–171. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Yang, N.; Wang, T.; Dai, B.; Shang, Y. Vitamin D reduces inflammatory response in asthmatic mice through HMGB1/TLR4/NF-κB signaling pathway. Mol. Med. Rep. 2018, 17, 2915–2920. [Google Scholar] [CrossRef] [PubMed]
- Mechoud, M.A.; Mateos, M.V.; Valdez, G.F.D.; Vilena, J.; Salvador, G.A.; Rodriguez, A.V. Lactobacillus reuteri, CRL1098 soluble factors modulate tumor necrosis factor alpha production in peripheral blood mononuclear cells: Involvement of lipid rafts. Int. Immunopharmacol. 2012, 14, 446–453. [Google Scholar] [CrossRef] [PubMed]
- Nobuhiro Ueno, M.D.; Mikihiro Fujiya, M.D.; Segawa, S.; Moriichi, K.; Tanabe, H.; Mizukami, Y.; Kobayashi, N.; Ito, K.; Kohgo, Y. Heat-killed body of lactobacillus brevis SBC8803 ameliorates intestinal injury in a murine model of colitis by enhancing the intestinal barrier function. Inflamm. Bowel Dis. 2011, 17, 2235–2250. [Google Scholar] [CrossRef] [PubMed]
- Colcombet, J.; Hirt, H. Arabidopsis MAPKs: A complex signalling network involved in multiple biological processes. Biochem. J. 2008, 413, 217–226. [Google Scholar] [CrossRef] [PubMed]
- Foligne, B.; Nutten, S.; Grangette, C.; Dennin, V.; Gourdercout, D.; Poiret, S.; Dewulf, J.; Brassart, D.; Mercenier, A.; Pot, B. Correlation between in vitro and in vivo immunomodulatory properties of lactic acid bacteria. World J. Gastroenterol. 2007, 13, 236–243. [Google Scholar] [CrossRef] [PubMed]
- Naqid, I.A.; Owen, J.P.; Maddison, B.C.; Gardner, D.S.; Foster, N.; Tchozervska, A.M.; la Ragione, R.; Gough, K.C. Prebiotic and probiotic agents enhance antibody-based immune responses to Salmonella, Typhimurium infection in pigs. Anim. Feed Sci. Technol. 2015, 201, 57–65. [Google Scholar] [CrossRef]
- Ewaschuk, J.; Endersby, R.; Thiel, D.; Diaz, H.; Backer, J.; Ma, M.; Curchill, T.; Madsen, K. Probiotic bacteria prevent hepatic damage and maintain colonic barrier function in a mouse model of sepsis. Hepatology 2010, 46, 841–850. [Google Scholar] [CrossRef]
- Wang, Y.; Liu, Y.; Sidhu, A.; Zhenhua, M.; McClain, C.; Feng, W. Lactobacillus rhamnosus GG culture supernatant ameliorates acute alcohol-induced intestinal permeability and liver injury. Am. J. Physiol. Gastrointest. Liver Physiol. 2012, 303, 32–41. [Google Scholar] [CrossRef] [PubMed]
- Loguercio, C.; Federico, A.; Tuccillo, C.; Terraciano, F.; D’Auria, D.V.; de Simone, C.; del Vecchio Blanco, C. Beneficial effects of a probiotic VSL#3 on parameters of liver dysfunction in chronic liver diseases. J. Clin. Gastroenterol. 2005, 39, 540–543. [Google Scholar] [CrossRef] [PubMed]
- Miettinen, M.; Lehtonen, A.; Julkunen, I.; Sampsa, M. Lactobacilli and Streptococci Activate NF-κB and STAT Signaling Pathways in Human Macrophages. J. Immunol. 2000, 164, 3733–3740. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Huang, W.; Gu, J.; Du, X.; Lei, L.; Yuan, X.; Sun, G.; Wang, Z.; Li, X.; Liu, G. SREBP-1c overactivates ROS-mediated hepatic NF-κB inflammatory pathway in dairy cows with fatty liver. Cell. Signal. 2015, 27, 2099–2109. [Google Scholar] [CrossRef] [PubMed]
- Martins, G.R.; Gelaleti, G.B.; Moschetta, M.G.; Maschio-Signorini, L.B.; Zuccari, D.A. Proinflammatory and Anti-Inflammatory Cytokines Mediated by NF-κB Factor as Prognostic Markers in Mammary Tumors. Mediat. Inflamm. 2016, 2016, 9512743. [Google Scholar] [CrossRef] [PubMed]
- Haskard, C.A.; Elnezami, H.S.; Kankaanpää, P.E.; Salminen, S.; Ahokas, J.T. Surface Binding of Aflatoxin B1 by Lactic Acid Bacteria. Appl. Environ. Microbiol. 2001, 67, 3086–3091. [Google Scholar] [CrossRef] [PubMed]
- Peltonen, K.; Elnezami, H.; Haskard, C.; Ahodas, J.T.; Salminen, S.J. Aflatoxin B1 binding by dairy strains of lactic acid bacteria and bifidobacteria. J. Dairy Sci. 2001, 84, 2152–2156. [Google Scholar] [CrossRef]
- Wang, Y.; Liu, Y.; Kirpich, I.; Ma, Z.; Wang, C.; Zhang, M.; Suttles, J.; Mcclain, C.; Feng, W. Lactobacillus rhamnosus GG reduces hepatic TNFα production and inflammation in chronic alcohol-induced liver injury. J. Nutr. Biochem. 2013, 24, 1609–1615. [Google Scholar] [CrossRef]
- Ahmad, M.; Lim, C.P.; Akowuah, G.A.; Ismai, N.N.; Hashim, A.; Hor, S.Y.; Ang, L.F.; Yam, M.F. Safety assessment of standardised methanol extract of Cinnamomum burmannii. Phytomedicine 2013, 20, 1124–1130. [Google Scholar] [CrossRef]
- Hinson, J.A.; Roberts, D.W.; James, L.P. Mechanisms of Acetaminophen-Induced Liver Necrosis. Handb. Exp. Pharmacol. 2010, 196, 369. [Google Scholar] [CrossRef]
- Lin, L.; Cui, F.; Zhang, J.; Gao, X.; Zhou, M.; Liu, M.; Yhang, C.; Jia, L. Antioxidative and renoprotective effects of residue polysaccharides from Flammulina velutipes. Carbohydr. Polym. 2016, 146, 388–395. [Google Scholar] [CrossRef] [PubMed]
- Yu, M.; Xue, J.; Li, Y.; Zhang, W.; Ma, D.; Liu, L.; Zhang, Z. Resveratrol protects against arsenic trioxide-induced nephrotoxicity by facilitating arsenic metabolism and decreasing oxidative stress. Arch. Toxicol. 2013, 87, 1025–1035. [Google Scholar] [CrossRef] [PubMed]
- Ilic, Z.; Crawford, D.; Vakharia, D.; Egner, P.A.; Sell, S. Glutathione-S-transferase A3 knockout mice are sensitive to acute cytotoxic and genotoxic effects of aflatoxin B1. Toxicol. Appl. Pharmacol. 2010, 242, 241–246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hathout, A.S.; Mohamed, S.R.; El-Nekeety, A.A.; Hasan, N.S.; Aly, S.E.; Abdel-Wahhab, M.A. Ability of Lactobacillus casei and Lactobacillus reuteri to protect against oxidative stress in rats fed aflatoxins-contaminated diet. Toxicon 2011, 58, 179–186. [Google Scholar] [CrossRef] [PubMed]
- Poór, M.; Bálint, M.; Hetényi, C.; Goder, B.; Kunsagi-Mate, S.; Koszegi, T.; Lemil, B. Investigation of Non-Covalent Interactions of Aflatoxins (B1, B2, G1, G2, and M1) with Serum Albumin. Toxins 2017, 9, 339. [Google Scholar] [CrossRef] [PubMed]
- Weninger, W.; von Andrian, U.H. Chemokine regulation of naïve T cell traffic in health and disease. Semin. Immunol. 2003, 15, 257–270. [Google Scholar] [CrossRef] [PubMed]
- Shanmugam, G.; Narasimhan, M.; Sakthivel, R.; Kumar, R.R.; Davidson, C.; Palaniapoan, S.; Claycomb, W.W.; Hoidal, J.R.; Darley-Usmar, V.M.; Rajasekaran, N.S. A biphasic effect of TNF-α in regulation of the Keap1/Nrf2 pathway in cardiomyocytes. Redox Biol. 2016, 9, 77–89. [Google Scholar] [CrossRef] [Green Version]
- Rottenberg, S.; Schmuckli-Maurer, J.; Grimm, S.; Heussler, V.T.; Dobelaere, D.A. Characterization of the bovine IkappaB kinases (IKK)alpha and IKKbeta, the regulatory subunit NEMO and their substrate IkappaBalpha. Gene 2002, 299, 293–300. [Google Scholar] [CrossRef]
- Matsumoto, S.; Hara, T.; Hori, T.; Mitsuyama, K.; Nagaoka, M.; Tomiyasu, N.; Suzuki, A.; Sata, M. Probiotic Lactobacillus-induced improvement in murine chronic inflammatory bowel disease is associated with the down-regulation of pro-inflammatory cytokines in lamina propria mononuclear cells. Clin. Exp. Immunol. 2010, 140, 417–426. [Google Scholar] [CrossRef]
- Zhang, L.; Li, N.; Caicedo, R.; Neu, J. Alive and Dead Lactobacillus rhamnosus GG Decrease Tumor Necrosis Factor-α–Induced Interleukin-8 Production in Caco-2 Cells. J. Nutr. 2005, 135, 1752–1756. [Google Scholar] [CrossRef]
- Ma, D.; Forsythe, P.; Bienenstock, J. Live Lactobacillus reuteri Is Essential for the Inhibitory Effect on Tumor Necrosis Factor Alpha-Induced Interleukin-8 Expression. Infect. Immun. 2004, 72, 5308–5314. [Google Scholar] [CrossRef] [PubMed]
- Yu, L.; Li, L.; Medeiros, L.J.; Young, K.H. NF-κB signaling pathway and its potential as a target for therapy in lymphoid neoplasms. Blood Rev. 2016, 31, 77. [Google Scholar] [CrossRef] [PubMed]
- Janus, P.; Szołtysek, K.; Zając, G.; Stokowy, T.; Walaszczyk, A.; Widlak, W.; Wojtas, B.; Gielniewski, B.; Iwanasazko, M.; Braun, R. Pro-inflammatory cytokine and high doses of ionizing radiation have similar effects on the expression of NF-kappaB-dependent genes. Cell. Signal. 2018, 46. [Google Scholar] [CrossRef] [PubMed]
- Kaulmann, A.; Legay, S.; Schneider, Y.J.; Hoffmann, L.; Bohn, T. Inflammation related responses of intestinal cells to plum and cabbage digesta with differential carotenoid and polyphenol profiles following simulated gastro-intestinal digestion. Mol. Nutr. Food Res. 2016, 60, 992–1005. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Ding, H.; Wang, X.; Liu, L.; Huang, D.; Zhang, R.; Guo, L.; Wang, Z.; Li, X.; Liu, G.; et al. High levels of acetoacetate and glucose increase expression of cytokines in bovine hepatocytes, through activation of the NF-κB signalling pathway. J. Dairy Res. 2016, 83, 51–57. [Google Scholar] [CrossRef] [PubMed]
- Chang, I.; Kim, J.D. Inhibition of Aflatoxin Production of Aspergillus flavus by Lactobacillus casei. Mycobiology 2007, 35, 76–81. [Google Scholar] [CrossRef] [PubMed]
- Gratz, S.; Täubel, M.; Juvonen, R.O.; Viluksela, L.; Turner, P.C.; Mykkanen, H.; El-Nezami, H. Lactobacillus rhamnosus Strain GG Modulates Intestinal Absorption, Fecal Excretion, and Toxicity of Aflatoxin B1 in Rats. Appl. Environ. Microbiol. 2006, 72, 7398. [Google Scholar] [CrossRef] [PubMed]
- Kuisma, J.; Mentula, S.; Jarvinen, H.; Kahri, A.; Saxelin, M.; Farkila, M. Effect of Lactobacillus rhamnosus, GG on ileal pouch inflammation and microbial flora. Aliment. Pharmacol. Ther. 2003, 17, 509–515. [Google Scholar] [CrossRef] [PubMed]
- Gerbaldo, G.A.; Barberis, C.; Pascual, L.; Dalcero, A.M.; Barberis, L. Antifungal activity of two Lactobacillus, strains with potential probiotic properties. FEMS Microbiol. Lett. 2012, 332, 27–33. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Groups | Body Weights (g) | Liver Index (%) | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Day 7 | Day 14 | Day 21 | Day 28 | Day 35 | Day 42 | Day 49 | Day 56 | ||
| C | 36.04 ± 0.54 | 37.07 ± 3.17 | 37.69 ± 3.72 | 41.53 ± 2.10 | 42.83 ± 1.88 | 43.74 ± 2.82 | 45.93 ± 2.32 | 46.19 ± 3.31 | 4.02 ± 0.243 |
| D | 36.57 ± 1.03 | 38.53 ± 1.99 | 39.73 ± 1.17 | 42.20 ± 1.61 | 43.63 ± 2.84 | 43.83 ± 2.34 | 46.27 ± 2.48 | 45.47 ± 1.50 | 3.96 ± 0.189 # |
| A | 35.47 ± 1.14 | 41.02 ± 1.62 * | 39.82 ± 1.48 | 39.63 ± 2.79 | 41.05 ± 4.28 | 41.20 ± 4.84 | 44.17 ± 1.96 | 45.47 ± 1.86 | 4.27 ± 0.171 * |
| A + L1 | 36.40 ± 2.25 | 36.16 ± 2.97 # | 36.58 ± 3.21 # | 40.33 ± 4.96 | 42.79 ± 2.70 | 43.66 ± 3.91 | 44.53 ± 3.12 | 46.47 ± 2.29 | 4.12 ± 0.191 |
| A + L2 | 36.70 ± 2.80 | 35.05 ± 2.02 # | 39.35 ± 1.82 | 39.30 ± 2.68 | 38.38 ± 3.52 * | 42.00 ± 1.90 | 43.43 ± 2.80 | 45.35 ± 2.80 | 4.09 ± 0.168 |
| Gene | Primer Sequence (5′→3′) | Product Size (bp) |
|---|---|---|
| β-actin | F: 5′-GAGACCTTCAACACCCCAGC-3′ | 263 |
| R: 5′-ATGTCACGCACGATTTCCC-3′ | ||
| p65 | F: 5′-GCTCCTGTTCGAGTCTCCATG-3′ | 91 |
| R: 5′-CATCTGTGTCTGGCAAGTACTGG-3′ | ||
| IL-1β | F: 5′-AGCTTCAAATCTCGCAGCAG-3′ | 72 |
| R: 5′-TCTCCACAGCCACAATGAGT-3′ | ||
| TNF-α | F: 5′-CTCATGCACCACCATCAAGG-3′ | 96 |
| R: 5′-ACCTGACCACTCTCCCTTTG-3′ |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, Y.; Li, R.; Chang, Q.; Dong, Z.; Yang, H.; Xu, C. Lactobacillus bulgaricus or Lactobacillus rhamnosus Suppresses NF-κB Signaling Pathway and Protects against AFB1-Induced Hepatitis: A Novel Potential Preventive Strategy for Aflatoxicosis? Toxins 2019, 11, 17. https://doi.org/10.3390/toxins11010017
Chen Y, Li R, Chang Q, Dong Z, Yang H, Xu C. Lactobacillus bulgaricus or Lactobacillus rhamnosus Suppresses NF-κB Signaling Pathway and Protects against AFB1-Induced Hepatitis: A Novel Potential Preventive Strategy for Aflatoxicosis? Toxins. 2019; 11(1):17. https://doi.org/10.3390/toxins11010017
Chicago/Turabian StyleChen, Yuanyuan, Ruirui Li, Qiaocheng Chang, Zhihao Dong, Huanmin Yang, and Chuang Xu. 2019. "Lactobacillus bulgaricus or Lactobacillus rhamnosus Suppresses NF-κB Signaling Pathway and Protects against AFB1-Induced Hepatitis: A Novel Potential Preventive Strategy for Aflatoxicosis?" Toxins 11, no. 1: 17. https://doi.org/10.3390/toxins11010017