Endothelium-Dependent Relaxation Effect of Apocynum venetum Leaf Extract via Src/PI3K/Akt Signalling Pathway

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Experimental Section
2.1. Materials
2.2. Animals
2.3. Aortic Ring Preparation and Functional Study
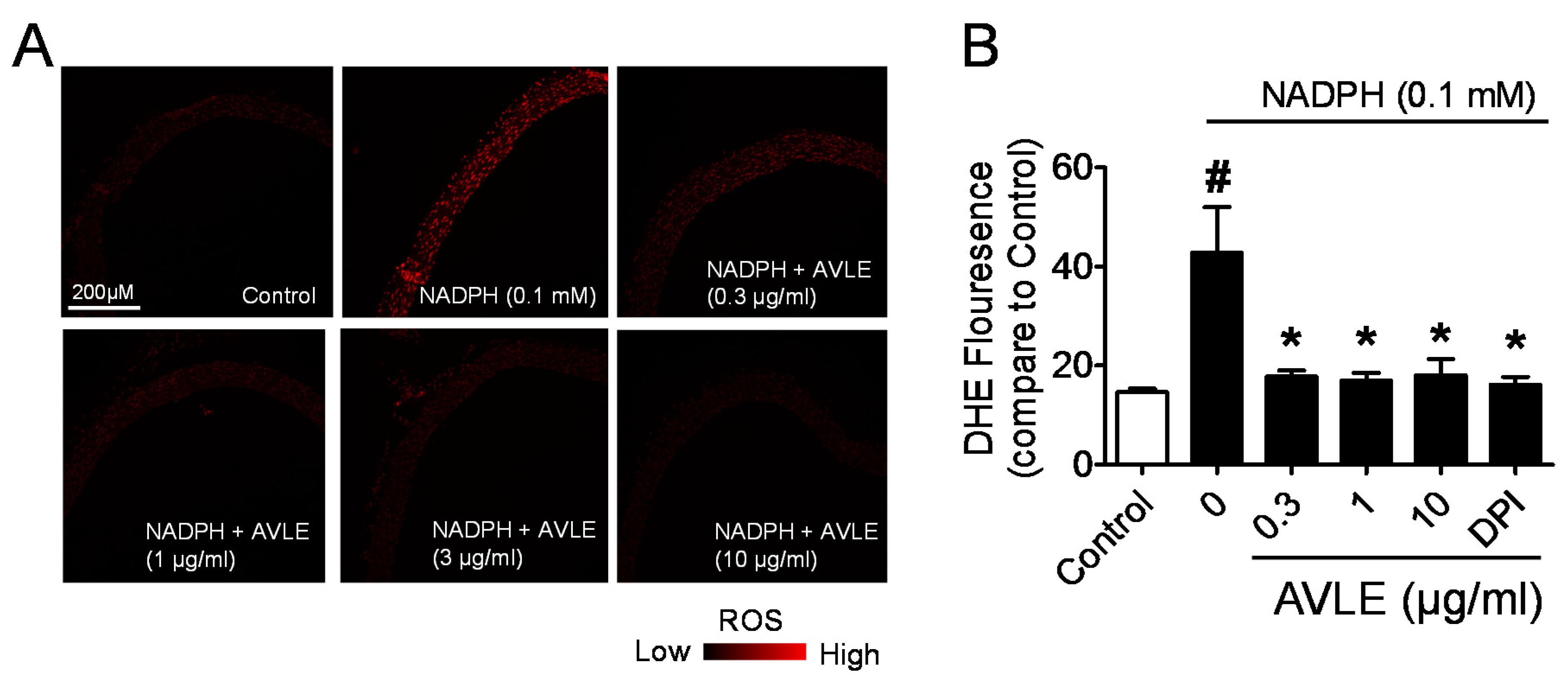
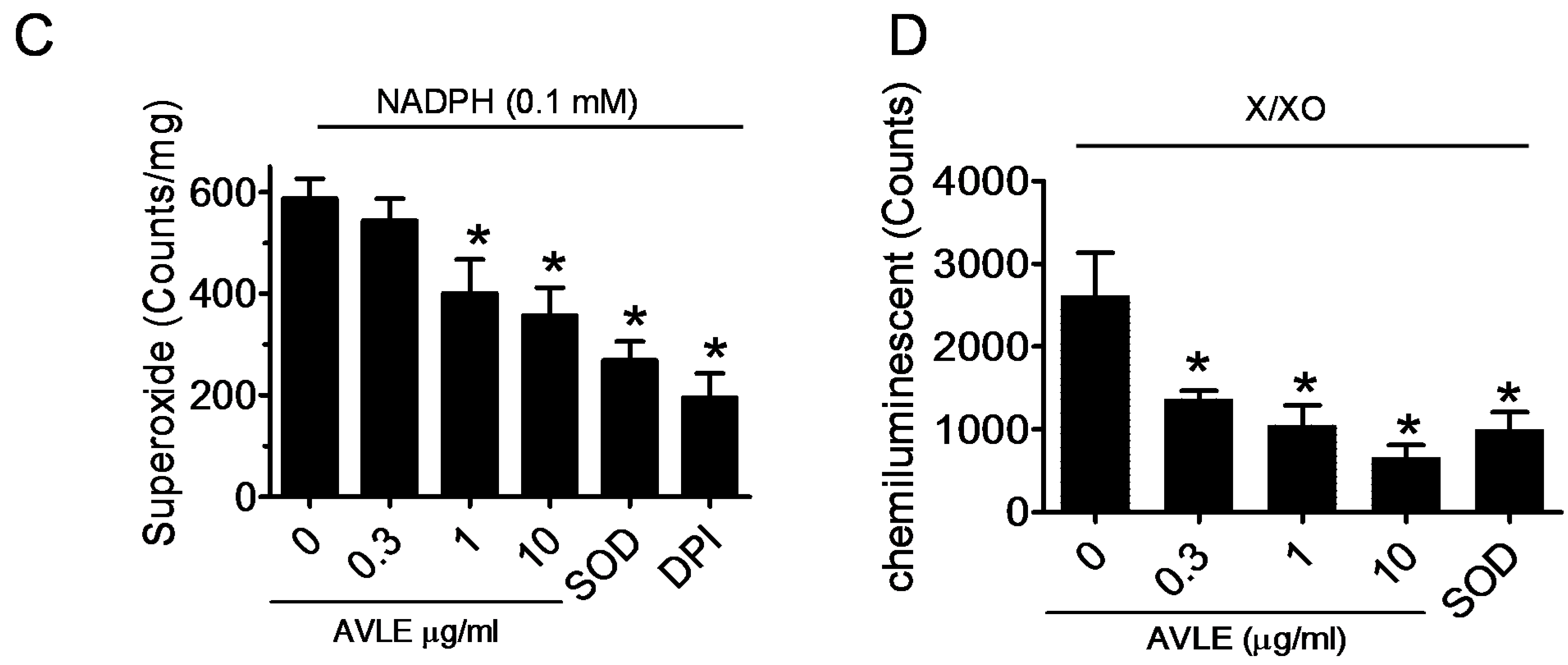
2.4. Measurement of Vascular Superoxide Production
2.4.1. Measurement of NADPH-Mediated Superoxide Production in Aortic Ring and Cryostat Section
2.4.2. Measurement of X/XO Superoxide Production
2.5. Culture of Human Umbilical Vein Endothelial Cells
2.6. Determination of the Phosphorylation Level of Akt and eNOS
2.7. Total Nitrite and Nitrate Detection
2.8. Statistical Analysis
3. Results
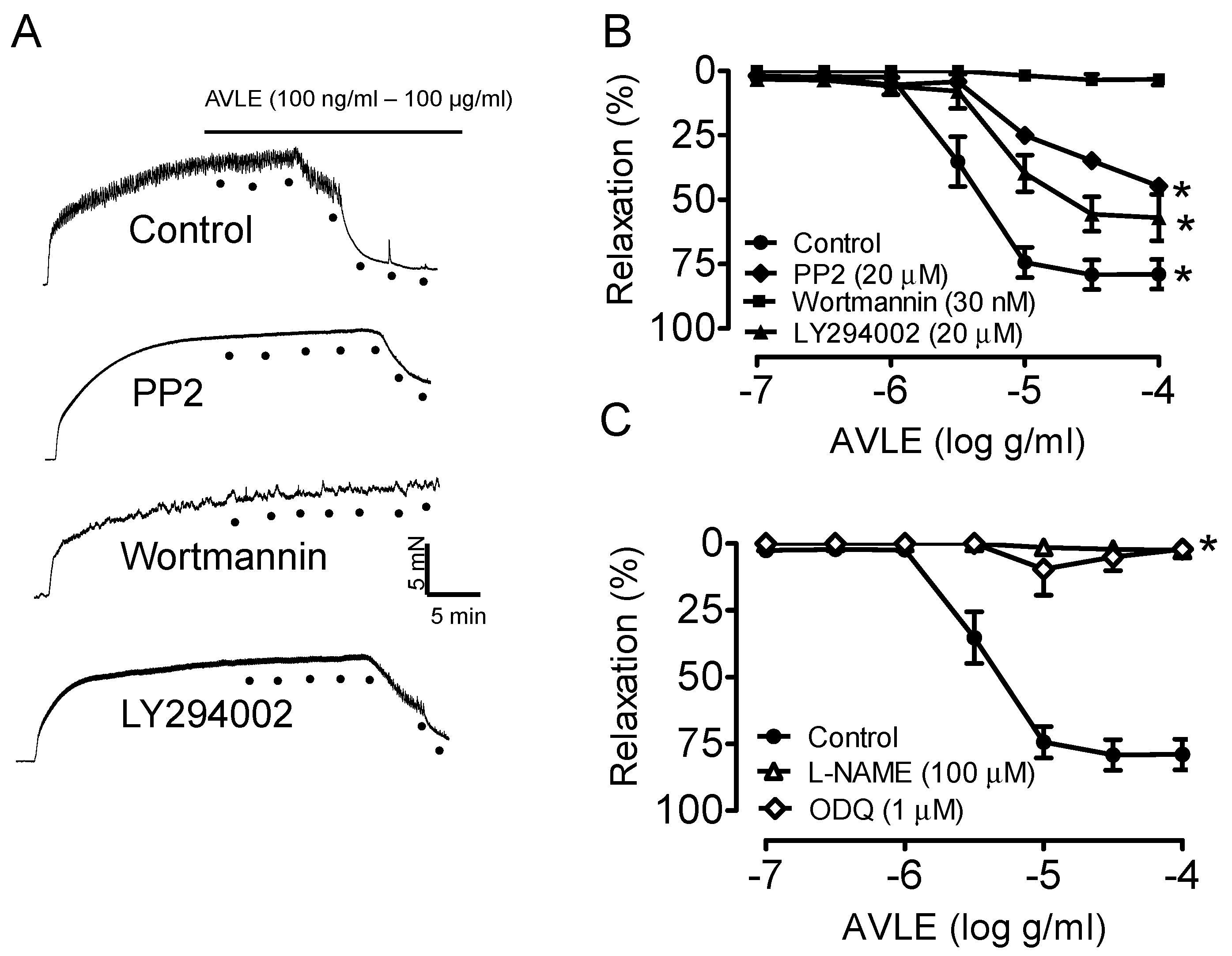
3.1. AVLE Induces Endothelium-Dependent Relaxation in Rat Aortas
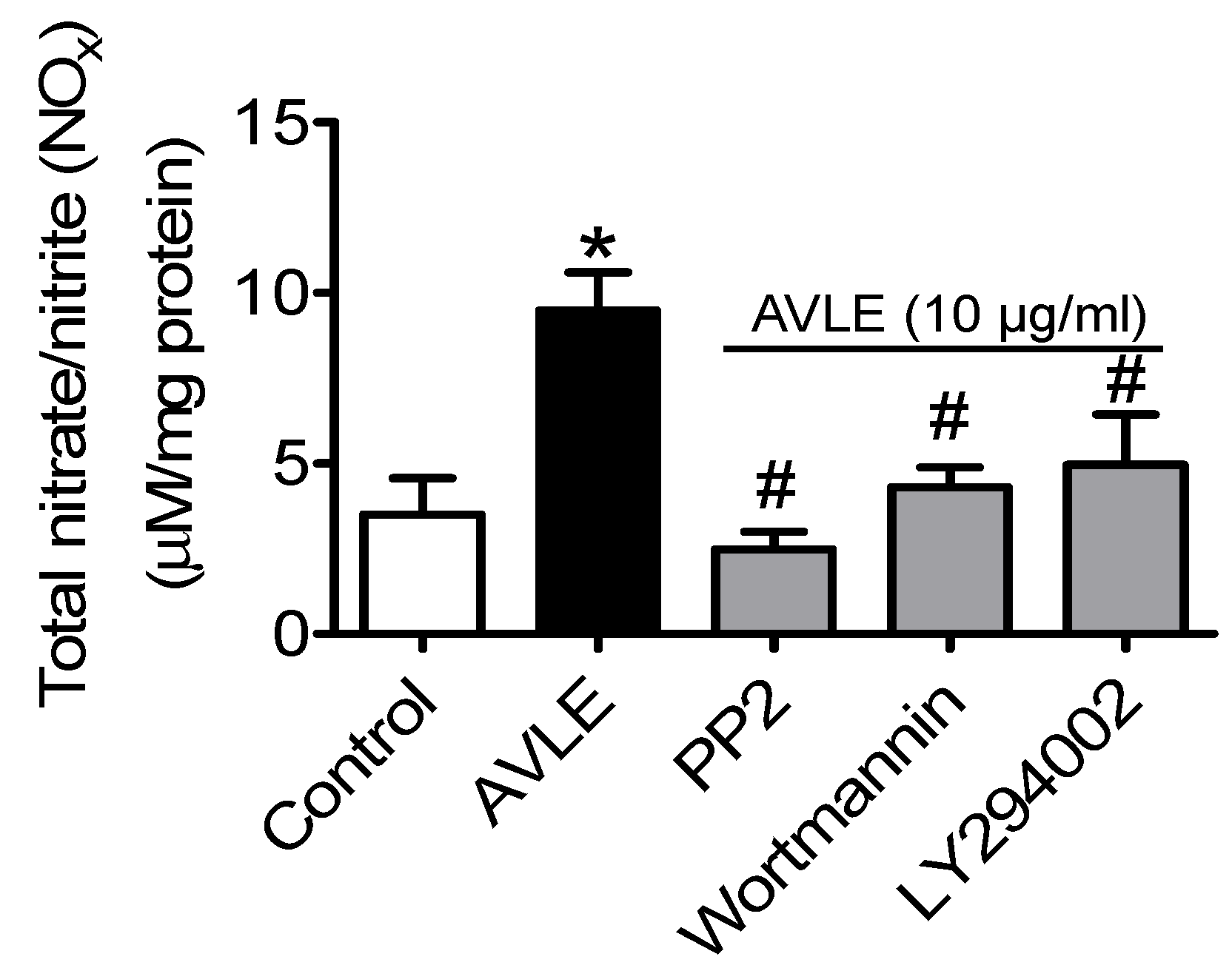
3.2. AVLE Increases Total Nitrite and Nitrate (NOx) Levels in Rat Aortas
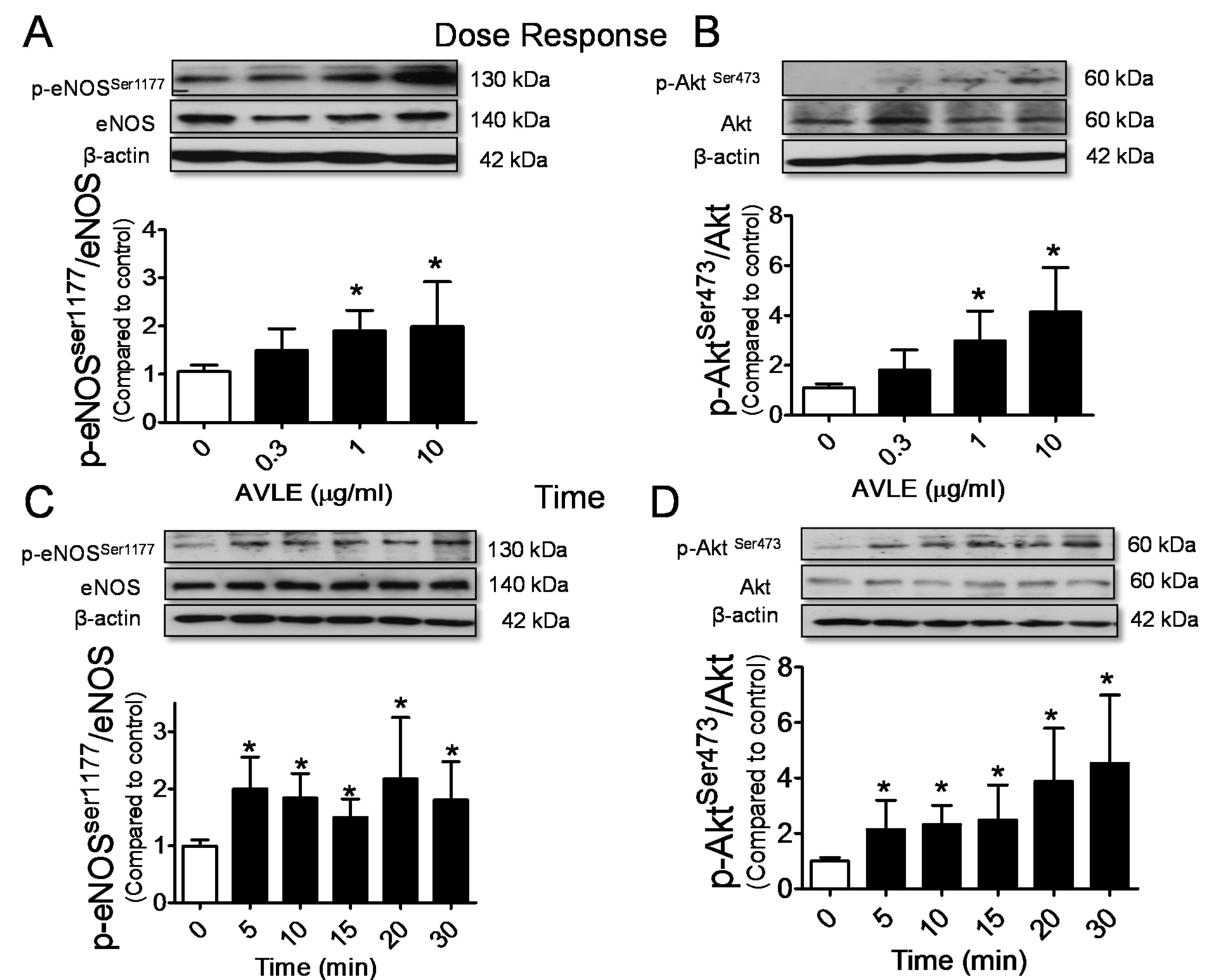
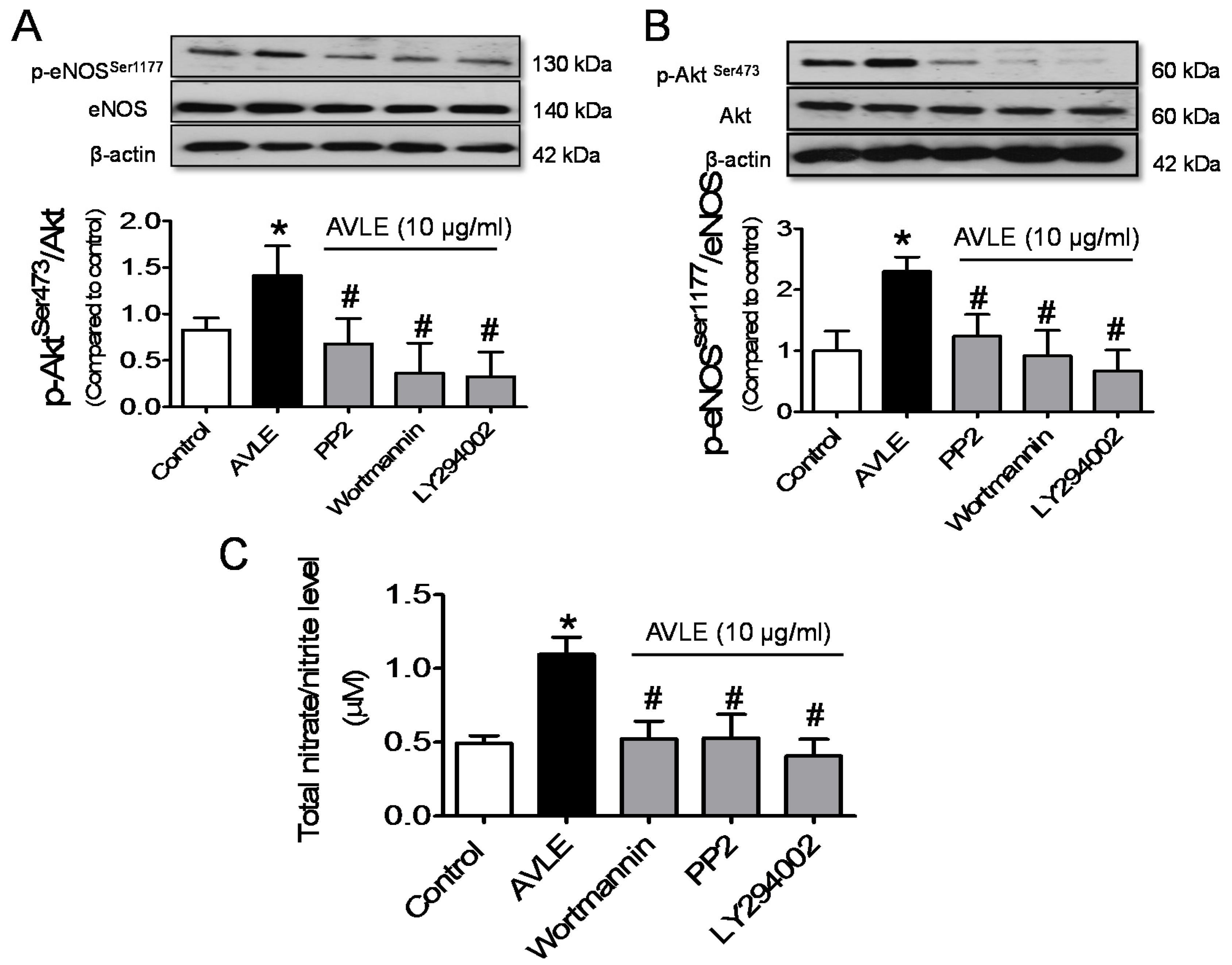
3.3. AVLE Increases Phosphorylated Akt, eNOS and NO Production in HUVECs



3.4. AVLE Inhibits Production of Superoxide Anion in Rat Aortas



4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Oparil, S.; Zaman, M.A.; Calhoun, D.A. Pathogenesis of hypertension. Ann. Intern. Med. 2003, 139, 761–776. [Google Scholar] [CrossRef] [PubMed]
- Intengan, H.D.; Schiffrin, E.L. Vascular remodeling in hypertension: Roles of apoptosis, inflammation, and fibrosis. Hypertension 2001, 38, 581–587. [Google Scholar] [CrossRef] [PubMed]
- Kwan, C.Y. Vascular effects of selected antihypertensive drugs derived from traditional medicinal herbs. Clin. Exp. Pharmacol. Physiol. Suppl. 1995, 22, S297–S299. [Google Scholar] [CrossRef] [PubMed]
- Sutter, M.C.; Wang, Y.X. Recent cardiovascular drugs from chinese medicinal plants. Cardiovasc. Res. 1993, 27, 1891–1901. [Google Scholar] [CrossRef] [PubMed]
- Xiong, X.; Yang, X.; Liu, Y.; Zhang, Y.; Wang, P.; Wang, J. Chinese herbal formulas for treating hypertension in traditional chinese medicine: Perspective of modern science. Hypertens. Res.: Off. J. Jpn. Soc. Hypertens. 2013, 36, 570–579. [Google Scholar] [CrossRef] [PubMed]
- Xiong, Q.; Fan, W.; Tezuka, Y.; Adnyana, I.K.; Stampoulis, P.; Hattori, M.; Namba, T.; Kadota, S. Hepatoprotective effect of apocynum venetum and its active constituents. Planta Med. 2000, 66, 127–133. [Google Scholar] [CrossRef] [PubMed]
- Achike, F.I.; Kwan, C.Y. Nitric oxide, human diseases and the herbal products that affect the nitric oxide signalling pathway. Clin. Exp. Pharmacol. Physiol. 2003, 30, 605–615. [Google Scholar] [CrossRef] [PubMed]
- Schini-Kerth, V.B.; Auger, C.; Kim, J.H.; Etienne-Selloum, N.; Chataigneau, T. Nutritional improvement of the endothelial control of vascular tone by polyphenols: Role of no and edhf. Pflug. Arch. 2010, 459, 853–862. [Google Scholar] [CrossRef] [PubMed]
- Anselm, E.; Chataigneau, M.; Ndiaye, M.; Chataigneau, T.; Schini-Kerth, V.B. Grape juice causes endothelium-dependent relaxation via a redox-sensitive src- and akt-dependent activation of enos. Cardiovasc. Res. 2007, 73, 404–413. [Google Scholar] [CrossRef] [PubMed]
- Madeira, S.V.; Auger, C.; Anselm, E.; Chataigneau, M.; Chataigneau, T.; Soares de Moura, R.; Schini-Kerth, V.B. Enos activation induced by a polyphenol-rich grape skin extract in porcine coronary arteries. J. Vasc. Res. 2009, 46, 406–416. [Google Scholar] [CrossRef] [PubMed]
- Edirisinghe, I.; Burton-Freeman, B.; Varelis, P.; Kappagoda, T. Strawberry extract caused endothelium-dependent relaxation through the activation of pi3 kinase/akt. J. Agric. Food Chem. 2008, 56, 9383–9390. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.; Yokozawa, T.; Hattori, M.; Kadota, S.; Namba, T. Effects of aqueous extracts of apocynum venetum leaves on spontaneously hypertensive, renal hypertensive and nacl-fed-hypertensive rats. J. Ethnopharmacol. 2000, 72, 53–59. [Google Scholar] [CrossRef]
- Kwan, C.Y.; Zhang, W.B.; Nishibe, S.; Seo, S. A novel in vitro endothelium-dependent vascular relaxant effect of apocynum venetum leaf extract. Clin. Exp. Pharmacol. Physiol. 2005, 32, 789–795. [Google Scholar] [CrossRef] [PubMed]
- Cao, Y.; Chu, Q.; Ye, J. Determination of hydroxyl radical by capillary electrophoresis and studies on hydroxyl radical scavenging activities of chinese herbs. Anal. Bioanal. Chem. 2003, 376, 691–695. [Google Scholar] [CrossRef] [PubMed]
- Yokozawa, T.; Kashiwada, Y.; Hattori, M.; Chung, H.Y. Study on the components of luobuma with peroxynitrite-scavenging activity. Biol. Pharm. Bull. 2002, 25, 748–752. [Google Scholar] [CrossRef] [PubMed]
- Irie, K.; Sato, T.; Tanaka, I.; Nakajima, J.; Kawaguchi, M.; Himi, T. Cardiotonic effect of apocynum venetum l. Extracts on isolated guinea pig atrium. J. Natural Med. 2009, 63, 111–116. [Google Scholar] [CrossRef] [PubMed]
- Butterweck, V.; Nishibe, S.; Sasaki, T.; Uchida, M. Antidepressant effects of apocynum venetum leaves in a forced swimming test. Biol. Pharm. Bull. 2001, 24, 848–851. [Google Scholar] [CrossRef] [PubMed]
- Lau, Y.S.; Tian, X.Y.; Mustafa, M.R.; Murugan, D.; Liu, J.; Zhang, Y.; Lau, C.W.; Huang, Y. Boldine improves endothelial function in diabetic db/db mice through inhibition of angiotensin ii-mediated bmp4-oxidative stress cascade. Br. J. Pharmacol. 2013, 170, 1190–1198. [Google Scholar] [CrossRef] [PubMed]
- Fang, J.; Iyer, A.K.; Seki, T.; Nakamura, H.; Greish, K.; Maeda, H. Sma-copolymer conjugate of ahpp: A polymeric inhibitor of xanthine oxidase with potential antihypertensive effect. J. Controll. Release: Off. J. Controll. Release Soc. 2009, 135, 211–217. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Du, L.-j.; Ding, Y.; Xing, D.M.; Wang, W. Studies on chemical constituents from flowers of apocynum venetum. China J. Chin. Mater. Med. 2005, 30, 1340–1342. [Google Scholar]
- Khan, F.; Ray, S.; Craigie, A.M.; Kennedy, G.; Hill, A.; Barton, K.L.; Broughton, J.; Belch, J.J. Lowering of oxidative stress improves endothelial function in healthy subjects with habitually low intake of fruit and vegetables: A randomized controlled trial of antioxidant- and polyphenol-rich blackcurrant juice. Free Radic. Biol. Med. 2014, 72, 232–237. [Google Scholar] [CrossRef]
- Ndiaye, M.; Chataigneau, T.; Chataigneau, M.; Schini-Kerth, V.B. Red wine polyphenols induce edhf-mediated relaxations in porcine coronary arteries through the redox-sensitive activation of the pi3-kinase/akt pathway. Br. J. Pharmacol. 2004, 142, 1131–1136. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, J.A.; Ali, F.; Bailey, L.; Moreno, L.; Harrington, L.S. Role of nitric oxide and prostacyclin as vasoactive hormones released by the endothelium. Exp. Physiol. 2008, 93, 141–147. [Google Scholar] [CrossRef] [PubMed]
- Stuehr, D.J. Enzymes of the l-arginine to nitric oxide pathway. J. Nutr. 2004, 134, 2748S–2751S. [Google Scholar] [PubMed]
- Vallance, P.; Chan, N. Endothelial function and nitric oxide: Clinical relevance. Heart 2001, 85, 342–350. [Google Scholar] [CrossRef] [PubMed]
- Sessa, W.C. Enos at a glance. J. Cell Sci. 2004, 117, 2427–2429. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.P.; Mitchelhill, K.I.; Michell, B.J.; Stapleton, D.; Rodriguez-Crespo, I.; Witters, L.A.; Power, D.A.; Ortiz de Montellano, P.R.; Kemp, B.E. Amp-activated protein kinase phosphorylation of endothelial no synthase. FEBS Lett. 1999, 443, 285–289. [Google Scholar] [CrossRef]
- Feil, R.; Lohmann, S.M.; de Jonge, H.; Walter, U.; Hofmann, F. Cyclic gmp-dependent protein kinases and the cardiovascular system: Insights from genetically modified mice. Circ. Res. 2003, 93, 907–916. [Google Scholar] [CrossRef] [PubMed]
- Dell’Agli, M.; Busciala, A.; Bosisio, E. Vascular effects of wine polyphenols. Cardiovasc. Res. 2004, 63, 593–602. [Google Scholar] [CrossRef] [PubMed]
- Mizutani, K.; Ikeda, K.; Kawai, Y.; Yamori, Y. Extract of wine phenolics improves aortic biomechanical properties in stroke-prone spontaneously hypertensive rats (shrsp). J. Nutr. Sci. Vitaminol. 1999, 45, 95–106. [Google Scholar] [CrossRef] [PubMed]
- Diebolt, M.; Bucher, B.; Andriantsitohaina, R. Wine polyphenols decrease blood pressure, improve no vasodilatation, and induce gene expression. Hypertension 2001, 38, 159–165. [Google Scholar] [CrossRef] [PubMed]
- Edirisinghe, I.; Burton-Freeman, B.; Tissa Kappagoda, C. Mechanism of the endothelium-dependent relaxation evoked by a grape seed extract. Clin. Sci. (Lond.) 2008, 114, 331–337. [Google Scholar] [CrossRef] [PubMed]
- Rubanyi, G.M.; Vanhoutte, P.M. Superoxide anions and hyperoxia inactivate endothelium-derived relaxing factor. Am. J. Physiol. 1986, 250, H822–H827. [Google Scholar] [PubMed]
- Lundberg, J.O.; Weitzberg, E. No generation from nitrite and its role in vascular control. Arterioscler. Thromb. Vasc. Biol. 2005, 25, 915–922. [Google Scholar] [CrossRef] [PubMed]
- Lau, Y.S.; Kwan, C.Y.; Ku, T.C.; Hsieh, W.T.; Wang, H.D.; Nishibe, S.; Dharmani, M.; Mustafa, M.R. Apocynum venetum leaf extract, an antihypertensive herb, inhibits rat aortic contraction induced by angiotensin ii: A nitric oxide and superoxide connection. J. Ethnopharmacol. 2012, 143, 565–571. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lau, Y.S.; Ling, W.C.; Murugan, D.; Kwan, C.Y.; Mustafa, M.R. Endothelium-Dependent Relaxation Effect of Apocynum venetum Leaf Extract via Src/PI3K/Akt Signalling Pathway. Nutrients 2015, 7, 5239-5253. https://doi.org/10.3390/nu7075220
Lau YS, Ling WC, Murugan D, Kwan CY, Mustafa MR. Endothelium-Dependent Relaxation Effect of Apocynum venetum Leaf Extract via Src/PI3K/Akt Signalling Pathway. Nutrients. 2015; 7(7):5239-5253. https://doi.org/10.3390/nu7075220
Chicago/Turabian StyleLau, Yeh Siang, Wei Chih Ling, Dharmani Murugan, Chiu Yin Kwan, and Mohd Rais Mustafa. 2015. "Endothelium-Dependent Relaxation Effect of Apocynum venetum Leaf Extract via Src/PI3K/Akt Signalling Pathway" Nutrients 7, no. 7: 5239-5253. https://doi.org/10.3390/nu7075220