Ginseng Berry Extract Attenuates Dextran Sodium Sulfate-Induced Acute and Chronic Colitis



Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Chemicals
2.3. Comparison and Standardization of Ginsenoside Composition
2.4. DSS-Induced Acute Colitis
2.5. DSS-Induced Chronic Colitis
2.6. Hematoxylin and Eosin Staining
2.7. Antibodies
2.8. Preparation of Lamina Propria (LP) Single Cell Suspensions
2.9. Ex Vivo Stimulation of LP Cells and Intracellular Cytokine Staining
2.10. Intestinal Macrophage and DC Analysis
2.11. Real-Time qPCR
2.12. ELISA
2.13. In Vitro T Cell Stimulation
2.14. Statistical Analysis
3. Results
3.1. The Micronutrient Content is Higher in GB Than in GR
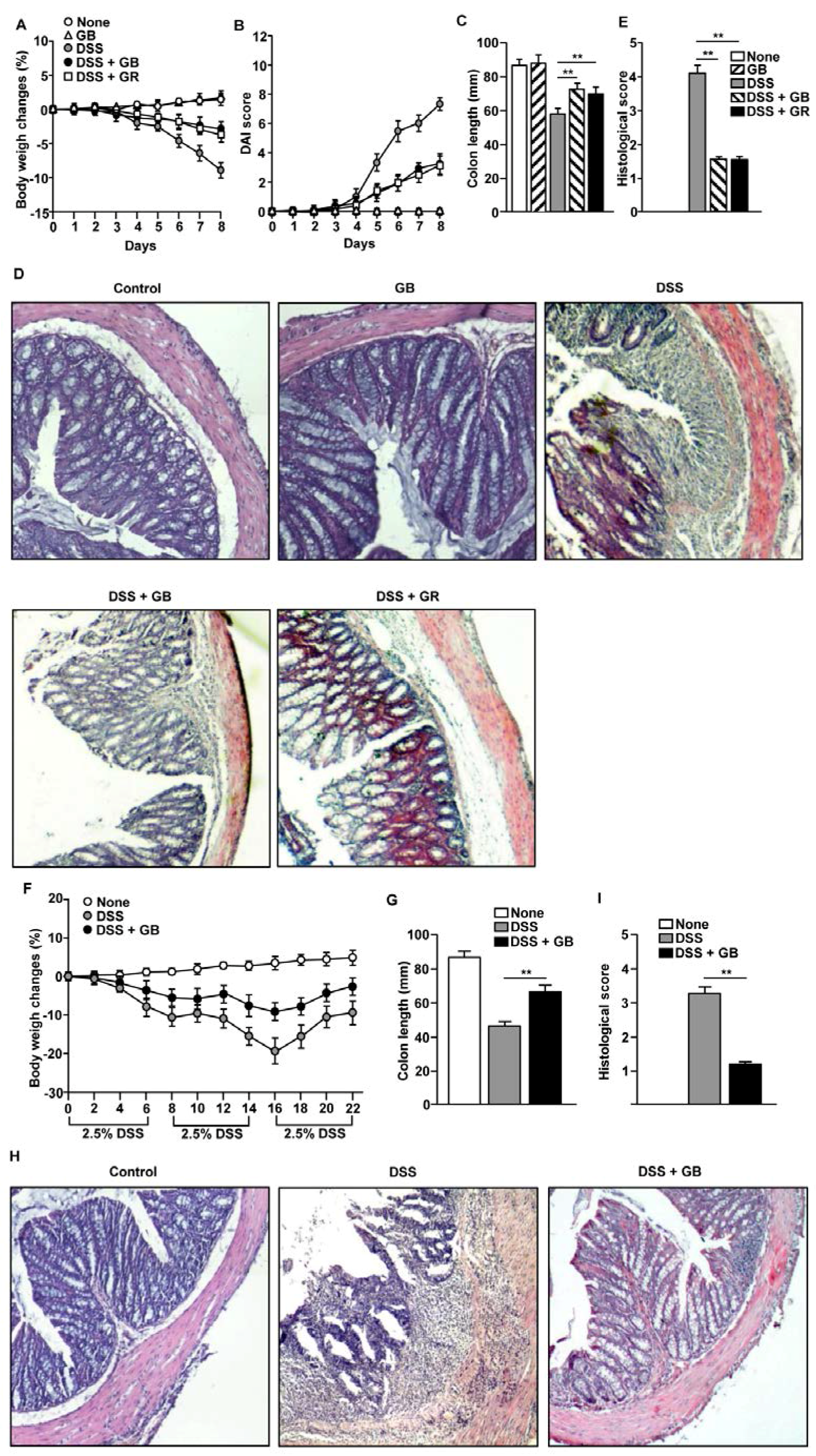
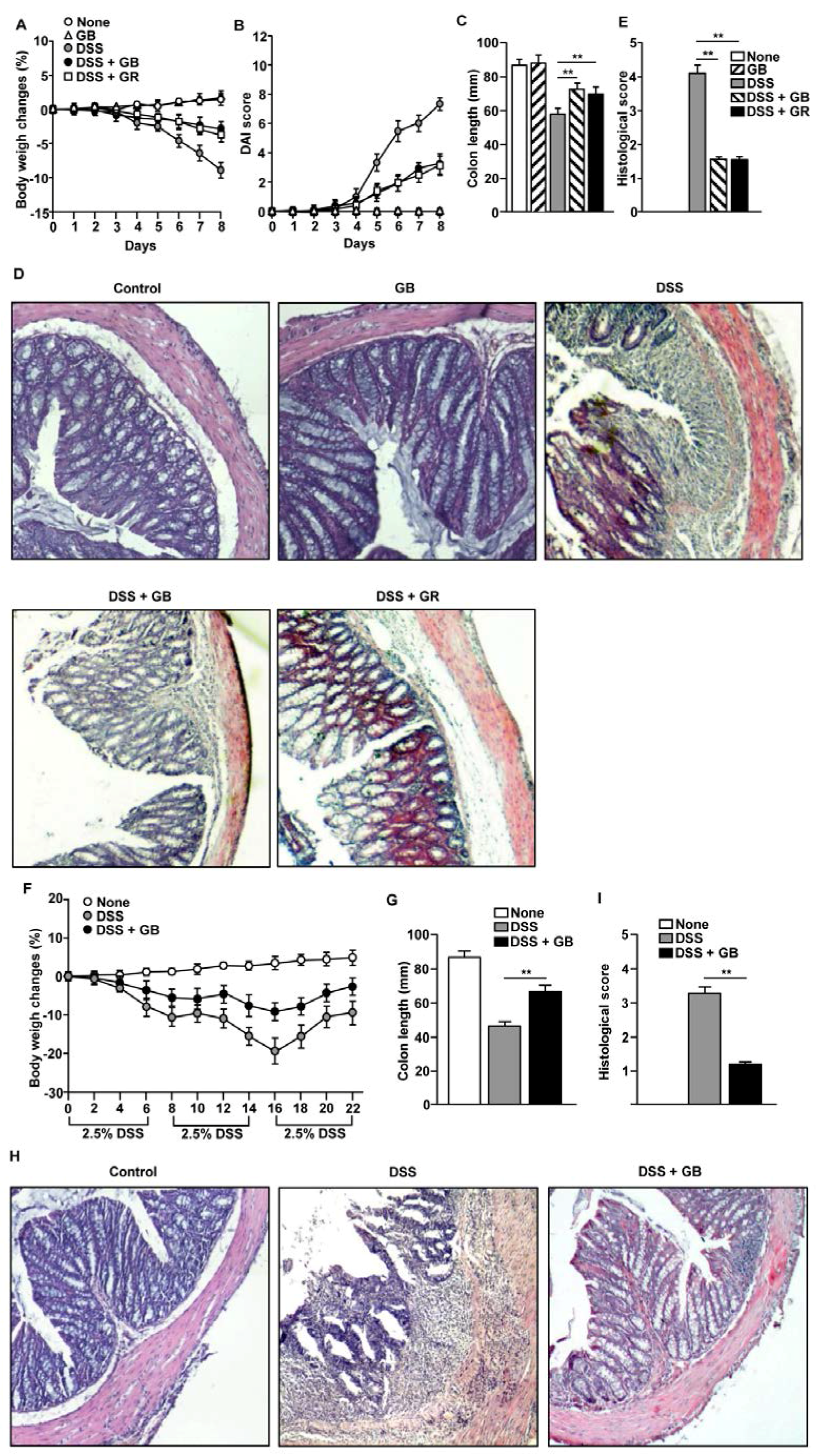
3.2. GB Attenuates DSS-Induced Acute and Chronic Colitis
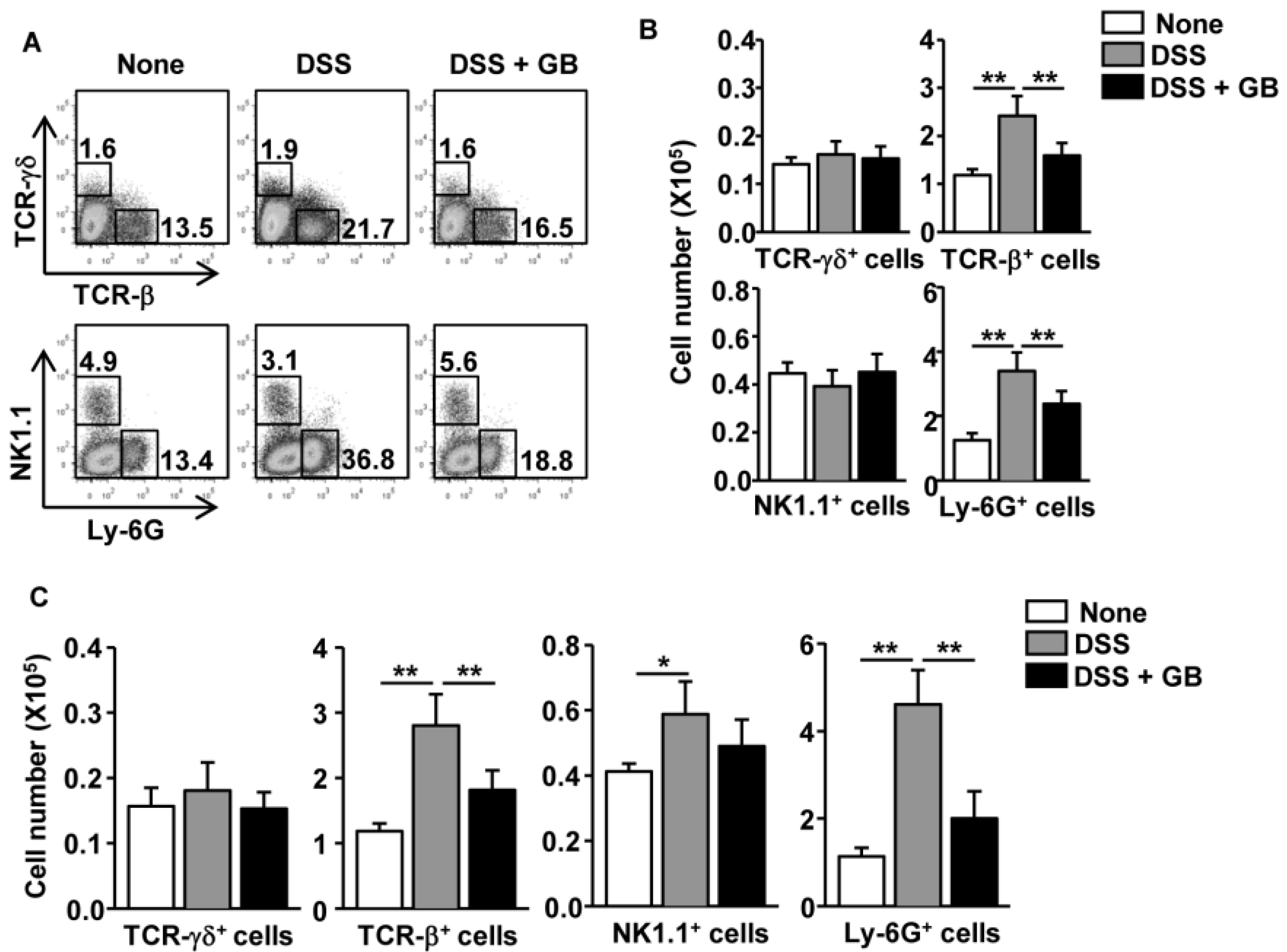
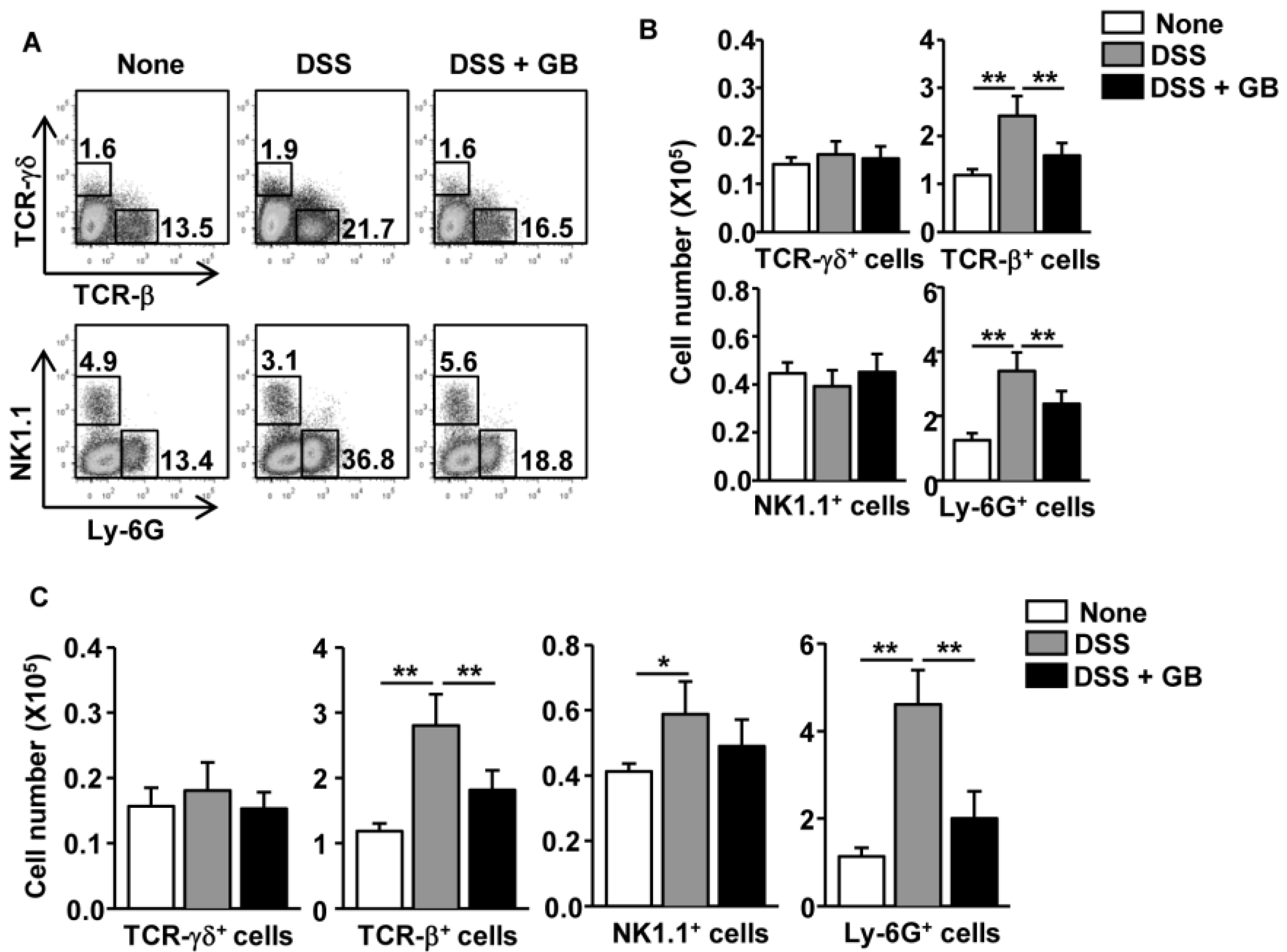
3.3. GB Reduces the Number of T Cells and Neutrophils in the Colon
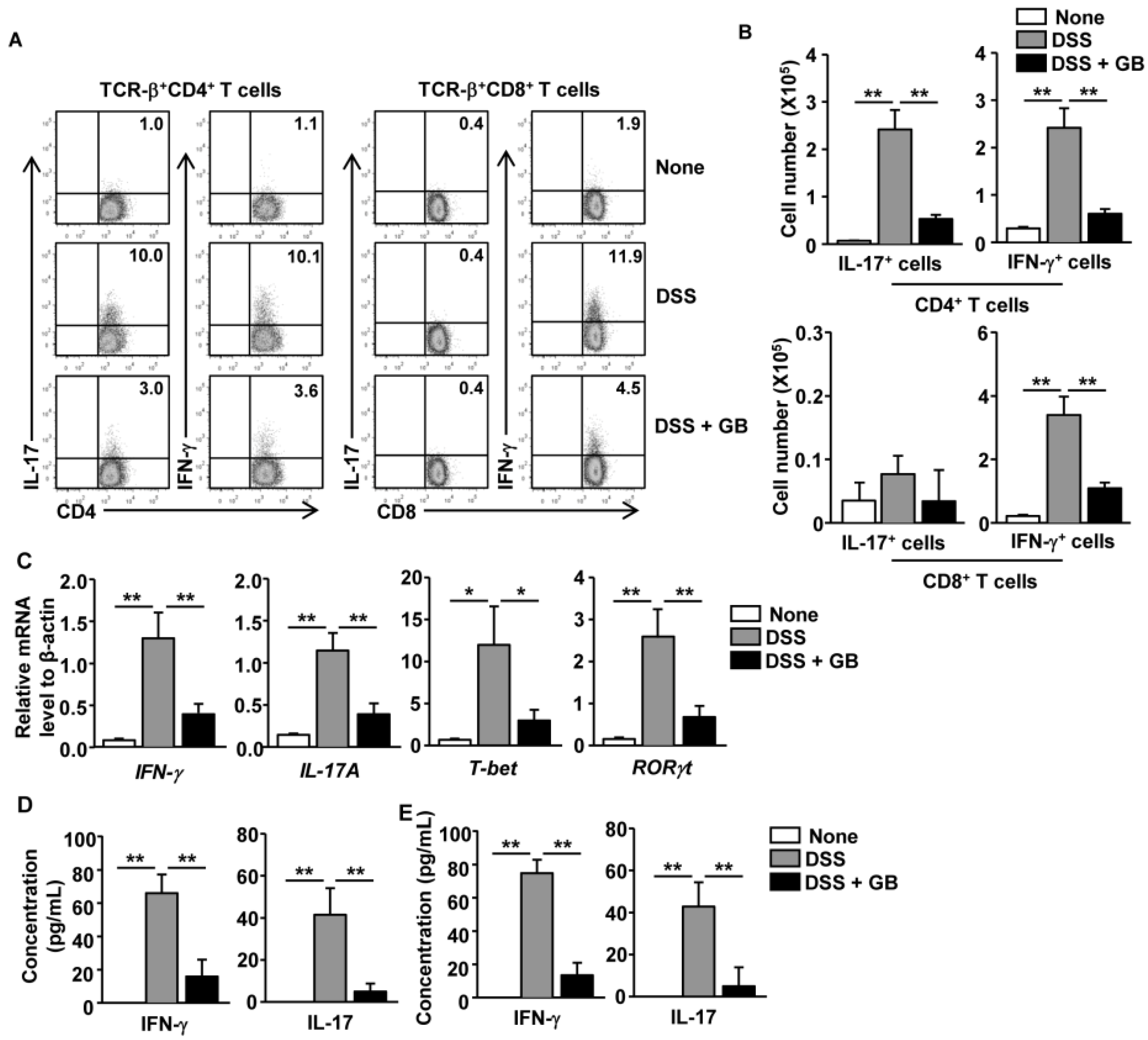
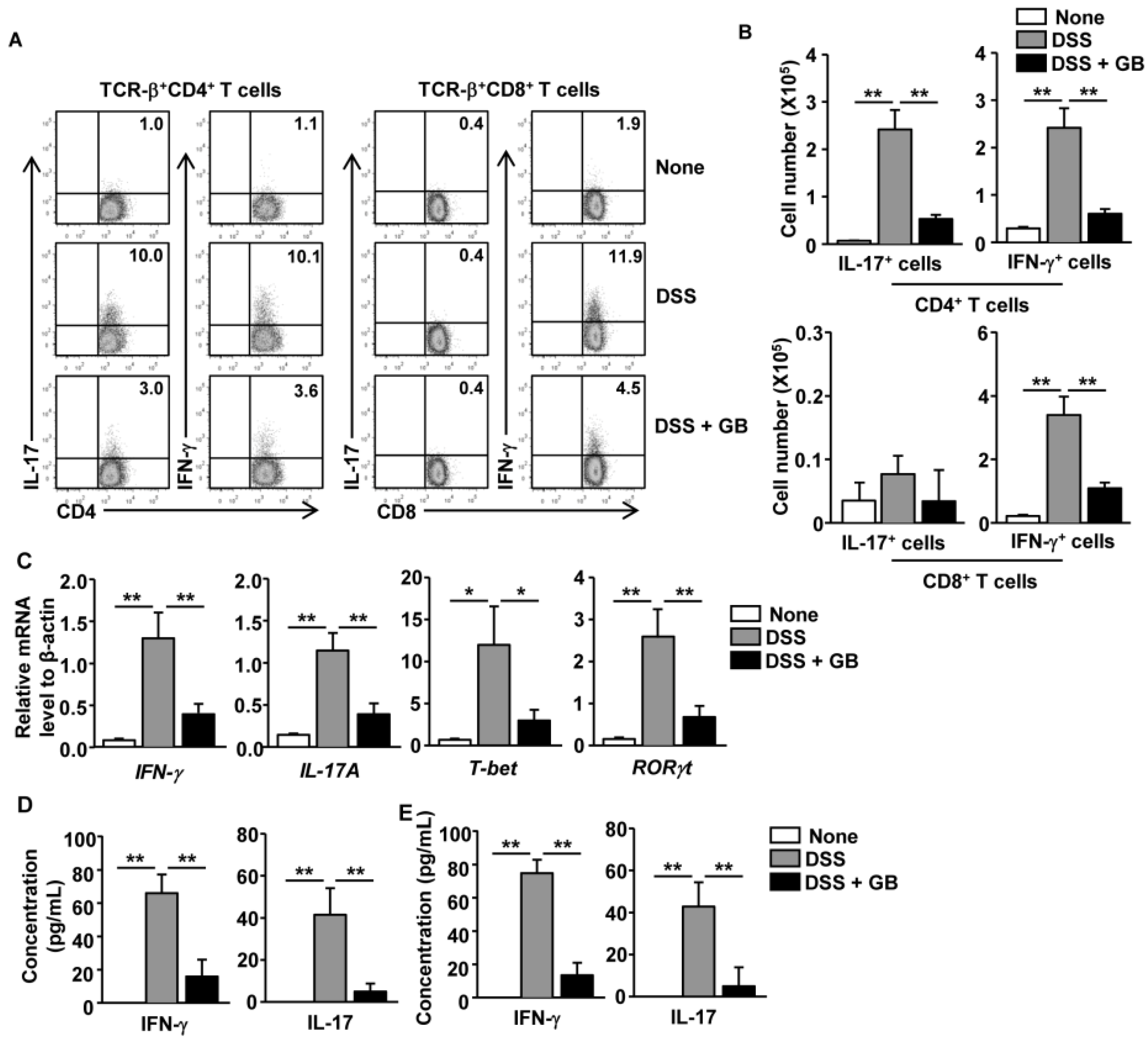
3.4. GB Inhibits DSS-Induced Th1 and Th17 Responses
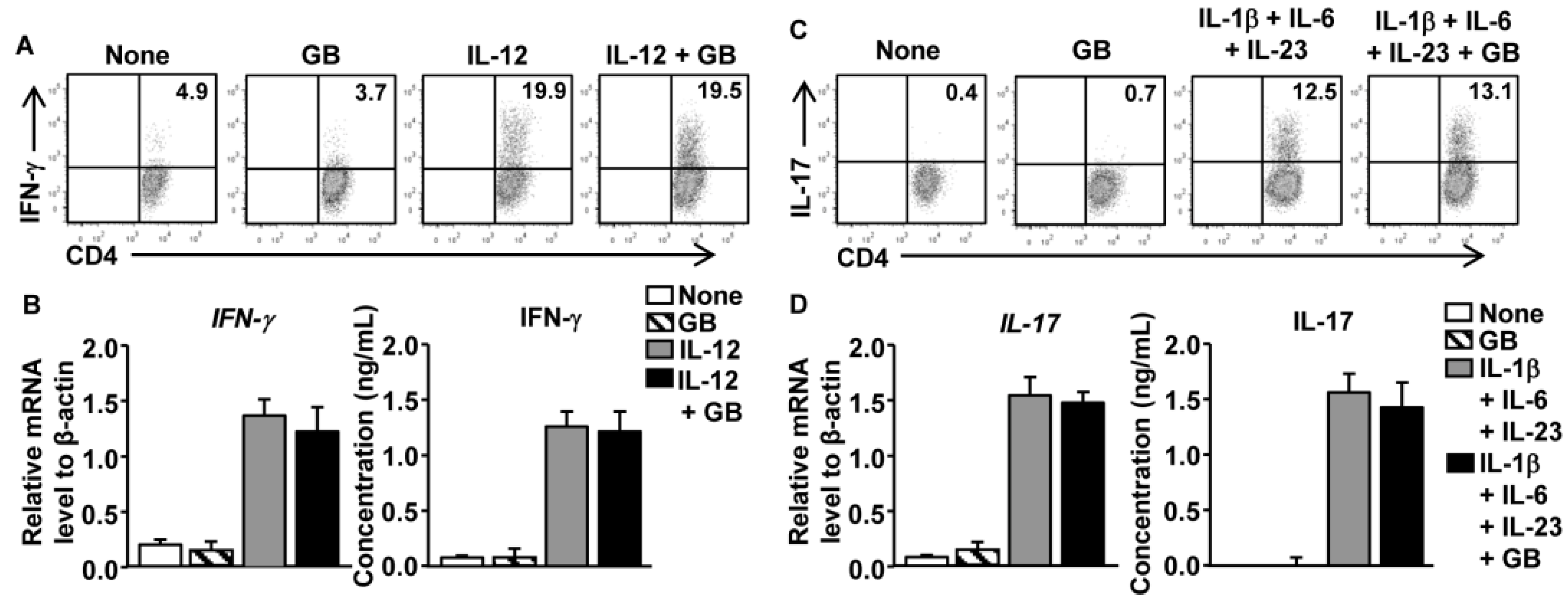
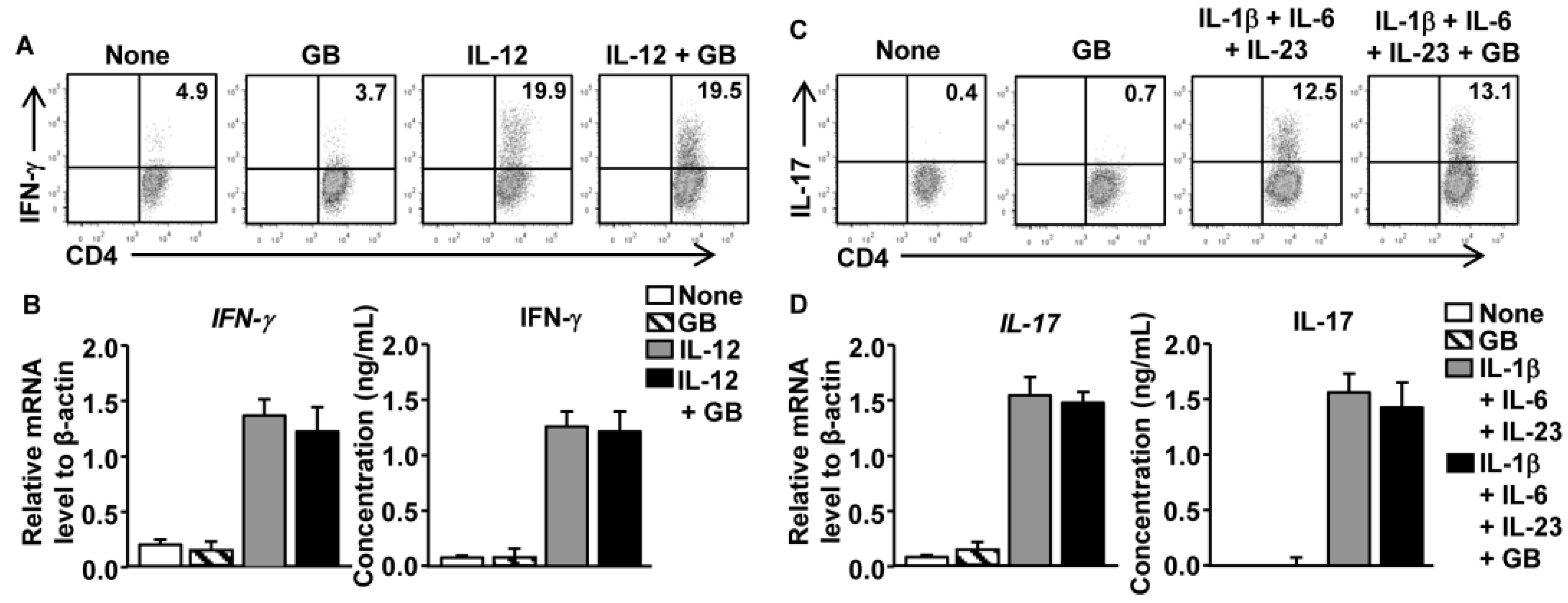
3.5. GB Does Not Directly Affect Th1 and Th17 Cell Differentiation
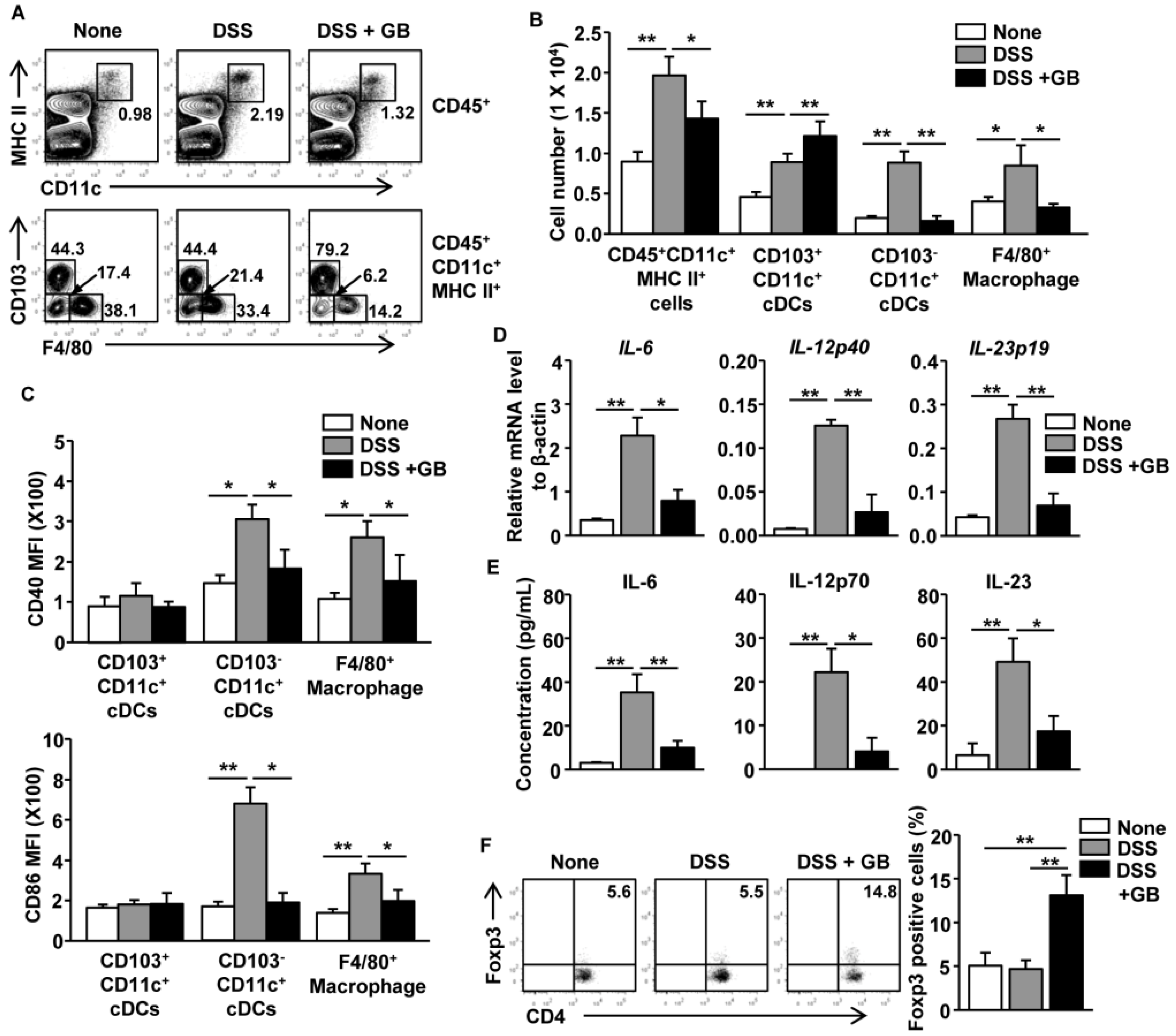
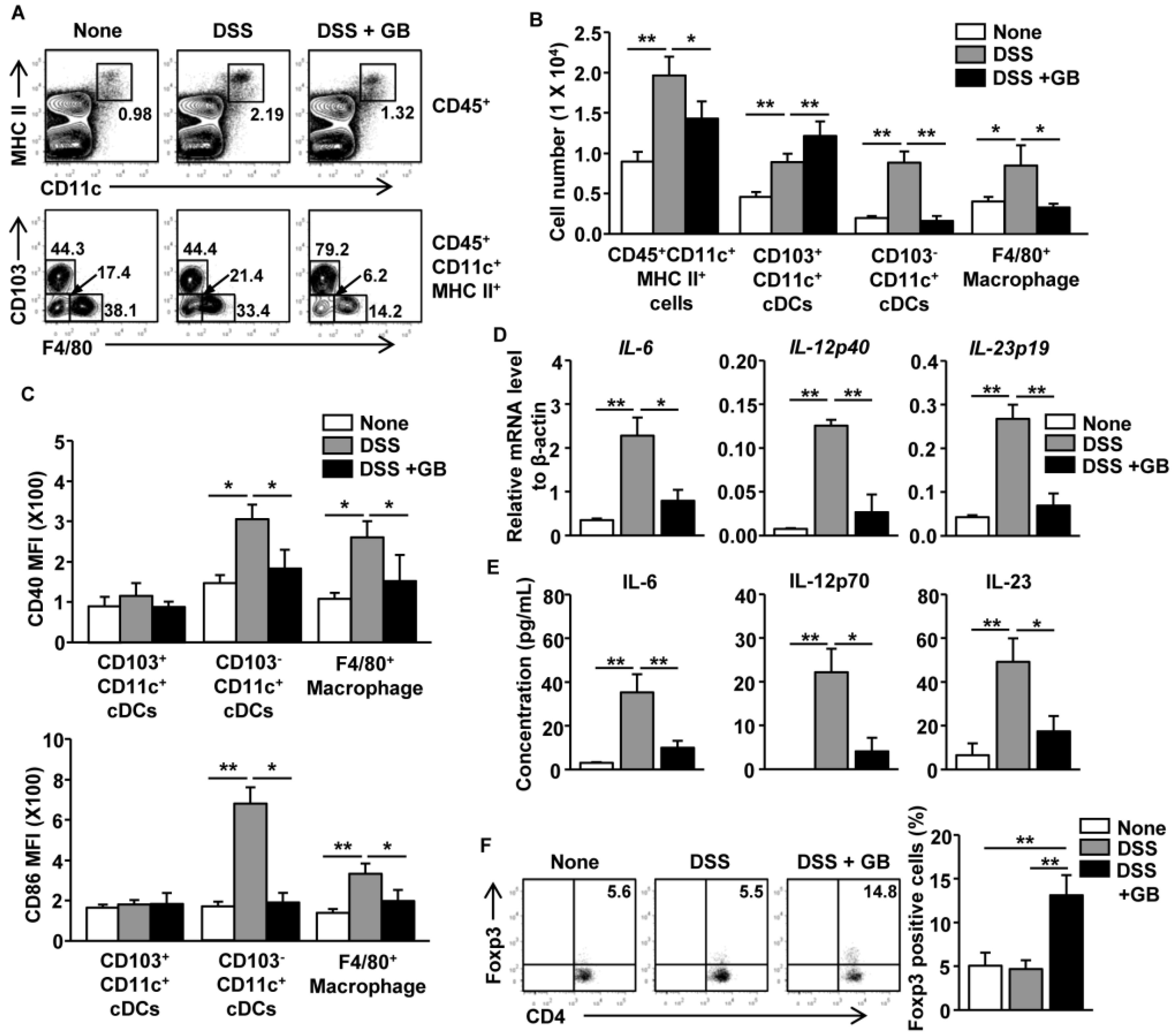
3.6. GB Inhibits DSS-Induced Activation of Intestinal DCs and Macrophages and Induces the Expansion of Regulatory T Cells
4. Discussion
5. Conclusions
Supplementary Files
Supplementary File 1Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| GB | ginseng berry extract |
| DSS | dextran sodium sulfate |
| UC | ulcerative colitis |
| GR | ginseng root extract |
| IBD | inflammatory bowel diseases |
| CD | Crohn’s disease |
| DC | dendritic cell |
| Th | T helper |
| Treg | regulatory T |
| LAL | limulus amebocyte lysate |
| Ca | calcium |
| Mg | magnesium |
| Zn | zinc |
| Fe | iron |
| K | potassium |
| Vit. | vitamin |
| DAI | disease activity index |
| PMA | phorbol 12-myristate 13-acetate |
References
- Blumenthal, M. Asian ginseng: Potential therapeutic uses. Adv. Nurse. Pract. 2001, 9, 26–28. [Google Scholar] [PubMed]
- Park, B.G.; Jung, H.J.; Cho, Y.W.; Lim, H.W.; Lim, C.J. Potentiation of antioxidative and anti-inflammatory properties of cultured wild ginseng root extract through probiotic fermentation. J. Pharm. Pharmacol. 2013, 65, 457–464. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Yu, X.; Yang, X.; Li, Y.; Yao, Y.; Lui, E.M.; Ren, G. Structural and anti-inflammatory characterization of a novel neutral polysaccharide from North American ginseng (Panax quinquefolius). Int. J. Biol. Macromol. 2015, 74, 12–17. [Google Scholar] [CrossRef] [PubMed]
- Yu, C.; Wen, X.D.; Zhang, Z.; Zhang, C.F.; Wu, X.H.; Martin, A.; Du, W.; He, T.C.; Wang, C.Z.; Yuan, C.S. American ginseng attenuates azoxymethane/dextran sodium sulfate-induced colon carcinogenesis in mice. J. Ginseng Res. 2015, 39, 14–21. [Google Scholar] [CrossRef] [PubMed]
- Park, E.Y.; Kim, H.J.; Kim, Y.K.; Park, S.U.; Choi, J.E.; Cha, J.Y.; Jun, H.S. Increase in Insulin Secretion Induced by Panax ginseng Berry Extracts Contributes to the Amelioration of Hyperglycemia in Streptozotocininduced Diabetic Mice. J. Ginseng Res. 2012, 36, 153–160. [Google Scholar] [CrossRef] [PubMed]
- Lu, J.M.; Yao, Q.; Chen, C. Ginseng compounds: An update on their molecular mechanisms and medical applications. Curr. Vasc. Pharmacol. 2009, 7, 293–302. [Google Scholar] [CrossRef] [PubMed]
- Cho, K.S.; Park, C.W.; Kim, C.K.; Jeon, H.Y.; Kim, W.G.; Lee, S.J.; Kim, Y.M.; Lee, J.Y.; Choi, Y.D. Effects of Korean ginseng berry extract (GB0710) on penile erection: evidence from in vitro and in vivo studies. Asian J. Androl. 2013, 15, 503–507. [Google Scholar] [CrossRef] [PubMed]
- Dey, L.; Xie, J.T.; Wang, A.; Wu, J.; Maleckar, S.A.; Yuan, C.S. Anti-hyperglycemic effects of ginseng: Comparison between root and berry. Phytomedicine 2003, 10, 600–605. [Google Scholar] [CrossRef] [PubMed]
- Danese, S.; Panes, J. Development of drugs to target interactions between leukocytes and endothelial cells and treatment algorithms for inflammatory bowel diseases. Gastroenterology 2014, 147, 981–989. [Google Scholar] [CrossRef] [PubMed]
- Steinbach, E.C.; Plevy, S.E. The role of macrophages and dendritic cells in the initiation of inflammation in IBD. Inflam. Bowel Dis. 2014, 20, 166–175. [Google Scholar] [CrossRef] [PubMed]
- Kamada, N.; Hisamatsu, T.; Okamoto, S.; Sato, T.; Matsuoka, K.; Arai, K.; Nakai, T.; Hasegawa, A.; Inoue, N.; Watanabe, N.; et al. Abnormally differentiated subsets of intestinal macrophage play a key role in Th1-dominant chronic colitis through excess production of IL-12 and IL-23 in response to bacteria. J. Immunol. 2005, 175, 6900–6908. [Google Scholar] [CrossRef] [PubMed]
- Uhlig, H.H.; Schwerd, T.; Koletzko, S.; Shah, N.; Kammermeier, J.; Elkadri, A.; Ouahed, J.; Wilson, D.C.; Travis, S.P.; Turner, D.; et al. The diagnostic approach to monogenic very early onset inflammatory bowel disease. Gastroenterology 2014, 147, 990–1007. [Google Scholar] [CrossRef] [PubMed]
- Abe, K.; Nguyen, K.P.; Fine, S.D.; Mo, J.H.; Shen, C.; Shenouda, S.; Corr, M.; Jung, S.; Lee, J.; Eckmann, L.; et al. Conventional dendritic cells regulate the outcome of colonic inflammation independently of T cells. Proc. Natl. Acad. Sci. USA 2007, 104, 17022–17027. [Google Scholar] [CrossRef] [PubMed]
- Hart, A.L.; Al-Hassi, H.O.; Rigby, R.J.; Bell, S.J.; Emmanuel, A.V.; Knight, S.C.; Kamm, M.A.; Stagg, A.J. Characteristics of intestinal dendritic cells in inflammatory bowel diseases. Gastroenterology 2005, 129, 50–65. [Google Scholar] [CrossRef] [PubMed]
- Ko, H.J.; Chang, S.Y. Regulation of intestinal immune system by dendritic cells. Immune Netw. 2015, 15, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Qualls, J.E.; Tuna, H.; Kaplan, A.M.; Cohen, D.A. Suppression of experimental colitis in mice by CD11c+ dendritic cells. Inflamm. Bowel Dis. 2009, 15, 236–247. [Google Scholar] [CrossRef] [PubMed]
- Mottet, C.; Uhlig, H.H.; Powrie, F. Cutting edge: Cure of colitis by CD4+CD25+ regulatory T cells. J. Immunol. 2003, 170, 3939–3943. [Google Scholar] [CrossRef] [PubMed]
- Uematsu, S.; Fujimoto, K.; Jang, M.H.; Yang, B.G.; Jung, Y.J.; Nishiyama, M.; Sato, S.; Tsujimura, T.; Yamamoto, M.; Yokota, Y.; et al. Regulation of humoral and cellular gut immunity by lamina propria dendritic cells expressing Toll-like receptor 5. Nat. Immunol. 2008, 9, 769–776. [Google Scholar] [CrossRef] [PubMed]
- Feng, T.; Qin, H.; Wang, L.; Benveniste, E.N.; Elson, C.O.; Cong, Y. Th17 cells induce colitis and promote Th1 cell responses through IL-17 induction of innate IL-12 and IL-23 production. J. Immunol. 2011, 186, 6313–6318. [Google Scholar] [CrossRef] [PubMed]
- Kim, C.K.; Cho, D.H.; Lee, K.S.; Lee, D.K.; Park, C.W.; Kim, W.G.; Lee, S.J.; Ha, K.S.; Goo Taeg, O.; Kwon, Y.G.; et al. Ginseng Berry Extract Prevents Atherogenesis via Anti-Inflammatory Action by Upregulating Phase II Gene Expression. Evid. Based Complement. Alternat. Med. 2012, 2012, 490301. [Google Scholar] [CrossRef] [PubMed]
- Al-Masri, M.R.; Al-Bachir, M. Microbial load, acidity, lipid oxidation and volatile basic nitrogen of irradiated fish and meat-bone meals. Bioresour. Technol. 2007, 98, 1163–1166. [Google Scholar] [CrossRef] [PubMed]
- Goldschmidt, R.J.; Wolf, W.R. Simultaneous determination of water-soluble vitamins in SRM 1849 Infant/Adult Nutritional Formula powder by liquid chromatography-isotope dilution mass spectrometry. Anal. Bioanal. Chem. 2010, 397, 471–481. [Google Scholar] [CrossRef] [PubMed]
- Bonvehi, J.S.; Coll, F.V.; Rius, I.A. Liquid chromatographic determination of tocopherols and tocotrienols in vegetable oils, formulated preparations, and biscuits. J. AOAC Int. 2000, 83, 627–634. [Google Scholar] [PubMed]
- Ghia, J.E.; Blennerhassett, P.; Kumar-Ondiveeran, H.; Verdu, E.F.; Collins, S.M. The vagus nerve: A tonic inhibitory influence associated with inflammatory bowel disease in a murine model. Gastroenterology 2006, 131, 1122–1130. [Google Scholar] [CrossRef] [PubMed]
- Takedatsu, H.; Michelsen, K.S.; Wei, B.; Landers, C.J.; Thomas, L.S.; Dhall, D.; Braun, J.; Targan, S.R. TL1A (TNFSF15) regulates the development of chronic colitis by modulating both T-helper 1 and T-helper 17 activation. Gastroenterology 2008, 135, 552–567. [Google Scholar] [CrossRef] [PubMed]
- Jin, J.O.; Kawai, T.; Cha, S.; Yu, Q. Interleukin-7 enhances the Th1 response to promote the development of Sjogren’s syndrome-like autoimmune exocrinopathy in mice. Arthritis Rheum. 2013, 65, 2132–2142. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Du, J.Y.; Yu, Q.; Jin, J.O. IL-7 Produced by Intestinal Epithelial Cells in Response to Citrobacter rodentium Infection Plays a Major Role in the Innate Immunity against This Pathogen. Infect. Immun. 2015. [Google Scholar] [CrossRef] [PubMed]
- Jakubzick, C.; Bogunovic, M.; Bonito, A.J.; Kuan, E.L.; Merad, M.; Randolph, G.J. Lymph-migrating, tissue-derived dendritic cells are minor constituents within steady-state lymph nodes. J. Exp. Med. 2008, 205, 2839–2850. [Google Scholar] [CrossRef] [PubMed]
- Ohl, L.; Mohaupt, M.; Czeloth, N.; Hintzen, G.; Kiafard, Z.; Zwirner, J.; Blankenstein, T.; Henning, G.; Forster, R. CCR7 governs skin dendritic cell migration under inflammatory and steady-state conditions. Immunity 2004, 21, 279–288. [Google Scholar] [CrossRef] [PubMed]
- Jin, J.O.; Han, X.; Yu, Q. Interleukin-6 induces the generation of IL-10-producing Tr1 cells and suppresses autoimmune tissue inflammation. J. Autoimmun. 2013, 40, 28–44. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, T.; Okamoto, S.; Hisamatsu, T.; Kamada, N.; Chinen, H.; Saito, R.; Kitazume, M.T.; Nakazawa, A.; Sugita, A.; Koganei, K.; et al. IL23 differentially regulates the Th1/Th17 balance in ulcerative colitis and Crohn’s disease. Gut 2008, 57, 1682–1689. [Google Scholar] [CrossRef] [PubMed]
- Ito, R.; Kita, M.; Shin-Ya, M.; Kishida, T.; Urano, A.; Takada, R.; Sakagami, J.; Imanishi, J.; Iwakura, Y.; Okanoue, T.; et al. Involvement of IL-17A in the pathogenesis of DSS-induced colitis in mice. Biochem. Biophys. Res. Commun. 2008, 377, 12–16. [Google Scholar] [CrossRef] [PubMed]
- Fournier, B.M.; Parkos, C.A. The role of neutrophils during intestinal inflammation. Mucosal Immunol. 2012, 5, 354–366. [Google Scholar] [CrossRef] [PubMed]
- Ye, P.; Rodriguez, F.H.; Kanaly, S.; Stocking, K.L.; Schurr, J.; Schwarzenberger, P.; Oliver, P.; Huang, W.; Zhang, P.; Zhang, J.; et al. Requirement of interleukin 17 receptor signaling for lung CXC chemokine and granulocyte colony-stimulating factor expression, neutrophil recruitment, and host defense. J. Exp. Med. 2001, 194, 519–527. [Google Scholar] [CrossRef] [PubMed]
- Atarashi, K.; Nishimura, J.; Shima, T.; Umesaki, Y.; Yamamoto, M.; Onoue, M.; Yagita, H.; Ishii, N.; Evans, R.; Honda, K.; et al. ATP drives lamina propria T(H)17 cell differentiation. Nature 2008, 455, 808–812. [Google Scholar] [CrossRef] [PubMed]
- Del Rio, M.L.; Bernhardt, G.; Rodriguez-Barbosa, J.I.; Forster, R. Development and functional specialization of CD103+ dendritic cells. Immunol. Rev. 2010, 234, 268–281. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.Z.; Zhang, B.; Song, W.X.; Wang, A.; Ni, M.; Luo, X.; Aung, H.H.; Xie, J.T.; Tong, R.; He, T.C.; et al. Steamed American ginseng berry: Ginsenoside analyses and anticancer activities. J. Agric. Food Chem. 2006, 54, 9936–9942. [Google Scholar] [CrossRef] [PubMed]
- Attele, A.S.; Zhou, Y.P.; Xie, J.T.; Wu, J.A.; Zhang, L.; Dey, L.; Pugh, W.; Rue, P.A.; Polonsky, K.S.; Yuan, C.S. Antidiabetic effects of Panax ginseng berry extract and the identification of an effective component. Diabetes 2002, 51, 1851–1858. [Google Scholar] [CrossRef] [PubMed]
- Graham, T.O.; Kandil, H.M. Nutritional factors in inflammatory bowel disease. Gastroenterol. Clin. North Am. 2002, 31, 203–218. [Google Scholar] [CrossRef]
- Lomer, M.C. Dietary and nutritional considerations for inflammatory bowel disease. Proc. Nutr. Soc. 2011, 70, 329–335. [Google Scholar] [CrossRef] [PubMed]
- Brown, K.; DeCoffe, D.; Molcan, E.; Gibson, D.L. Diet-induced dysbiosis of the intestinal microbiota and the effects on immunity and disease. Nutrients 2012, 4, 1095–1119. [Google Scholar] [CrossRef] [PubMed]
- Castro-Quezada, I.; Roman-Vinas, B.; Serra-Majem, L. The Mediterranean diet and nutritional adequacy: A review. Nutrients 2014, 6, 231–248. [Google Scholar] [CrossRef] [PubMed]
- Lagishetty, V.; Misharin, A.V.; Liu, N.Q.; Lisse, T.S.; Chun, R.F.; Ouyang, Y.; McLachlan, S.M.; Adams, J.S.; Hewison, M. Vitamin D deficiency in mice impairs colonic antibacterial activity and predisposes to colitis. Endocrinology 2010, 151, 2423–2432. [Google Scholar] [CrossRef] [PubMed]
- Benight, N.M.; Stoll, B.; Chacko, S.; da Silva, V.R.; Marini, J.C.; Gregory, J.F., III; Stabler, S.P.; Burrin, D.G. B-vitamin deficiency is protective against DSS-induced colitis in mice. Am. J. Physiol. Gastrointest. Liver Physiol. 2011, 301, G249–G259. [Google Scholar] [CrossRef] [PubMed]
- Cho, J.H.; Brant, S.R. Recent insights into the genetics of inflammatory bowel disease. Gastroenterology 2011, 140, 1704–1712. [Google Scholar] [CrossRef] [PubMed]
- Brown, J.B.; Cheresh, P.; Zhang, Z.; Ryu, H.; Managlia, E.; Barrett, T.A. P-selectin glycoprotein ligand-1 is needed for sequential recruitment of T-helper 1 (Th1) and local generation of Th17 T cells in dextran sodium sulfate (DSS) colitis. Inflamm. Bowel Dis. 2012, 18, 323–332. [Google Scholar] [CrossRef] [PubMed]
- Hanai, H.; Takeuchi, K.; Iida, T.; Kashiwagi, N.; Saniabadi, A.R.; Matsushita, I.; Sato, Y.; Kasuga, N.; Nakamura, T. Relationship between fecal calprotectin, intestinal inflammation, and peripheral blood neutrophils in patients with active ulcerative colitis. Dig. Dis. Sci. 2004, 49, 1438–1443. [Google Scholar] [CrossRef] [PubMed]
- Araki, Y.; Mukaisyo, K.; Sugihara, H.; Fujiyama, Y.; Hattori, T. Increased apoptosis and decreased proliferation of colonic epithelium in dextran sulfate sodium-induced colitis in mice. Oncol. Rep. 2010, 24, 869–874. [Google Scholar] [CrossRef] [PubMed]
- Hunter, M.M.; Wang, A.; Parhar, K.S.; Johnston, M.J.; Van Rooijen, N.; Beck, P.L.; McKay, D.M. In vitro-derived alternatively activated macrophages reduce colonic inflammation in mice. Gastroenterology 2010, 138, 1395–1405. [Google Scholar] [CrossRef] [PubMed]
- Arranz, A.; Doxaki, C.; Vergadi, E.; Martinez de la Torre, Y.; Vaporidi, K.; Lagoudaki, E.D.; Ieronymaki, E.; Androulidaki, A.; Venihaki, M.; Margioris, A.N.; et al. Akt1 and Akt2 protein kinases differentially contribute to macrophage polarization. Proc. Natl. Acad. Sci. USA 2012, 109, 9517–9522. [Google Scholar] [CrossRef] [PubMed]
- Annacker, O.; Coombes, J.L.; Malmstrom, V.; Uhlig, H.H.; Bourne, T.; Johansson-Lindbom, B.; Agace, W.W.; Parker, C.M.; Powrie, F. Essential role for CD103 in the T cell-mediated regulation of experimental colitis. J. Exp. Med. 2005, 202, 1051–1061. [Google Scholar] [CrossRef] [PubMed]
- Ruane, D.T.; Lavelle, E.C. The role of CD103(+) dendritic cells in the intestinal mucosal immune system. Front. Immunol. 2011, 2, 25. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Cho, S.Y.; Xiang, G.; Min, K.J.; Yu, Q.; Jin, J.O. Ginseng Berry Extract Promotes Maturation of Mouse Dendritic Cells. PLoS ONE 2015, 10, e0130926. [Google Scholar] [CrossRef] [PubMed]
- Lean, Q.Y.; Eri, R.D.; Fitton, J.H.; Patel, R.P.; Gueven, N. Fucoidan Extracts Ameliorate Acute Colitis. PLoS ONE 2015, 10, e0128453. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Oda, T.; Yu, Q.; Jin, J.O. Fucoidan from Macrocystis pyrifera has powerful immune-modulatory effects compared to three other fucoidans. Mar. Drugs 2015, 13, 1084–1104. [Google Scholar] [CrossRef] [PubMed]
- Jin, J.O.; Zhang, W.; Du, J.Y.; Wong, K.W.; Oda, T.; Yu, Q. Fucoidan can function as an adjuvant in vivo to enhance dendritic cell maturation and function and promote antigen-specific T cell immune responses. PLoS ONE 2014, 9, e99396. [Google Scholar] [CrossRef] [PubMed]
- Chassaing, B.; Aitken, J.D.; Malleshappa, M.; Vijay-Kumar, M. Dextran sulfate sodium (DSS)-induced colitis in mice. Curr. Protoc. Immunol 2014. [Google Scholar] [CrossRef]
- Melgar, S.; Karlsson, L.; Rehnstrom, E.; Karlsson, A.; Utkovic, H.; Jansson, L.; Michaelsson, E. Validation of murine dextran sulfate sodium-induced colitis using four therapeutic agents for human inflammatory bowel disease. Int. Immunopharmacol. 2008, 8, 836–844. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.S.; Lee, M.H.; Ju, A.S.; Rhee, K.J. Th17 responses are not induced in dextran sodium sulfate model of acute colitis. Immune Netw. 2011, 11, 416–419. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| K | Ca | Fe | Mg | Zn | Folate | PA | Niacin | Vit. | |||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| A | B1 | B2 | B6 | E | |||||||||
| GB | 5860 | 820 | 59.3 | 354 | 178 | 345 | 5.8 | 5.8 | 213 | 12.3 | 8.4 | 10.5 | 23.6 |
| GR | 1460 | 340 | 9.8 | 160 | 2 | n.d. | 0.7 | 1 | n.d. | 0.2 | n.d. | n.d. | n.d. |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, W.; Xu, L.; Cho, S.-Y.; Min, K.-J.; Oda, T.; Zhang, L.; Yu, Q.; Jin, J.-O. Ginseng Berry Extract Attenuates Dextran Sodium Sulfate-Induced Acute and Chronic Colitis. Nutrients 2016, 8, 199. https://doi.org/10.3390/nu8040199
Zhang W, Xu L, Cho S-Y, Min K-J, Oda T, Zhang L, Yu Q, Jin J-O. Ginseng Berry Extract Attenuates Dextran Sodium Sulfate-Induced Acute and Chronic Colitis. Nutrients. 2016; 8(4):199. https://doi.org/10.3390/nu8040199
Chicago/Turabian StyleZhang, Wei, Li Xu, Si-Young Cho, Kyung-Jin Min, Tatsuya Oda, LiJun Zhang, Qing Yu, and Jun-O Jin. 2016. "Ginseng Berry Extract Attenuates Dextran Sodium Sulfate-Induced Acute and Chronic Colitis" Nutrients 8, no. 4: 199. https://doi.org/10.3390/nu8040199