Protective Activity of Total Polyphenols from Genista quadriflora Munby and Teucrium polium geyrii Maire in Acetaminophen-Induced Hepatotoxicity in Rats

 , ,
, ,
Abstract
:
1. Introduction
2. Materials and Methods
2.1. General Experimental Procedures
2.2. Plant Material Collect and Extracts Preparation
2.3. Determination of Total Polyphenol Content
2.4. TLC-Fingerprint Analysis
2.5. Animals
2.6. Biochemical Analysis
2.7. Histological Analysis
2.8. Oxidative Stress Markers
2.9. Mitochondria Isolation
2.10. Mitochondrial Respiration
2.11. Mitochondrial Respiratory Complexes and Citrate Synthase Activities
2.12. Western Blot Analysis
2.13. Analysis of mRNA Levels by Real-Time PCR
2.14. Protein Levels
2.15. Statistical Analysis
3. Results
3.1. Total Polyphenols Content (TPC) and TLC -Fingerprint of Gq and Tp Extracts
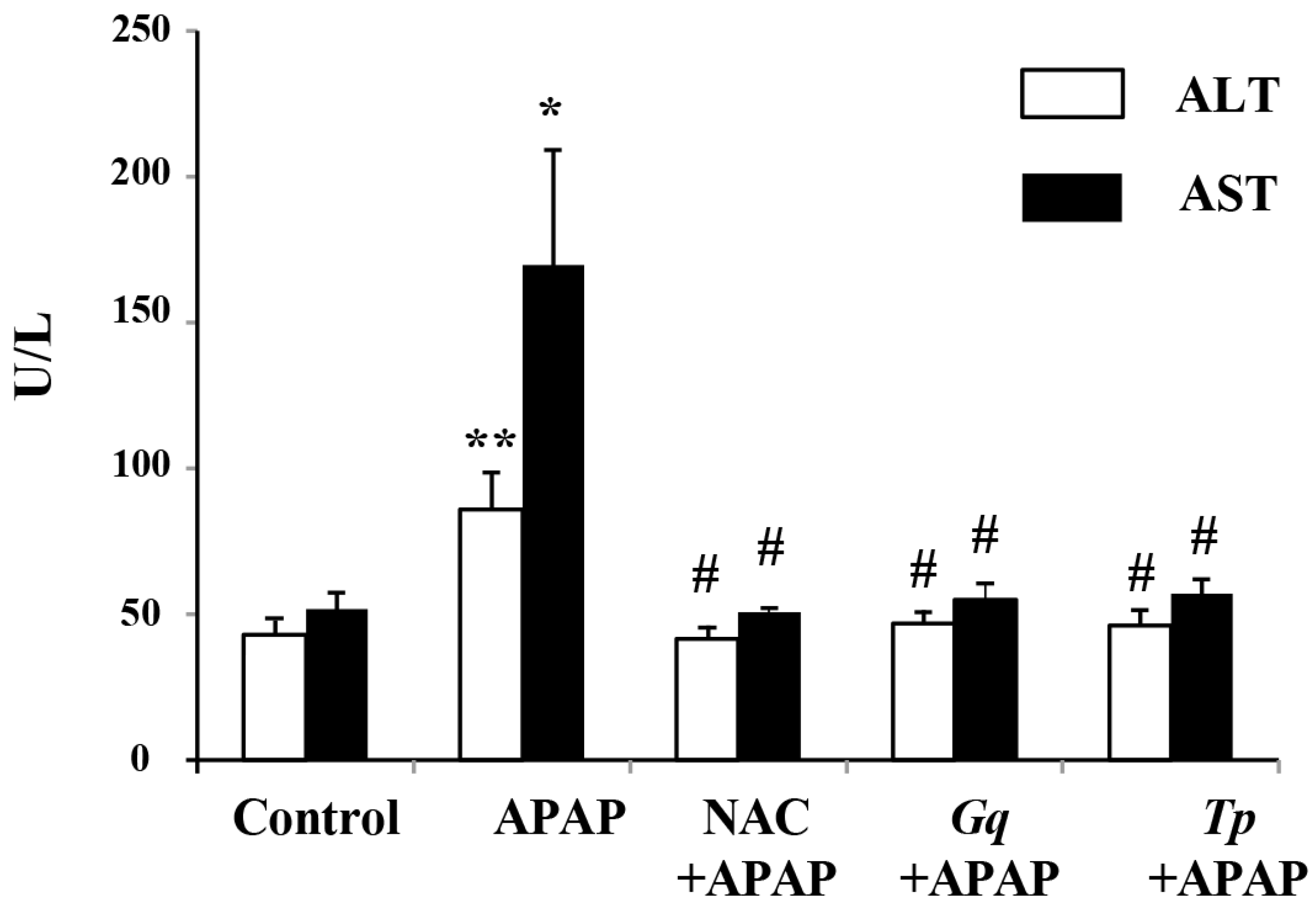
3.2. Influence of Gq and Tp Extracts on Blood Transaminases Levels
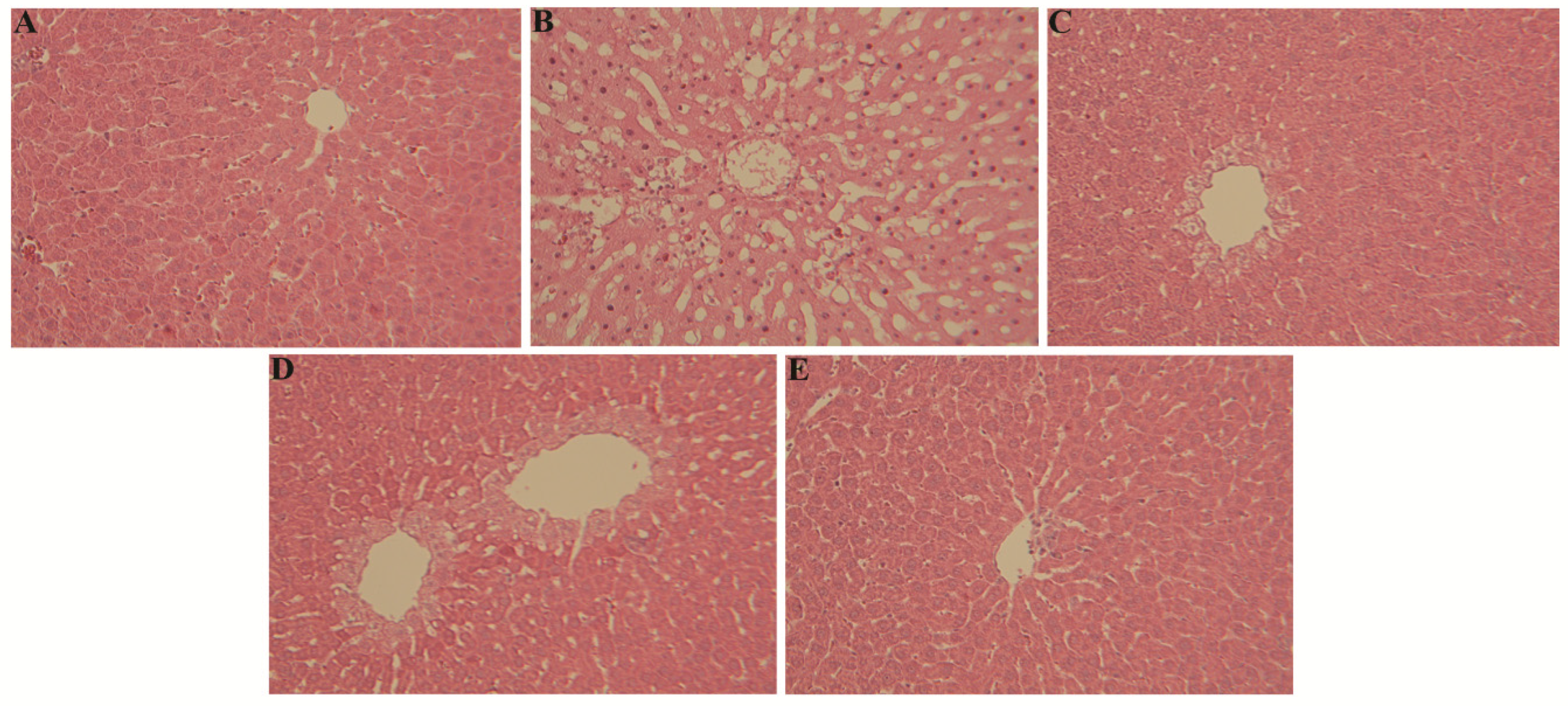
3.3. Influence of Gq and Tp Extracts on Liver Histology
3.4. Effect of Gq and Tp Extracts on Liver Oxidative Stress Markers
3.5. Effect of Gq and Tp Extracts on Liver Mitochondrial Activity
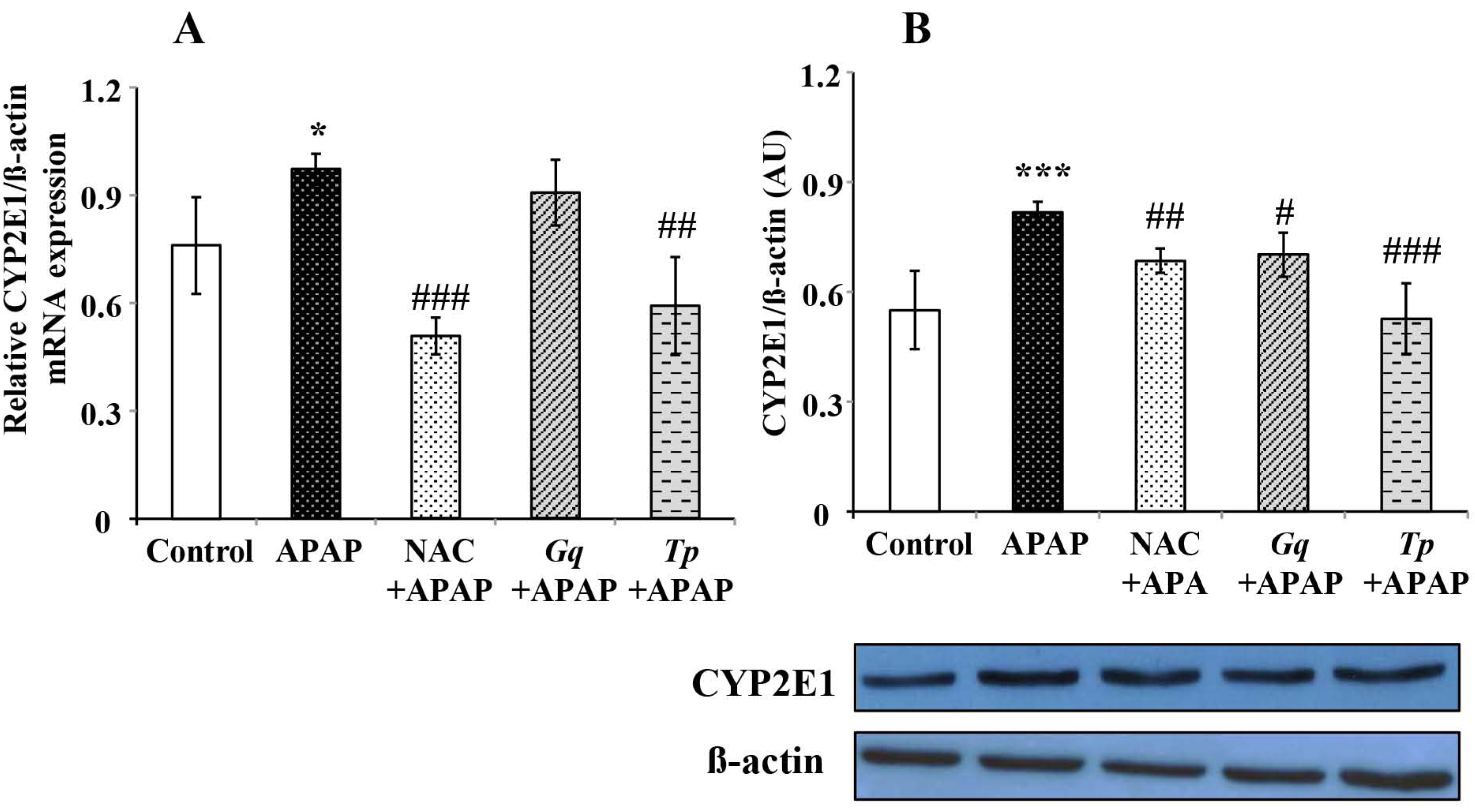
3.6. Effect of Gq and Tp Extracts on Liver CYP2E1 Protein and mRNA Levels
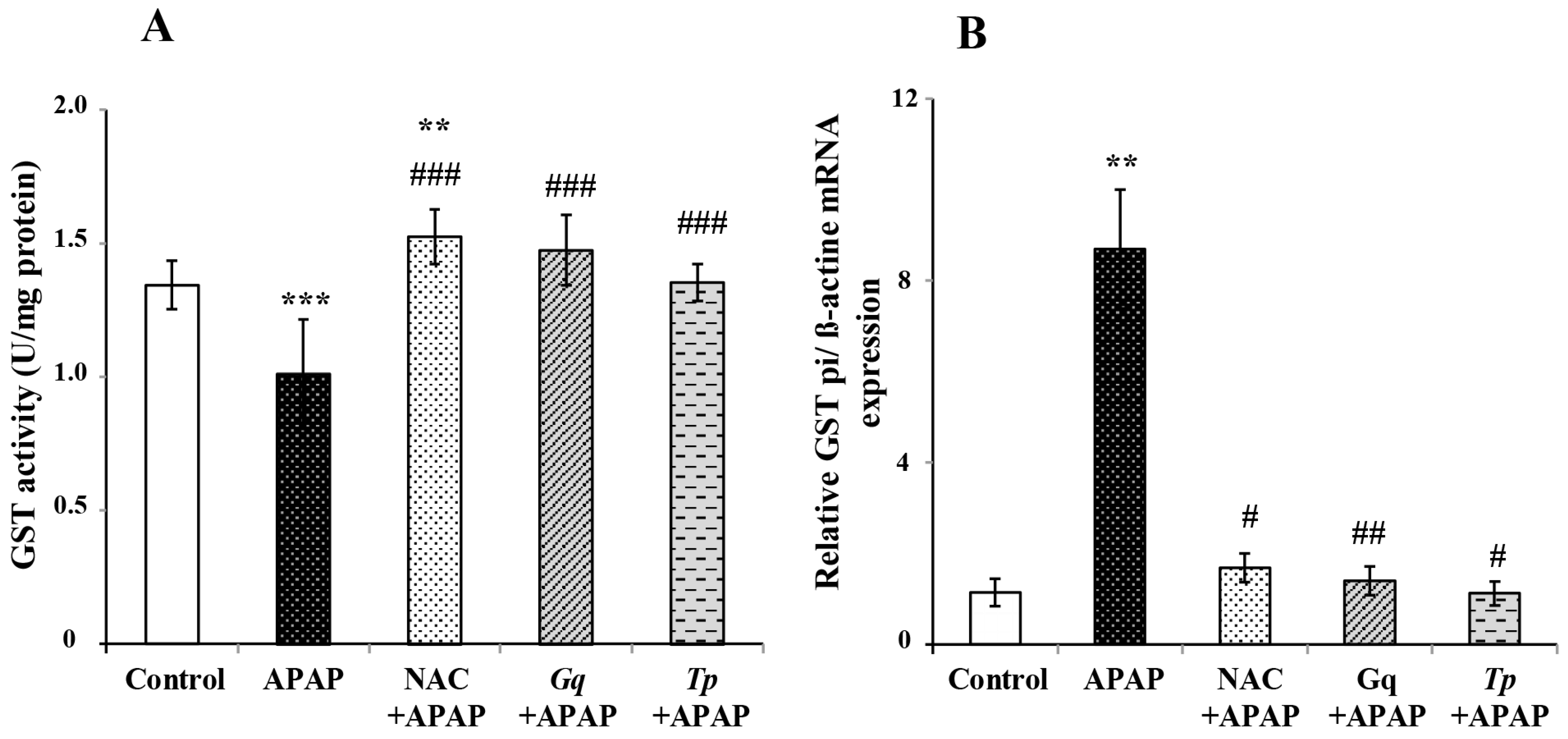
3.7. Effect of Gq and Tp Extracts on Liver GST activity and GSTpi mRNA Levels
3.8. Effect of Gq and Tp Extracts on Liver TNF-α Expression Levels
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Prescott, L.F. Hepatotoxicity of mild analgesics. Br. J. Clin. Pharmacol. 1980, 10 (Suppl. 2), 373S–379S. [Google Scholar] [CrossRef] [PubMed]
- Dahlin, D.C.; Miwa, G.T.; Lu, A.Y.; Nelson, S.D. N-acetyl-p-benzoquinone imine: A cytochrome P-450-mediated oxidation product of acetaminophen. Proc. Natl. Acad. Sci. USA 1984, 81, 1327–1331. [Google Scholar] [CrossRef] [PubMed]
- Meyers, L.L.; Beierschmitt, W.P.; Khairallah, E.A.; Cohen, S.D. Acetaminophen-induced inhibition of hepatic mitochondrial respiration in mice. Toxicol. Appl. Pharmacol. 1988, 93, 378–387. [Google Scholar] [CrossRef]
- Katyare, S.S.; Satav, J.G. Impaired mitochondrial oxidative energy metabolism following paracetamol-induced hepatotoxicity in the rat. Br. J. Pharmacol. 1989, 96, 51–58. [Google Scholar] [CrossRef] [PubMed]
- Jaeschke, H.; Knight, T.R.; Bajt, M.L. The role of oxidant stress and reactive nitrogen species in acetaminophen hepatotoxicity. Toxicol. Lett. 2003, 144, 279–288. [Google Scholar] [CrossRef]
- Reid, A.B.; Kurten, R.C.; McCullough, S.S.; Brock, R.W.; Hinson, J.A. Mechanisms of acetaminophen-induced hepatotoxicity: Role of oxidative stress and mitochondrial permeability transition in freshly isolated mouse hepatocytes. J. Pharmacol. Exp. Ther. 2005, 312, 509–516. [Google Scholar] [CrossRef] [PubMed]
- Saito, C.; Zwingmann, C.; Jaeschke, H. Novel mechanisms of protection against acetaminophen hepatotoxicity in mice by glutathione and N-acetylcysteine. Hepatology 2010, 51, 246–254. [Google Scholar] [CrossRef] [PubMed]
- Vendemiale, G.; Grattagliano, I.; Altomare, E.; Turturro, N.; Guerrieri, F. Effect of acetaminophen administration on hepatic glutathione compartmentation and mitochondrial energy metabolism in the rat. Biochem. Pharmacol. 1996, 52, 1147–1154. [Google Scholar] [CrossRef]
- Kon, K.; Kim, J.S.; Uchiyama, A.; Jaeschke, H.; Lemasters, J.J. Lysosomal iron mobilization and induction of the mitochondrial permeability transition in acetaminophen-induced toxicity to mouse hepatocytes. Toxicol. Sci. 2010, 117, 101–108. [Google Scholar] [CrossRef] [PubMed]
- Smilkstein, M.J.; Knapp, G.L.; Kulig, K.W.; Rumack, B.H. Efficacy of oral N-acetylcysteine in the treatment of acetaminophen overdose: Analysis of the national multicenter study (1976 to 1985). N. Engl. J. Med. 1988, 319, 1557–1562. [Google Scholar] [CrossRef] [PubMed]
- Grimble, G.K. Adverse gastrointestinal effects of arginine and related amino acids. J. Nutr. 2007, 137, 1693S–1701S. [Google Scholar] [PubMed]
- Luper, S. A review of plants used in the treatment of liver disease: Part 1. Altern. Med. Rev. 1998, 3, 410–421. [Google Scholar] [PubMed]
- Wang, A.Y.; Lian, L.H.; Jiang, Y.Z.; Wu, Y.L.; Nan, J.X. Gentiana manshurica Kitagawa prevents acetaminophen-induced acute hepatic injury in mice via inhibiting JNK/ERK MAPK pathway. World J. Gastroenterol. 2010, 16, 384–391. [Google Scholar] [CrossRef] [PubMed]
- Yousef, M.I.; Omar, S.A.; El-Guendi, M.I.; Abdelmegid, L.A. Potential protective effects of quercetin and curcumin on paracetamol-induced histological changes, oxidative stress, impaired liver and kidney functions and haematotoxicity in rat. Food Chem. Toxicol. 2010, 48, 3246–3261. [Google Scholar] [CrossRef] [PubMed]
- Rice-Evans, C.A.; Miller, N.J.; Bolwell, P.G.; Bramley, P.M.; Pridham, J.B. The relative antioxidant activities of plant-derived polyphenolic flavonoids. Free Radic. Res. 1995, 22, 375–383. [Google Scholar] [CrossRef] [PubMed]
- Yazdanparast, R.; Amin Ardestani, A. Suppressive effect of ethyl acetate extract of Teucrium polium on cellular oxidative damages and apoptosis induced by 2-deoxy-d-ribose: Role of de novo synthesis of glutathione. Food Chem. 2009, 114, 1222–1230. [Google Scholar] [CrossRef]
- Quezel, P.; Santa, S. Nouvelle Flore de l’Algérie et des Régions Désertiques Méridionales; Tome, I., Ed.; Centre National de la Recherche Scientifique (CNRS): Paris, France, 1962; pp. 470–471. [Google Scholar]
- Maire, R. Flore de l’Afrique du Nord (Maroc, Algerie, Tunisie, Tripolitaine, Cyrenaique et Sahara); Lechevalier: Paris, France, 1987. [Google Scholar]
- Rainova, L.; Nakov, N.; Bogdanova, S.; Minkov, E.; Staneva-Stoytcheva, D. Ulceroprotective activity of the flavonoids of Genista rumelica Vel. Phytother. Res. 1988, 2, 137–139. [Google Scholar] [CrossRef]
- Rauter, A.P.; Martins, A.; Lopes, R.; Ferreira, J.; Serralheiro, L.M.; Araújo, M.E.; Borges, C.; Justino, J.; Silva, F.V.; Goulart, M.; et al. Bioactivity studies and chemical profile of the antidiabetic plant Genista tenera. J. Ethnopharmacol. 2009, 122, 384–393. [Google Scholar] [CrossRef] [PubMed]
- Garritano, S.; Pinto, B.; Giachi, I.; Pistelli, L.; Reali, D. Assessment of estrogenic activity of flavonoids from Mediterranean plants using an in vitro short-term test. Phytomedicine 2005, 12, 143–147. [Google Scholar] [CrossRef] [PubMed]
- Rigano, D.; Cardile, V.; Formisano, C.; Maldini, M.T.; Piacente, S.; Bevelacqua, J.; Russo, A. Genista sessilifolia DC. and Genista tinctoria L. inhibit UV light and nitric oxide-induced DNA damage and human melanoma cell growth. Chem. Biol. Interact. 2009, 180, 211–219. [Google Scholar] [CrossRef] [PubMed]
- Orhan, E.I.; Tosun, F.; Tamer, U.; Duran, A.; Alan, B.; Kök, A.F. Quantification of genistein and daidzein in two endemic Genista species and their antioxidant activity. J. Serb. Chem. Soc. 2011, 76, 35–42. [Google Scholar] [CrossRef]
- Martins, A.; Wink, M.; Tei, A.; Brum-Bousquet, M.; Tillequin, F.; Rauter, A.P. A phytochemical study of the quinolizidine alkaloids from Genista tenera by gas chromatography-mass spectrometry. Phytochem. Anal. 2005, 16, 264–266. [Google Scholar] [CrossRef] [PubMed]
- Ozenda, P. Flore du Sahara, 2nd ed.; Centre National de la Recherche Scientifique (CNRS): Paris, France, 1983. [Google Scholar]
- Sadraei, H.; Hajhashemi, V.; Ghannadi, A.; Mohseni, M. Antispasmodic effect of aerial part of Teucrium polium. essential oil on rat isolated ileum in vitro. Med. J. Islamic. Rep. Iran. 2001, 14, 355–358. [Google Scholar]
- Stankovic, M.S.; Niciforovic, N.; Mihailovic, V.; Topuzovic, M.; Solujic, S. Antioxidant activity, total phenolic content and flavonoid concentrations of different plant parts of Teucrium polium. Subsp. polium. Acta Soc. Bot. Pol. 2012, 81, 117–122. [Google Scholar] [CrossRef]
- Bahramikia, S.; Yazdanparast, R. Phytochemistry and medicinal properties of Teucrium polium L. (Lamiaceae). Phytother. Res. 2012, 26, 1581–1593. [Google Scholar] [CrossRef] [PubMed]
- Parsaee, H.; Shafiee-Nick, R. Anti-Spasmodic and Anti-Nociceptive Effects of Teucrium polium Aqueous Extract. Iran. Biomed. J. 2006, 10, 145–149. [Google Scholar]
- Allahtavakoli, M.; Moradi, R.; Shamsi, S.; Afsharmanesh, K. Effect of hydro-alcoholic extract of Teucrium polium on castor oil-induced diarrhea in male rat. Zahedan. J. Res. Med. Sci. 2013, 15, 54–57. [Google Scholar]
- Mehrabani, D.; Rezaee, A.; Azarpira, N.; Fattahi, M.R.; Amini, M.; Tanideh, N.; Panjehshahin, M.R.; Saberi-Firouzi, M. The healing effects of Teucrium polium in the repair of indomethacin-induced gastric ulcer in rats. Saudi. Med. J. 2009, 30, 494–499. [Google Scholar] [PubMed]
- Gülçin, I.; Uğuz, M.T.; Oktay, M.; Beydemir, S.; Küfrevioğlu, Ö.I. Antioxidant and antimicrobial activities of Teucrium polium L. J. Food. Technol. 2003, 1, 9–16. [Google Scholar]
- Movahedi, A.; Basir, R.; Rahmat, A.; Charaffedine, M.; Othman, F. Remarkable anticancer activity of Teucrium polium on hepatocellular carcinogenic rats. Evid. Based Complement. Alternat. Med. 2014, 2014, 726724. [Google Scholar] [CrossRef] [PubMed]
- Forouzandeh, H.; Azemi, M.E.; Rashidi, I.; Goudarzi, M.; Kalantari, H. Study of the protective effect of Teucrium polium L. extract on acetaminophen-induced hepatotoxicity in mice. Iran. J. Pharm. Res. 2013, 12, 123–129. [Google Scholar] [PubMed]
- Belmekki, N.; Bendimerad, N.; Bekhechi, C.; Fernandez, X. Chemical analysis and antimicrobial activity of Teucrium polium L. essential oil from Western Algeria. J. Med. Plants Res. 2013, 7, 897–902. [Google Scholar]
- Panovska, K.T.; Kulevanova, S.; Stefova, M. In vitro antioxidant activity of some Teucrium species (Lamiaceae). Acta Pharm. 2005, 55, 207–214. [Google Scholar]
- Sarr, M.; Ngom, S.; Kane, M.O.; Wele, A.; Diop, D.; Sarr, B.; Gueye, L.; Andriantsitohaina, R.; Diallo, A.S. In vitro vasorelaxation mechanisms of bioactive compounds extracted from Hibiscus sabdariffa on rat thoracic aorta. Nutr. Metab. 2009, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singleton, V.L.; Rossi, J.A. Colorimetry of total phenolics with phosphomolybdic-phosphotungstic acid reagents. Am. J. Enol. Vitic. 1965, 16, 144–158. [Google Scholar]
- Wagner, H. Plant Drug Analysis: A Thin Layer Chromatography Atlas; Bladt, S., Ed.; Springer: Berlin/Heidelberg, Germany, 1996. [Google Scholar]
- Markham, K.R. Techniques of Flavonoid Identification; Treherne, J.E., Rubery, P.H., Eds.; Academic Press: New York, NY, USA, 1982; pp. 1–113. [Google Scholar]
- Naglaa, A.B.; Saad, H.E.; Mohammed, M.M.; Ahmed, M.S.H. Protective effect of curcumin versus N-acetylcystein ona induced hepatotoxicity in adult Albino rats. J. Cytol. Histol. 2015. [Google Scholar] [CrossRef]
- Nithianantham, K.; Shyamala, M.; Chen, Y.; Latha, L.Y.; Jothy, S.L.; Sasidharan, S. Hepatoprotective potential of Clitoria ternatea leaf extract against paracetamol induced damage in mice. Molecules 2011, 16, 10134–10145. [Google Scholar] [CrossRef] [PubMed]
- El-Shenawya, S.M.; Marzouk, M.S.; El Dib, R.A.; Elyazed, H.A.; Shaffie, N.S.; Moharram, F.A. Polyphenols and biological activities of feijoa sellowiana leaves and twigs. Rev. Latinoamer. Quim. 2008, 36, 103–120. [Google Scholar]
- Sunderman, F.W.; Marzouk, A.; Hopfer, S.M.; Zaharia, O.; Reid, M.C. Increased lipid peroxidation in tissues of nickel chloride-treated rats. Ann. Clin. Lab. Sci. 1985, 15, 229–236. [Google Scholar] [PubMed]
- Griffith, O.W. Determination of glutathione and glutathione disulfide using glutathione reductase and 2-vinylpyridine. Anal. Biochem. 1980, 106, 207–212. [Google Scholar] [CrossRef]
- Marklund, S.; Marklund, G. Involvement of the superoxide anion radical in the autoxidation of pyrogallol and a convenient assay for superoxide dismutase. Eur. J. Biochem. 1974, 47, 469–474. [Google Scholar] [CrossRef] [PubMed]
- Flohé, L.; Günzler, W.A. Assays of glutathione peroxidase. Methods Enzymol. 1984, 105, 114–121. [Google Scholar] [PubMed]
- Carlberg, I.; Mannervik, B. Glutathione reductase. Methods Enzymol. 1985, 113, 484–490. [Google Scholar] [PubMed]
- Habig, W.H.; Pabst, M.J.; Jakoby, W.B. Glutathione S-transferases the first enzymatic step in mercapturic acid formation. J. Biol. Chem. 1974, 249, 7130–7139. [Google Scholar] [PubMed]
- Frezza, C.; Cipolat, S.; Scorrano, L. Organelle isolation: Functional mitochondria from mouse liver, muscle and cultured fibroblasts. Nat. Protoc. 2007, 2, 287–295. [Google Scholar] [CrossRef] [PubMed]
- Estabrook, R.W. Mitochondrial respiratory control and the polarographic measurement of ADP: O ratios. Methods Enzymol. 1967, 10, 41–47. [Google Scholar]
- Janssen, A.J.M.; Trijbels, F.J.M.; Sengers, R.C.A.; Smeitink, J.A.M.; Van den Heuvel, L.P.; Wintjes, L.T.; Stoltenborg-Hogenkamp, B.J.M.; Rodenburg, R.J.T. Spectrophotometric assay for complex I of the respiratory chain in tissue samples and cultured fibroblasts. Clin. Chem. 2007, 53, 729–734. [Google Scholar] [CrossRef] [PubMed]
- Rustin, P.; Chretien, D.; Bourgeron, T.; Gérard, B.; Rötig, A.; Saudubray, J.M.; Munnich, A. Biochemical and molecular investigations in respiratory chain deficiencies. Clin. Chim. Acta 1994, 228, 35–51. [Google Scholar] [CrossRef]
- Wharton, D.C.; Tzagoloff, A. Cytochrome oxidase from beef heart mitochondria. Methods Enzymol. 1967, 10, 245–250. [Google Scholar]
- Srere, P. Citrate synthase: (EC 4.1.3.7. Citrate oxaloacetate-lyase (CoA-acetylating)). Methods Enzymol. 1969, 13, 3–11. [Google Scholar]
- Andrè, C.M.; Oufir, M.; Hoffmann, L.; Hausman, J.F.; Rogez, H.; Larondelle, Y.; Evers, D. Influence of environment and genotype on polyphenol compounds and in vitro antioxidant capacity of native Andean potatoes (Solanum tuberosum L.). J. Food Compos. Anal. 2009, 22, 517–524. [Google Scholar] [CrossRef]
- Shtukmaster, S.; Ljubuncic, P.; Bomzon, A. The Effect of an aqueous extract of Teucrium polium on glutathione homeostasis in vitro: A possible mechanism of its hepatoprotectant action. Adv. Pharmacol. Sci. 2010, 2010, 938324. [Google Scholar] [CrossRef] [PubMed]
- Oz, H.S.; McClain, C.J.; Nagasawa, H.T.; Ray, M.B.; De Villiers, W.J.; Chen, T.S. Diverse antioxidants protect against acetaminophen hepatotoxicity. J. Biochem. Mol. Toxicol. 2004, 18, 361–368. [Google Scholar] [CrossRef] [PubMed]
- Soliman, M.M.; Abdo Nassan, M.; Ismail, T.A. Immunohistochemical and molecular study on the protective effect of curcumin against hepatic toxicity induced by paracetamol in Wistar rats. BMC Complement. Altern. Med. 2014, 29. [Google Scholar] [CrossRef] [PubMed]
- Xiao, J.; Sun, G.B.; Sun, B.; Wu, Y.; He, L.; Wang, X.; Chen, R.C.; Cao, L.; Ren, X.Y.; Sun, X.B. Kaempferol protects against doxorubicin-induced cardiotoxicity in vivo and in vitro. Toxicology 2012, 292, 53–62. [Google Scholar] [CrossRef] [PubMed]
- Dixon, M.F.; Fulker, M.J.; Walker, B.E.; Kelleher, J.; Losowsky, M.S. Serum transaminase levels after experimental paracetamol-induced hepatic necrosis. Gut. 1975, 16, 800–807. [Google Scholar] [CrossRef] [PubMed]
- Bajt, M.L.; Knight, T.R.; Lemasters, J.J.; Jaeschke, H. Acetaminophen-induced oxidant stress and cell injury in cultured mouse hepatocytes: protection by N-acetyl cysteine. Toxicol. Sci. 2004, 80, 343–349. [Google Scholar] [CrossRef] [PubMed]
- Albano, E.; Poli, G.; Chiarpotto, E.; Biasi, F.; Dianzani, M.U. Paracetamol-stimulated lipid peroxidation in isolated rat and mouse hepatocytes. Chem. Biol. Interact. 1983, 47, 249–263. [Google Scholar] [CrossRef]
- Knight, T.R.; Fariss, M.W.; Farhood, A.; Jaeschke, H. Role of lipid peroxidation as a mechanism of liver injury after acetaminophen overdose in mice. Toxicol. Sci. 2003, 76, 229–236. [Google Scholar] [CrossRef] [PubMed]
- Panovska, T.K.; Kulevanova, S.; Gjorgoski, I.; Bogdanova, M.; Petrushevska, G. Hepatoprotective effect of the ethyl acetate extract of Teucrium polium L. against carbontetrachloride-induced hepatic injury in rats. Acta Pharm. 2007, 57, 241–248. [Google Scholar] [CrossRef] [PubMed]
- Amini, R.; Nosrati, N.; Yazdanparast, R.; Molaei, M. Teucrium polium in prevention of steatohepatitis in rats. Liver Int. 2009, 29, 1216–1221. [Google Scholar] [CrossRef] [PubMed]
- Ramsay, R.R.; Rashed, M.S.; Nelson, S.D. In vitro effects of acetaminophen metabolites and analogs on the respiration of mouse liver mitochondria. Arch. Biochem. Biophys. 1989, 273, 449–457. [Google Scholar] [CrossRef]
- Burcham, P.C.; Harman, A.W. Acetaminophen toxicity results in site-specific mitochondrial damage in isolated mouse hepatocytes. J. Biol. Chem. 1991, 266, 5049–5054. [Google Scholar] [PubMed]
- Abdelmegeed, M.A.; Jang, S.; Banerjee, A.; Hardwick, J.P.; Song, B.J. Robust protein nitration contributes to acetaminophen-induced mitochondrial dysfunction and acute liver injury. Free Radic. Biol. Med. 2013, 60, 211–222. [Google Scholar] [CrossRef] [PubMed]
- Guzy, J.; Chovanova, Z.; Marekova, M.; Chavkova, Z.; Tomeckova, V.; Mojzisova, G.; Kusnir, J. Effect of quercetin on paracetamol-induced rat liver mitochondria dysfunction. Biol. Bratisl. 2004, 59, 399–403. [Google Scholar]
- Du, K.; McGill, M.R.; Xie, Y.; Bajt, M.L.; Jaeschke, H. Resveratrol prevents protein nitration and release of endonucleases from mitochondria during acetaminophen hepatotoxicity. Food Chem. Toxicol. 2015, 81, 62–70. [Google Scholar] [CrossRef] [PubMed]
- Franco, J.L.; Posser, T.; Missau, F.; Pizzolatti, M.G.; Santos, A.R.S.; Souza, D.O.; Aschner, M.; Rocha, J.B.T.; Dafre, A.L.; Farina, M. Structure-activity relationship of flavonoids derived from medicinal plants in preventing methylmercury-induced mitochondrial dysfunction. Environ. Toxicol. Pharmacol. 2010, 30, 272–278. [Google Scholar] [CrossRef] [PubMed]
- Funakoshi-Tago, M.; Nakamura, K.; Tago, K.; Mashino, T.; Kasahara, T. Anti-inflammatory activity of structurally related flavonoids, Apigenin, Luteolin and Fisetin. Int. Immunopharmacol. 2011, 11, 1150–1159. [Google Scholar] [CrossRef] [PubMed]
- Raza, H.; John, A. Implications of altered glutathione metabolism in aspirin-induced oxidative stress and mitochondrial dysfunction in HepG2 cells. PLoS ONE 2012, 7. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, J.; Das, J.; Manna, P.; Sil, P.C. Arjunolic acid, a triterpenoid saponin, prevents acetaminophen (APAP)-induced liver and hepatocyte injury via the inhibition of APAP bioactivation and JNK-mediated mitochondrial protection. Free Radic. Biol. Med. 2010, 48, 535–553. [Google Scholar] [CrossRef] [PubMed]
- Das, J.; Ghosh, J.; Manna, P.; Sil, P.C. Acetaminophen induced acute liver failure via oxidative stress and JNK activation: Protective role of taurine by the suppression of cytochrome P450 2E1. Free Radic. Res. 2010, 44, 340–355. [Google Scholar] [CrossRef] [PubMed]
- Zheng, J.; Zhou, H.H. Effects of the flavonoids on cytochrome P-450 CYP1, 2E1, 3A4 and 19. Yao Xue Xue Bao 2007, 42, 8–12. [Google Scholar] [PubMed]
- Hau, D.K.P.; Gambari, R.; Wong, R.S.M.; Yuen, M.C.W.; Cheng, G.Y.M.; Tong, C.S.W.; Zhu, G.Y.; Leung, A.K.M.; Lai, P.B.S.; Lau, F.Y.; et al. Phyllanthus urinaria extract attenuates acetaminophen induced hepatotoxicity: involvement of cytochrome P450 CYP2E1. Phytomedicine 2009, 16, 751–760. [Google Scholar] [CrossRef] [PubMed]
- Coles, B.; Wilson, I.; Wardman, P.; Hinson, J.A.; Nelson, S.D.; Ketterer, B. The spontaneous and enzymatic reaction of N-acetyl-p-benzoquinonimine with glutathione: A stopped-flow kinetic study. Arch. Biochem. Biophys. 1988, 264, 253–260. [Google Scholar] [CrossRef]
- Henderson, C.J.; Wolf, C.R.; Kitteringham, N.; Powell, H.; Otto, D.; Park, B.K. Increased resistance to acetaminophen hepatotoxicity in mice lacking glutathione S-transferase Pi. Proc. Natl. Acad. Sci. USA 2000, 97, 12741–12745. [Google Scholar] [CrossRef] [PubMed]
- Kitteringham, N.R.; Powell, H.; Jenkins, R.E.; Hamlett, J.; Lovatt, C.; Elsby, R.; Henderson, C.J.; Wolf, C.R.; Pennington, S.R.; Park, B.K. Protein expression profiling of glutathione S-transferase pi null mice as a strategy to identify potential markers of resistance to paracetamol-induced toxicity in the liver. Proteomics 2003, 3, 191–207. [Google Scholar] [CrossRef] [PubMed]
- Bourdi, M.; Masubuchi, Y.; Reilly, T.P.; Amouzadeh, H.R.; Martin, J.L.; George, J.W.; Shah, A.G.; Pohl, L.R. Protection against acetaminophen-induced liver injury and lethality by interleukin 10: Role of inducible nitric oxide synthase. Hepatology 2002, 35, 289–298. [Google Scholar] [CrossRef] [PubMed]
- Dambach, D.M.; Durham, S.K.; Laskin, J.D.; Laskin, D.L. Distinct roles of NF-kappaB p50 in the regulation of acetaminophen-induced inflammatory mediator production and hepatotoxicity. Toxicol. Appl. Pharmacol. 2006, 211, 157–165. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.L.; Wang, Y.J.; Zhang, Q.Y.; Liu, B.; Wang, F.Y.; Li, J.J.; Zhu, R.Z. Hepatoprotective effects of baicalein against CCl4-induced acute liver injury in mice. World J. Gastroenterol. 2012, 18, 6605–6613. [Google Scholar] [CrossRef] [PubMed]
- Taslipinar, M.Y.; Aydin, I.; Kaldirim, U.; Aydin, F.N.; Agilli, M.; Eyi, Y.E.; Tuncer, S.K.; Altayli, E.; Ucar, F.; Macit, E.; et al. Hyperbaric oxygen treatment and N-acetylcysteine ameliorate acetaminophen-induced liver injury in a rat model. Hum. Exp. Toxicol. 2013, 32, 1107–1116. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.H.; Kang, K.S.; Lee, Y.S. The inhibitory effect of genistein on hepatic steatosis is linked to visceral adipocyte metabolism in mice with diet-induced non-alcoholic fatty liver disease. Br. J. Nutr. 2010, 104, 1333–1342. [Google Scholar] [CrossRef] [PubMed]
- Demirel, U.; Harputluoglu, M.M.M.; Us, S.B.; Kaya, E.; Sahin, N.; Aydin, N.E.; Gursoy, S.; Bilgic, Y.; Demirel, M.; Bulut, T.; et al. The effects of Teucrium polium on ionizing radiation-induced intestinal damage in rats. Acta GastroEnterol. Belg. 2011, 74, 491–496. [Google Scholar] [PubMed]
- Henderson, N.C.; Pollock, K.J.; Frew, J.; Mackinnon, A.C.; Flavell, R.A.; Davis, R.J.; Sethi, T.; Simpson, K.J. Critical role of c-jun (NH2) terminal kinase in paracetamol- induced acute liver failure. Gut 2007, 56, 982–990. [Google Scholar] [CrossRef] [PubMed]
- Gandhi, A.; Guo, T.; Ghose, R. Role of c-Jun N-terminal kinase (JNK) in regulating tumor necrosis factor-alpha (TNF-alpha) mediated increase of acetaminophen (APAP) and chlorpromazine (CPZ) toxicity in murine hepatocytes. J. Toxicol. Sci. 2010, 35, 163–173. [Google Scholar] [CrossRef] [PubMed]
- Ferret, P.J.; Hammoud, R.; Tulliez, M.; Tran, A.; Trebeden, H.; Jaffray, P.; Malassagne, B.; Calmus, Y.; Weill, B.; Batteux, F. Detoxification of reactive oxygen species by a nonpeptidyl mimic of superoxide dismutase cures acetaminophen-induced acute liver failure in the mouse. Hepatolog 2001, 33, 1173–1180. [Google Scholar] [CrossRef] [PubMed]
- Manov, I.; Hirsh, M.; Iancu, T.C. N-acetylcysteine does not protect HepG2 cells against acetaminophen-induced apoptosis. Basic Clin. Pharmacol. Toxicol. 2004, 94, 213–225. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Neu-Reagent UV/365 nm | ||||
|---|---|---|---|---|
| Band N° | Rf | Color | Type of Phenol/Possible Flavonoid | |
| Gq | 1 | 0.03 | Blue-white | Flavonols, flavones, isoflavones, flavonones |
| 2 | 0.05 | Orange | Flavonols glycosides | |
| 3 | 0.09 | Yellow-green | Favonols | |
| 4 | 0.14 | Yellow-pale | Flavonols, flavones, isoflavones, flavonones | |
| 5 | 0.18 | Blue | Phenolic acid | |
| 6 | 0.26 | Blue | Flavonoïd aglycones | |
| 7 | 0.32 | Orange | Flavonols, flavones, isoflavones, flavonones | |
| 8 | 0.36 | Red | Methylated flavones | |
| 9 | 0.42 | Orange | Not identified | |
| 10 | 0.50 | Yellow | Phenolic acid | |
| 11 | 0.55 | Yellow-pale | Phenolic acid | |
| 12 | 0.78 | Blue fluorescent | Phenolic acid | |
| Tp | 1′ | 0.05 | Blue-white fluorescent | Phenolic acid |
| 2′ | 0.12 | Yellow-green | Not identified | |
| 3′ | 0.19 | Blue-white | Phenolic acid, isoflavones, flavonones | |
| 4′ | 0.26 | Yellow-orange | Flavonols | |
| 5′ | 0.31 | Yellow-green | Flavonols, flavones, isoflavones, flavonones | |
| 6′ | 0.36 | Orange | Flavonols-glycosides | |
| 7′ | 0.44 | Red | Not identified | |
| 8′ | 0.54 | Blue-white | Not identified | |
| 9′ | 0.98 | Blue-white fluorescent | Phenolic acid | |
| Groups | Control | APAP | NAC+APAP | Gq+APAP | Tp+APAP |
|---|---|---|---|---|---|
| TBARs (nmol/g liver) | 53 ± 8 | 87 ± 13 *** | 48 ± 11 ## | 56 ± 9 ## | 58 ± 19 # |
| SOD (U/mg prot) | 14 ± 2 | 11 ± 1 *** | 15 ± 1 ## | 16 ± 2 ## | 15 ± 1 ## |
| GPx (mU/mg prot) | 4192 ± 865 | 2453 ± 487 *** | 4289 ± 398 ## | 5021 ± 69 ## | 4594 ± 448 ## |
| GR (mU/mg prot) | 102 ± 9 | 75 ± 12 *** | 114 ± 10 ## | 123 ± 4 ***,## | 116 ± 8 *,## |
| GSH (nmol/g liver) | 1010 ± 235 | 445 ± 79 *** | 930 ± 213 ## | 807 ± 182 ## | 923 ± 201 ## |
| Groups | Control | APAP | NAC + APAP | Gq + APAP | Tp + APAP |
|---|---|---|---|---|---|
| State 4 § | 110.0 ± 0.8 | 44.6 ± 6.2 *** | 81.9 ± 3.2 ## | 58.7 ± 7.1 | 87.5 ± 2.8 ## |
| State 3 § | 687 ± 122 | 136 ± 14.9 ** | 387 ± 67.1## | 255 ± 63.7 | 577 ± 47.8 ### |
| RCR @ | 6.2 ± 1.1 | 3.0 ± 0.6 * | 4.7 ± 0.7 | 4.3 ± 0.7 | 6.6 ± 0.8 # |
| CI $ | 112 ± 25 | 82 ± 10 ** | 96 ± 15 | 134 ± 28 ### | 147 ± 17 **,### |
| CII $ | 178 ± 43 | 132 ± 30 * | 164 ± 40 | 271 ± 64 **,### | 248 ± 23 **,### |
| CII+III $ | 160 ± 37 | 145 ± 60 | 183 ± 31 | 173 ± 44 | 200 ± 37 |
| CIV $ | 749 ± 155 | 729 ± 155 | 699 ± 113 | 1063 ± 184 **,## | 994 ± 115 **,## |
| CS $ | 324 ± 39 | 262 ± 32 ** | 304 ± 30 # | 372 ± 65 ## | 387 ± 91 ## |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Baali, N.; Belloum, Z.; Baali, S.; Chabi, B.; Pessemesse, L.; Fouret, G.; Ameddah, S.; Benayache, F.; Benayache, S.; Feillet-Coudray, C.; et al. Protective Activity of Total Polyphenols from Genista quadriflora Munby and Teucrium polium geyrii Maire in Acetaminophen-Induced Hepatotoxicity in Rats. Nutrients 2016, 8, 193. https://doi.org/10.3390/nu8040193
Baali N, Belloum Z, Baali S, Chabi B, Pessemesse L, Fouret G, Ameddah S, Benayache F, Benayache S, Feillet-Coudray C, et al. Protective Activity of Total Polyphenols from Genista quadriflora Munby and Teucrium polium geyrii Maire in Acetaminophen-Induced Hepatotoxicity in Rats. Nutrients. 2016; 8(4):193. https://doi.org/10.3390/nu8040193
Chicago/Turabian StyleBaali, Nacera, Zahia Belloum, Samiya Baali, Beatrice Chabi, Laurence Pessemesse, Gilles Fouret, Souad Ameddah, Fadila Benayache, Samir Benayache, Christine Feillet-Coudray, and et al. 2016. "Protective Activity of Total Polyphenols from Genista quadriflora Munby and Teucrium polium geyrii Maire in Acetaminophen-Induced Hepatotoxicity in Rats" Nutrients 8, no. 4: 193. https://doi.org/10.3390/nu8040193