1. Introduction
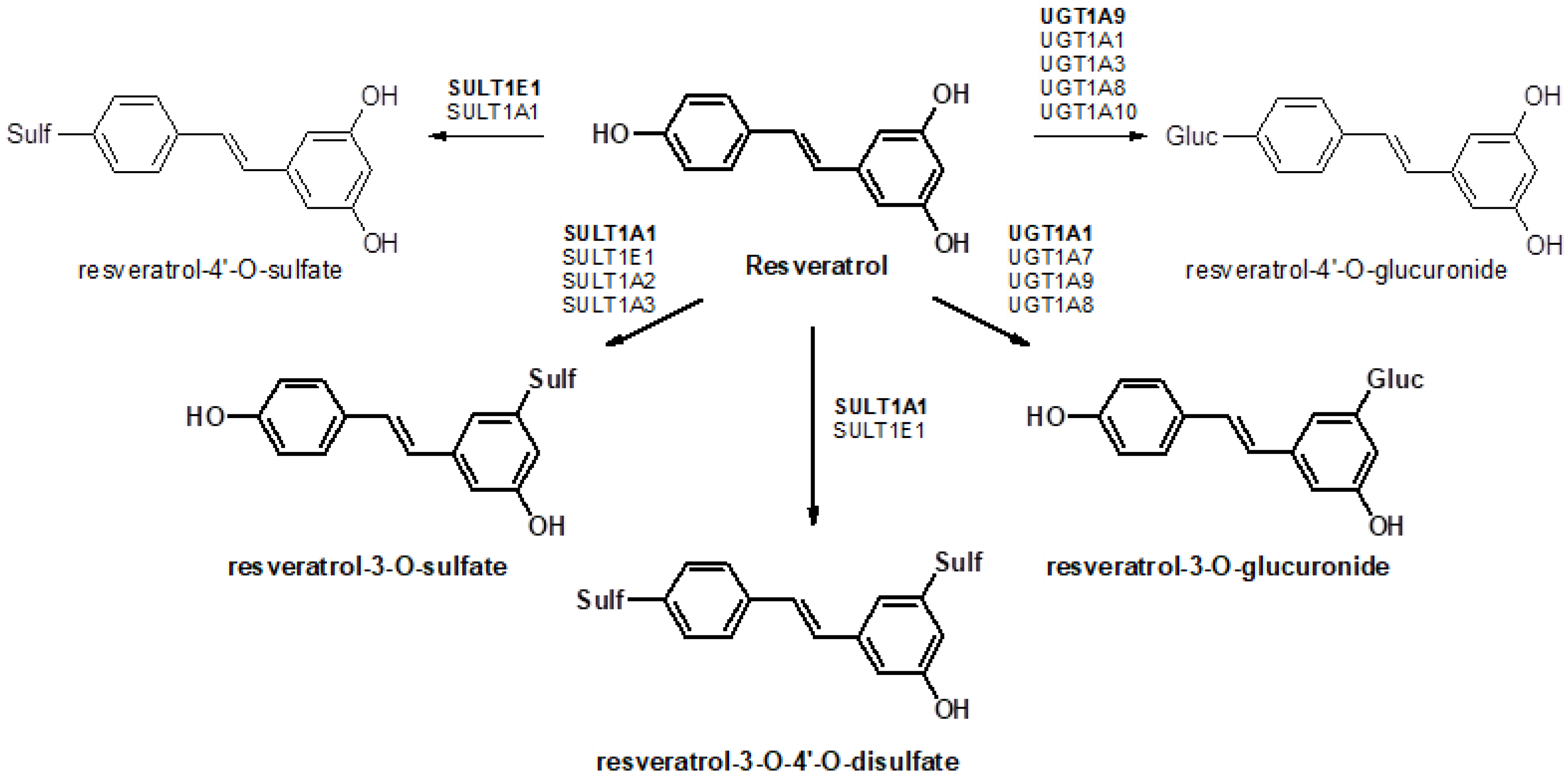
Resveratrol (3,4′,5-trihydroxy-trans-stilbene) is a naturally occurring compound that is produced by a wide variety of plants, berries, and fruits, and is mainly found in the skin of grapes and in red wine [
1]. Interest in resveratrol has increased due to various studies reporting its role in improving the outcomes of various pathologies, e.g., cardiovascular and neurodegenerative diseases, inflammation, viral infections, and cancer [
2,
3,
4]. These beneficial effects on health are observed despite extremely low bioavailability and rapid clearance from the circulation [
4,
5], based on extensive metabolism in humans and animal models [
6,
7,
8,
9]. Previously, data from our laboratory demonstrated that resveratrol is biotransformed in the rat liver into six different products [
10], with resveratrol-3-O-glucuronide, resveratrol-3-O-4′-O-disulfate, and resveratrol-3-O-sulfate being the most abundant metabolites. In humans, resveratrol glucuronides are the predominant biotransformation products when low doses of resveratrol are ingested, whereas for higher doses, sulfates are mainly generated [
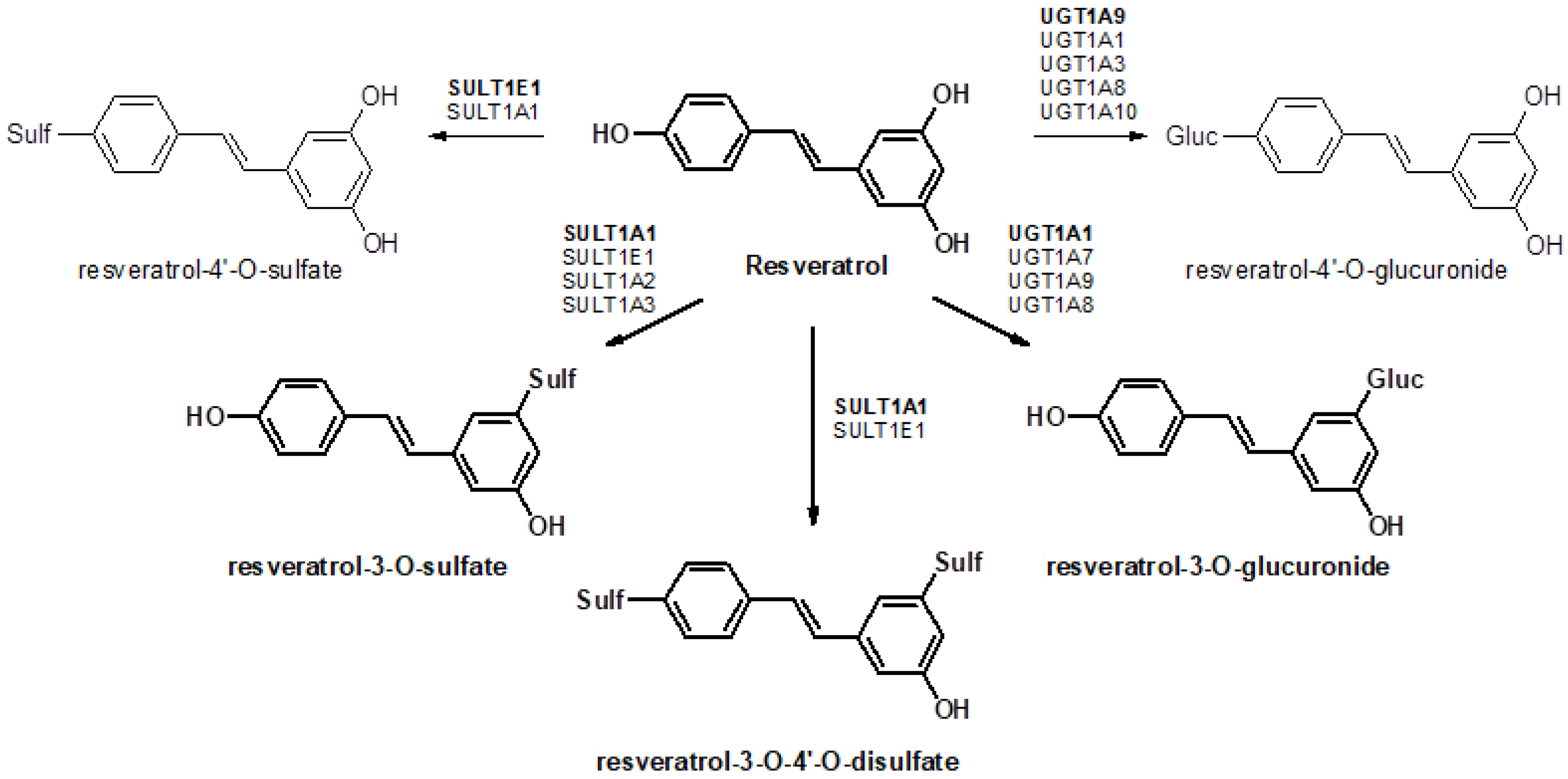
11]. As most resveratrol conjugates have decreased pharmacological activity, first-pass metabolism in the gut and liver has a significant influence on the efficacy of resveratrol. Sulfation and glucuronidation of resveratrol are carried out by UDP-glucuronosyltransferases (UGTs) and cytosolic sulfotransferases (SULTs), respectively, and the expression levels of these enzymes have a major influence on the profile of resveratrol conjugates generated in different organs of the body (
Figure 1). Therefore, it is important to determine the tissue-specific expression patterns of these enzymes. Previous investigations have mainly focused on the analysis of resveratrol and its metabolites in plasma, urine, or feces [
4,
6,
12], as well as the distribution in various organs [
9,
13,
14]. To the best of our knowledge, there are no data in the literature regarding the expression of resveratrol-metabolizing enzymes in tissue samples and their correlation with resveratrol concentrations.
Previously, we have shown that resveratrol is selectively metabolized to resveratrol-3-O-sulfate in 12 of 13 human breast cancer tissue specimens [
15]. Reverse transcription quantitative PCR (RT-qPCR) analysis of paired samples from control and tumor tissues revealed mRNA expression of SULT1A2, SULT1A3, and SULT1E1, which have all been shown to catalyze resveratrol biotransformation [
16]. As SULT1A1 mRNA expression was below the detection limit in all samples and SULT1E1 mRNA was only detectable in a few samples, the cellular localization of SULT1A3 was assessed by indirect immunofluorescence on paraffin-embedded sections from control and malignant breast tissues, clearly indicating a correlation of RT-qPCR data with the protein expression of this enzyme. Other studies identified the UGT1A family, including UGT1A1, UGT1A6, UGT1A7, UGT1A9, and UGT1A10, to be responsible for catalyzing the formation of resveratrol-3-
O-glucuronide and resveratrol-4′-
O-glucuronide [
17,
18].
However, no data are available regarding the association of UGT and SULT mRNA expression with resveratrol glucuronidation and sulfation in tissue samples. Therefore, in the present study, we conducted feeding studies in mice, and time-dependently quantified resveratrol and its main glucuronides and sulfates in plasma and selected tissues by using a sensitive high-performance liquid chromatography (HPLC) assay. Furthermore, the mRNA expression levels of the major isoenzymes responsible for resveratrol metabolism were investigated in order to determine the association between tissue-specific expression and metabolite formation.
2. Material and Methods
2.1. Chemicals
Resveratrol (3,4′,5-trihydroxy-trans-stilbene) was obtained from Sigma-Aldrich (Munich, Germany). Resveratrol-3-O-glucuronide, resveratrol-3-O-sulfate, and resveratrol-3-O-4′-O-disulfate were obtained from Santa Cruz Biotechnology (Dallas, TX, USA). Methanol and water were of HPLC grade and were purchased from Merck (Darmstadt, Germany). All other chemicals and solvents were commercially available, of analytical grade, and used without further purification.
2.2. Animal Experiments
Male wild-type C57BL/6 Oca2p−un (weight, 23–28 g) were obtained from the University of California at Los Angeles (UCLA). Animals were housed in a temperature-, light-, and humidity-controlled environment and fed a SSNIFF™ standard diet and water (pH 3.5) ad libitum. Resveratrol, as a PBS solution containing 1% Tween 20 and 1% ethanol, was administered intragastrically by oral gavage feeding in overnight-fasted mice (aged 1.5–2.5 months) at a single dose of 10 mg/kg resveratrol. Blood samples (300 µL each) were obtained prior to sacrifice of the animals, and were collected into EDTA-coated tubes. For the tissue distribution study, groups of mice (n = 3 per group) were sacrificed at 0.5, 1, 1.5, 2, 4, 6, and 8 h post-dosing, respectively. Subsequently, the spleen, liver, kidney, heart, lung, stomach, duodenum, jejunum, ileum, cecum, colon, thymus, muscle, and brain were rapidly excised and washed with NaCl (0.9%) to remove residual blood containing resveratrol and its metabolites. Tissues were wiped with filter paper, weighed, immediately frozen in liquid nitrogen, and maintained at −80 °C until homogenization. For RT-qPCR studies, groups of four mice were used without previous gavage. The experimental protocol was approved by the Committee on the Ethics of the Austrian Ministry of Science (No. 66.009/122-II/10b/2010).
2.3. Sample Preparation
Following the addition of 200 μL of methanol to 100 μL of each plasma sample, samples were centrifuged and an 80 μL aliquot of each supernatant was injected into the HPLC column. Mouse tissue samples were thawed on ice at 4 °C, mixed with a threefold amount of 5 mM ammonium acetate/acetic acid buffer (pH 7.4), and then minced using an Ultra-Turrax®-homogenizer, four times for 10 s each, on ice. Tissue homogenates were first centrifuged at 13,000× g for 10 min, and proteins were eliminated through a second centrifugation by adding methanol (500 µL) to 250 µL of the supernatant. Subsequently, 80 µL of the remaining supernatant was injected onto the HPLC column. In all cases, sample manipulation was performed while avoiding direct light exposure to prevent possible photochemical isomerization of trans-resveratrol to the cis form.
2.4. High-Performance Liquid Chromatography (HPLC) Analysis
Resveratrol and its glucuronidated and sulfated biotransformation products were quantified by HPLC as described previously [
19] using a Dionex UltiMate 3000 system (Sunnyvale, CA, USA) equipped with an L-7250 injector, an L-7100 pump, an L-7300 column oven (set at 15 °C), a D-7000 interface, and an L-7400 UV detector (Thermo Fisher Scientific, Waltham, MA, USA) set at a wavelength of 307 nm. Calibration of the chromatogram was accomplished using the external standard method. Linear calibration curves were performed by spiking drug-free cell culture medium with standard solutions of resveratrol, resveratrol-3-
O-sulfate, resveratrol-3-
O-4′-
O-disulfate, and resveratrol-3-
O-glucuronide to give a concentration range from 0.001 to 10 µg/mL (average correlation coefficients: >0.999). For this method, the lower limits of quantification for resveratrol and resveratrol conjugates were 5 ng and 7 ng, respectively. Coefficients of accuracy and precision for these compounds were <11%.
2.5. Reverse Transcription Quantitative PCR (RT-qPCR)
Total RNA was extracted from mouse tissue samples using peqGOLD Trifast™ reagent (peqLAB, Erlangen, Germany), according to the manufacturer’s instructions. The concentration, purity, and integrity of RNA samples were determined by UV absorbance and gel electrophoresis. Total RNA samples (2 µg) were reverse transcribed to cDNA using random hexamer primers and a MultiScribe™ Reverse Transcriptase Kit (ThermoFisher Scientific, Waltham, MA, USA), in accordance with the manufacturer’s instructions. RT-qPCR was performed using RT2 Profiler PCR Arrays (PAMM-069ZA, Qiagen, Hilden, Germany) and RT2 SYBR Green qPCR Master Mix (Qiagen, Hilden, Germany) on 96-well plates in an ABI Prism® 7900HT Sequence Detection System (Thermo Fisher Scientific, Waltham, MA, USA) to analyze mouse phase II enzymes involved in drug metabolism. In order to determine appropriate reference genes, 5 different mouse housekeeping genes were included in the arrays, and Hprt (validated with the Mm01545399TaqMan® Gene Expression Assays, ThermoFisher Scientific, Waltham, MA, USA) was selected as the most appropriate reference gene. Gene expression levels were quantified relative to the values obtained for Hprt. Data analyses were done using the web-based analysis software from Quiagen.
2.6. Data Analysis
For the quantifications of resveratrol and its metabolites in plasma, urine, feces, and various organs, a group of three mice was used. The area under the concentration curves from 0 to 8 h (AUCs
0–8h) was calculated using the linear trapezoidal rule with the program Phoenic WinNonlin version 1.5 (Centera Inc., Princeton, NJ, USA). The maximal plasma, urine, feces, and tissue concentration (C
max) and the time after administration of resveratrol when C
max was reached (T
max) were estimated from
Table 1,
Table 2,
Table 3 and
Table 4. All concentrations and the values for C
max and AUC were expressed as mean ± SD calculated using the GraphPad Prism 6.0 (GraphPad, San Diego, CA, USA).
4. Discussion
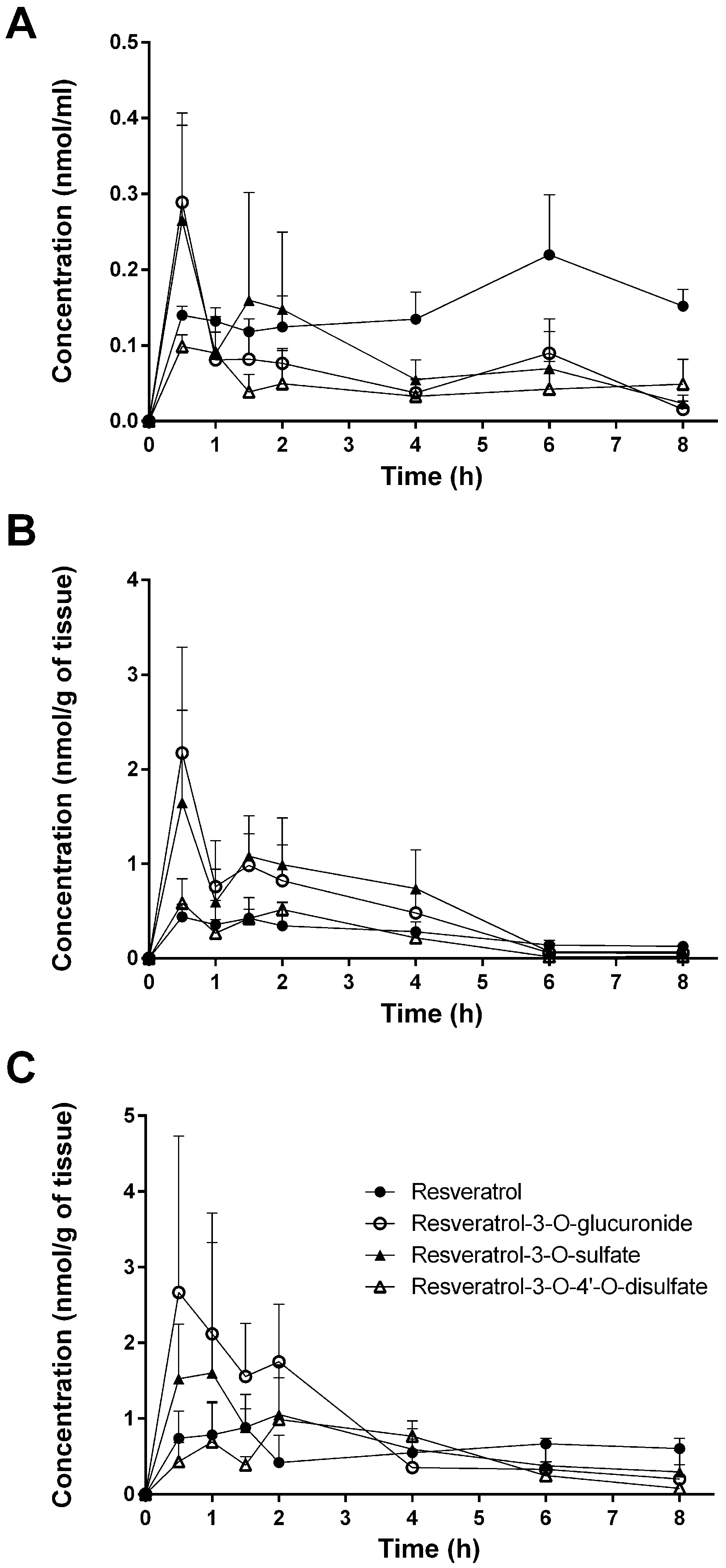
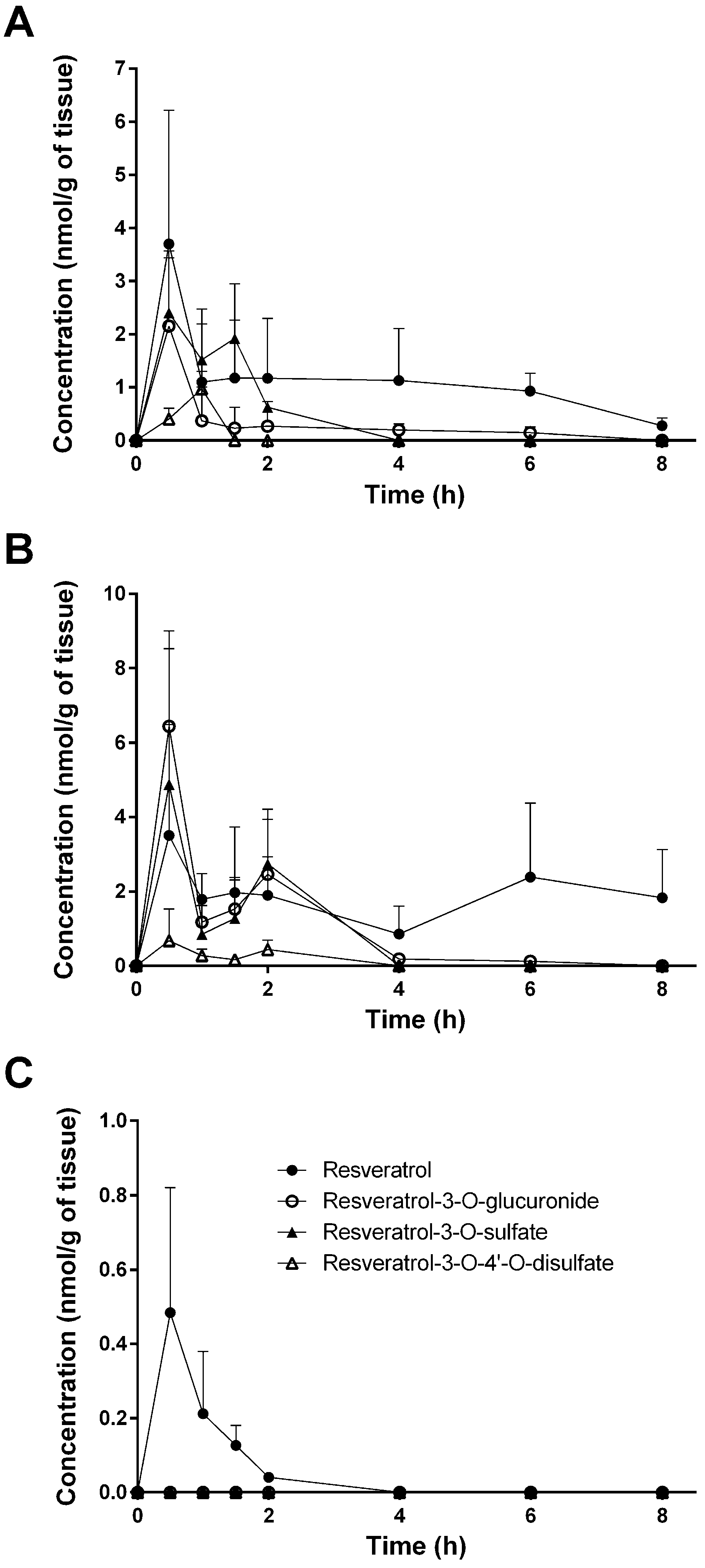
In the present study, tissue distribution of resveratrol and its three major metabolites (resveratrol-3-O-sulfate, resveratrol-3-O-4′-O-disulfate, and resveratrol-3-O-glucuronide) in the plasma, urine, feces, and tissues of mice was investigated in male mice after oral administration of resveratrol (10 mg/kg).
A single dose of 10 mg/kg was chosen based on the daily intake of resveratrol as dietary supplement (100–500 mg/day) to humans. Furthermore, data from our lab also showed that higher resveratrol concentrations inhibit the formation of resveratrol-3-O-sulfate, which might also be observed in mice [
24]. We decided to use male and not female mice in all experiments as recent papers [
25,
26] described significant gender-dependent differences in the glucuronidation of drugs and flavonoids in mice, with a higher formation rate of glucuonides in females than in males, which might be also true for resveratrol. In humans, such pronounced gender-difference in glucuronidation has not been observed so far.
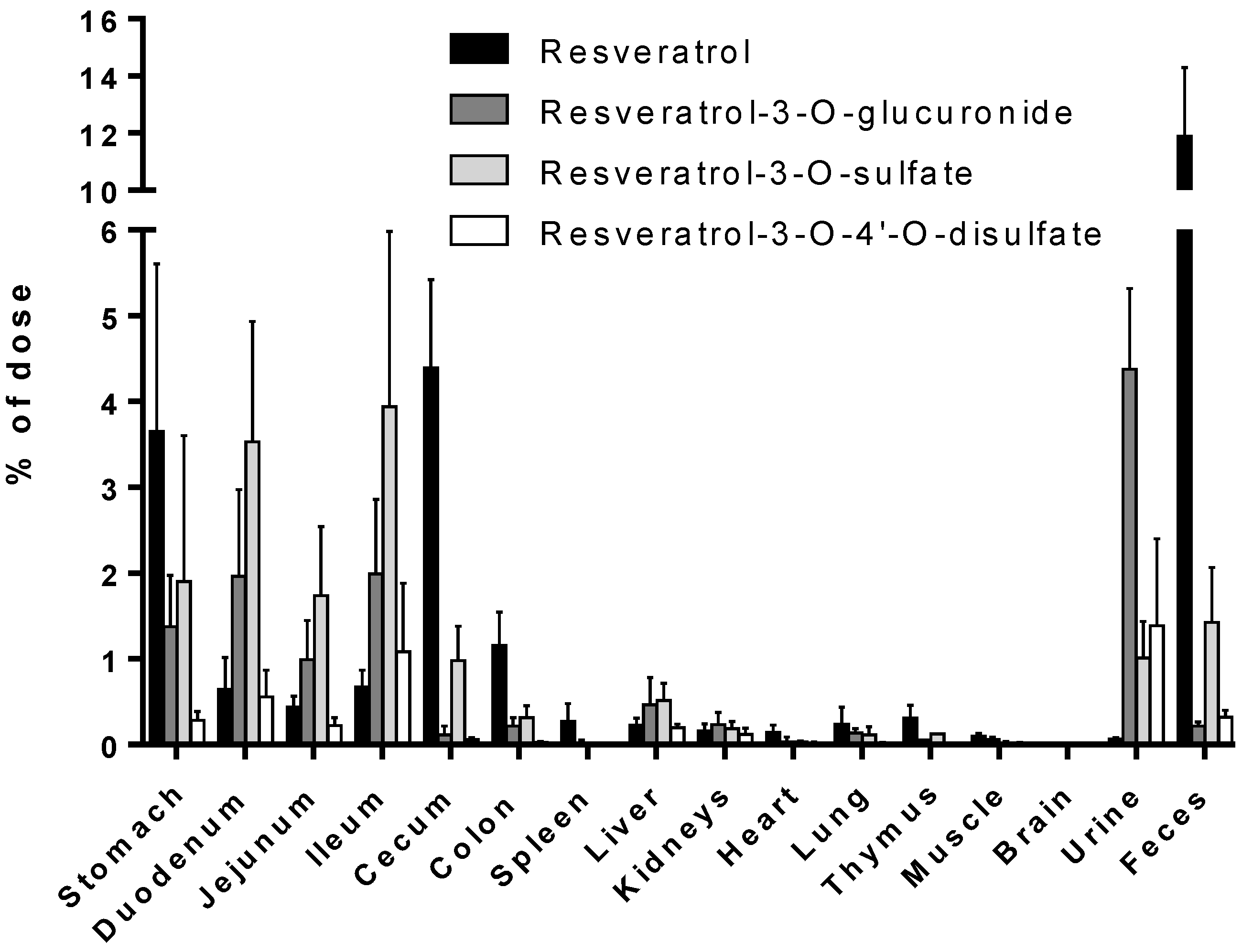
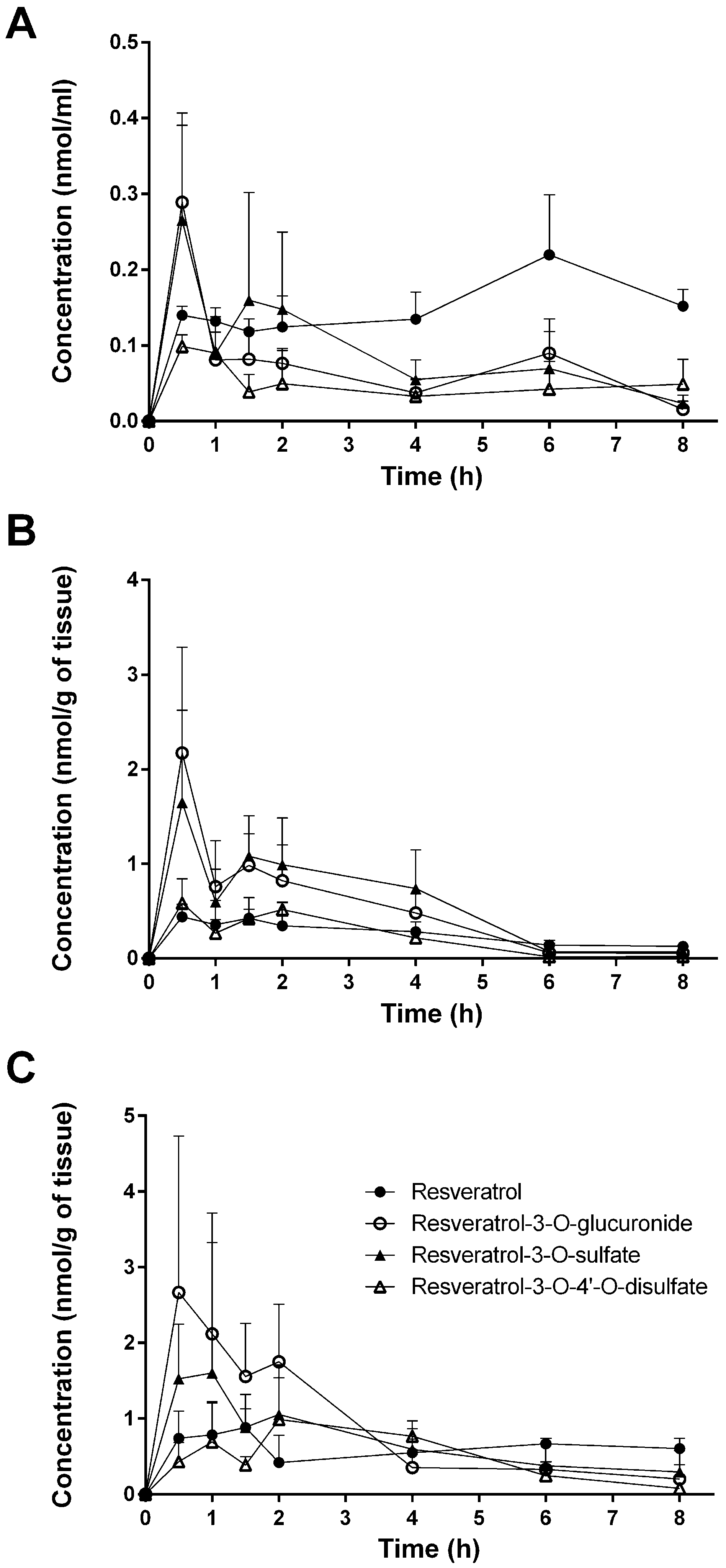
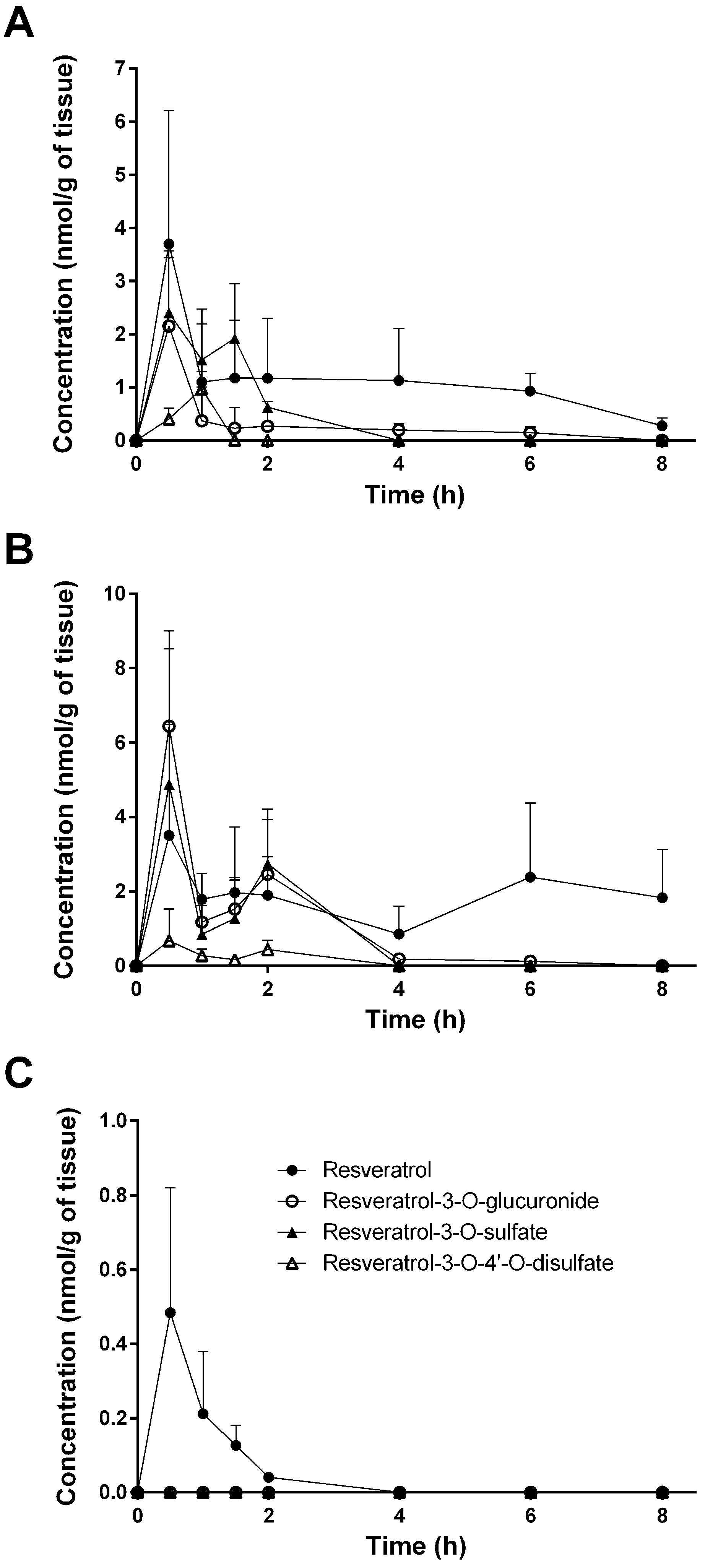
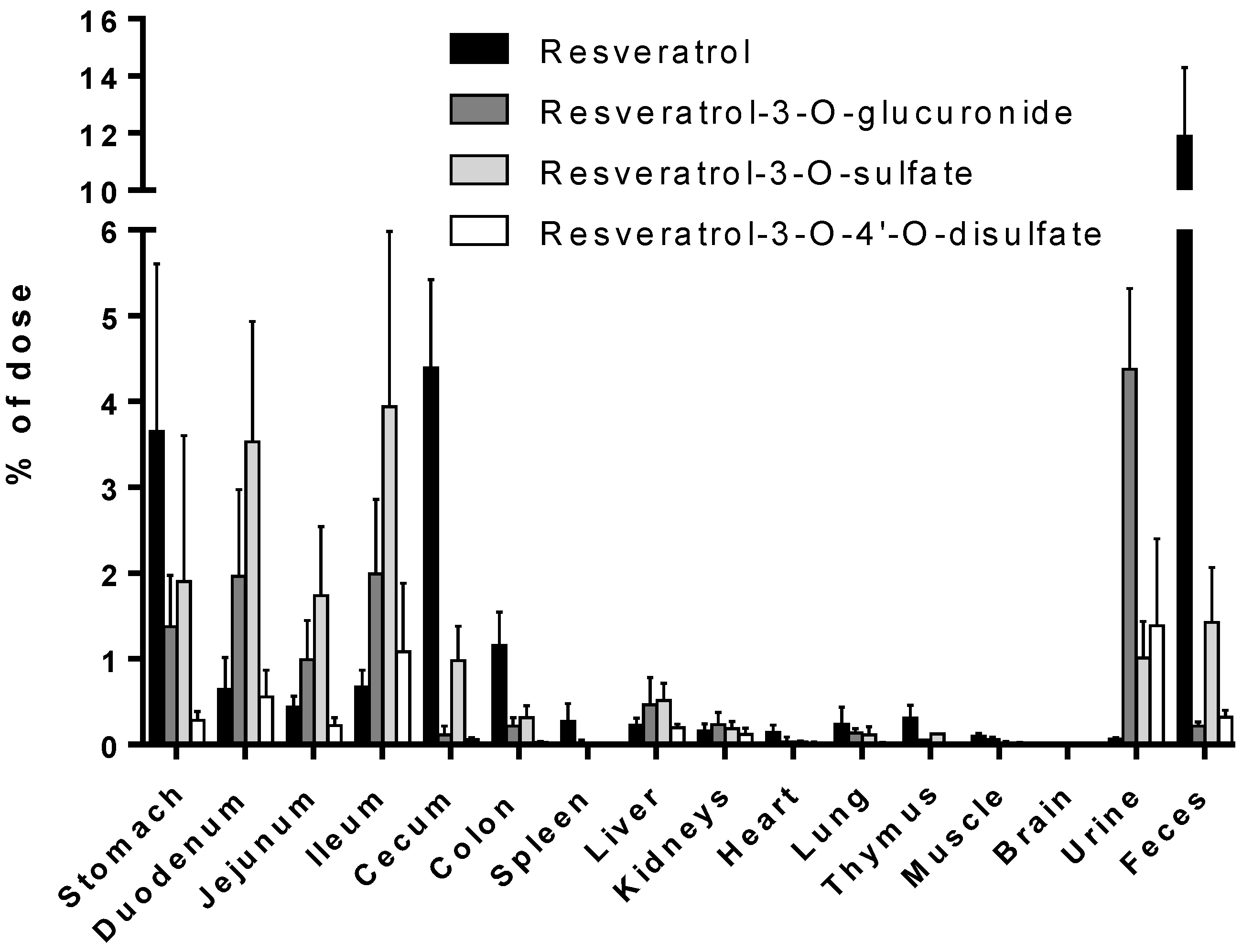
Despite the numerous studies that have investigated the plasmatic levels of resveratrol and its metabolites in mice, very few have attempted to assess the tissue distribution of this natural compound. Therefore, to the best of our knowledge, the present study constitutes the first report of the time-dependent distribution of resveratrol and its major metabolites in 14 tissues as well as in feces and urine in mice following intragastric administration. No data about the concentration of resveratrol and its conjugates in the spleen, thymus, and muscle have been published so far. Furthermore, we also performed, for the first time, a comparison of concentrations with the presence of phase II metabolizing enzymes in appropriate tissues and organs. The liver, kidneys, and gastrointestinal tract are organs with a high phase II metabolizing activity [
27]. After oral administration of resveratrol, our experiments showed that resveratrol was detected predominantly in its glucuronic-acid- and sulfate-conjugated forms in these organs (
Figure 4). High levels of resveratrol conjugates could be detected already in the stomach by resveratrol-metabolizing gastric cells. The lower concentration of the three conjugates in the cecum and colon might be due to a decreased expression of the efflux transporters multidrug resistance protein 2 (MRP2) and breast cancer resistance protein (BCRP), as both membrane proteins are substrates for resveratrol and its conjugates [
28,
29]. Both transporters are able to pump resveratrol glucuronides and sulfates back into the lumen as previously shown in rats [
30]. Interestingly, the overall concentration of resveratrol was higher than that of its metabolites in the stomach, cecum, colon, spleen, heart, lung, thymus, and muscle, as well as in the fecal samples (
Figure 4). A recent paper of Menet et al. [
31] confirmed our results also showing higher resveratrol concentration in mouse heart tissue. This is in contrast to other authors who found higher levels of conjugates in the hearts of rats and pigs [
9,
13].
The same is true for the lung, for which Juan et al. [
32] also showed higher resveratrol concentrations, whereas Lin et al. [
13] could only detect conjugates. Interestingly, in the lung, we could only quantify resveratrol at low levels; the concentration of all three metabolites was below the detection limit. The observed poor accumulation of resveratrol in the brain might be explained by the presence of MRP2 and BCRP in the blood–brain barrier, preventing a pronounced uptake into the brain. No detectable resveratrol conjugates in the brain are in line with findings in pigs shown after intragastric administration of 6.5 mg/kg of resveratrol [
9]. Other authors, however, could not detect resveratrol and its conjugates in rat and mouse brains [
13] or found up to six-fold higher resveratrol-3-
O-sulfate concentration compared with that of native resveratrol [
33]. This discrepancy might be explained by altered doses or different sample preparation steps prior to analysis.
As mentioned above, higher resveratrol concentrations might inhibit resveratrol-3-O-sulfate formation and also change the pattern of metabolites. Indeed, a single dose of 150 mg/kg to mice resulted in up to 5.8-fold higher resveratrol-3-O-glucuronide concentrations in mouse hearts 30 min and 60 min after dosing [
31], whereas our data showed slightly higher concentrations of resveratrol-3-O-sulfate levels at the same time points.
Interestingly, tissue, feces, and urine levels of resveratrol and its conjugates did not show the expected decline with time after dosing. This might be explained either with a delayed transport of resveratrol from the stomach to the colon or by a redistribution between organs. This would explain the high concentration in the thymus for free resveratrol after 8 h. A second peak in the plasma after 6 h for unconjugated resveratrol is possibly caused by enterohepatic recirculation of free resveratrol cleaved by β-glucuronidase from resveratrol 3-O-glucuronide after biliary excretion. The maximum concentration of the final metabolic product resveratrol-3-O-4′-O-sulfate in the urine after 6 h, however, can be explained by its longer formation time starting from resveratrol via resveratrol-3-O-sulfate and resveratrol-4′-O-sulfate.
In humans, resveratrol glucuronidation is mainly catalyzed by UGT1A1 and UGT1A9, and, to a minor extent, by UGT1A6, UGT1A7, and UGT1A10 [
18,
34], whereas sulfation is performed by SULT1A1, SULT1A2, SULT1A3, and SULT1E1 [
15,
16]. Although there are no data available in the literature regarding the conjugation of resveratrol glucuronides and sulfates by mouse-specific Ugts and Sults, several studies have demonstrated high homology between mouse and human isoenzymes, at least for Ugt1a1, Ugt1a6a, and Sult1A1, with a similarity in the amino acid sequence of 75–90% [
35,
36], which suggests comparable enzymatic activity for resveratrol.
In the present study, the formation of resveratrol glucuronides and sulfates correlated with the expression of certain
Ugts and
Sults. RT-qPCR analysis revealed high mRNA expression of
Ugt1a1 and
Ugt1a6a in the liver, duodenum, jejunum, and ileum, leading to high concentrations of resveratrol-3-O-glucuronide in these organs. High expression of
Ugt1a1 and
Ugt1a6a in the liver and small intestines of mice is in accordance with previously reported data [
20,
22]. A good correlation of resveratrol-3-O-sulfate and resveratrol-3-O-4′-O-disulfate formation with
Sult1a1 mRNA expression was also observed, particularly in the liver and colon.
Ugt1a2 and
Ugt2b34 were highly expressed in the liver, duodenum, jejunum, and ileum, whereas Sult4a1 mRNA was almost exclusively detected in the brain. Whether Sult2b1, Ugt1a2, and Ugt2b34 also catalyze resveratrol sulfation and glucuronidation is not yet known. However, based on their high expression in the liver and gastrointestinal tract, we assume the involvement of these isoforms in the metabolism of resveratrol in mouse organs. Our data are in line with previous reports also demonstrating high mRNA expression of
Ugt2b5 and
Ugt2b1 in the liver and of
Ugt2b38 in the kidney [
36]. Contrary to a study by Saiki et al., which reported the expression of Sult1b1 mRNA exclusively in the liver, the present findings revealed
Sult1b1 mRNA expression in the small and large intestines in mice [
37]. This discrepancy may be explained by the fact that the authors used a Northern blot analysis and not the more sensitive RT-qPCR. RT-qPCR analysis may therefore be predictive for the metabolism in tissue samples. Low or no detectable expression of Ugts and Sults, as observed in the spleen, heart, and lung, consequently led to an overall far higher percentage of resveratrol compared with those of its conjugates.
The different distribution of resveratrol and its conjugates in various tissues may be predictive for the observed pharmacological activities of resveratrol. In organs like the spleen, heart, lung, muscle, and brain, resveratrol concentrations might be far too low for any significant biological effect. Although resveratrol glucuronides and sulfates are less active than resveratrol, they may act as a depot, easily converted back to resveratrol by ubiquitously existing human β-glucuronidase and sulfatases. The expression of UGTs and SULTs in the tissues therefore helps to predict the overall pharmacological activity of resveratrol.
Another option to improve oral bioavailibity of resveratrol and, consequently, its efficacy is the inhibition of the glucuronidation in the intestine by UGT-inhibitors. Co-administration of the UGT inhibitor piperin increased the AUC and C
max of resveratrol in rat plasma by 229% and 1544%, respectively [
38]. This pronounced effect was also observed when resveratrol was administered by a nanoemulsion composed of soybean oil, soy lecithin, and the UGT inhibitor labrasol, again increasing the C
max and bioavailability approximately 11- and 5-fold, respectively [
39]. These strategies might help to increase concentrations in organs for which we show minor accumulation of resveratrol.




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}