C-Type Lectin-20 Interacts with ALP1 Receptor to Reduce Cry Toxicity in Aedes aegypti

 and
and
Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
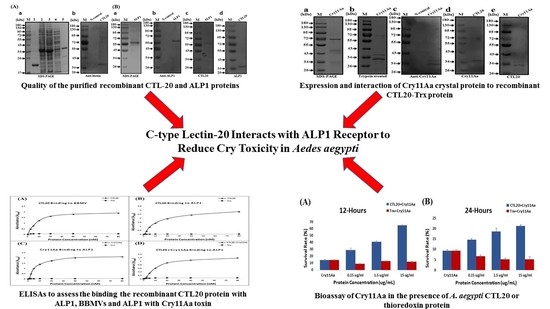
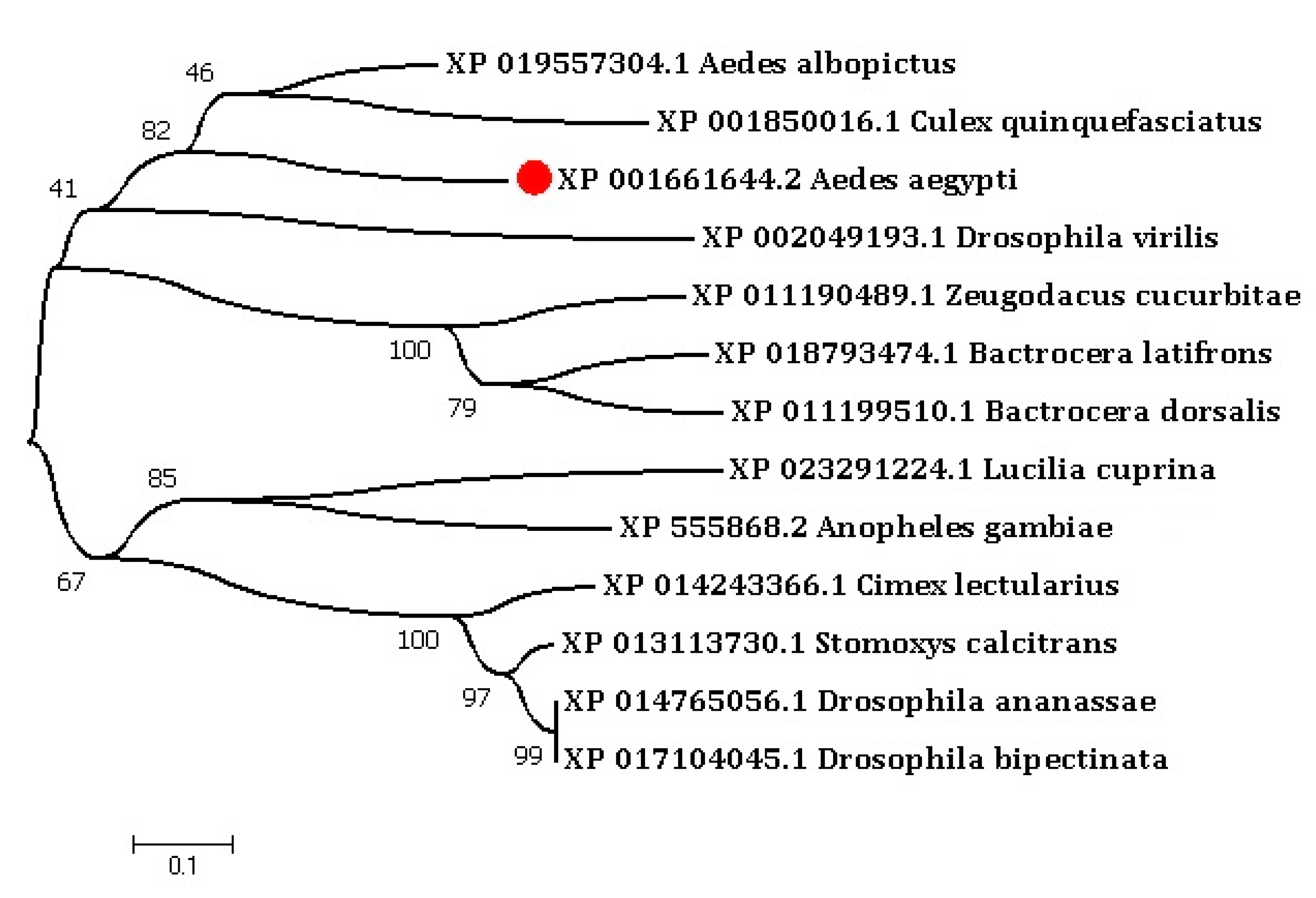
2.1. Characterization of A. aegypti CTL-20
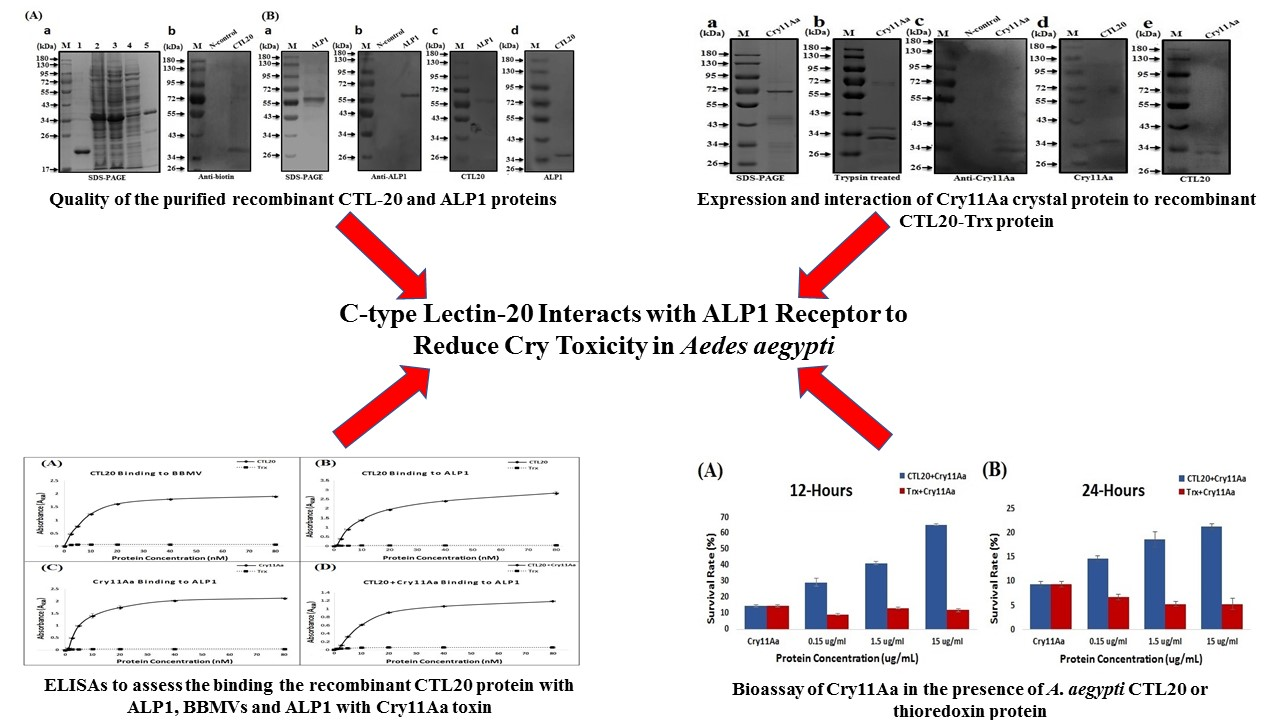
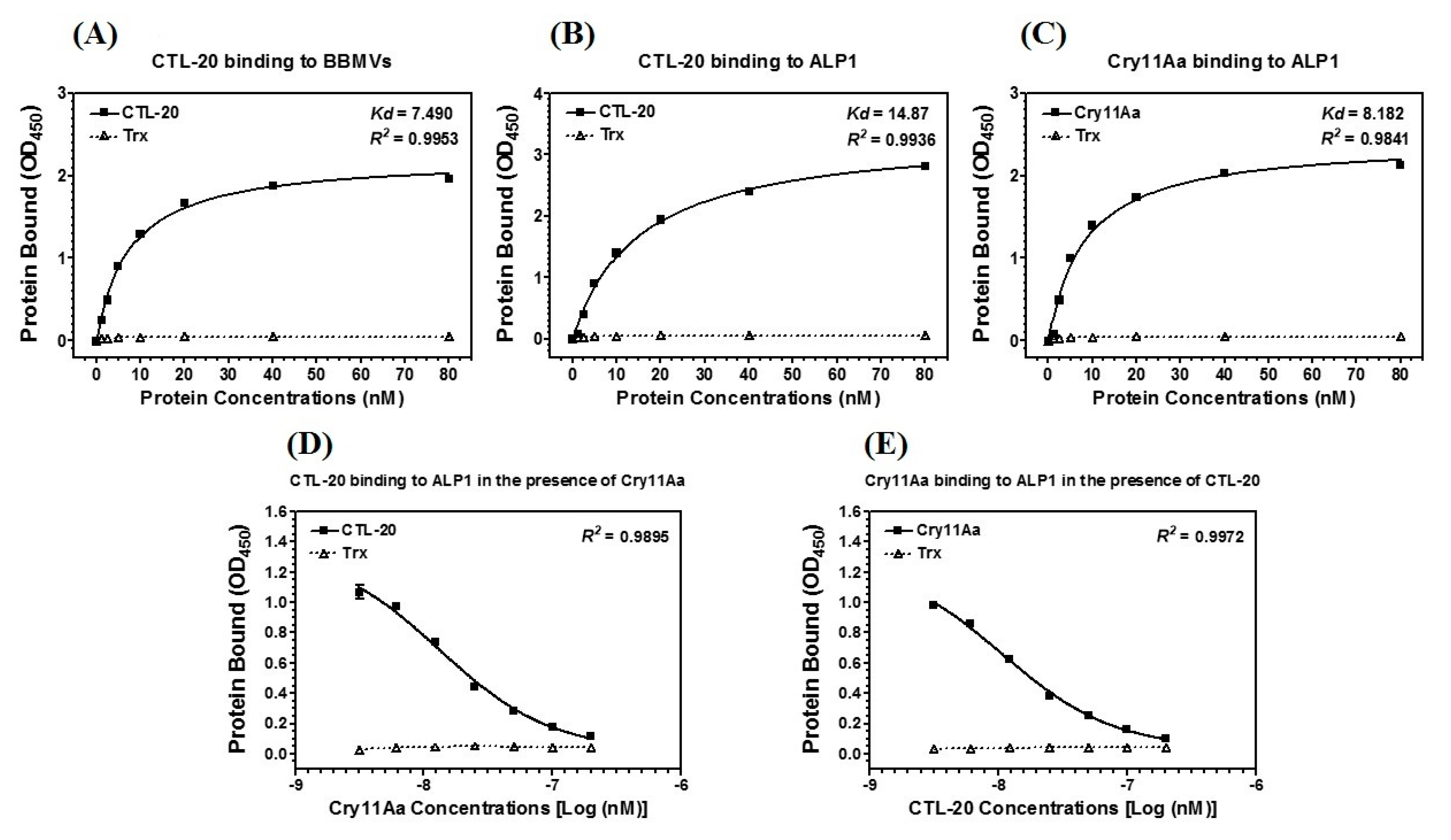
2.2. CTL-20 Binds to ALP 1 and Cry11Aa
2.3. CTL-20 Binds to BBMVs and ALP1
2.4. CTL-20 Inhibits Larvicidal Activity of Cry Toxins
3. Discussion
4. Materials and Methods
4.1. Mosquitoes and Sample Collection
4.2. Cry11Aa Toxin Purification and Activation
4.3. CTL-20 Full-Length cDNA Cloning
4.4. Bioinformatics Analysis
4.5. Prokaryotic Expression and Purification of Recombinant CTL-20
4.6. Insect Feeding Assay
4.7. Preparation of A. aegypti BBMVs
4.8. Western Blot and Far-Western Blot Analyses
4.9. Plate ELISA Assays
Author Contributions
Funding
Conflicts of Interest
References
- Christophers, S. Aedes aegypti (L.) the Yellow Fever Mosquito: Its Life History, Bionomics and Structure; Cambridge University Press: New York, NY, USA, 1960. [Google Scholar]
- Gubler, D.J. Epidemic dengue/dengue hemorrhagic fever as a public health, social and economic problem in the 21st century. Trends Microbiol. 2002, 10, 100–103. [Google Scholar] [CrossRef]
- Ligon, B.L. In Dengue fever and dengue hemorrhagic fever: A review of the history, transmission, treatment, and prevention. Semin. Pediatr. Infect. Dis. 2005, 16, 60–65. [Google Scholar] [CrossRef] [PubMed]
- Ligon, B.L. In Reemergence of an unusual disease: The Chikungunya epidemic. Semin. Pediatr. Infect. Dis. 2006, 17, 99–104. [Google Scholar] [CrossRef] [PubMed]
- Tomori, O. Yellow fever: The recurring plague. Crit. Rev. Clin. Lab. Sci. 2004, 41, 391–427. [Google Scholar] [CrossRef]
- Rajah, M.M.; Pardy, R.D.; Condotta, S.A.; Richer, M.J.; Sagan, S.M. Zika virus: Emergence, phylogenetics, challenges, and opportunities. ACS Infect. Dis. 2016, 2, 763–772. [Google Scholar] [CrossRef] [PubMed]
- TuiskunenBäck, A.; Lundkvist, Å. Dengue viruses—An overview. Infect. Ecol. Epidemiol. 2013, 3, 19839. [Google Scholar] [CrossRef]
- Guzman, M.G.; Halstead, S.B.; Artsob, H.; Buchy, P.; Farrar, J.; Gubler, D.J.; Hunsperger, E.; Kroeger, A.; Margolis, H.S.; Martínez, E.; et al. Dengue: A continuing global threat. Nat. Rev. Microbiol. 2010, 8, S7–S16. [Google Scholar] [CrossRef] [PubMed]
- Bhatt, S.; Gething, P.W.; Brady, O.J.; Messina, J.P.; Farlow, A.W.; Moyes, C.L.; Drake, J.M.; Brownstein, J.S.; Hoen, A.G.; Sankoh, O.; et al. The global distribution and burden of dengue. Nature 2013, 496, 504–507. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilder-Smith, A.; Gubler, D.J. Dengue vaccines at a crossroad. Science 2015, 350, 626–627. [Google Scholar] [CrossRef]
- Wang, P.; Zhang, C.; Guo, M.; Guo, S.; Zhu, Y.; Zheng, J.; Zhu, L.; Ruan, L.; Peng, D.; Sun, M. Complete genome sequence of Bacillus thuringiensis ybt-1518, a typical strain with high toxicity to nematodes. J. Biotechnol. 2014, 171, 1–2. [Google Scholar] [CrossRef]
- Bravo, A.; Gill, S.S.; Soberón, M. Mode of action of Bacillus thuringiensis Cry and Cyt toxins and their potential for insect control. Toxicon 2007, 49, 423–435. [Google Scholar] [CrossRef] [PubMed]
- Palma, L.; Muñoz, D.; Berry, C.; Murillo, J.; Caballero, P. Bacillus thuringiensis toxins: An overview of their biocidal activity. Toxins 2014, 6, 3296–3325. [Google Scholar] [CrossRef] [PubMed]
- Poncet, S.; Bernard, C.; Dervyn, E.; Cayley, J.; Klier, A.; Rapoport, G. Improvement of Bacillus sphaericus toxicity against dipteran larvae by integration, via homologous recombination, of the Cry11A toxin gene from Bacillus thuringiensis subsp. israelensis. Appl. Environ. Microbiol. 1997, 63, 4413–4420. [Google Scholar] [PubMed]
- Zhang, L.; Huang, E.; Lin, J.; Gelbič, I.; Zhang, Q.; Guan, Y.; Huang, T.; Guan, X. A novel mosquitocidal Bacillus thuringiensis strain LLP29 isolated from the phylloplane of Magnolia denudata. Microbiol. Res. 2010, 165, 133–141. [Google Scholar] [CrossRef] [PubMed]
- Johnson, S.; Dureja, P.; Dhingra, S. Photostabilizers for azadirachtin-a (a neem-based pesticide). J. Environ. Sci. 2003, 38, 451–462. [Google Scholar] [CrossRef]
- Fernandez, L.E.; Aimanova, K.G.; Gill, S.S.; Bravo, A.; Soberón, M. A GPI-anchored Alkaline phosphatase is a functional midgut receptor of Cry11Aa toxin in Aedes aegypti larvae. Biochem. J. 2006, 394, 77–84. [Google Scholar] [CrossRef]
- Park, Y.; González-Martínez, R.M.; Navarro-Cerrillo, G.; Chakroun, M.; Kim, Y.; Ziarsolo, P.; Blanca, J.; Cañizares, J.; Ferré, J.; Herrero, S. ABCC transporters mediate insect resistance to multiple Bt toxins revealed by bulk segregant analysis. BMC Biol. 2014, 12, 46. [Google Scholar] [CrossRef]
- Guo, Z.; Kang, S.; Chen, D.; Wu, Q.; Wang, S.; Xie, W.; Zhu, X.; Baxter, S.W.; Zhou, X.; Jurat-Fuentes, J.L.; et al. MAPK signaling pathway alters expression of midgut ALP and ABCC genes and causes resistance to Bacillus thuringiensis Cry1Ac toxin in diamondback moth. PLoS Genet. 2015, 11, 1–32. [Google Scholar] [CrossRef]
- Xia, X.; You, M.; Rao, X.J.; Yu, X.Q. Insect C-type lectins in innate immunity. Dev. Comp. Immunol. 2018, 83, 70–79. [Google Scholar] [CrossRef]
- Weis, W.I.; Taylor, M.E.; Drickamer, K. The C-type lectin superfamily in the immune system. Immunol. Rev. 1998, 163, 19–34. [Google Scholar] [CrossRef]
- Mayer, S.; Raulf, M.-K.; Lepenies, B. C-type lectins: Their network and roles in pathogen recognition and immunity. Histochem. Cell Biol. 2017, 147, 223–237. [Google Scholar] [CrossRef] [PubMed]
- Shi, X.Z.; Yu, X.Q. The extended loop of the C-terminal carbohydrate-recognition domain of Manduca sexta immulectin-2 is important for ligand binding and functions. J. Amino Acids 2012, 42, 2383–2391. [Google Scholar] [CrossRef] [PubMed]
- Drickamer, K. Engineering galactose-binding activity into a C-type mannose-binding protein. Nature 1992, 360, 183–186. [Google Scholar] [CrossRef] [PubMed]
- Rao, X.J.; Cao, X.; He, Y.; Hu, Y.; Zhang, X.; Chen, Y.R.; Blissard, G.; Kanost, M.R.; Yu, X.Q.; Jiang, H. Structural features, evolutionary relationships, and transcriptional regulation of C-type lectin-domain proteins in Manduca sexta. Insect Biochem. Mol. Biol. 2015, 62, 75–85. [Google Scholar] [CrossRef] [PubMed]
- Ao, J.; Ling, E.; Yu, X.Q. Drosophila C-type lectins enhance cellular encapsulation. Mol. Immunol. 2007, 44, 2541–2548. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schnitger, A.K.; Yassine, H.; Kafatos, F.C.; Osta, M.A. Two C-type lectins cooperate to defend Anopheles gambiae against gram-negative bacteria. J. Biol. Chem. 2009, 284, 17616–17624. [Google Scholar] [CrossRef]
- Chai, L.Q.; Tian, Y.Y.; Yang, D.T.; Wang, J.X.; Zhao, X.F. Molecular cloning and characterization of a C-type lectin from the cotton bollworm, Helicoverpa armigera. Dev. Comp. Immunol. 2008, 32, 71–83. [Google Scholar] [CrossRef]
- Hillyer, J.F. Mosquito immunity. Adv. Exp. Med. Biol. 2010, 708, 218–238. [Google Scholar]
- Hoffmann, J.A. The immune response of drosophila. Nature 2003, 426, 33–38. [Google Scholar] [CrossRef]
- Iwanaga, S.; Lee, B.L. Recent advances in the innate immunity of invertebrate animals. Biochem. Mol. Biol. J. 2005, 38, 128–150. [Google Scholar] [CrossRef]
- Osta, M.A.; Christophides, G.K.; Kafatos, F.C. Effects of mosquito genes on plasmodium development. Science 2004, 303, 2030–2032. [Google Scholar] [CrossRef] [PubMed]
- Aliota, M.T.; Fuchs, J.F.; Mayhew, G.F.; Chen, C.C.; Christensen, B.M. Mosquito transcriptome changes and filarial worm resistance in Armigeres subalbatus. BMC Genom. 2007, 8, 463. [Google Scholar] [CrossRef] [PubMed]
- Christensen, B.M.; Li, J.; Chen, C.C.; Nappi, A.J. Melanization immune responses in mosquito vectors. Trends Parasitol. 2005, 21, 192–199. [Google Scholar] [CrossRef] [PubMed]
- Waterhouse, R.M.; Kriventseva, E.V.; Meister, S.; Xi, Z.; Alvarez, K.S.; Bartholomay, L.C.; Barillas-Mury, C.; Bian, G.; Blandin, S.; Christensen, B.M.; et al. Evolutionary dynamics of immune-related genes and pathways in disease-vector mosquitoes. Science 2007, 316, 1738–1743. [Google Scholar] [CrossRef] [PubMed]
- Yassine, H.; Osta, M.A. Anopheles gambiae innate immunity. Cell Microbiol. 2010, 12, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Zhao, G.; Hu, X.; Liu, J.; Li, M.; Batool, K.; Chen, M.; Wang, J.; Xu, J.; Huang, T.; et al. Cry11Aa interacts with the ATP-binding protein from Culex quinquefasciatus to improve the toxicity. J. Agric. Food Chem. 2017, 65, 10884–10890. [Google Scholar] [CrossRef] [PubMed]
- Ben-Dov, E. Bacillus thuringiensis subsp. israelensis and its dipteran-specific toxins. Toxins 2014, 6, 1222–1243. [Google Scholar] [CrossRef]
- Chilcott, C.N.; Ellar, D.J. Comparative toxicity of Bacillus thuringiensis var. israelensis crystal proteins in vivo and in vitro. J. Gen. Microbiol. 1988, 134, 2551–2558. [Google Scholar]
- Despres, L.; Stalinski, R.; Tetreau, G.; Paris, M.; Bonin, A.; Navratil, V.; Reynaud, S.; David, J.P. Gene expression patterns and sequence polymorphisms associated with mosquito resistance to Bacillus thuringiensis israelensis toxins. BMC Genom. 2014, 15, 926. [Google Scholar] [CrossRef]
- Lee, S.B.; Aimanova, K.G.; Gill, S.S. Alkaline phosphatases and Aminopeptidases are altered in a Cry11Aa resistant strain of Aedes aegypti. Insect Biochem. Mol. Biol. 2014, 54, 112–121. [Google Scholar] [CrossRef]
- Bourne, Y.; Roig-Zamboni, V.; Barre, A.; Peumans, W.J.; Astoul, C.H.; Van Damme, E.J.; Rougé, P. The crystal structure of the Calystegiasepium agglutinin reveals a novel quaternary arrangement of lectin subunits with a β-prism fold. J. Biol. Chem. 2004, 279, 527–533. [Google Scholar] [CrossRef]
- Meagher, J.L.; Winter, H.C.; Ezell, P.; Goldstein, I.J.; Stuckey, J.A. Crystal structure of banana lectin reveals a novel second sugar binding site. Glycobiology 2005, 15, 1033–1042. [Google Scholar] [CrossRef] [Green Version]
- Darboux, I.; Nielsen-LeRoux, C.; Charles, J.F.; Pauron, D. The receptor of Bacillus sphaericus binary toxin in Culex pipiens (diptera: Culicidae) midgut: Molecular cloning and expression. Insect Biochem. Mol. Biol. 2001, 31, 981–990. [Google Scholar] [CrossRef]
- Fernandez, L.E.; Martinez-Anaya, C.; Lira, E.; Chen, J.; Evans, A.; Hernandez-Martinez, S.; Lanz-Mendoza, H.; Bravo, A.; Gill, S.S.; Soberon, M. Cloning and epitope mapping of Cry11Aa-binding sites in the Cry11Aa-receptor Alkaline phosphatase from Aedes aegypti. Biochemistry 2009, 48, 8899–8907. [Google Scholar] [CrossRef] [PubMed]
- Lereclus, D.; Agaisse, H.; Gominet, M.; Chaufaux, J. Overproduction of encapsulated insecticidal crystal proteins in a Bacillus thuringiensis spo0a mutant. Biotechnology (N.Y.) 1995, 13, 67–71. [Google Scholar]
- Chen, J.; Aimanova, K.; Gill, S.S. Functional characterization of Aedes aegypti Alkaline phosphatase ALP1 involved in the toxicity of cry toxins from Bacillus thuringiensis subsp. Israelensis and jegathesan. Peptides 2017, 98, 78–85. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Aimanova, K.G.; Pan, S.; Gill, S.S. Identification and characterization of Aedes aegypti Aminopeptidase N as a putative receptor of Bacillus thuringiensis Cry11A toxin. Insect Biochem. Mol. Biol. 2009, 39, 688–696. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Likitvivatanavong, S.; Aimanova, K.G.; Gill, S.S. A 104 kDa Aedes aegypti Aminopeptidase N is a putative receptor for the Cry11Aa toxin from Bacillus thuringiensis subsp. israelensis. Insect Biochem. Mol. Biol. 2013, 43, 1201–1208. [Google Scholar] [CrossRef] [PubMed]
- Dai, S.M.; Gill, S.S. In vitro and in vivo proteolysis of the Bacillus thuringiensis subsp. israelensis CryIVD protein by Culex quinquefasciatus larval midgut proteases. Insect Biochem. Mol. Biol. 1993, 23, 273–283. [Google Scholar]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. Mega6: Molecular evolutionary genetics analysis version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef]
- Carroll, J.; Ellar, D.J. An analysis of Bacillus thuringiensis delta-endotoxin action on insect-midgut-membrane permeability using a light-scattering assay. Eur. J. Biochem. 1993, 214, 771–778. [Google Scholar] [CrossRef] [PubMed]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]





© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Batool, K.; Alam, I.; Zhao, G.; Wang, J.; Xu, J.; Yu, X.; Huang, E.; Guan, X.; Zhang, L. C-Type Lectin-20 Interacts with ALP1 Receptor to Reduce Cry Toxicity in Aedes aegypti. Toxins 2018, 10, 390. https://doi.org/10.3390/toxins10100390
Batool K, Alam I, Zhao G, Wang J, Xu J, Yu X, Huang E, Guan X, Zhang L. C-Type Lectin-20 Interacts with ALP1 Receptor to Reduce Cry Toxicity in Aedes aegypti. Toxins. 2018; 10(10):390. https://doi.org/10.3390/toxins10100390
Chicago/Turabian StyleBatool, Khadija, Intikhab Alam, Guohui Zhao, Junxiang Wang, Jin Xu, Xiaoqiang Yu, Enjiong Huang, Xiong Guan, and Lingling Zhang. 2018. "C-Type Lectin-20 Interacts with ALP1 Receptor to Reduce Cry Toxicity in Aedes aegypti" Toxins 10, no. 10: 390. https://doi.org/10.3390/toxins10100390