Protective Effect of Gallotannin-Enriched Extract Isolated from Galla Rhois against CCl4-Induced Hepatotoxicity in ICR Mice

 and
and 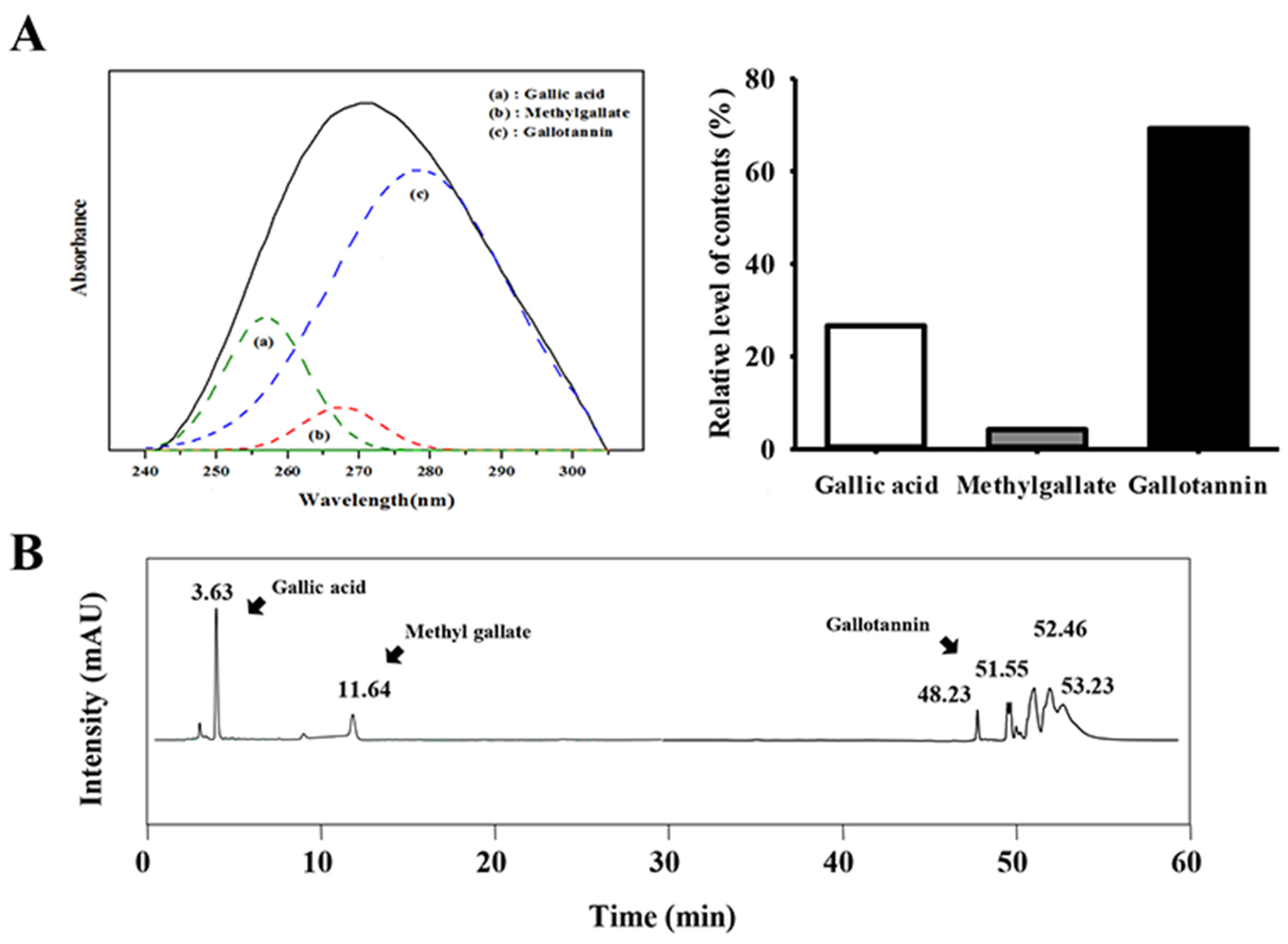{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Experimental Section
2.1. Preparation of GEGR
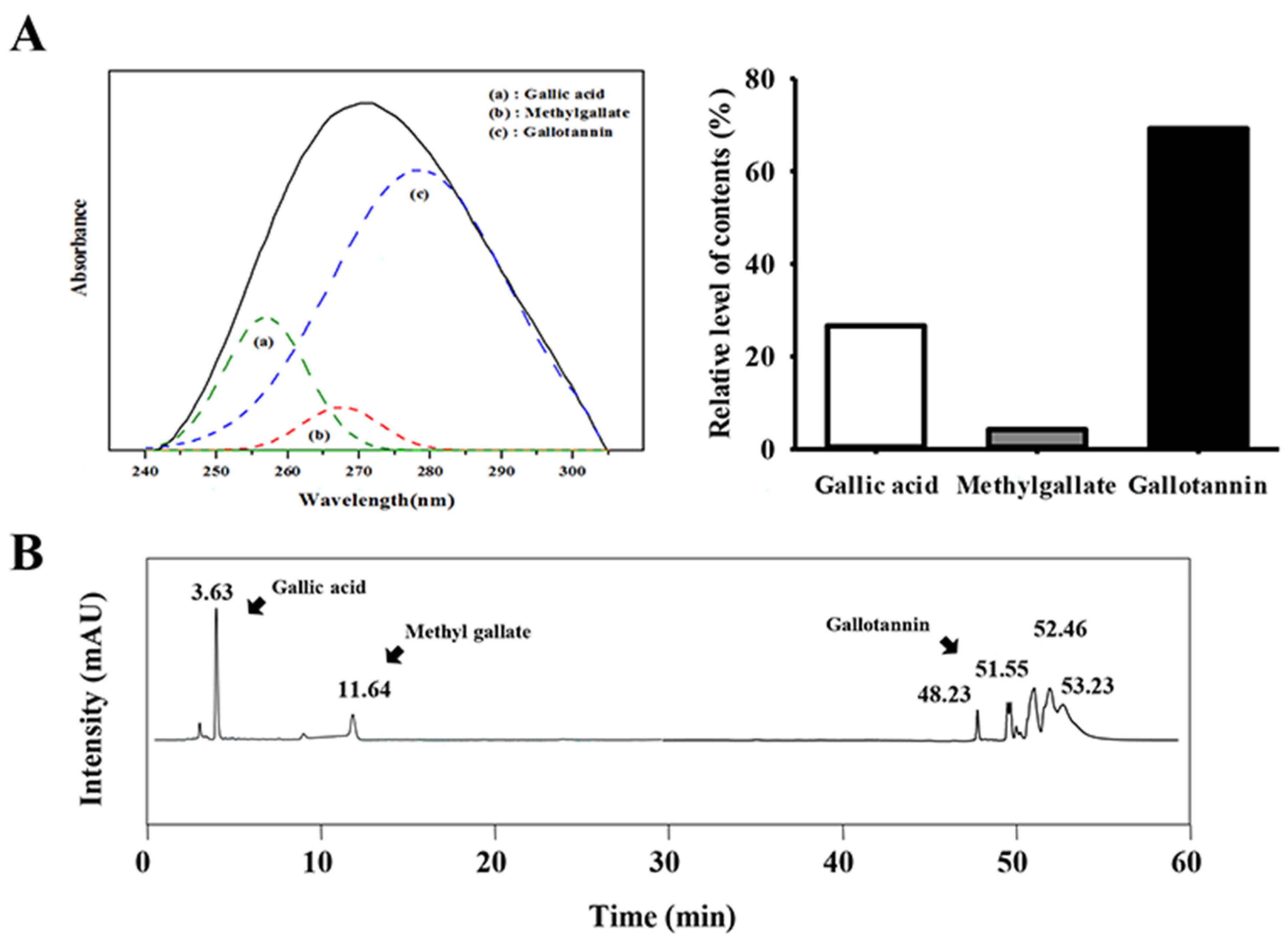
2.2. Analysis of Gallic Acid, Methyl Gallate, and Gallotanin Concentration
2.3. Design of Animal Experiment
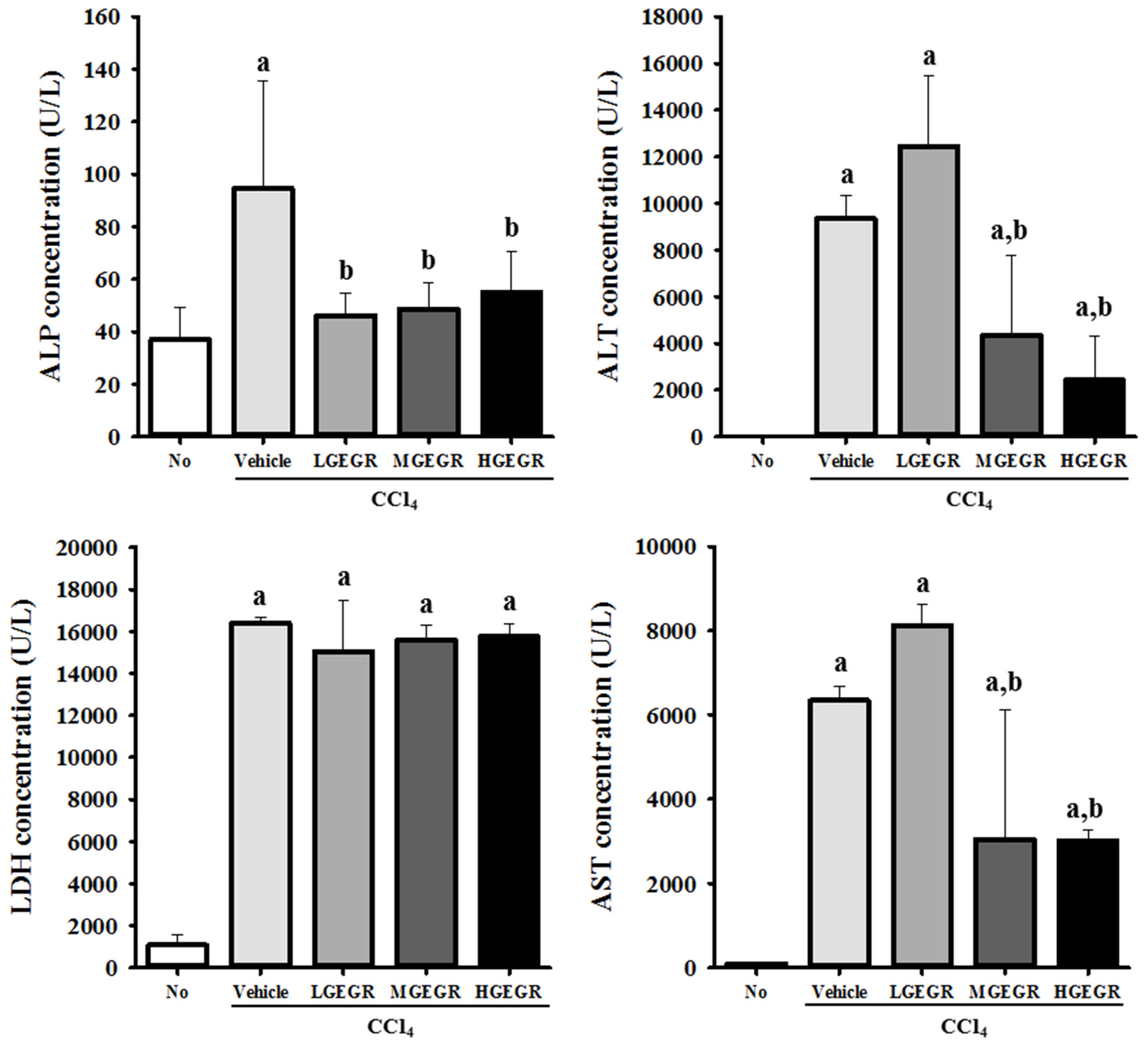
2.4. Serum Biochemistry
2.5. Histological Analysis
2.6. Immunohistochemical Analysis
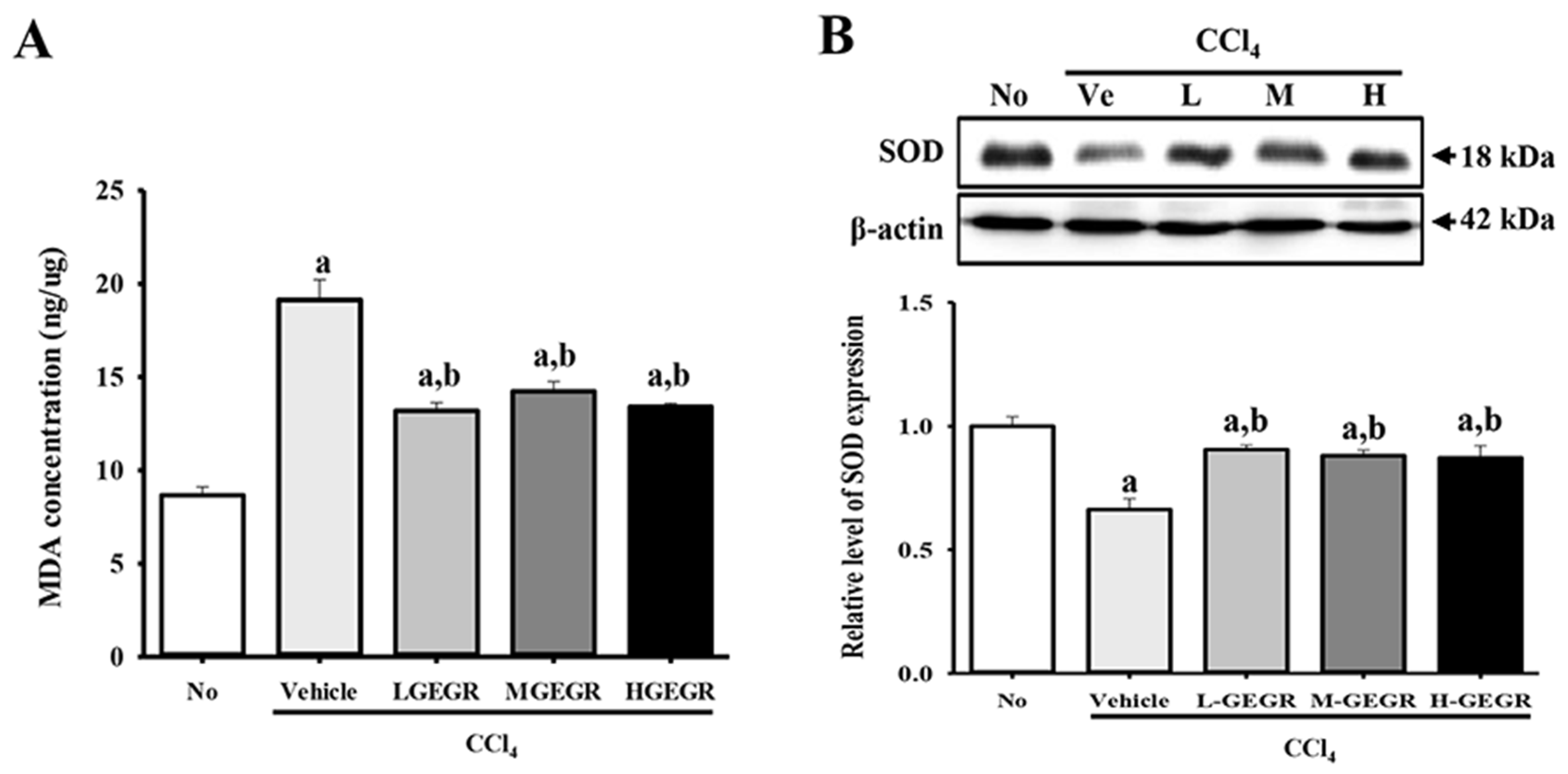
2.7. Determination of Malondialdehyde (MDA) Levels
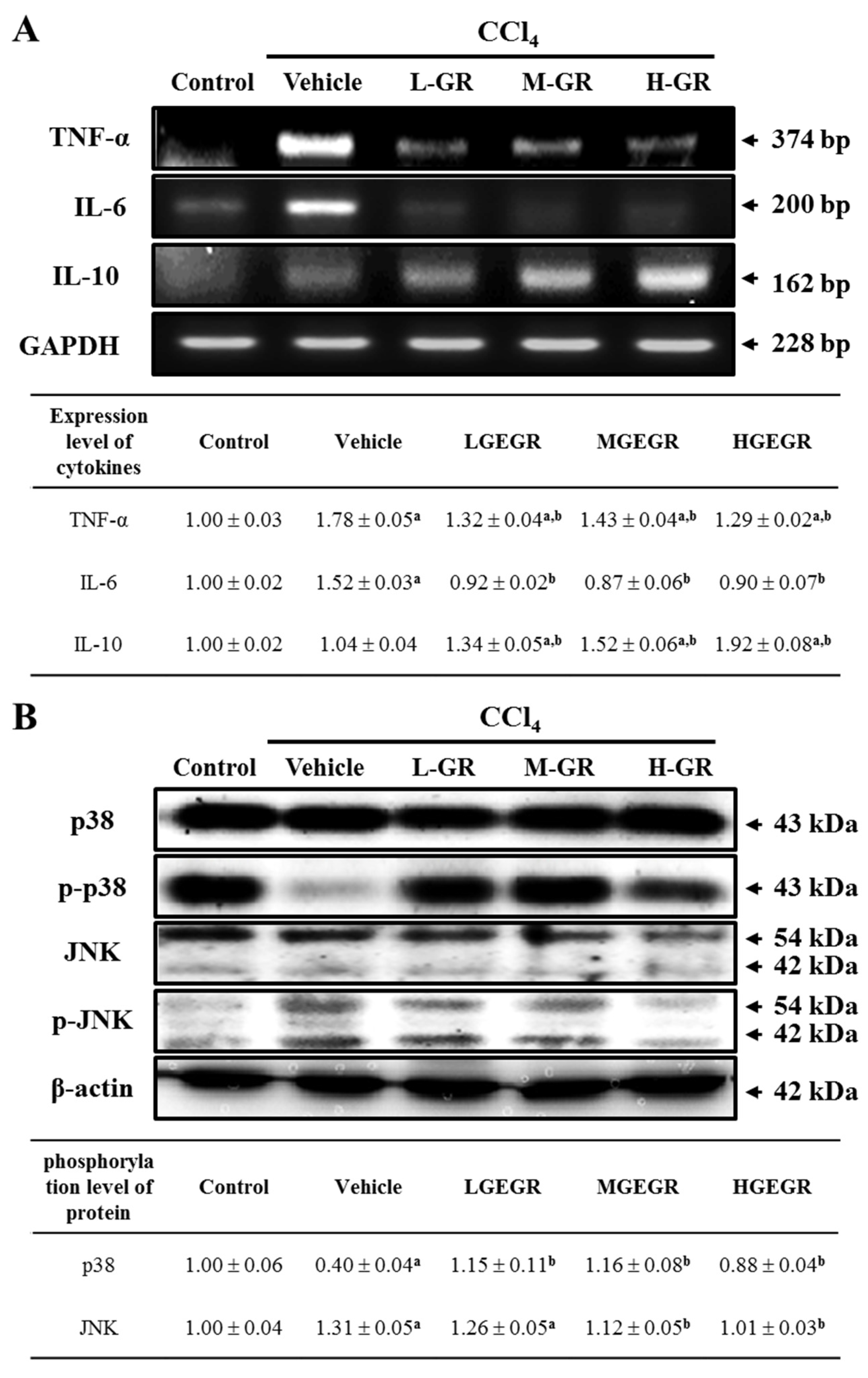
2.8. Western Blot
2.9. Enzyme-Linked Immunosorbent Assay (ELISA) for Tumor Growth Factor (TGF)-β1
2.10. RT-PCR
2.11. Statistical Analysis
3. Results
3.1. Detection of Gallotannin in GEGR
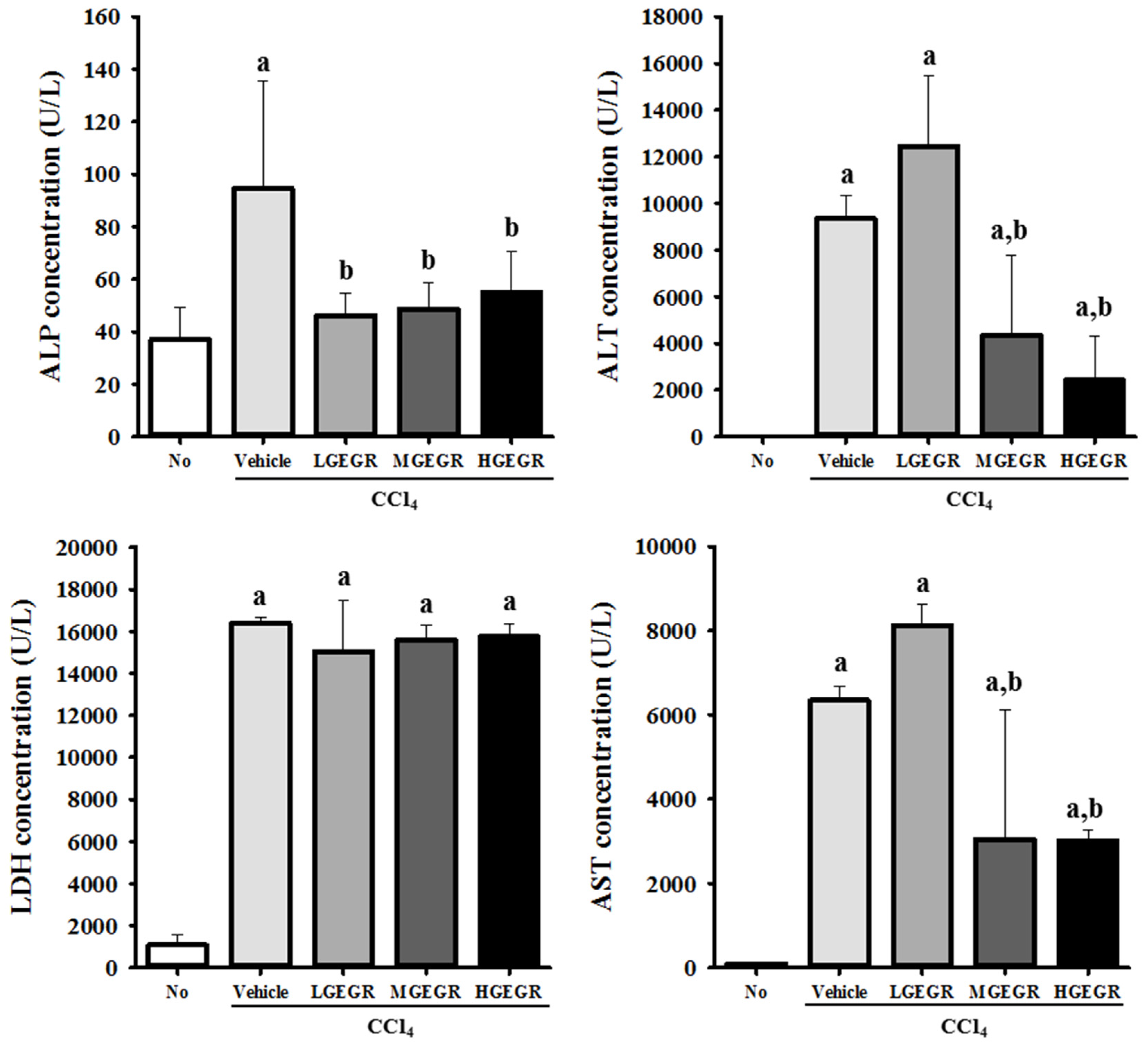
3.2. Protective Effects of GEGR against Hepatic Toxicity
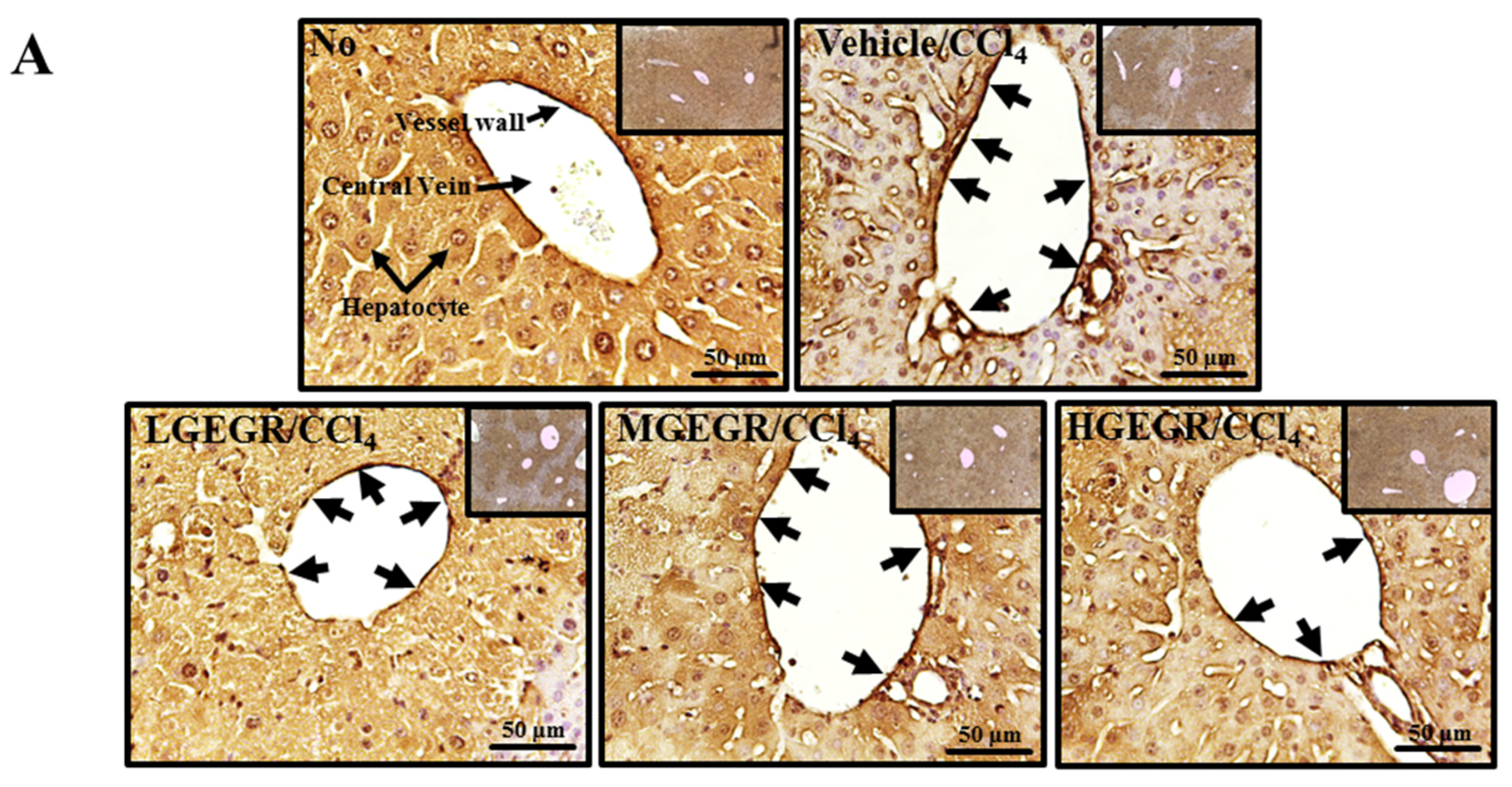
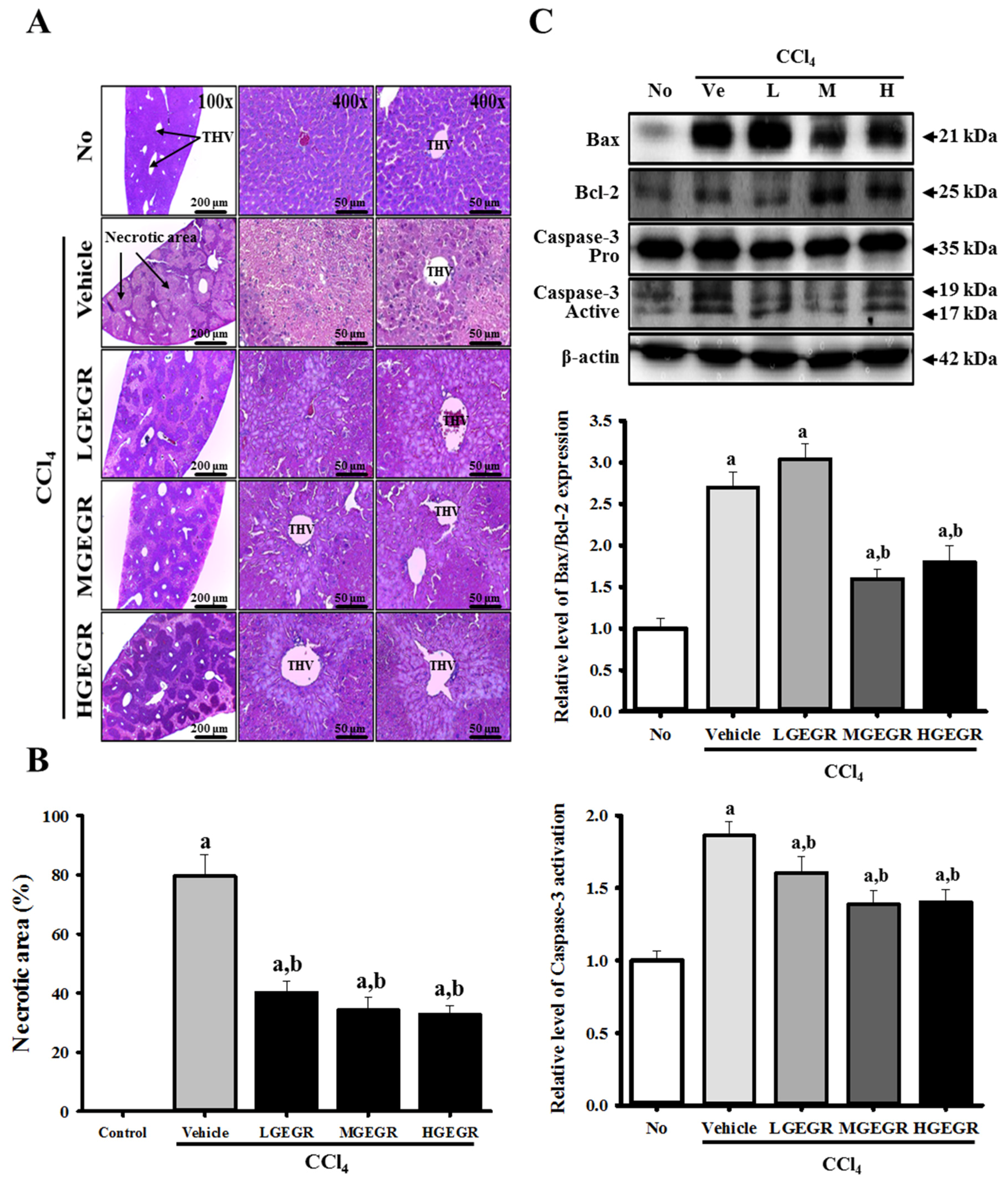
3.3. Protective Effects of GEGR against Alteration of Histopathological Structure
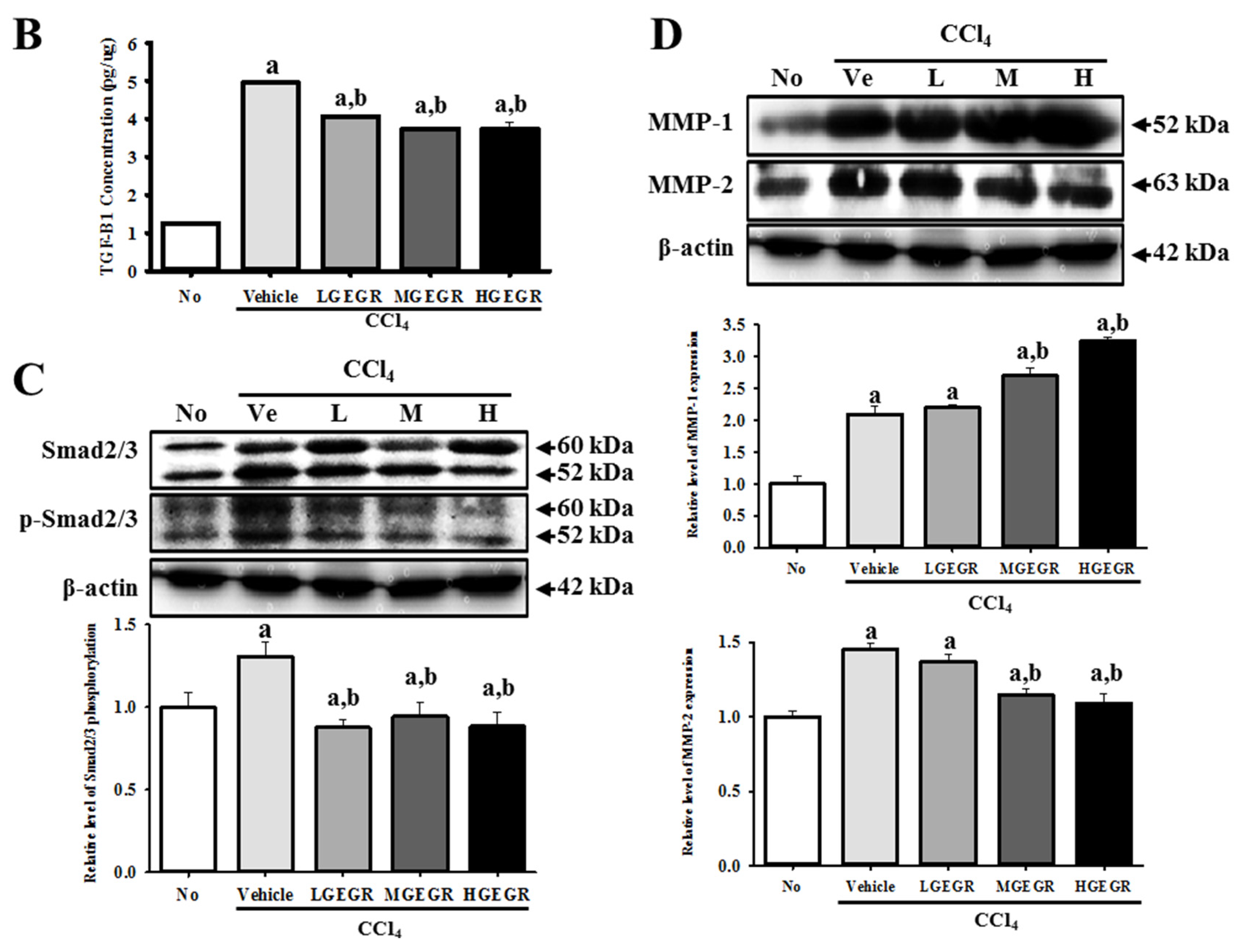
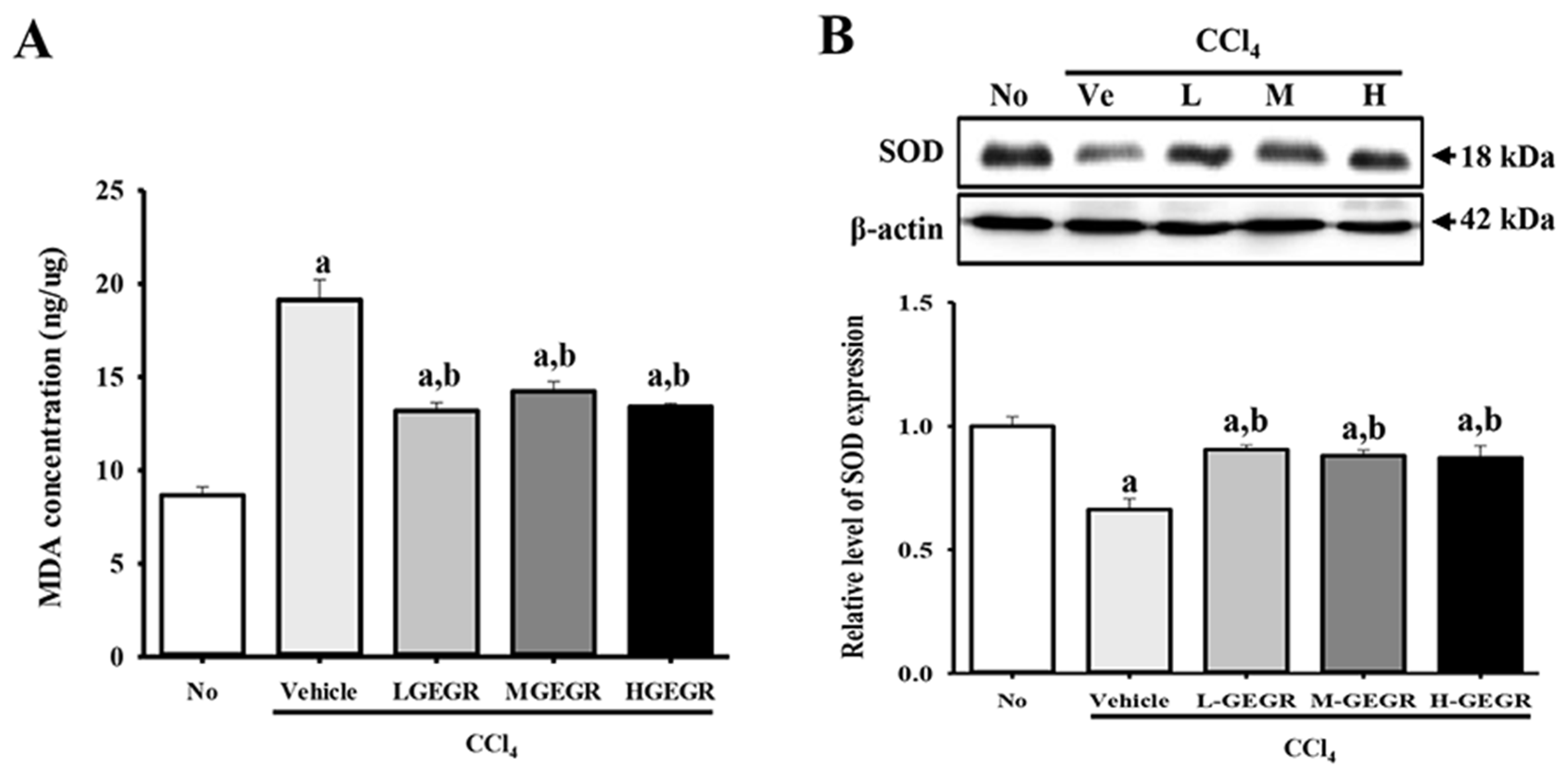
3.4. Protective Effects of GEGR on the Regulation of Oxidative Stress
3.5. Anti-Apoptosis Effects of GEGR Treatment
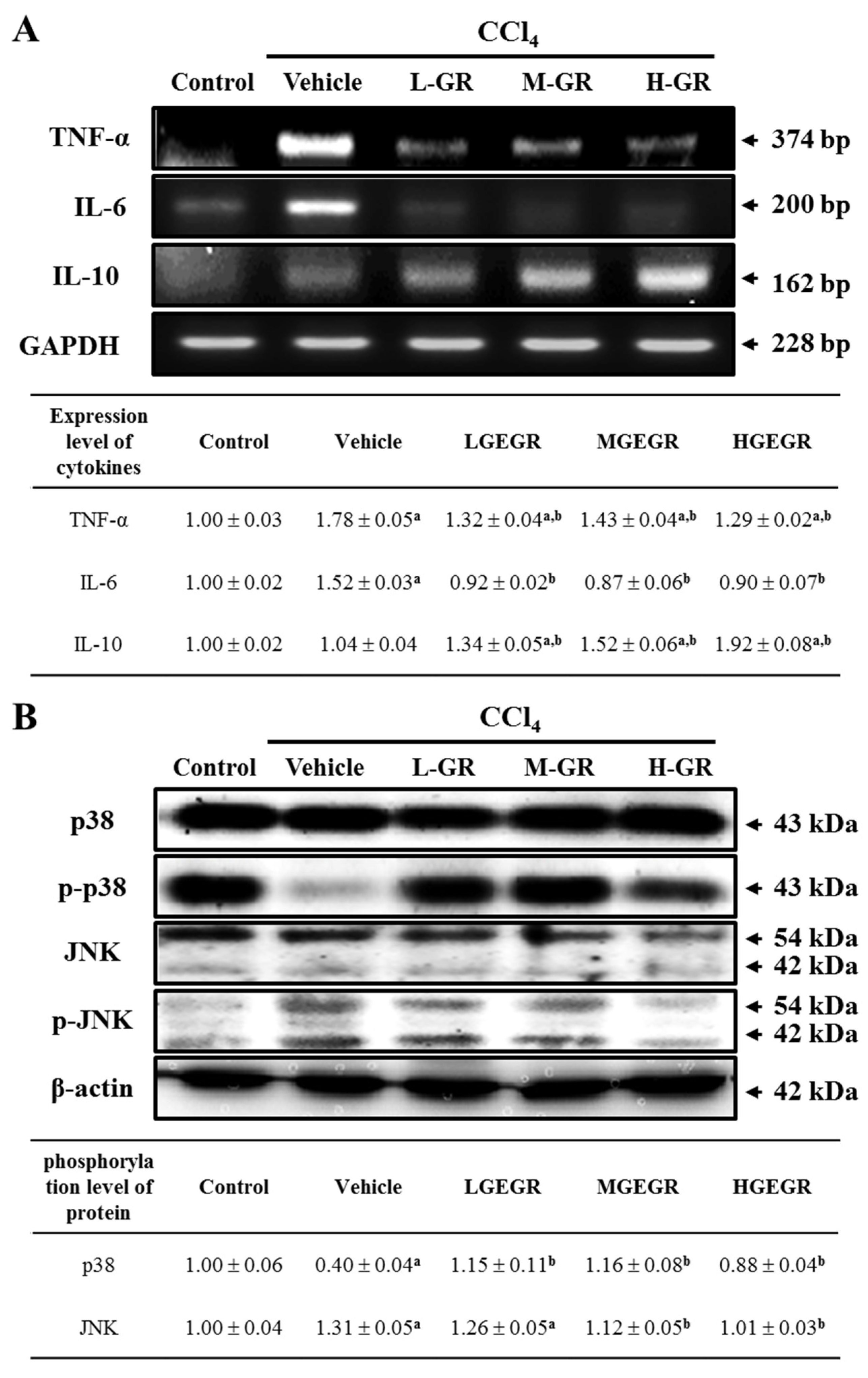
3.6. Anti-Inflammation Effects of GEGR Treatment
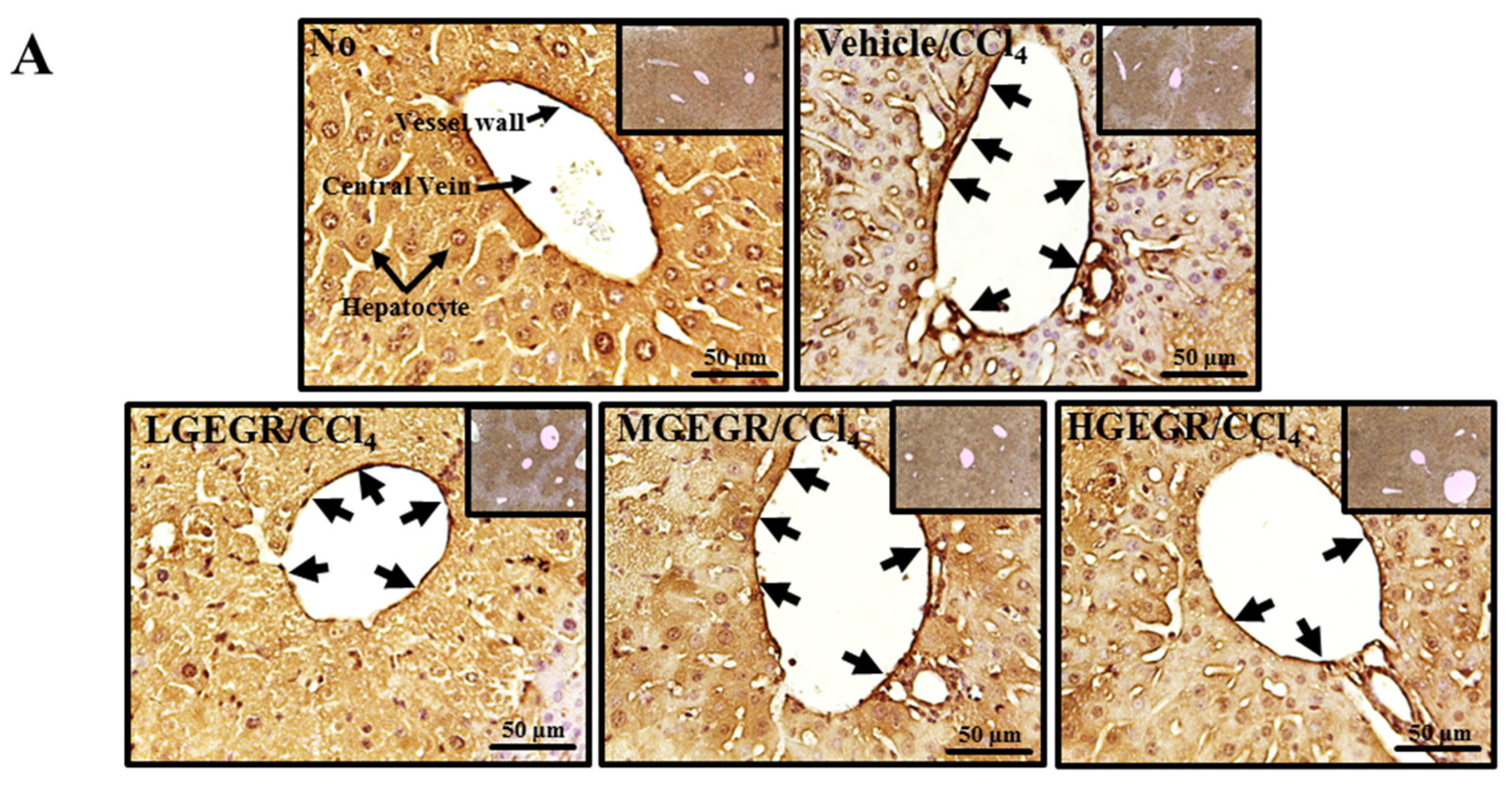
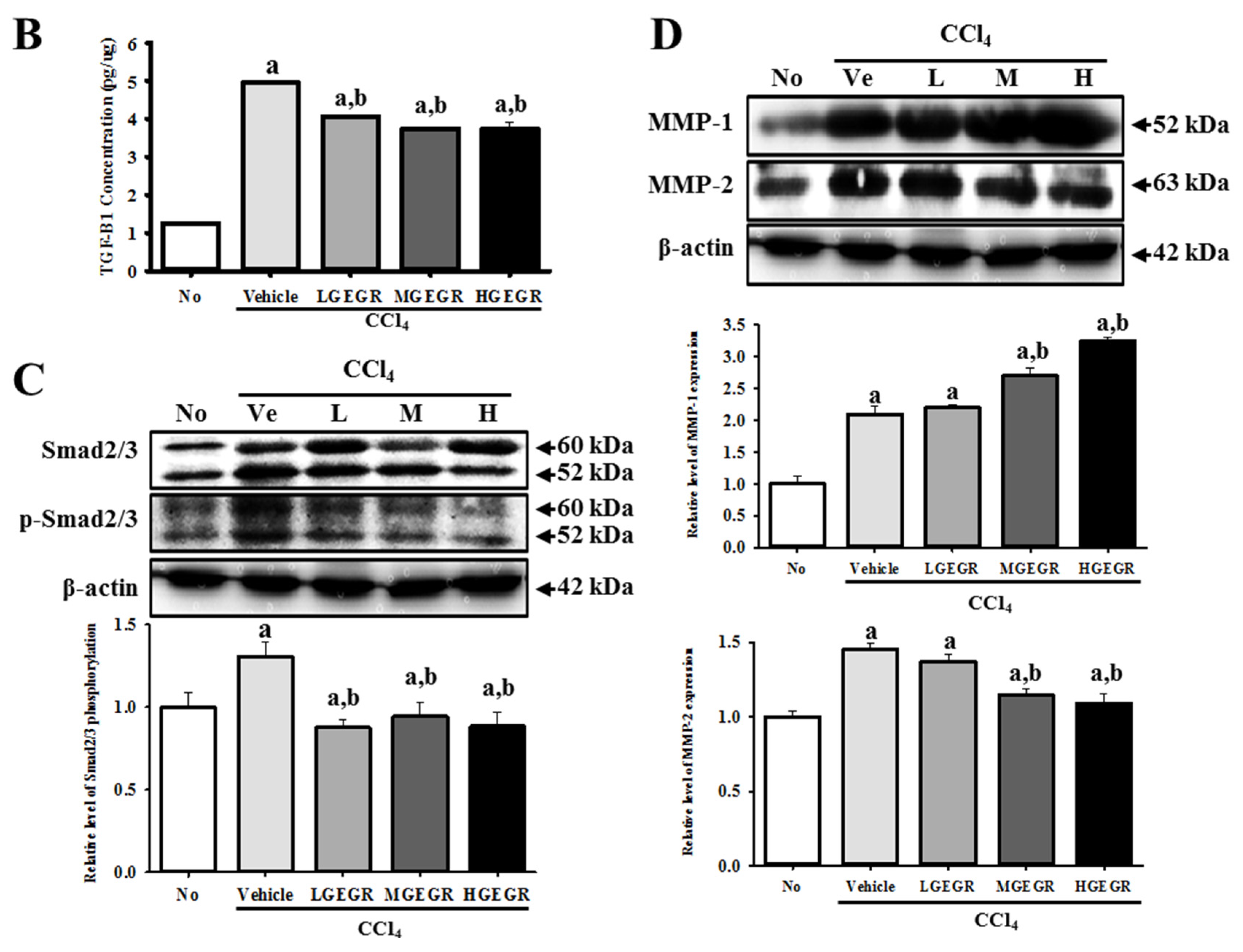
3.7. Protective Effects of GEGR against Liver Fibrosis
4. Discussion
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Besednova, N.N.; Zaporozhets, T.S.; Kuznetsova, T.A.; Kryzhanovskiĭ, S.P.; Kovalev, N.N.; Zviagintseva, T.N. Hepatoprotective effects of extracts and polysaccharides from seaweed. Antibiot. Khimioterapiia 2014, 59, 30–37. [Google Scholar]
- Siviková, K.; Piesová, E.; Dianovský, J. The protection of Vitamin E and selenium against carbon tetrachloride-induced genotoxicity in ovine peripheral blood lymphocytes. Mutat. Res. 2001, 494, 135–142. [Google Scholar] [CrossRef]
- Shi, J.; Aisaki, K.; Ikawa, Y.; Wake, K. Evidence of hepatocyte apoptosis in rat liver after the administration of carbon tetrachloride. Am. J. Pathol. 1998, 153, 515–525. [Google Scholar] [CrossRef]
- Wernke, M.J.; Schell, J.D. Solvents and malignancy. Clin. Occup. Environ. Med. 2004, 4, 513–527. [Google Scholar] [CrossRef] [PubMed]
- Singh, N.; Kamath, V.; Narasimhamurthy, K.; Rajini, P.S. Protective effect of potato peel extract against carbon tetrachloride-induced liver injury in rats. Environ. Toxicol. Pharmacol. 2008, 26, 241–246. [Google Scholar] [CrossRef] [PubMed]
- Sreelatha, S.; Padma, P.R.; Umadevi, M. Protective effects of Coriandrum sativum extracts on carbon tetrachloride-induced hepatotoxicity in rats. Food Chem. Toxicol. 2009, 47, 702–708. [Google Scholar] [CrossRef] [PubMed]
- Srivastava, A.; Shivanandappa, T. Hepatoprotective effect of the aqueous extract of the roots of Decalepis hamiltonii against carbon tetrachloride-induced oxidative stress in rats. Food Chem. 2010, 118, 411–417. [Google Scholar] [CrossRef]
- Sahreen, S.; Khan, M.R.; Khan, R.A. Evaluation of antioxidant activities of various solvent extracts of Carissa opaca fruits. Food Chem. 2010, 122, 1205–1211. [Google Scholar] [CrossRef]
- Lee, S.M.; Lee, D.W.; Park, J.D.; Kim, J.I. Study on Formation and Development of Schlechtendalis chinensis Gall in Rhus javanica. Korean J. Appl. Entomol. 1997, 36, 83–87. [Google Scholar]
- Ren, Z.; Zhu, B.; Wang, D.; Ma, E.; Su, D.; Zhong, Y. Comparative population structure of Chinese sumac aphid Schlechtendalia chinensis and its primary host-plant Rhus chinensis. Genetica 2008, 132, 103–112. [Google Scholar] [CrossRef] [PubMed]
- Song, G.Y.; Park, B.J.; Kim, S.H. Antithrombotic effect of galla rhois. Korean J. Pharmacogn. 2002, 33, 120–123. [Google Scholar]
- Shim, Y.J.; Doo, H.K.; Ahn, S.Y.; Kim, Y.S.; Seong, J.K.; Park, I.S.; Min, B.H. Inhibitory effect of aqueous extract from the gall of Rhus chinensis on alpha-glucosidase activity and postprandial blood glucose. J. Ethnopharmacol. 2003, 85, 283–287. [Google Scholar] [CrossRef]
- Kim, S.H.; Park, H.H.; Lee, S.; Jun, C.D.; Choi, B.J.; Kim, S.Y.; Kim, S.H.; Kim, D.K.; Park, J.S.; Chae, B.S.; et al. The anti-anaphylactic effect of the gall of Rhus javanica is mediated through inhibition of histamine release and inflammatory cytokine secretion. Int. Immunopharmacol. 2005, 5, 1820–1829. [Google Scholar] [CrossRef] [PubMed]
- Ahn, Y.J.; Lee, C.O.; Kweon, J.H.; Ahn, J.W.; Park, J.H. Growth-inhibitory effects of Galla Rhois-derived tannins on intestinal bacteria. J. Appl. Microbiol. 1998, 84, 439–443. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.G.; Kang, O.H.; Lee, Y.S.; Oh, Y.C.; Chae, H.S.; Jang, H.J.; Kim, J.H.; Sohn, D.H.; Shin, D.W.; Park, H.; et al. In vitro activity of methyl gallate isolated from galla rhois alone and in combination with ciprofloxacin against clinical isolates of salmonella. J. Microbiol. Biotechnol. 2008, 18, 1848–1852. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.G.; Kang, O.H.; Lee, Y.S.; Oh, Y.C.; Chae, H.S.; Jang, H.J.; Shin, D.W.; Kwon, D.Y. Antibacterial activity of methyl gallate isolated from Galla Rhois or carvacrol combined with nalidixic acid against nalidixic acid resistant bacteria. Molecules 2009, 14, 1773–1780. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.G.; Mun, S.H.; Chahar, H.S.; Bharaj, P.; Kang, O.H.; Kim, S.G.; Shin, D.W.; Kwon, D.Y. Methyl gallate from Galla rhois successfully controls clinical isolates of Salmonella infection in both in vitro and in vivo systems. PLoS ONE 2014, 9. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.J.; Bae, J.H.; Kim, D.H.; Lim, J.J.; Kim, D.G.; Lee, H.J.; Min, W.; Rhee, M.H.; Chang, H.H.; Park, H.; et al. Intracellular replication inhibitory effects of Galla Rhois ethanol extract for Brucella abortus infection. J. Ethnopharmacol. 2011, 138, 602–609. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.J.; Kim, D.H.; Lim, J.J.; Kim, D.G.; Min, W.; Kim, G.S.; Lee, H.J.; Rhee, M.H.; Park, H.; Kim, S.C.; et al. Anticoccidial effect of supplemental dietary Galla Rhois against infection with Eimeria tenella in chickens. Avian Pathol. 2012, 41, 403–407. [Google Scholar] [CrossRef] [PubMed]
- Chae, H.S.; Kang, O.H.; Choi, J.G.; Oh, Y.C.; Lee, Y.S.; Brice, O.O.; Chong, M.S.; Lee, K.N.; Shin, D.W.; Kwon, D.Y. Methyl gallate inhibits the production of interleukin-6 and nitric oxide via down-regulation of extracellular-signal regulated protein kinase in RAW 264.7 cells. Am. J. Chin. Med. 2010, 38, 973–983. [Google Scholar] [CrossRef] [PubMed]
- Park, P.H.; Hur, J.; Kim, Y.C.; An, R.B.; Sohn, D.H. Involvement of heme oxygenase-1 induction in inhibitory effect of ethyl gallate isolated from Galla Rhois on nitric oxide production in RAW 264.7 macrophages. Arch. Pharm. Res. 2011, 34, 1545–1552. [Google Scholar] [CrossRef] [PubMed]
- Ata, N.; Oku, T.; Hattori, M.; Fujii, H.; Nakajima, M.; Saiki, I. Inhibition by galloylglucose (GG6-10) of tumor invasion through extracellular matrix and gelatinase-mediated degradation of type IV collagens by metastatic tumor cells. Oncol. Res. 1996, 8, 503–511. [Google Scholar] [PubMed]
- Xiang, Q.; Fan, C.; Xiao, S.; Pan, H.; Wang, J.; Zhao, N.; Tian, J. Effect of gallnut extract on nasopharyngeal carcinoma 5–8F cells and its mechanism. Zhong Nan da Xue Xue Bao Yi Xue Ban 2012, 37, 871–875. [Google Scholar] [PubMed]
- Lee, T.Y.; Chang, H.H.; Wang, G.J.; Chiu, J.H.; Yang, Y.Y.; Lin, H.C. Water-soluble extract of Salvia miltiorrhiza ameliorates carbon tetrachloride-mediated hepatic apoptosis in rats. J. Pharm. Pharmacol. 2006, 58, 659–665. [Google Scholar] [CrossRef] [PubMed]
- An, R.B.; Oh, H.C.; Kim, Y.C. Phenolic constituents of galla Rhois with hepatoprotective effects on tacrine- and nitrofurantoin-induced cytotoxicity in Hep G2 cells. Biol. Pharm. Bull. 2005, 28, 2155–2157. [Google Scholar] [CrossRef] [PubMed]
- Park, E.J.; Zhao, Y.Z.; An, R.B.; Kim, Y.C.; Sohn, D.H. 1,2,3,4,6-penta-O-galloyl-beta-d-glucose from Galla Rhois protects primary rat hepatocytes from necrosis and apoptosis. Planta Med. 2008, 11, 1380–1383. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.H. Dyeing, fastness, and deodorizing properties of cotton, silk, and wool fabrics dyed with coffee sludge (Coffea arabica L.) extract. J. Appl. Polym. Sci. 2007, 103, 251–257. [Google Scholar] [CrossRef]
- Hwang, E.K.; Lee, Y.H.; Kim, H.D. Dyeing and deodorizing properties of cotton, silk, and wool fabrics dyed with various natural colorants. J. Korean Soc. Dyers Finish. 2007, 19, 12–20. [Google Scholar]
- Lee, Y.H.; Hwang, E.K.; Jung, Y.J.; Do, S.K.; Kim, H.D. Dyeing and deodorizing properties of cotton, silk, wool fabrics dyed with Amur Cork tree, Dryopteris crassirhizoma, Chrysanthemum boreale, Artemisia Extracts. J. Appl. Polym. Sci. 2010, 115, 2246–2253. [Google Scholar] [CrossRef]
- Lee, Y.H.; Hwang, E.K.; Baek, Y.M.; Lee, M.S.; Lee, D.J.; Jung, Y.J.; Kim, H.D. Deodorizing and antibacterial performance of cotton, silk and wool fabrics dyed with Punica granatum L. extracts. Fiber Polym. 2013, 14, 1445–1453. [Google Scholar] [CrossRef]
- Albu, M.G.; Ghica, M.V.; Giurginca, M.; Trandafir, V.; Popa, L.; Cotrut, C. Spectral characteristics and antioxidant properties of tannic acid immobilized on collagen drug-delivery systems. Rev. Chim. 2009, 60, 666–672. [Google Scholar]
- Lee, Y.H.; Hwang, E.K.; Baek, Y.M.; Kim, H.D. Deodorizing function and antibacterial activity of fabrics dyed with gallnut (Galla chinensis) extract. Text. Res. J. 2015, 85, 1045–1054. [Google Scholar] [CrossRef]
- Hwang, D.Y.; Chae, K.R.; Kang, T.S.; Hwang, J.H.; Lim, C.H.; Kang, H.K.; Goo, J.S.; Lee, M.R.; Lim, H.J.; Min, S.H.; et al. Alterations in behavior, amyloid beta-42, caspase-3, and Cox-2 in mutant PS2 transgenic mouse model of Alzheimer’s disease. FASEB J. 2002, 16, 805–813. [Google Scholar] [CrossRef] [PubMed]
- Joensuu, H.; Pylkkänen, L.; Toikkanen, S. Bcl-2 protein expression and long-term survival in breast cancer. Am. J. Pathol. 1994, 145, 1191–1198. [Google Scholar] [PubMed]
- Apakama, I.; Robinson, M.C.; Walter, N.M.; Charlton, R.G.; Royds, J.A.; Fuller, C.E.; Neal, D.E.; Hamdy, F.C. Bcl-2 overexpression combined with p53 protein accumulation correlates with hormone-refractory prostate cancer. Br. J. Urol. 1996, 74, 1258–1262. [Google Scholar] [CrossRef]
- Bissell, D.M. Cell-matrix interaction and hepatic fibrosis. Prog. Liver Dis. 1990, 9, 143–155. [Google Scholar] [PubMed]
- Friedman, S.L.; Millward-Sadler, G.H.; Arthur, M.J.P. Wright’s Liver and Biliary Disease, 3rd ed.; W.B. Saunders: Philadelphia, PA, USA, 1992; pp. 821–881. [Google Scholar]
- Liu, G.T.; Zhang, T.M.; Wang, B.E.; Wang, Y.W. Protective action of seven natural phenolic compounds against peroxidative damage to biomembranes. Biochem Pharmacol. 1992, 43, 147–152. [Google Scholar] [CrossRef]
- Domitrović, R.; Jakovac, H.; Blagojević, G. Hepatoprotective activity of berberine is mediated by inhibition of TNF-α, COX-2, and iNOS expression in CCl4-intoxicated mice. Toxicology 2001, 280, 33–43. [Google Scholar] [CrossRef] [PubMed]
- Tian, F.; Li, B.; Ji, B.; Zhang, G.; Luo, Y. Identification and structure-activity relationship of gallotannins separated from Galla chinensis. Food Sci. Technol. 2009, 42, 1289–1295. [Google Scholar] [CrossRef]
- Beasley, T.H.; Ziegler, H.W.; Bell, A.D. Determination and characterization of gallotannin by high performance liquid chromatography. Anal. Chem. 1977, 49, 238–243. [Google Scholar] [CrossRef]
- Xiang, P.; Lin, Y.M.; Lin, P.; Xiang, C.; Yang, Z.W.; Lu, Z.M. Effect of cationization reagents on the matrix-assisted laser desorption/ionization time-of-flight mass spectrum of Chinese gallotannins. J. Appl. Polym. Sci. 2007, 105, 859–864. [Google Scholar] [CrossRef]
- Sahreen, S.; Khan, M.R.; Khan, R.A. Hepatoprotective effects of methanol extract of Carissa opaca leaves on CCl4-induced damage in rat. BMC Complement. Altern. Med. 2011, 11, 48–56. [Google Scholar] [CrossRef] [PubMed]
- Brai, B.I.; Adisa, R.A.; Odetola, A.A. Hepatoprotective properties of aqueous leaf extract of Persea Americana, Mill (Lauraceae) “avocado” against CCl4-induced damage in rats. Afr. J. Tradit. Complement. Altern. Med. 2014, 28, 237–244. [Google Scholar] [CrossRef]
- Li, C.; Yi, L.T.; Geng, D.; Han, Y.Y.; Weng, L.J. Hepatoprotective effect of ethanol extract from Berchemia lineate against CCl4-induced acute hepatotoxicity in mice. Pharm. Biol. 2015, 53, 767–772. [Google Scholar] [CrossRef] [PubMed]
- Al-Sayed, E.; El-Lakkany, N.M.; Seif El-Din, S.H.; Sabra, A.N.; Hammam, O.A. Hepatoprotective and antioxidant activity of Melaleuca styphelioides on carbon tetrachloride-induced hepatotoxicity in mice. Pharm. Biol. 2014, 52, 1581–1590. [Google Scholar] [CrossRef] [PubMed]
- Cui, Y.; Yang, X.; Lu, X.; Chen, J.; Zhao, Y. Protective effects of polyphenols-enriched extract from Huangshan Maofeng green tea against CCl4-induced liver injury in mice. Chem. Biol. Interact. 2014, 220, 75–83. [Google Scholar] [CrossRef] [PubMed]
- Srilaxmi, P.; Sareddy, G.R.; Kavi Kishor, P.B.; Setty, O.H.; Babu, P.P. Protective efficacy of natansnin, a dibenzoyl glycoside from Salvinia natans against CCl4 induced oxidative stress and cellular degeneration in rat liver. BMC Pharmacol. 2010, 10, 13–25. [Google Scholar] [CrossRef] [PubMed]
- Lu, B.; Xu, Y.; Xu, L.; Cong, X.; Yin, L.; Li, H.; Peng, J. Mechanism investigation of dioscin against CCl4-induced acute liver damage in mice. Environ. Toxicol. Pharmacol. 2012, 34, 127–135. [Google Scholar] [CrossRef] [PubMed]
- Guo, X.; Liang, B.; Wang, X.; Fan, F.; Jin, J.; Lan, R.; Yang, J.; Wang, X.; Jin, L.; Cao, Q. Glycyrrhizic acid attenuates CCl4-induced hepatocyte apoptosis in rats via a p53-mediated pathway. World J. Gastroenterol. 2013, 19, 3781–3791. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Lu, B.; Han, X.; Xu, L.; Qi, Y.; Yin, L.; Xu, Y.; Zhao, Y.; Liu, K.; Peng, J. Protection of the flavonoid fraction from Rosa laevigata Michx fruit against carbon tetrachloride-induced acute liver injury in mice. Food Chem. Toxicol. 2013, 55, 60–69. [Google Scholar] [CrossRef] [PubMed]
- Iyer, A.; Fairlie, D.P.; Prins, J.B.; Hammock, B.D.; Brown, L. Inflammatory lipid mediators in adipocyte function and obesity. Nat. Rev. Endocrinol. 2010, 6, 71–82. [Google Scholar] [CrossRef] [PubMed]
- Muñoz, A.; Costa, M. Nutritionally mediated oxidative stress and inflammation. Oxid. Med. Cell Longev. 2013, 2013. [Google Scholar] [CrossRef] [PubMed]
- Reyes-Gordillo, K.; Segovia, J.; Shibayama, M.; Vergara, P.; Moreno, M.G.; Muriel, P. Curcumin protects against acute liver damage in the rat by inhibiting NF-kappaB, proinflammatory cytokines production and oxidative stress. Biochim. Biophys. Acta 2007, 1770, 989–996. [Google Scholar] [CrossRef] [PubMed]
- Lin, X.; Liu, X.; Huang, Q.; Zhang, S.; Zheng, L.; Wei, L.; He, M.; Jiao, Y.; Huang, J.; Fu, S.; et al. Hepatoprotective effects of the polysaccharide isolated from Tarphochlamys affinis (Acanthaceae) against CCl4-induced hepatic injury. Biol. Pharm. Bull. 2012, 35, 1574–1580. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.Q.; Ding, J.; Zhang, L.; Liu, C.M. Ursolic acid protects mouse liver against CCl4-induced oxidative stress and inflammation by the MAPK/NF-κB pathway. Environ. Toxicol. Pharmacol. 2014, 37, 975–983. [Google Scholar] [CrossRef] [PubMed]
- Hou, Y.L.; Tsai, Y.H.; Lin, Y.H.; Chao, J.C. Ginseng extract and ginsenoside Rb1 attenuate carbon tetrachloride-induced liver fibrosis in rats. BMC Complement. Altern. Med. 2014, 14, 415–425. [Google Scholar] [CrossRef] [PubMed]
- Hazem, S.H.; Shaker, M.E.; Ashamallah, S.A. The novel Janus kinase inhibitor ruxolitinib confers protection against carbon tetrachloride-induced hepatotoxicity via multiple mechanisms. Chem. Biol. Interact. 2014, 220, 116–127. [Google Scholar] [CrossRef] [PubMed]
- Friedman, S.L.; Maher, J.J.; Bissell, D.M. Mechanisms and therapy of hepatic fibrosis: report of the AASLD Single Topic Basic Research Conference. Hepatology 2000, 32, 1403–1408. [Google Scholar] [CrossRef] [PubMed]
- Wells, R.G. The role of matrix stiffness in hepatic stellate cell activation and liver fibrosis. J. Clin. Gastroenterol. 2005, 46, 295–303. [Google Scholar] [CrossRef]
- Lin, X.; Zhang, S.; Huang, Q.; Wei, L.; Zheng, L.; Chen, Z.; Jiao, Y.; Huang, J.; Fu, S.; Huang, R. Protective effect of Fufang-Liu-Yue-Qing, a traditional Chinese herbal formula, on CCl4 induced liver fibrosis in rats. J. Ethnopharmacol. 2012, 142, 548–556. [Google Scholar] [CrossRef] [PubMed]
- Tipoe, G.L.; Leung, T.M.; Liong, E.C.; Lau, T.Y.; Fung, M.L.; Nanji, A.A. Epigallocatechin-3-gallate (EGCG) reduces liver inflammation, oxidative stress and fibrosis in carbon tetrachloride (CCl4)-induced liver injury in mice. Toxicology 2010, 273, 45–52. [Google Scholar] [CrossRef] [PubMed]
- Mata-Santos, H.A.; Lino, F.G.; Rocha, C.C.; Paiva, C.N.; Castelo Branco, M.T.; Pyrrho Ados, S. Silymarin treatment reduces granuloma and hepatic fibrosis in experimental schistosomiasis. Parasitol. Res. 2010, 107, 1429–1434. [Google Scholar] [CrossRef] [PubMed]
- Fu, Y.; Zheng, S.; Lin, J.; Ryerse, J.; Chen, A. Curcumin protects the rat liver from CCl4-caused injury and fibrogenesis by attenuating oxidative stress and suppressing inflammation. Mol. Pharmacol. 2008, 73, 339–409. [Google Scholar] [CrossRef] [PubMed]
- Pu, X.; Fan, W.; Yu, S.; Li, Y.; Ma, X.; Liu, L.; Ren, J.; Zhang, W. Polysaccharides from Angelica and Astragalus exert hepatoprotective effects against carbon-tetrachloride-induced intoxication in mice. Can. J. Physiol. Pharmacol. 2015, 93, 39–43. [Google Scholar] [CrossRef] [PubMed]
- Pareek, A.; Godavarthi, A.; Issarani, R.; Nagori, B.P. Antioxidant and hepatoprotective activity of Fagonia schweinfurthii (Hadidi) Hadidi extract in carbon tetrachloride induced hepatotoxicity in HepG2 cell line and rats. J. Ethnopharmacol. 2013, 150, 973–981. [Google Scholar] [CrossRef] [PubMed]
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Go, J.; Kim, J.E.; Koh, E.K.; Song, S.H.; Sung, J.E.; Lee, H.A.; Lee, Y.H.; Lim, Y.; Hong, J.T.; Hwang, D.Y. Protective Effect of Gallotannin-Enriched Extract Isolated from Galla Rhois against CCl4-Induced Hepatotoxicity in ICR Mice. Nutrients 2016, 8, 107. https://doi.org/10.3390/nu8030107
Go J, Kim JE, Koh EK, Song SH, Sung JE, Lee HA, Lee YH, Lim Y, Hong JT, Hwang DY. Protective Effect of Gallotannin-Enriched Extract Isolated from Galla Rhois against CCl4-Induced Hepatotoxicity in ICR Mice. Nutrients. 2016; 8(3):107. https://doi.org/10.3390/nu8030107
Chicago/Turabian StyleGo, Jun, Ji Eun Kim, Eun Kyoung Koh, Sung Hwa Song, Ji Eun Sung, Hyun Ah Lee, Young Hee Lee, Yong Lim, Jin Tae Hong, and Dae Youn Hwang. 2016. "Protective Effect of Gallotannin-Enriched Extract Isolated from Galla Rhois against CCl4-Induced Hepatotoxicity in ICR Mice" Nutrients 8, no. 3: 107. https://doi.org/10.3390/nu8030107