Assessing the Association between Natural Food Folate Intake and Blood Folate Concentrations: A Systematic Review and Bayesian Meta-Analysis of Trials and Observational Studies

 ,
,
Abstract
:1. Introduction
2. Subjects and Methods
2.1. Search Strategy
2.2. Inclusion and Exclusion Criteria
2.3. Selection of Articles
2.4. Data Extraction
2.5. Quality Assessment of Studies
2.6. Data Standardization
RBC folate (nmol/L): Y = 0.7297 × X + 352.219
2.7. Meta-Analysis Study Inclusion/Exclusion
2.8. Statistical Analyses
2.9. Statistical Modeling
3. Results
3.1. Study Characteristics
3.2. Risk of Bias
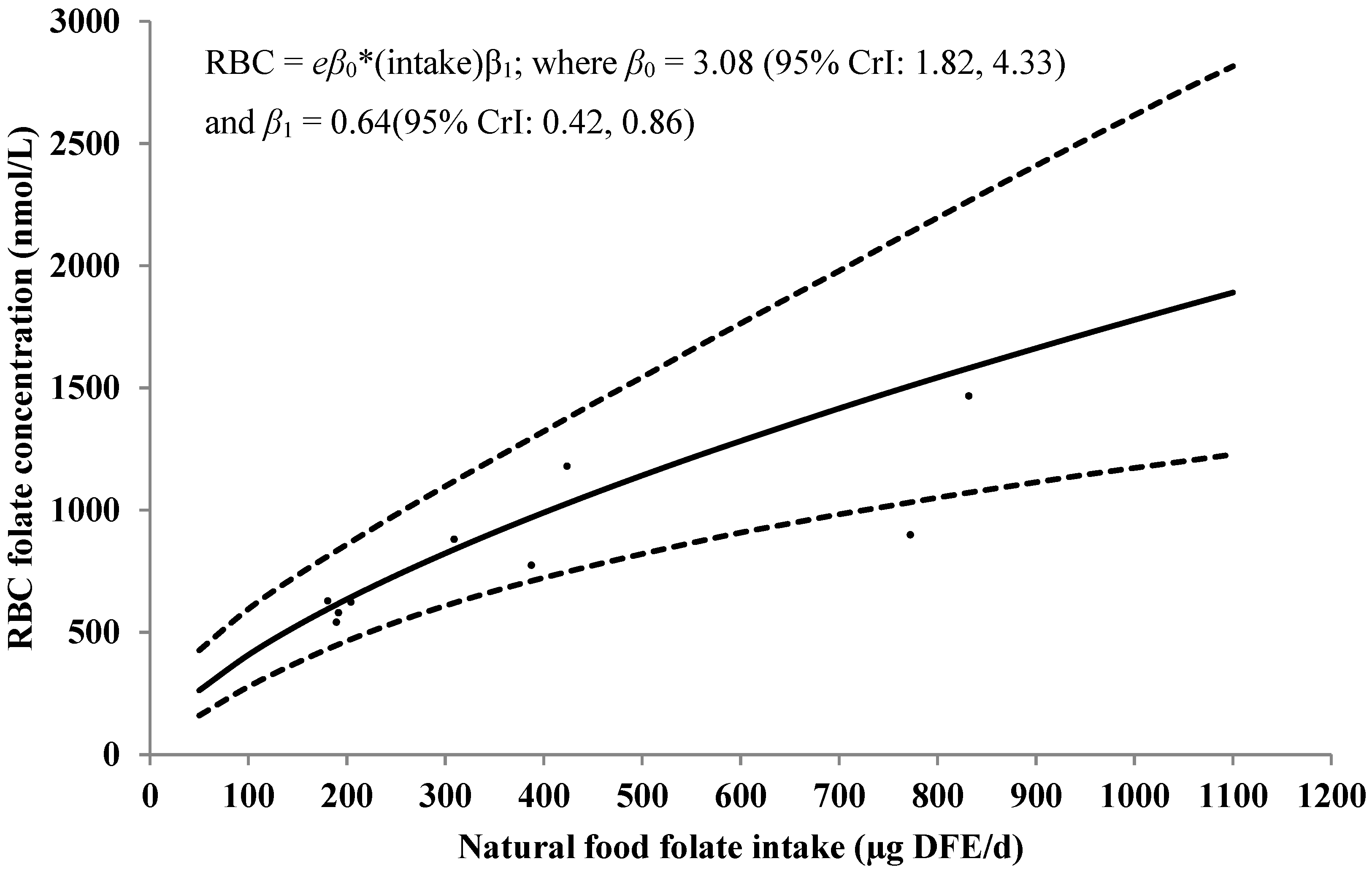
3.3. RBC Folate Concentrations
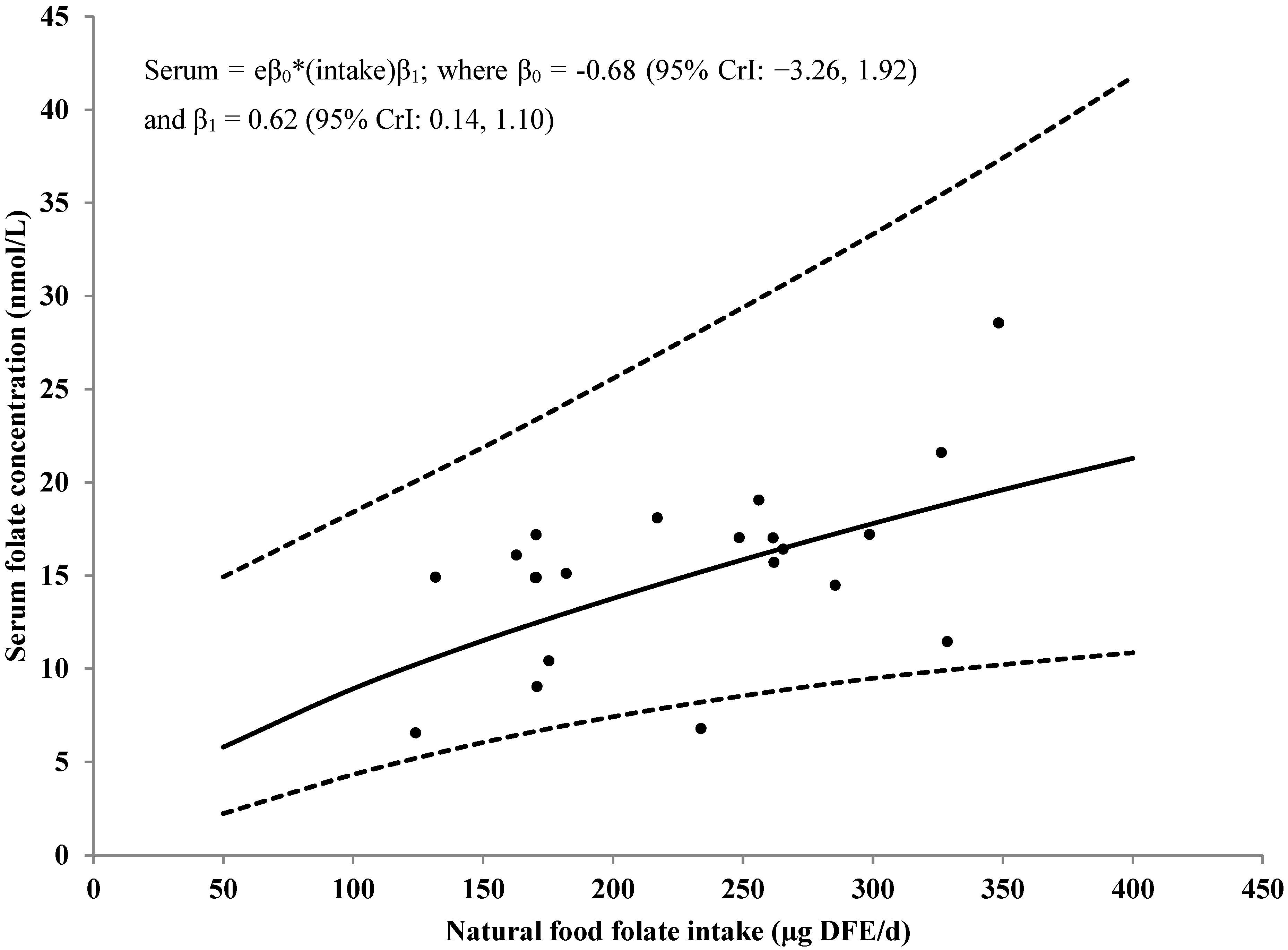
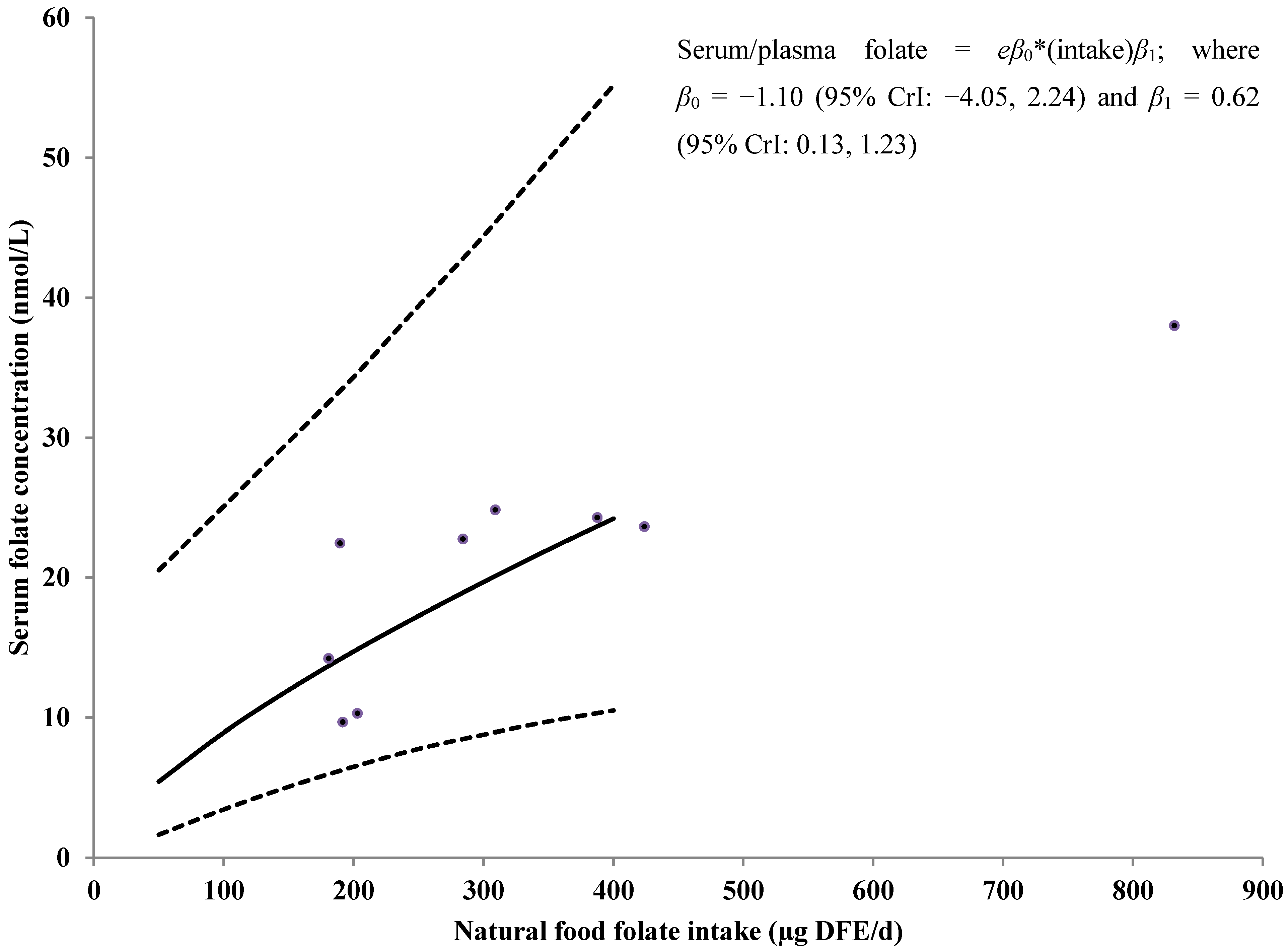
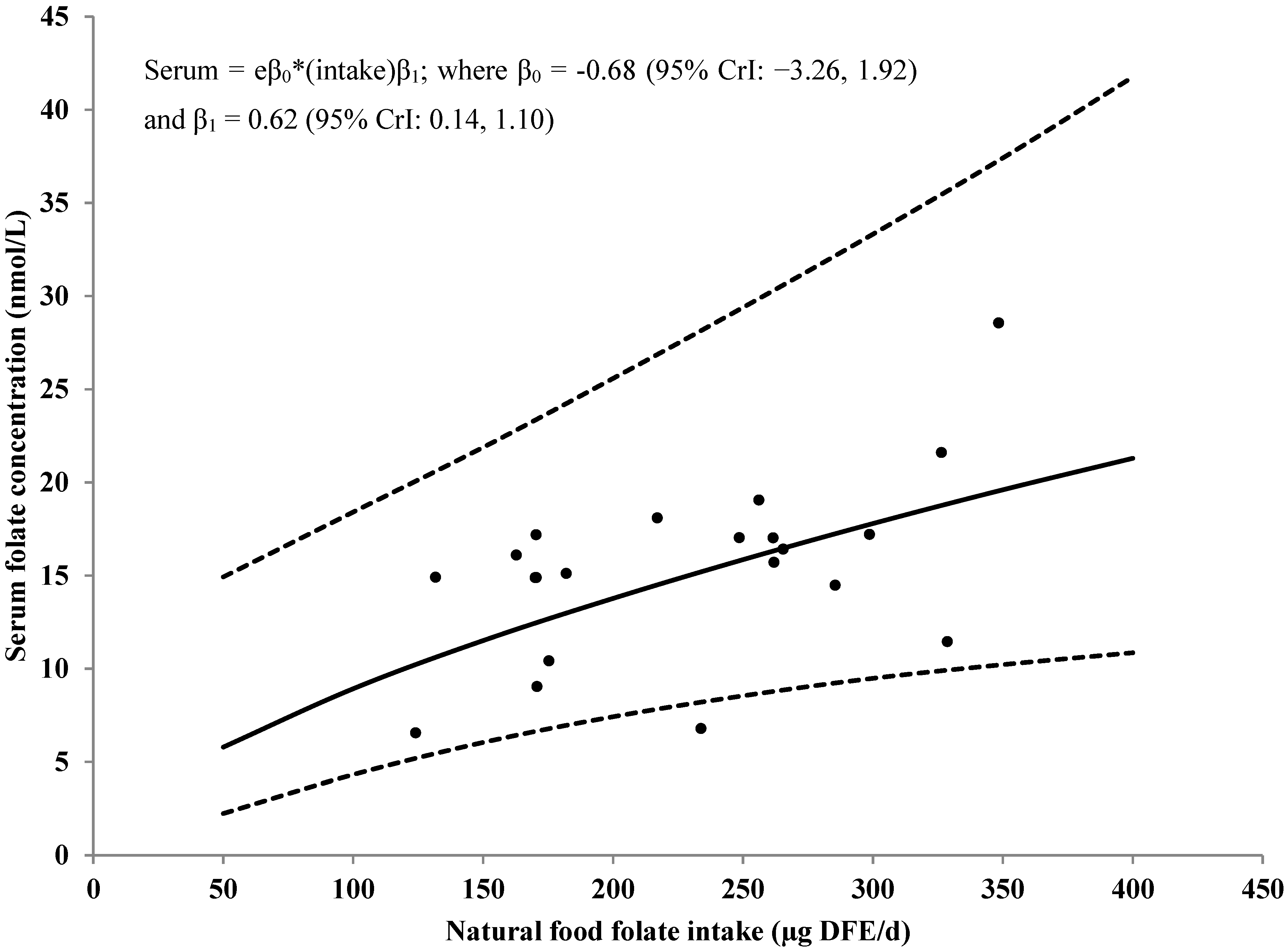
3.4. Serum/Plasma Folate Concentrations
3.5. Sensitivity Analyses

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Natural Food | Median | 95% Credible Interval |
|---|---|---|
| Folate Intake (μg DFE/day) | RBC Folate (nmol/L) | |
| 50 | 260 | (160, 420) |
| 100 | 410 | (280, 590) |
| 150 | 530 | (380, 730) |
| 200 | 640 | (460, 860) |
| 250 | 730 | (540, 980) |
| 300 | 820 | (610, 1100) |
| 350 | 910 | (670, 1210) |
| 400 | 990 | (720, 1330) |
| 700 | 1420 | (980, 1970) |
| 750 | 1480 | (1030, 2080) |
| 800 | 1550 | (1060, 2180) |
| 850 | 1600 | (1080, 2300) |
| 900 | 1670 | (1120, 2390) |
| 950 | 1730 | (1150, 2520) |
| 1000 | 1780 | (1190, 2610) |
| 1050 | 1840 | (1210, 2700) |
| 1100 | 1900 | (1230, 2810) |
4. Discussion
5. Conclusions
Supplementary Files
Supplementary File 1Acknowledgments
Author Contributions
Conflicts of Interest
Disclaimer
References
- National Institutes of Health: Office of Dietary Supplements Folate. Available online: http://ods.od.nih.gov/factsheets/Folate-HealthProfessional/ (accessed on 4 September 2014).
- Institute of Medicine. Dietary Reference Intakes for Thiamin, Riboflavin, Niacin, Vitamin B6, Folate, Vitamin B12, Pantothenic Acid, Biotin, and Choline; National Academy Press: Washington, DC, USA, 1998. [Google Scholar]
- Gibson, R.S. Principles of Nutritional Assessment, 2nd ed.; Oxford University Press: New York, NY, USA, 2005. [Google Scholar]
- World Health Organization. Serum and Red Blood Cell Folate Concentrations for Assessing Folate Status in Populations; World Health Organization: Geneva, Switzerland, 2013. [Google Scholar]
- Daly, L.E.; Kirke, P.N.; Molloy, A.; Weir, D.G.; Scott, J.M. Folate levels and neural tube defects: Implications for prevention. JAMA 1995, 274, 1698–1702. [Google Scholar] [CrossRef] [PubMed]
- Crider, K.S.; Devine, O.; Hao, L.; Dowling, N.F.; Li, S.; Molloy, A.M.; Li, Z.; Zhu, J.-H.; Berry, R.J. Population red blood cell folate concentrations for prevention of neural tube defects: Bayesian model. BMJ 2014, 349, g4554. [Google Scholar] [CrossRef] [PubMed]
- Pfeiffer, C.; Hughes, J.P.; Lacher, D.A.; Bailey, R.L.; Berry, R.J.; Zhang, M.; Yetley, E.A.; Rader, J.I.; Sempos, C.T.; Johnson, C.L. Estimation of trends in serum and RBC folate in the U.S. population from pre- to postfortification using assay-adjusted data from the NHANES 1988–2010. J. Nutr. 2012, 142, 886–893. [Google Scholar] [CrossRef] [PubMed]
- Crider, K.S.; Zhu, J.-H.; Hao, L.; Yang, Q.-H.; Yang, T.P.; Gindler, J.; Maneval, D.R.; Quinlivan, E.P.; Li, Z.; Bailey, L.B.; et al. MTHFR 677→T genotype is associated with folate and homocysteine concentrations in a large, population-based, double-blind trial of folic acid supplementation. Am. J. Clin. Nutr. 2011, 93, 1365–1372. [Google Scholar] [CrossRef]
- Berry, R.J.; Li, Z.; Erickson, J.D.; Li, S.; Moore, C.A.; Wang, H.; Mulinare, J.; Zhao, P.; Wong, L.Y.; Gindler, J.; et al. Prevention of neural-tube defects with folic acid in China. China-U.S. collaborative project for neural tube defect prevention. N. Engl. J. Med. 1999, 341, 1485–1490. [Google Scholar] [CrossRef]
- Centers for Disease Control and Prevention. Recommendations for the use of folic acid to reduce the number of cases of spina bifida and other neural tube defects. MMWR Recomm. Rep. 1992, 41, 1–7. [Google Scholar]
- MRC Vitamin Study Research Group. Prevention of neural tube defects: Results of the Medical Research Council Vitamin Study. Lancet 1991, 338, 131–137. [Google Scholar]
- Centers for Disease Control and Prevention. CDC grand rounds: Additional opportunities to prevent neural tube defects with folic acid fortification. MMWR Morb. Mortal. Wkly. Rep. 2010, 59, 980–984. [Google Scholar]
- Williams, L.J.; Rasmussen, S.A.; Flores, A.; Kirby, R.S.; Edmonds, L.D. Decline in the prevalence of spina bifida and anencephaly by race/ethnicity: 1995–2002. Pediatrics 2005, 116, 580–586. [Google Scholar] [CrossRef] [PubMed]
- Tsang, B.L.; Devine, O.J.; Cordero, A.M.; Marchetta, C.M.; Mulinare, J.; Mersereau, P.; Guo, J.; Qi, Y.P.; Berry, R.J.; Rosentha, J.; et al. Assessing the association between the methylenetetrahydrofolate reductase (MTHFR) 677C>T polymorphism and blood folate concentrations: A systematic review and meta-analysis of trials and observational studies. Am. J. Clin. Nutr. 2015. [Google Scholar] [CrossRef]
- Higgins, J.P.T.; Green, S. (Eds.) Cochrane Handbook for Systematic Reviews of Interventions; The Cochrane Collaboration: Oxford, UK, 2011. Available online: http://www.cochrane-handbook.org (accessed on 2 April 2013).
- Moher, D.; Liberati, A.; Tetzlaff, J.; Altman, D.G.; Group, P. Preferred reporting items for systematic reviews and meta-analyses: The PRISMA statement. J. Clin. Epidemiol. 2009, 62, 1006–1012. [Google Scholar] [CrossRef] [PubMed]
- Viswanathan, M.; Berkman, N.D. Development of the RTI item bank on risk of bias and precision of observational studies. J. Clin. Epidemiol. 2012, 65, 163–178. [Google Scholar] [CrossRef] [PubMed]
- Souverein, O.W.; Dullemeijer, C.; van’t Veer, P.; van der Voet, H. Transformations of summary statistics as input in meta-analysis for linear dose-response models on a logarithmic scale: A methodology developed within EURRECA. BMC Med. Res. Methodol. 2012, 12, 57. [Google Scholar] [CrossRef] [PubMed]
- Pfeiffer, C.M.; Zhang, M.; Lacher, D.A.; Molloy, A.M.; Tamura, T.; Yetley, E.A.; Picciano, M.F.; Johnson, C.L. Comparison of serum and red blood cell folate microbiologic assays for national population surveys. J. Nutr. 2011, 141, 1402–1409. [Google Scholar] [CrossRef] [PubMed]
- Tamura, T. Microbiological assay of folates. In Folic Acid Metabolism in Health and Disease; Picciano, M.F., Stokstad, E.L.R., Eds.; Wiley: New York, NY, USA, 1990; pp. 121–137. [Google Scholar]
- O’Broin, S.; Kelleher, B. Microbiological assay on microtitre plates of folate in serum and red cells. J. Clin. Pathol. 1992, 45, 344–347. [Google Scholar] [CrossRef]
- Molloy, A.M.; Scott, J.M. Microbiological assay for serum, plasma and red cell folate using cryopreserved, microtiter plate method. Methods Enzymol. 1997, 281, 43–53. [Google Scholar] [PubMed]
- Rothman, K.; Greenland, S.; Lash, T. Modern Epidemiology, 3rd ed.; Lippincott Williams & Wilkins: New York, NY, USA, 2012. [Google Scholar]
- Gunter, E.W.; Bowman, B.A.; Caudill, S.P.; Twite, D.B.; Adams, M.J.; Sampson, E.J. Results of an international round robin for serum and whole-blood folate. Clin. Chem. 1996, 42, 1689–1694. [Google Scholar] [PubMed]
- Owen, W.E.; Roberts, W.L. Comparison of five automated serum and whole blood folate assays. Am. J. Clin. Pathol. 2003, 120, 121–126. [Google Scholar] [CrossRef] [PubMed]
- Satija, A.; Yu, E.; Willett, W.C.; Hu, F.B. Understanding nutritional epidemiology and its role in policy. Adv. Nutr. 2015, 6, 5–18. [Google Scholar] [CrossRef] [PubMed]
- Chew, S.C.; Khor, G.L.; Loh, S.P. Association between dietary folate intake and blood status of folate and homocysteine in Malaysian adults. J. Nutr. Sci. Vitaminol. 2011, 57, 150–155. [Google Scholar] [CrossRef] [PubMed]
- Rasmussen, L.B.; Ovesen, L.; Bulow, I.; Knudsen, N.; Laurberg, P.; Perrild, H. Folate intake, lifestyle factors, and homocysteine concentrations in younger and older women. Am. J. Clin. Nutr. 2000, 72, 1156–1163. [Google Scholar] [PubMed]
- Agodi, A.; Barchitta, M.; Quattrocchi, A.; Marchese, A.E.; Boffetta, P. Folate deficiency is not associated with increased mitochondrial genomic instability: Results from dietary intake and lymphocytic mtDNA 4977-bp deletion in healthy young women in Italy. Mutagenesis 2014, 29, 101–106. [Google Scholar] [CrossRef] [PubMed]
- Abratte, C.M.; Wang, W.; Li, R.; Moriarty, D.J.; Caudill, M.A. Folate intake and the MTHFR C677T genotype influence choline status in young Mexican American women. J. Nutr. Biochem. 2008, 19, 158–165. [Google Scholar] [CrossRef] [PubMed]
- Hung, J.; Yang, T.L.; Urrutia, T.F.; Li, R.; Perry, C.A.; Hata, H.; Cogger, E.A.; Moriarty, D.J.; Caudill, M.A. Additional food folate derived exclusively from natural sources improves folate status in young women with the MTHFR 677 CC or TT genotype. J. Nutr. Biochem. 2006, 17, 728–734. [Google Scholar] [CrossRef] [PubMed]
- Perry, C.A.; Renna, S.A.; Khitun, E.; Ortiz, M.; Moriarty, D.J.; Caudill, M.A. Ethnicity and race influence the folate status response to controlled folate intakes in young women. J. Nutr. 2004, 134, 1786–1792. [Google Scholar] [PubMed]
- Shelnutt, K.P.; Kauwell, G.P.; Chapman, C.M.; Gregory, J.F., 3rd; Maneval, D.R.; Browdy, A.A.; Theriaque, D.W.; Bailey, L.B. Folate status response to controlled folate intake is affected by the methylenetetrahydrofolate reductase 677C→T polymorphism in young women. J. Nutr. 2003, 133, 4107–4111. [Google Scholar]
- Wright, A.J.; King, M.J.; Wolfe, C.A.; Powers, H.J.; Finglas, P.M. Comparison of (6 S)-5-methyltetrahydrofolic acid v. folic acid as the reference folate in longer-term human dietary intervention studies assessing the relative bioavailability of natural food folates: Comparative changes in folate status following a 16-week placebo-controlled study in healthy adults. Br. J. Nutr. 2010, 103, 724–729. [Google Scholar] [CrossRef] [PubMed]
- Kwanbunjan, K.; Thepouyporn, A.; Songmuaeng, K.; Nakosiri, W.; Cheeramakara, C.; Chusongsang, Y.; Laisupasin, P.; Tunsakul, S.; Chantaranipapong, Y.; Pooudong, S.; et al. Food behavior and folate status of hill-tribe schoolchildren and women of childbearing age on the northern border of Thailand. Southeast Asian J. Trop. Med. Public Health 2008, 39, 353–361. [Google Scholar]
- Pathak, P.; Saxena, R.; Kapoor, S.K.; Dwivedi, S.N.; Singh, R.; Kapil, U. Status of serum ferritin and folate levels amongst young women in a rural community of Haryana, India. Nepal Med. Coll. J. 2004, 6, 13–16. [Google Scholar] [PubMed]
- Jang, H.B.; Han, Y.H.; Piyathilake, C.J.; Kim, H.; Hyun, T. Intake and blood concentrations of folate and their association with health-related behaviors in Korean college students. Nutr. Res. Pract. 2013, 7, 216–223. [Google Scholar] [CrossRef] [PubMed]
- Han, Y.H.; Yon, M.; Hyun, T.H. Folate intake estimated with an updated database and its association to blood folate and homocysteine in Korean college students. Eur. J. Clin. Nutr. 2005, 59, 246–254. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.A.; Lim, H.S. Dietary folate intake, blood folate status, and urinary folate catabolite excretion in Korean women of childbearing age. J. Nutr. Sci. Vitaminol. 2008, 54, 291–297. [Google Scholar] [CrossRef] [PubMed]
- Khor, G.L.; Duraisamy, G.; Peng Loh, S.; Green, T.J.; Skeaff, C.M. Dietary and blood folate status of Malaysian women of childbearing age. Asia Pac. J. Clin. Nutr. 2006, 15, 341–349. [Google Scholar] [PubMed]
- Dominguez-Salas, P.; Moore, S.E.; Cole, D.; da Costa, K.A.; Cox, S.E.; Dyer, R.A.; Fulford, A.J.; Innis, S.M.; Waterland, R.A.; Zeisel, S.H.; et al. DNA methylation potential: Dietary intake and blood concentrations of one-carbon metabolites and cofactors in rural African women. Am. J. Clin. Nutr. 2013, 97, 1217–1227. [Google Scholar] [CrossRef] [PubMed]
- Yang, Q.; Bailey, L.; Clarke, R.; Flanders, W.D.; Liu, T.; Yesupriya, A.; Khoury, M.J.; Friedman, J.M. Prospective study of methylenetetrahydrofolate reductase (MTHFR) variant C677T and risk of all-cause and cardiovascular disease mortality among 6000 US adults. Am. J. Clin. Nutr. 2012, 95, 1245–1253. [Google Scholar] [CrossRef] [PubMed]
- Costa de Carvalho, M.J.; Guilland, J.C.; Moreau, D.; Boggio, V.; Fuchs, F. Vitamin status of healthy subjects in Burgundy (France). Ann. Nutr. Metab. 1996, 40, 24–51. [Google Scholar]
- Glew, R.H.; Williams, M.; Conn, C.A.; Cadena, S.M.; Crossey, M.; Okolo, S.N.; VanderJagt, D.J. Cardiovascular disease risk factors and diet of Fulani pastoralists of northern Nigeria. Am. J. Clin. Nutr. 2001, 74, 730–736. [Google Scholar] [PubMed]
- Haddad, E.H.; Berk, L.S.; Kettering, J.D.; Hubbard, R.W.; Peters, W.R. Dietary intake and biochemical, hematologic, and immune status of vegans compared with nonvegetarians. Am. J. Clin. Nutr. 1999, 70, 586S–593S. [Google Scholar] [PubMed]
- Liu, J.J.; Prescott, J.; Giovannucci, E.; Hankinson, S.E.; Rosner, B.; de Vivo, I. One-carbon metabolism factors and leukocyte telomere length. Am. J. Clin. Nutr. 2013, 97, 794–799. [Google Scholar] [CrossRef] [PubMed]
- Sofi, F.; Innocenti, G.; Dini, C.; Masi, L.; Battistini, N.C.; Brandi, M.L.; Rotella, C.M.; Gensini, G.F.; Abbate, R.; Surrenti, C.; et al. Low adherence of a clinically healthy Italian population to nutritional recommendations for primary prevention of chronic diseases. Nutr. Metab. Cardiovasc. Dis. 2006, 16, 436–444. [Google Scholar] [CrossRef] [PubMed]
- Vandevijvere, S.; Geelen, A.; Gonzalez-Gross, M.; van’t Veer, P.; Dallongeville, J.; Mouratidou, T.; Dekkers, A.; Bornhorst, C.; Breidenassel, C.; Crispim, S.P.; et al. Evaluation of food and nutrient intake assessment using concentration biomarkers in European adolescents from the Healthy Lifestyle in Europe by Nutrition in Adolescence study. Br. J. Nutr. 2013, 109, 736–747. [Google Scholar] [CrossRef] [PubMed]
- Castetbon, K.; Vernay, M.; Malon, A.; Salanave, B.; Deschamps, V.; Roudier, C.; Oleko, A.; Szego, E.; Hercberg, S. Dietary intake, physical activity and nutritional status in adults: The French nutrition and health survey (ENNS, 2006–2007). Br. J. Nutr. 2009, 102, 733–743. [Google Scholar] [CrossRef] [PubMed]
- Henriquez, P.; Doreste, J.; Diaz-Cremades, J.; Lopez-Blanco, F.; Alvarez-Leon, E.; Serra-Majem, L. Folate status of adults living in the Canary Islands (Spain). Int. J. Vitam. Nutr. Res. 2004, 74, 187–192. [Google Scholar] [CrossRef] [PubMed]
- Hiraoka, M. Nutritional status of vitamin A, E, C, B1, B2, B6, nicotinic acid, B12, folate, and beta-carotene in young women. J. Nutr. Sci. Vitaminol. 2001, 47, 20–27. [Google Scholar] [CrossRef] [PubMed]
- Hiraoka, M. Folate intake, serum folate, serum total homocysteine levels and methylenetetrahydrofolate reductase C677T polymorphism in young Japanese women. J. Nutr. Sci. Vitaminol. 2004, 50, 238–245. [Google Scholar] [CrossRef] [PubMed]
- Al Khatib, L.; Obeid, O.; Sibai, A.M.; Batal, M.; Adra, N. Folate deficiency is associated with nutritional anaemia in Lebanese women of childbearing age. Public Health Nutr. 2006, 9, 921–927. [Google Scholar]
- Planells, E.; Sanchez, C.; Montellano, M.A.; Mataix, J.; Llopis, J. Vitamins B6 and B12 and folate status in an adult Mediterranean population. Eur. J. Clin. Nutr. 2003, 57, 777–785. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, H.; Ishida, S.; Konno, Y.; Matsumoto, M.; Nomachi, S.; Masaki, K.; Okayama, H.; Nagai, Y. Impact of dietary folate intake on depressive symptoms in young women of reproductive age. J. Midwifery Womens Health 2012, 57, 43–48. [Google Scholar] [CrossRef] [PubMed]
- Kondo, A.; Tsushima Rehabilitation Hospital, Minami-Shinkai, Tsushima, Japan. Personal communication, 2012.
- Lee, Y.; Krawinkel, M. The nutritional status of iron, folate, and vitamin B-12 of Buddhist vegetarians. Asia Pac. J. Clin. Nutr. 2011, 20, 42–49. [Google Scholar] [PubMed]
- Lunn, D.; Spiegelhalter, D.; Thomas, A.; Best, N. The BUGS project: Evolution, critique and future directions (with discussion). Stat. Med. 2009, 28, 3049–3082. [Google Scholar] [CrossRef] [PubMed]
- Gelman, A.; Carlin, J.B.; Stern, H.S.; Dunson, D.B.; Vehtari, A.; Rubin, D.B. Bayesian Data Analysis; CDC Press: New York, NY, USA, 2013. [Google Scholar]
- Fazili, Z.; Pfeiffer, C.M.; Zhang, M. Comparison of serum folate species analyzed by LC-MS/MS with total folate measured by microbiologic assay and Bio-Rad radioassay. Clin. Chem. 2007, 53, 781–784. [Google Scholar] [CrossRef] [PubMed]
- Fazili, Z.; Pfeiffer, C.M.; Zhang, M.; Jain, R.B.; Koontz, D. Influence of 5,10-methylenetetrahydrofolate reductase polymorphism on whole-blood folate concentrations measured by LC-MS/MS, microbiologic assay, and Bio-Rad radioassay. Clin. Chem. 2008, 54, 197–201. [Google Scholar] [CrossRef] [PubMed]
- Molloy, A.M.; Mills, J.L.; Kirke, P.N.; Whitehead, A.S.; Weir, D.G.; Scott, J.M. Whole-blood folate values in subjects with different methylenetetrahydrofolate reductase genotypes: Differences between the radioassay and microbiological assays. Clin. Chem. 1998, 44, 186–188. [Google Scholar] [PubMed]
- Willett, W. Nutritional Epidemiology, 2nd ed.; Oxford University Press: New York, NY, USA, 1998; Volume 30. [Google Scholar]
- Food Fortification Initiative Country Profiles. Available online: http://www.ffinetwork.org/ (accessed on 27 June 2013).
- Brouwer, I.A.; van Dusseldorp, M.; West, C.E.; Meyboom, S.; Thomas, C.M.; Duran, M.; van het Hof, K.H.; Eskes, T.K.; Hautvast, J.G.; Steegers-Theunissen, R.P. Dietary folate from vegetables and citrus fruit decreases plasma homocysteine concentrations in humans in a dietary controlled trial. J. Nutr. 1999, 129, 1135–1139. [Google Scholar] [PubMed]
- Silaste, M.L.; Rantala, M.; Alfthan, G.; Aro, A.; Kesaniemi, A. Plasma homocysteine concentration is decreased by dietary intervention. Br. J. Nutr. 2003, 89, 295–301. [Google Scholar] [CrossRef] [PubMed]
- Mennen, L.I.; de Courcy, G.P.; Guilland, J.C.; Ducros, V.; Bertrais, S.; Nicolas, J.P.; Maurel, M.; Zarebska, M.; Favier, A.; Franchisseur, C.; et al. Homocysteine, cardiovascular disease risk factors, and habitual diet in the French Supplementation with Antioxidant Vitamins and Minerals Study. Am. J. Clin. Nutr. 2002, 76, 1279–1289. [Google Scholar] [PubMed]
- Nagata, C.; Shimizu, H.; Takami, R.; Hayashi, M.; Takeda, N.; Yasuda, K. Soy product intake is inversely associated with serum homocysteine level in premenopausal Japanese women. J. Nutr. 2003, 133, 797–800. [Google Scholar] [PubMed]
- Taguchi, T.; Mori, H.; Hamada, A.; Yamori, Y.; Mori, M. Serum folate, total homocysteine levels and methylenetetrahydrofolate reductase 677C>T polymorphism in young healthy female Japanese. Asia Pac. J. Clin. Nutr. 2012, 21, 291–295. [Google Scholar] [PubMed]
- Berti, C.; Fekete, K.; Dullemeijer, C.; Trovato, M.; Souverein, O.W.; Cavelaars, A.; Dhonukshe-Rutten, R.; Massari, M.; Decsi, T.; van’t Veer, P.; et al. Folate intake and markers of folate status in women of reproductive age, pregnant and lactating women: A meta-analysis. J. Nutr. Metab. 2012, 2012. [Google Scholar] [CrossRef]
- Duffy, M.E.; Hoey, L.; Hughes, C.F.; Strain, J.J.; Rankin, A.; Souverein, O.W.; Dullemeijer, C.; Collings, R.; Hooper, L.; McNulty, H. Biomarker responses to folic acid intervention in healthy adults: A meta-analysis of randomized controlled trials. Am. J. Clin. Nutr. 2014, 99, 96–106. [Google Scholar] [CrossRef] [PubMed]
- Wolff, T.; Witkop, C.T.; Miller, T.; Syed, S.B. Folic acid supplementation for the prevention of neural tube defects: An update of the evidence for the U.S. Preventive Services Task Force. Ann. Intern. Med. 2009, 150, 632–639. [Google Scholar] [CrossRef] [PubMed]
- Bentley, T.G.K.; Willett, W.C.; Weinstein, M.C.; Kuntz, K.M. Population-level changes in folate intake by age, gender, and race-ethnicity after folic acid fortification. Am. J. Public Health 2006, 96, 2040–2047. [Google Scholar] [CrossRef] [PubMed]
- Monsivais, P.; Drewnowski, A. The rising cost of low-energy-density foods. J. Am. Diet. Assoc. 2007, 107, 2071–2076. [Google Scholar] [CrossRef] [PubMed]
- Finzer, L.E.; Ajay, V.S.; Ali, M.K.; Shivashankar, R.; Goenka, S.; Sharma, P.; Pillai, D.S.; Khandelwal, S.; Tandon, N.; Reddy, K.S.; et al. Fruit and vegetable purchasing patterns and preferences in South Delhi. Ecol. Food Nutr. 2013, 52, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Krajcovicová-Kudlácková, M.; Valachovicová, M.; Blazícek, P. Seasonal folate serum concentrations at different nutrition. Cent. Eur. J. Public Health 2013, 21, 36–38. [Google Scholar] [PubMed]
- Finer, L.B.; Zolna, M.R. Unintended pregnancy in the United States: Incidence and disparities, 2006. Contraception 2011, 84, 478–485. [Google Scholar] [CrossRef] [PubMed]
- Tinker, S.C.; Cogswell, M.E.; Hamner, H.C.; Berry, R.J. Usual folic acid intakes: A modelling exercise assessing changes in the amount of folic acid in foods and supplements, National Health and Nutrition Examination Survey, 2003–2008. Public Health Nutr. 2012, 15, 1216–1227. [Google Scholar] [CrossRef] [PubMed]
- Molloy, A.M.; Daly, S.; Mills, J.L.; Kirke, P.N.; Whitehead, A.S.; Ramsbottom, D.; Conley, M.R.; Weir, D.G.; Scott, J.M. Thermolabile variant of 5,10-methylenetetrahydrofolate reductase associated with low red-cell folates: Implications for folate intake recommendations. Lancet 1997, 349, 1591–1593. [Google Scholar] [CrossRef] [PubMed]
- Park, J.Y.; Vollset, S.E.; Melse-Boonstra, A.; Chajes, V.; Ueland, P.M.; Slimani, N. Dietary intake and biological measurement of folate: A qualitative review of validation studies. Mol. Nutr. Food Res. 2013, 57, 562–581. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Marchetta, C.M.; Devine, O.J.; Crider, K.S.; Tsang, B.L.; Cordero, A.M.; Qi, Y.P.; Guo, J.; Berry, R.J.; Rosenthal, J.; Mulinare, J.; et al. Assessing the Association between Natural Food Folate Intake and Blood Folate Concentrations: A Systematic Review and Bayesian Meta-Analysis of Trials and Observational Studies. Nutrients 2015, 7, 2663-2686. https://doi.org/10.3390/nu7042663
Marchetta CM, Devine OJ, Crider KS, Tsang BL, Cordero AM, Qi YP, Guo J, Berry RJ, Rosenthal J, Mulinare J, et al. Assessing the Association between Natural Food Folate Intake and Blood Folate Concentrations: A Systematic Review and Bayesian Meta-Analysis of Trials and Observational Studies. Nutrients. 2015; 7(4):2663-2686. https://doi.org/10.3390/nu7042663
Chicago/Turabian StyleMarchetta, Claire M., Owen J. Devine, Krista S. Crider, Becky L. Tsang, Amy M. Cordero, Yan Ping Qi, Jing Guo, Robert J. Berry, Jorge Rosenthal, Joseph Mulinare, and et al. 2015. "Assessing the Association between Natural Food Folate Intake and Blood Folate Concentrations: A Systematic Review and Bayesian Meta-Analysis of Trials and Observational Studies" Nutrients 7, no. 4: 2663-2686. https://doi.org/10.3390/nu7042663
APA StyleMarchetta, C. M., Devine, O. J., Crider, K. S., Tsang, B. L., Cordero, A. M., Qi, Y. P., Guo, J., Berry, R. J., Rosenthal, J., Mulinare, J., Mersereau, P., & Hamner, H. C. (2015). Assessing the Association between Natural Food Folate Intake and Blood Folate Concentrations: A Systematic Review and Bayesian Meta-Analysis of Trials and Observational Studies. Nutrients, 7(4), 2663-2686. https://doi.org/10.3390/nu7042663