Development and Application of a Quantitative PCR Assay to Assess Genotype Dynamics and Anatoxin Content in Microcoleus autumnalis-Dominated Mats


Abstract
:1. Introduction
2. Results
2.1. Quantitative PCR
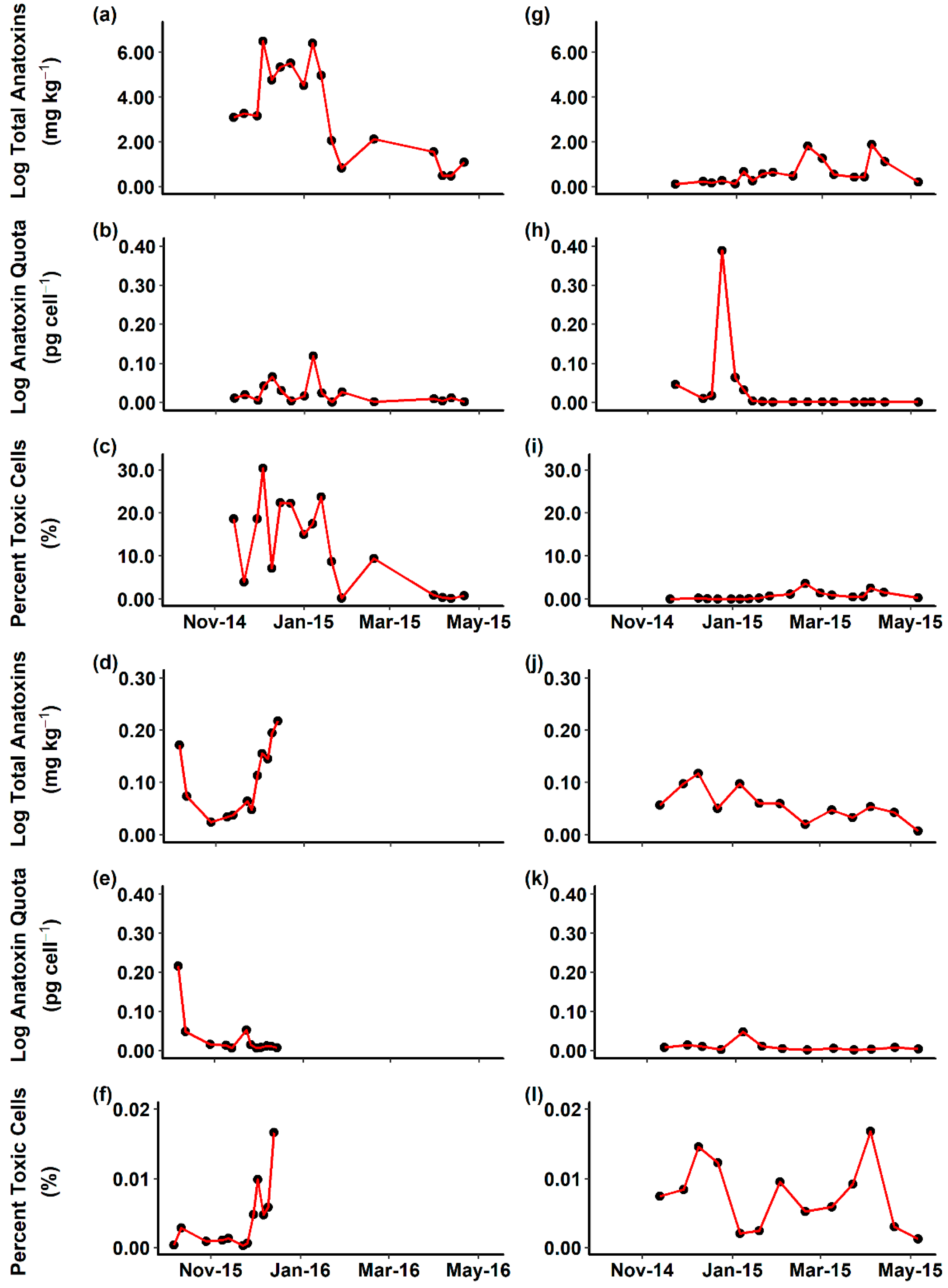
2.2. Environmental Samples
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Samples for Quantitative PCR Assay Development and Validation
5.2. Primer Design and Optimisation for Quantitative PCR Analyses
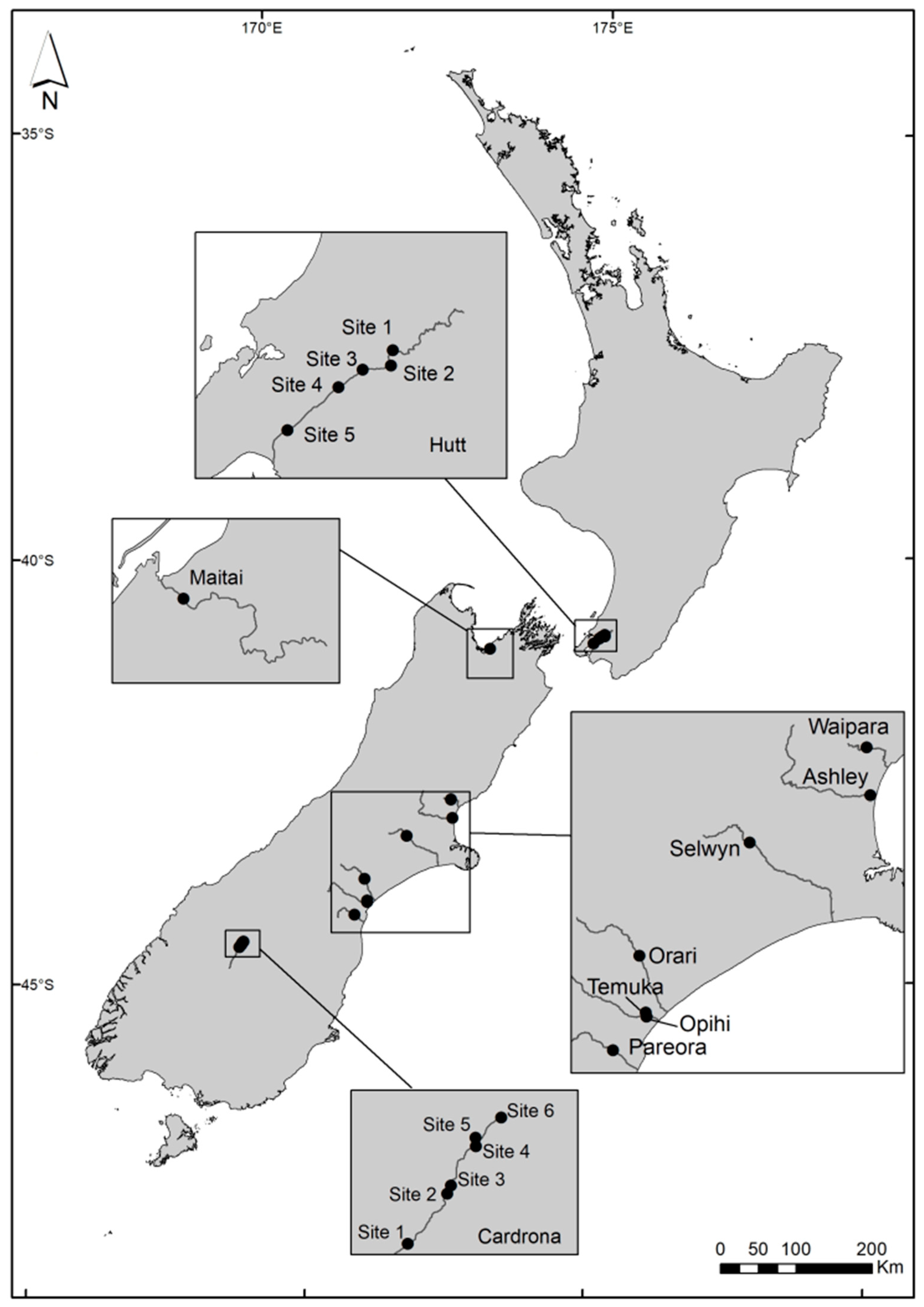
5.3. Environmental Sample Collection
5.4. DNA Extraction and Test for Inhibition
5.5. Quantitative PCR of 16S rRNA Reference Gene
5.6. Anatoxin Extraction and Analysis
5.7. Data and Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Culture Identification | Anatoxin Producer | Reference |
|---|---|---|---|
| Aphanocapsa sp. | CAWBG585 | − | |
| Aphanocapsa sp. | CAWBG615 | − | |
| Aphanizomenon sp. | CAWBG588 | − | |
| Aphanizomenon sp. | CAWBG594 | − | |
| Aphanizomenon sp. | CAWBG01 | ||
| Cuspidothrix issatchenkoi | CAWBG02 | + | [34] |
| Dolichospermum circinale | CAWBG571 | − | |
| Dolichospermum circinale | CAWBG601 | − | |
| Dolichospermum lemmermanii | CAWBG572 | − | |
| Dolichospermum lemmermanii | CAWBG593 | − | |
| Dolichospermum planktonicum | CAWBG589 | − | |
| Dolichospermum spiroides | CAWBG558 | − | |
| Dolichospermum spiroides | CAWBG583 | − | |
| Dolichospermum sp. | CAWBG608 | − | |
| Leptolyngbya sp. | CAWBG586 | − | |
| Leptolyngbya sp. | CAWBG700 | − | |
| Microcystis sp. | CAWBG580 | − | |
| Microcystis sp. | CAWBG561 | − | |
| Nostoc sp. | S. Wood submitted | − | |
| Oscillatoria sp. | PCC 6506 | + | [35] |
| Oscillatoria sp. | PCC 9029 | + | [36] |
| Planktothrix sp. | CAWBG616 | − | |
| Raphidiopsis raciborskii | CAWBG598 | − | |
| Raphidiopsis raciborskii | CAWBG614 | − | |
| Scytonema sp. | CAWBG699 | − | |
| Scytonema cf. crispum | CAWBG72 | − |
| Species | Culture Identification | Genbank Accession Number (16S rRNA Gene) | Reference |
|---|---|---|---|
| Microcoleus autumnalis | CAWBG 618 | KX016036 | [28] |
| M. autumnalis | CAWBG 619 | KX016037 | [28] |
| M. autumnalis | CAWBG 620 | KX016038 | [28] |
| M. autumnalis | CAWBG 621 | KX016039 | [28] |
| M. autumnalis | CAWBG 622 | KX016040 | [28] |
| M. autumnalis | CAWBG 623 | KX016041 | [28] |
| Oscillatoria sp. | PCC 10111 | JF803654.1 | [10] |
| Oscillatoria sp. | PCC 10601 | JF803652.1 | [10] |
| Oscillatoria sp. | PCC 6407 | JF803648.1 | [10] |
| Oscillatoria sp. | PCC 9240 | JF803651.1 | [10] |
| Oscillatoria sp. | PCC 9240-strain 193 | JF803653.1 | [10] |
References
- Heath, M.W.; Wood, S.A.; Ryan, K.G. Polyphasic assessment of fresh-water benthic mat-forming cyanobacteria isolated from New Zealand. FEMS Microbiol. Ecol. 2010, 73, 95–109. [Google Scholar] [CrossRef] [PubMed]
- Wood, S.A.; Selwood, A.I.; Rueckert, A.; Holland, P.T.; Milne, J.R.; Smith, K.F.; Smits, B.; Watts, L.F.; Cary, C.S. First report of homoanatoxin-a and associated dog neurotoxicosis in New Zealand. Toxicon 2007, 50, 292–301. [Google Scholar] [CrossRef] [PubMed]
- Teneva, I.; Dzhambazov, B.; Koleva, L.; Mladenov, R.; Schirmer, K. Toxic potential of five freshwater Phormidium species (Cyanoprokaryota). Toxicon 2005, 45, 711–725. [Google Scholar] [CrossRef] [PubMed]
- Wood, S.A.; Heath, M.W.; Kuhajek, J.; Ryan, K.G. Fine-scale spatial variability in anatoxin-a and homoanatoxin-a concentrations in benthic cyanobacterial mats: Implication for monitoring and management. J. Appl. Microbiol. 2010, 109, 2011–2018. [Google Scholar] [CrossRef] [PubMed]
- Gugger, M.; Lenoir, S.; Berger, C.; Ledreux, A.; Druart, J.-C.; Humbert, J.-F.; Guette, C.; Bernard, C. First report in a river in France of the benthic cyanobacterium Phormidium favosum producing anatoxin-a associated with dog neurotoxicosis. Toxicon 2005, 45, 919–928. [Google Scholar] [CrossRef] [PubMed]
- Quiblier, C.; Wood, S.; Echenique-Subiabre, I.; Heath, M.; Villeneuve, A.; Humbert, J.-F. A review of current knowledge on toxic benthic freshwater cyanobacteria–ecology, toxin production and risk management. Water Res. 2013, 47, 5464–5479. [Google Scholar]
- Wood, S.A.; Smith, F.M.; Heath, M.W.; Palfroy, T.; Gaw, S.; Young, R.G.; Ryan, K.G. Within-mat variability in anatoxin-a and homoanatoxin-a production among benthic Phormidium (cyanobacteria) strains. Toxins 2012, 4, 900–912. [Google Scholar] [CrossRef] [PubMed]
- Méjean, A.; Dalle, K.; Paci, G.; Bouchonnet, S.; Mann, S.; Pichon, V.; Ploux, O. Dihydroanatoxin-a Is Biosynthesized from Proline in Cylindrospermum stagnale PCC 7417: Isotopic Incorporation Experiments and Mass Spectrometry Analysis. J. Nat. Prod. 2016, 79, 1775–1782. [Google Scholar] [CrossRef] [PubMed]
- Méjean, A.; Paci, G.; Gautier, V.; Ploux, O. Biosynthesis of anatoxin-a and analogues (anatoxins) in cyanobacteria. Toxicon 2014, 91, 15–22. [Google Scholar] [CrossRef] [PubMed]
- Rantala-Ylinen, A.; Känä, S.; Wang, H.; Rouhiainen, L.; Wahlsten, M.; Rizzi, E.; Berg, K.; Gugger, M.; Sivonen, K. Anatoxin-a synthetase gene cluster of the cyanobacterium Anabaena sp. strain 37 and molecular methods to detect potential producers. Appl. Environ. Microbiol. 2011, 77, 7271–7278. [Google Scholar] [CrossRef] [PubMed]
- McAllister, T.G.; Wood, S.A.; Hawes, I. The rise of toxic benthic Phormidium proliferations: A review of their taxonomy, distribution, toxin content and factors regulating prevalence and increased severity. Harmful Algae 2016, 55, 282–294. [Google Scholar] [CrossRef] [PubMed]
- Heath, M.W.; Wood, S.A.; Ryan, K.G. Spatial and temporal variability in Phormidium mats and associated anatoxin-a and homoanatoxin-a in two New Zealand rivers. Aquat. Microb. Ecol. 2011, 64, 69. [Google Scholar] [CrossRef]
- Wood, S.; Puddick, J. The abundance of toxic genotypes is a key contributor to anatoxin variability in Phormidium-dominated benthic mats. Mar. Drugs 2017, 15, 307. [Google Scholar] [CrossRef] [PubMed]
- Wood, S.A.; Atalah, J.; Wagenhoff, A.; Brown, L.; Doehring, K.; Young, R.G.; Hawes, I. Effect of river flow, temperature, and water chemistry on proliferations of the benthic anatoxin-producing cyanobacterium Phormidium. Freshw. Sci. 2017, 36, 63–76. [Google Scholar] [CrossRef]
- Wood, S.A.; Depree, C.; Brown, L.; McAllister, T.; Hawes, I. Entrapped Sediments as a Source of Phosphorus in Epilithic Cyanobacterial Proliferations in Low Nutrient Rivers. PLoS ONE 2015, 10, e0141063. [Google Scholar] [CrossRef] [PubMed]
- Brasell, K.A.; Heath, M.W.; Ryan, K.G.; Wood, S.A. Successional Change in Microbial Communities of Benthic Phormidium-Dominated Biofilms. Microb. Ecol. 2015, 69, 254–266. [Google Scholar] [CrossRef] [PubMed]
- Heath, M.W.; Wood, S.A.; Barbieri, R.F.; Young, R.G.; Ryan, K.G. Effects of nitrogen and phosphorus on anatoxin-a, homoanatoxin-a, dihydroanatoxin-a and dihydrohomoanatoxin-a production by Phormidium autumnale. Toxicon 2014, 92, 179–185. [Google Scholar] [CrossRef] [PubMed]
- McAllister, T.G.; Wood, S.A.; Atalah, J.; Hawes, I. Spatiotemporal dynamics of Phormidium cover and anatoxin concentrations in eight New Zealand rivers with contrasting nutrient and flow regimes. Sci. Total Environ. 2018, 612, 71–80. [Google Scholar] [CrossRef] [PubMed]
- Al-Tebrineh, J.; Pearson, L.A.; Yasar, S.A.; Neilan, B.A. A multiplex qPCR targeting hepato-and neurotoxigenic cyanobacteria of global significance. Harmful Algae 2012, 15, 19–25. [Google Scholar] [CrossRef]
- Bouma-Gregson, K.; Kudela, R.M.; Power, M.E. Widespread anatoxin-a detection in benthic cyanobacterial mats throughout a river network. PLoS ONE 2018, 13, e0197669. [Google Scholar] [CrossRef] [PubMed]
- McAllister, T.G.; Wood, S.A.; Greenwood, M.J.; Broghammer, F.; Hawes, I. The effects of velocity and nitrate on Phormidium accrual cycles: A stream mesocosm experiment. Freshw. Sci. 2018, 37, 496–509. [Google Scholar] [CrossRef]
- Burford, M.A.; Davis, T.W.; Orr, P.T.; Sinha, R.; Willis, A.; Neilan, B.A. Nutrient-related changes in the toxicity of field blooms of the cyanobacterium, Cylindrospermopsis raciborskii. FEMS Microbiol. Ecol. 2014, 89, 135–148. [Google Scholar] [CrossRef] [PubMed]
- Davis, T.W.; Berry, D.L.; Boyer, G.L.; Gobler, C.J. The effects of temperature and nutrients on the growth and dynamics of toxic and non-toxic strains of Microcystis during cyanobacteria blooms. Harmful Algae 2009, 8, 715–725. [Google Scholar] [CrossRef]
- Baxa, D.V.; Kurobe, T.; Ger, K.A.; Lehman, P.W.; Teh, S.J. Estimating the abundance of toxic Microcystis in the San Francisco Estuary using quantitative real-time PCR. Harmful Algae 2010, 9, 342–349. [Google Scholar] [CrossRef]
- Bukowska, A.; Kaliński, T.; Koper, M.; Kostrzewska-Szlakowska, I.; Kwiatowski, J.; Mazur-Marzec, H.; Jasser, I. Predicting blooms of toxic cyanobacteria in eutrophic lakes with diverse cyanobacterial communities. Sci. Rep. 2017, 7, 8342. [Google Scholar] [CrossRef] [PubMed]
- Davis, T.W.; Harke, M.J.; Marcoval, M.A.; Goleski, J.; Orano-Dawson, C.; Berry, D.L.; Gobler, C.J. Effects of nitrogenous compounds and phosphorus on the growth of toxic and non-toxic strains of Microcystis during cyanobacterial blooms. Aquat. Microb. Ecol. 2010, 61, 149–162. [Google Scholar] [CrossRef]
- Heath, M.; Wood, S.A.; Young, R.G.; Ryan, K.G. The role of nitrogen and phosphorus in regulating Phormidium sp.(cyanobacteria) growth and anatoxin production. FEMS Microbiol. Ecol. 2016, 92, fiw021. [Google Scholar] [CrossRef] [PubMed]
- Wood, S.A.; Puddick, J.; Fleming, R.; Heussner, A.H. Detection of anatoxin-producing Phormidium in a New Zealand farm pond and an associated dog death. N. Z. J. Bot. 2017, 55, 36–46. [Google Scholar] [CrossRef]
- Thompson, J.D.; Higgins, D.G.; Gibson, T.J. CLUSTAL W: Improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res. 1994, 22, 4673–4680. [Google Scholar] [CrossRef] [PubMed]
- Haugland, R.A.; Siefring, S.C.; Wymer, L.J.; Brenner, K.P.; Dufour, A.P. Comparison of Enterococcus measurements in freshwater at two recreational beaches by quantitative polymerase chain reaction and membrane filter culture analysis. Water Res. 2005, 39, 559–568. [Google Scholar] [CrossRef] [PubMed]
- Al-Tebrineh, J.; Mihali, T.K.; Pomati, F.; Neilan, B.A. Detection of saxitoxin-producing cyanobacteria and Anabaena circinalis in environmental water blooms by quantitative PCR. Appl. Environ. Microbiol. 2010, 76, 7836–7842. [Google Scholar] [CrossRef] [PubMed]
- Jungblut, A.D.; Hawes, I.; Mountfort, D.; Hitzfeld, B.; Dietrich, D.R.; Burns, B.P.; Neilan, B.A. Diversity within cyanobacterial mat communities in variable salinity meltwater ponds of McMurdo Ice Shelf, Antarctica. Environ. Microbiol. 2005, 7, 519–529. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Větrovský, T.; Baldrian, P. The Variability of the 16S rRNA Gene in Bacterial Genomes and Its Consequences for Bacterial Community Analyses. PLoS ONE 2013, 8, e57923. [Google Scholar] [CrossRef] [PubMed]
- Wood, S.A.; Rasmussen, J.P.; Holland, P.T.; Campbell, R.; Crowe, A.L. First report of the cyanotoxin anatoxin-a from Aphanizomenon issatschenkoi (cyanobacteria). J. Phycol. 2007, 43, 356–365. [Google Scholar] [CrossRef]
- Méjean, A.; Mazmouz, R.; Mann, S.; Calteau, A.; Médigue, C.; Ploux, O. The genome sequence of the cyanobacterium Oscillatoria sp. PCC 6506 reveals several gene clusters responsible for the biosynthesis of toxins and secondary metabolites. J. Bacteriol. 2010, 192, 5264–5265. [Google Scholar] [CrossRef] [PubMed]
- Aráoz, R.; Nghiêm, H.-O.; Rippka, R.; Palibroda, N.; de Marsac, N.T.; Herdman, M. Neurotoxins in axenic oscillatorian cyanobacteria: Coexistence of anatoxin-a and homoanatoxin-a determined by ligand-binding assay and GC/MS. Microbiology 2005, 151, 1263–1273. [Google Scholar] [CrossRef] [PubMed]





| Strain | AnaC | AnaF * | Anatoxins ** (LC-MS/MS) |
|---|---|---|---|
| CAWBG24 | − | nt | − |
| CAWBG26 | − | nt | − |
| CAWBG32 | + | nt | nt |
| CAWBG36 | − | nt | nt |
| CAWBG37 | − | nt | nt |
| CAWBG38 | − | nt | − |
| CAWBG46 | − | nt | − |
| CAWBG48 | − | nt | − |
| CAWBG50 | − | nt | − |
| CAWBG51 | − | nt | − |
| CAWBG52 | − | nt | − |
| CAWBG53 | − | nt | − |
| CAWBG54 | − | nt | − |
| CAWBG55 | − | nt | − |
| CAWBG56 | − | nt | nt |
| CAWBG57 | − | nt | nt |
| CAWBG58 | + | nt | nt |
| CAWBG71 | − | nt | nt |
| CAWBG503 | + | + | + |
| CAWBG507 | − | − | − |
| CAWBG511 | − | − | − |
| CAWBG512 | − | − | − |
| CAWBG520 | + | + | + |
| CAWBG521 | + | + | + |
| CAWBG556 | + | + | + |
| CAWBG557 | + | + | + |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kelly, L.T.; Wood, S.A.; McAllister, T.G.; Ryan, K.G. Development and Application of a Quantitative PCR Assay to Assess Genotype Dynamics and Anatoxin Content in Microcoleus autumnalis-Dominated Mats. Toxins 2018, 10, 431. https://doi.org/10.3390/toxins10110431
Kelly LT, Wood SA, McAllister TG, Ryan KG. Development and Application of a Quantitative PCR Assay to Assess Genotype Dynamics and Anatoxin Content in Microcoleus autumnalis-Dominated Mats. Toxins. 2018; 10(11):431. https://doi.org/10.3390/toxins10110431
Chicago/Turabian StyleKelly, Laura T., Susanna A. Wood, Tara G. McAllister, and Ken G. Ryan. 2018. "Development and Application of a Quantitative PCR Assay to Assess Genotype Dynamics and Anatoxin Content in Microcoleus autumnalis-Dominated Mats" Toxins 10, no. 11: 431. https://doi.org/10.3390/toxins10110431