The Positive Effects of Yerba Maté (Ilex paraguariensis) in Obesity


Abstract
:1. Introduction
2. Background on Obesity and Its Mechanisms
3. The Effects of Yerba Maté in Obesity

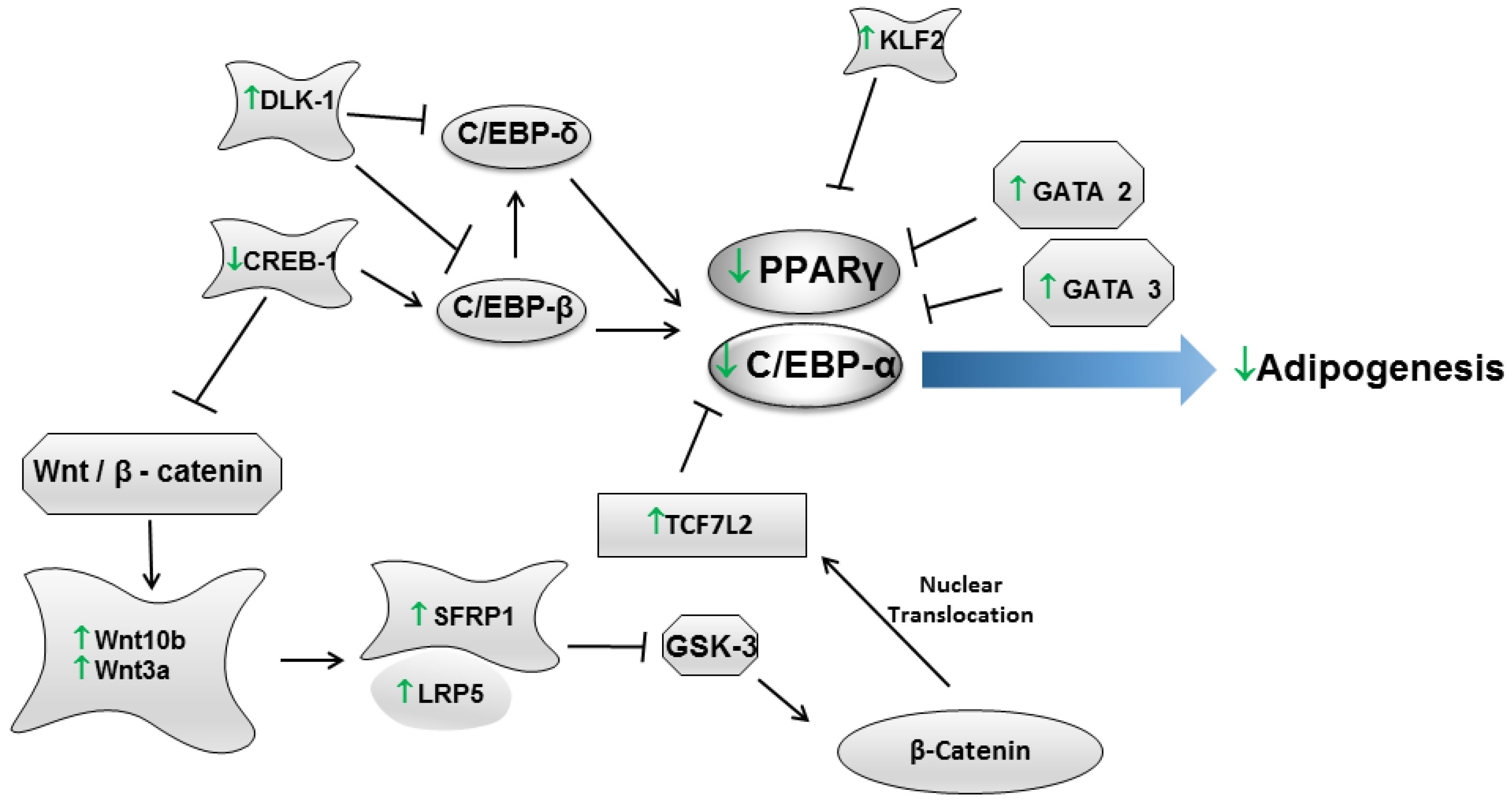
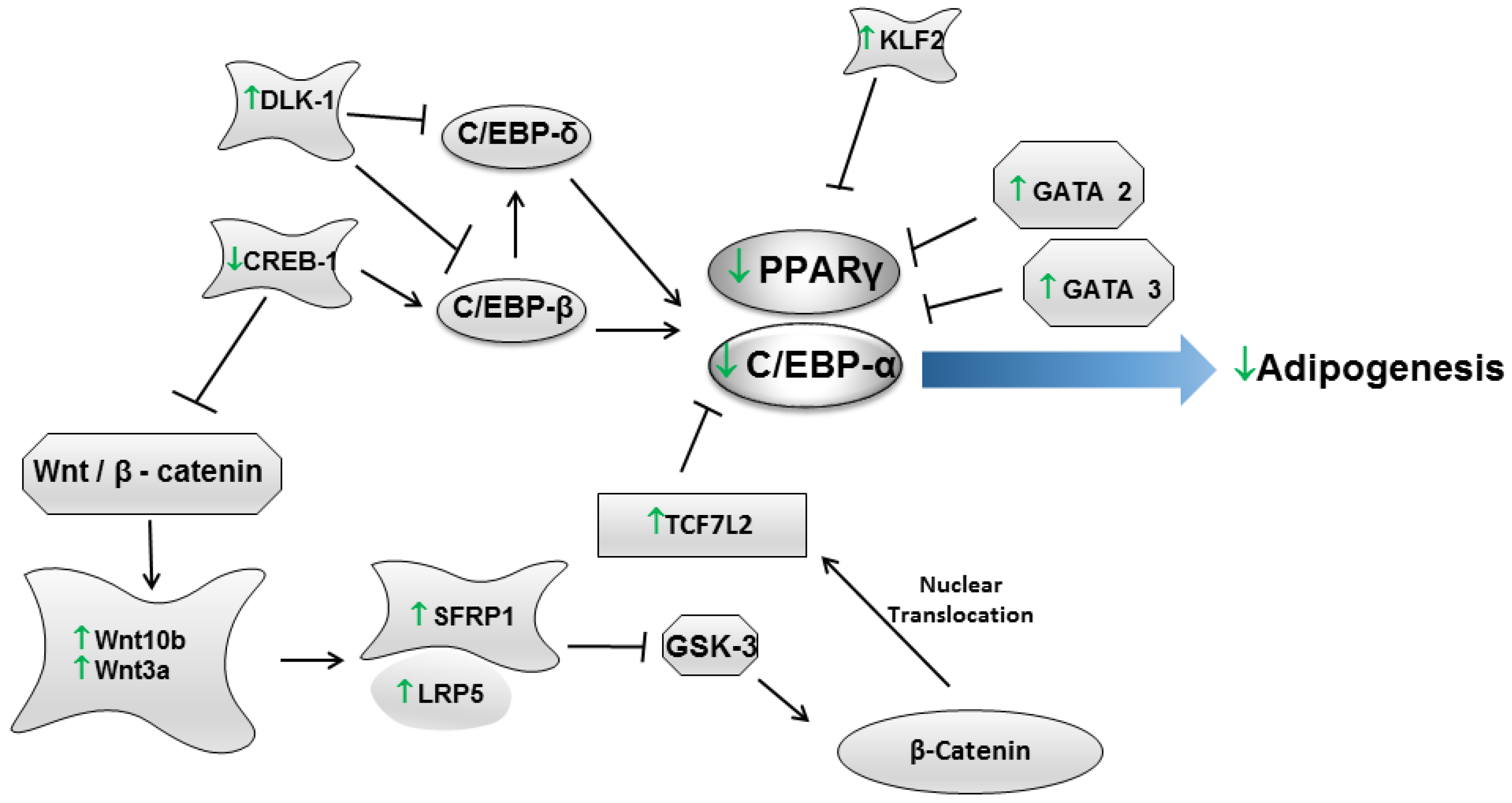{kind=link}
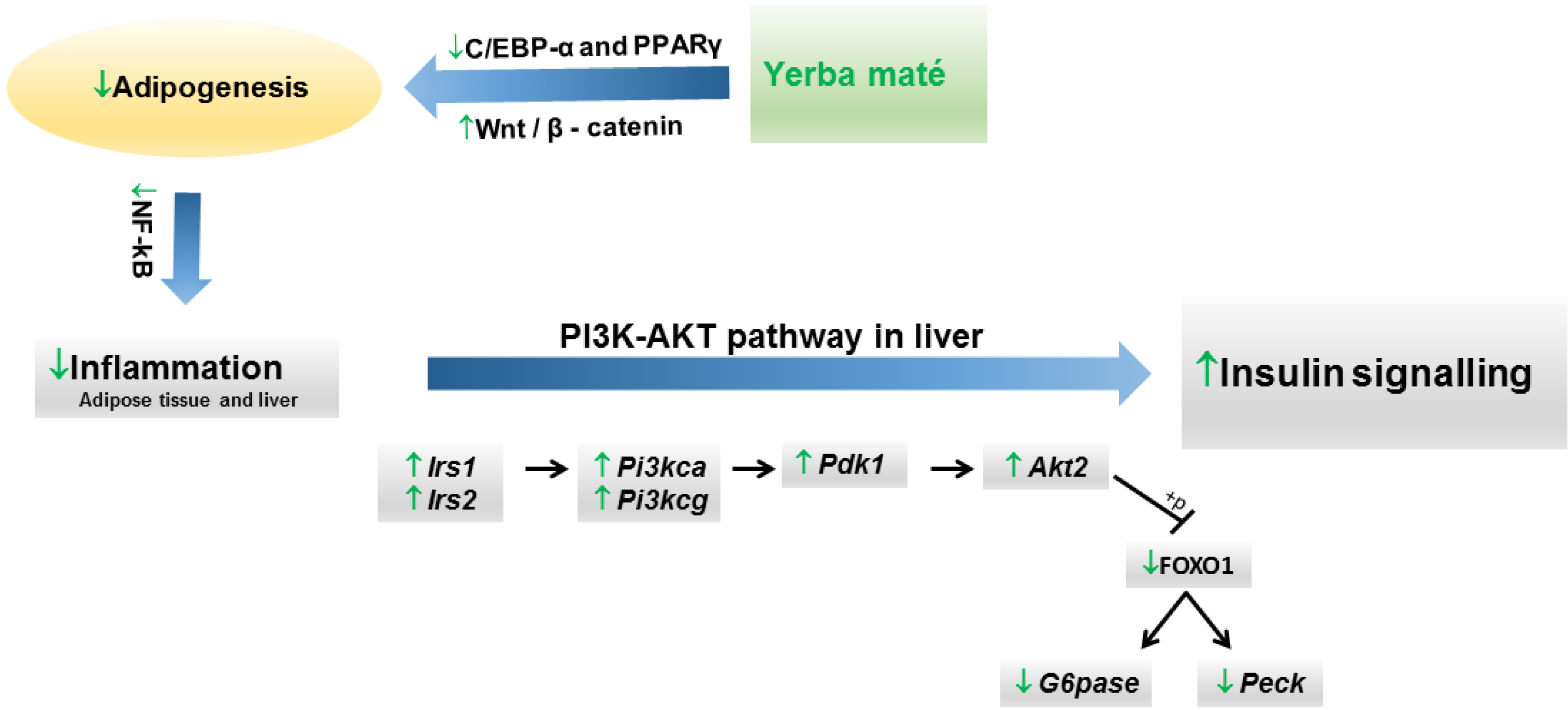{kind=link}
| Study | Type of Study | Population | Test Compounds (Daily Dosage) | Duration of Intake | Main Outcomes |
|---|---|---|---|---|---|
| Andersen and Fogh, 2001 [66] | Double-blind placebo-controlled parallel trial | Forty-seven healthy overweight (body mass index (BMI) range of 25.8 ± 30.4 kg/m2) volunteers. | Three tablets of YGD (112 mg yerba maté, 95 mg guarana and 36 mg damiana extract) before each main meal. | 10 days and 45 days and weight maintenance over 12 months | YGD significantly increased gastric emptying time of 58 ± 15 min compared to 38 ± 7.6 min after placebo; significantly increased body weight reductions over 10 days (0.8 ± 0.05 kg after YGD compared with 0.3 ± 0.03 kg after placebo) and over 45 days (5.1 ± 0.5 kg after YGD compared to 0.3 ± 0.08 kg after placebo); treatment with YGD resulted in weight maintenance (73 kg at the beginning and 72.5 kg at the end of 12 months). |
| De Morais et al., 2009 [68] | Single-blind controlled trial | One hundred and two volunteers (n = 36 male and 66 female; mean age = 48.4 ± 1.35 years). Divided into 3 groups: normolipidemic (n = 15), dyslipidemic (n = 57) and hypercholesterolemic subjects on long-term statin therapy (n = 30). | 330 mL, 3-times/day of green or roasted yerba maté infusions. | 40 days | Normolipidemic subject treated with yerba maté exhibited a significative reduction of 8.7% on LDL-cholesterol. Dyslipidemic individuals lowered LDL-cholesterol levels up to 8.6% and non-HDL cholesterol up to 6.5%. The apolipoprotein B level was reduced by 6.0%, and HDL-cholesterol was significantly increased by 4.4%. The yerba maté consumption by hypercholesterolemic individuals on statin therapy promoted an additional 13.1% reduction in LDL-C and increased HDL cholesterol by 6.2%. |
| Harrold et al., 2013 [67] | Double blind, placebo-controlled, crossover | Fifty-eight healthy women, aged 18–65, with a BMI between 18.5 and 29.9 kg/m2. | Three tablets of YGD (112 mg yerba maté, 95 mg guarana and 36 mg damiana extract) and inulin-based soluble fermentable fiber (SSF; 5 g in 100 mL water), 3 tablets of YGD and water (100 mL), SFF and placebo (3 tablets) or water and placebo 15 min before lunch. | 1 day | YGD and SFF significantly reduced food and energy intake (59.5 g, 16.3%; 112.4 kcal, 17.3%; and 31.9 g, 9.1%; 80 kcal, 11.7%, respectively) compared with conditions where they were absent. The lowest intake (gram and kcal) was observed in the YGD + SFF combination. In summary, YGD causes a robust short-term effect on caloric intake, an effect augmented by SFF. |
| Pang et al., 2008 [58] | Experimental | Sprague-Dawley rats fed with high-fat diet (3 groups, n = 8 each). | High-fat diet (HFD) supplemented with maté (ad libitum). | 8 weeks | Yerba maté extract significantly reduced the final body weight. It reversed the HFD-induced downregulation of the adipose tissue genes implicated in adipogenesis or thermogenesis. Significant decreases in the phospho-AMP-activated protein kinase (AMPK)/AMPK protein ratio were also observed. |
| Arçari et al., 2009 [49] | Experimental | Swiss mice fed with high-fat diet (3 groups, n = 10 each). | 1 g/kg of roasted yerba maté extract by oral route (once per day). | 8 weeks | Yerba maté significantly inhibited the inflammation in adipose tissue induced by obesity, modulating several pro- and anti-inflammatory genes and reducing macrophage infiltration. |
| Martins et al., 2010 [63] | Experimental | Swiss mice fed with high-fat diet (4 groups, n = 10 each). | 1 and 2 g/kg of roasted yerba maté extract by oral route (once per day). | 8 weeks | Yerba maté extract significantly reduced the final body weight. A reduction of total serum cholesterol and LDL-cholesterol levels was observed. Serum triglycerides were also significantly reduced. Yerba maté significantly reduced lipid accumulation in the liver (decreased by ~30%). |
| Arçari et al., 2011 [45] | Experimental | Swiss mice fed with high-fat diet (3 groups, n = 10 each). | 1 g/kg of roasted yerba maté extract by oral route (once per day). | 8 weeks | Yerba maté significantly improved insulin resistance by restoring hepatic and muscle IRS-1 and AKT phosphorylation and by controlling adipose tissue inflammation associated with obesity. |
| Hussein et al., 2011 [43] | Experimental | Male Tsumura Suzuki obese diabetic (TSOD) mice (3 groups, n = 5 each). | 100 mg/kg of an aqueous extract of yerba maté by oral route (once per day). | 7 weeks | Significantly ameliorated metabolic syndrome by improving peripheral insulin sensitivity and cellular glucose uptake and by modulating the level of circulating lipid metabolites and adiponectin. |
| Hussein et al., 2011 [61] | Experimental | DdY mice fed with high-fat diet. | Aqueous extract of maté by oral route (once per day). | 3 weeks | Administration of yerba maté induced significant increases in GLP-1 levels and leptin levels, generating anorexic effects by direct induction of satiety. |
| Kang et al., 2012 [44] | Experimental | C57BL/6J mice fed with high-fat diet (5 groups, n = 5 each). | 0.5, 1 or 2 g/kg of aqueous extract of yerba maté by oral route (once per day). | 4 weeks | Yerba maté consumption significantly reduced the body weight, adiposity, adipocyte size and leptin release by adipose tissue. Maté-treated mice also presented a significant reduction in serum levels of triglycerides and cholesterol. |
| Pimentel et al., 2012 [46] | Experimental | Wistar rats fed with high-fat diet (3 groups, n = 7 each). | From 100 to 200 mg/kg of yerba maté extract by oral route (once per day). | 8 weeks | Yerba maté significantly inhibited hypothalamic inflammation induced by obesity trough reducing the phosphorylation of hypothalamic IKK and NFκBp65 expression and significantly increasing the protein levels of IκBα and adiponectin receptor-1. Inflammation associated with obesity in liver and muscle was also significantly controlled by yerba maté. |
| Arçari et al., 2013 [57] | Experimental | Swiss mice fed with high-fat diet (3 groups, n = 10 each). | 1 g/kg of roasted yerba maté extract by oral route (once per day). | 8 weeks | Significantly improved insulin resistance by restoring hepatic FOXO1 nuclear translocation and upregulating gene expression of Akt2, Irs1, Irs2, Pi3kca, Pi3kcg and Pdk1. |
| Arçari et al., 2013 [59] | Experimental | Swiss mice fed with high-fat diet (3 groups, n = 10 each). | 1 g/kg of roasted yerba maté extract by oral route (once per day). | 8 weeks | Yerba maté extract significantly reduced the final body weight. It downregulated the expression of genes that regulate adipogenesis and upregulated the expression of genes related to the inhibition of adipogenesis. |
| Borges et al., 2013 [48] | Experimental | Wistar rats fed a with high-fat diet (4 groups, n = 8–12 each). | 1 g/kg of roasted yerba maté extract by oral route (once per day). | 4 weeks | The consumption of yerba maté promoted weight loss, attenuated the detrimental effects of HFD on adiposity and insulin sensitivity and decreased the blood levels of inflammatory biomarkers. Concerning peritoneal macrophages, maté decreased the mRNA production of Il-6, but did not influence the production of Il-1β, Tnf-α and nitric oxide. |
| Carmo et al., 2013 [47] | Experimental | Wistar rats fed a with high-fat diet (4 groups, n = 8–12 each). | 1 g/kg of roasted yerba maté extract by oral route (once per day) | 4 weeks | Significantly reduced body weight, body adiposity and cholesterol levels. Maté consumption reduced IL-1α, IL-6 and TNF-α production by bone marrow cells. |
| Gao et al., 2013 [69] | Experimental | Sprague-Dawley rats fed with high-fat diet (5 groups, n = 12 each). | 1 2 and 4% yerba maté extract. The animals had free access to bottles containing the prepared infusion as the only available liquid source. | 4 weeks | Yerba maté may regulate blood lipid and endothelial function in hyperlipidemia rats. The putative mechanism may include a reduction of endothelin and thromboxane A2 levels and an increase in nitric oxide and 6-keto-PGF1α levels in the blood, downregulating the expression of ICAM-1 (Intercellular Adhesion Molecule 1) protein and upregulating the expression of LDLR (LDL receptor) and SR-B1 (scavenger receptor class B member 1) genes, inhibiting the occurrence of atherosclerosis. |
| Lima et al., 2014 [56] | Experimental | Wistar rats primed by early weaning. | 1 g/kg of yerba maté extract by oral route (once per day). | 4 weeks | Yerba maté consumption significantly reduces body weight, adiposity and triglycerides levels in the blood. |
| Study | Type of Cell (Origin) | Assay Employed | Tested Compounds (Concentration) | Main Outcomes |
|---|---|---|---|---|
| Martins et al., 2010 [63] | None * | Determination of lipase activity inhibitory action against porcine and human lipases and the influence of several olive oil emulsifying reagents. | 0.5–5.0 mg/mL of roasted yerba maté. | Yerba maté significantly inhibited the enzyme activities in a dose-dependent way, and its inhibitory activity against both lipases reached a maximum at 3.0 mg/mL, corresponding to 9 mg of tea/g substrate. Kinetic results indicated that yerba maté competitively inhibited pancreatic lipase activity in a concentration-dependent manner with a half-maximal inhibitory concentration value of 1.5 mg yerba maté/mL (or 4.5 mg of yerba maté/g of substrate), whereas a drastic decrease in lipolytic activity (>80% that of the control) was observed in the presence of 3.0 mg yerba maté/mL. |
| Gosmann et al., 2012 [60] | 3T3-L1 (Mus musculus) | Determination of phenolic compounds (Folin-Ciocalteu method), Oil Red O staining and gene expression. | Extracts of both fresh and dried maté leaves subjected to chromatography in order to obtain the saponin (20% yield) and the polyphenol extracts (40% yield). | Among the yerba maté samples, the polyphenol extract of fresh leaves exhibited a higher content of total phenols, followed by the polyphenol extract and the ethanol extract of dried leaves. Saponin extracts of both fresh and dried leaves exhibited lower contents of phenolic compounds. Regarding adipogenesis, the highest anti-adipogenic effect was detected in the polyphenol extract of dried leaves at 50 μg/mL, followed by the saponin extract of fresh leaves at 100 μg/mL and by the polyphenol extract of fresh leaves at 500 μg/mL. All assayed samples restrained the expression of the PPARγ2, Lep, TNF-α and C/EBPα genes. |
| Arcari et al., 2013 [59] | 3T3-L1 (Mus musculus) | Oil Red O staining and gene expression (Mouse Adipogenesis RT2 Profiler™ PCR Array). | 50, 250 or 500 µg/mL of roasted yerba maté, chlorogenic acid, quercetin, and rutin (Sigma-Aldrich). | Yerba maté extract and chlorogenic acid inhibited adipogenesis at a concentration of 50 µg/mL. Quercetin and rutin had inhibitory activity at the highest concentration. The PCR array revealed that yerba maté modulated the expression of 14 genes (belonging to PPARγ and WNT signaling pathways) that are associated with adipogenesis. In addition, the bioactive compounds also modulated the expression of adipogenesis-associated genes. However, fewer genes were regulated by chlorogenic acid, quercetin and rutin than by yerba maté. The authors claim that the synergism between these compounds might be responsible for the results observed. |
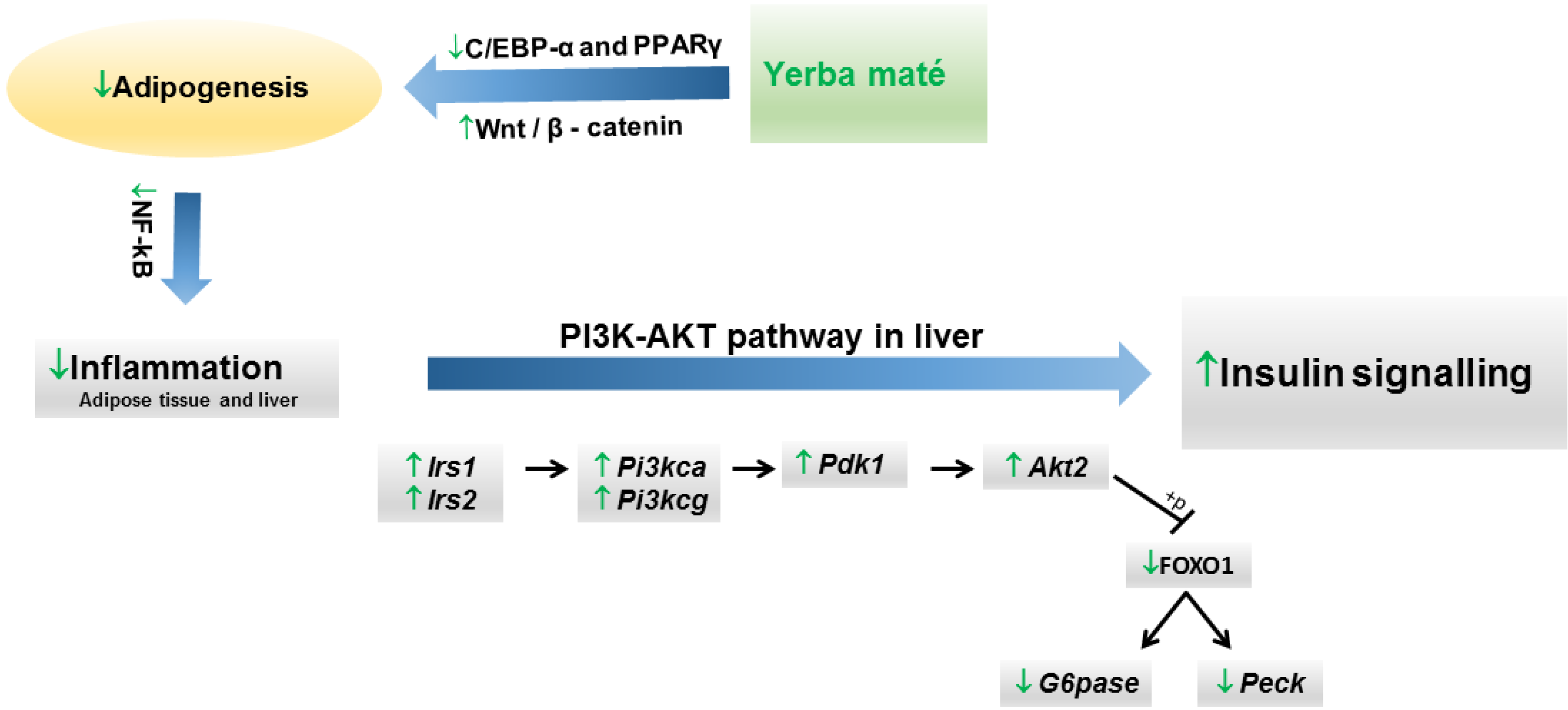
4. Conclusions
Author Contributions
Conflicts of Interest
References
- Berg, A.H.; Scherer, P.E. Adipose tissue, inflammation, and cardiovascular disease. Circ. Res. 2005, 96, 939–949. [Google Scholar] [CrossRef] [PubMed]
- Haslam, D.W.; James, W.P. Obesity. Lancet 2005, 366, 1197–1209. [Google Scholar] [CrossRef] [PubMed]
- Vaisse, C.; Halaas, J.L.; Horvath, C.M.; Darnell, J.E., Jr.; Stoffel, M.; Friedman, J.M. Leptin activation of Stat3 in the hypothalamus of wild-type and ob/ob mice but not db/db mice. Nat. Genet. 1996, 14, 95–97. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Proenca, R.; Maffei, M.; Barone, M.; Leopold, L.; Friedman, J.M. Positional cloning of the mouse obese gene and its human homologue. Nature 1994, 372, 425–432. [Google Scholar] [CrossRef] [PubMed]
- Loffreda, S.; Yang, S.Q.; Lin, H.Z.; Karp, C.L.; Brengman, M.L.; Wang, D.J.; Klein, A.S.; Bulkley, G.B.; Bao, C.; Noble, P.W.; et al. Leptin regulates proinflammatory immune responses. FASEB J. 1998, 12, 57–65. [Google Scholar] [PubMed]
- Hotamisligil, G.S.; Arner, P.; Caro, J.F.; Atkinson, R.L.; Spiegelman, B.M. Increased adipose tissue expression of tumor necrosis factor-alpha in human obesity and insulin resistance. J. Clin. Investig. 1995, 95, 2409–2415. [Google Scholar] [CrossRef] [PubMed]
- Weisberg, S.P.; McCann, D.; Desai, M.; Rosenbaum, M.; Leibel, R.L.; Ferrante, A.W., Jr. Obesity is associated with macrophage accumulation in adipose tissue. J. Clin. Investig. 2003, 112, 1796–1808. [Google Scholar] [CrossRef] [PubMed]
- Lumeng, C.N.; DelProposto, J.B.; Westcott, D.J.; Saltiel, A.R. Phenotypic switching of adipose tissue macrophages with obesity is generated by spatiotemporal differences in macrophage subtypes. Diabetes 2008, 57, 3239–3246. [Google Scholar] [CrossRef] [PubMed]
- Zeyda, M.; Farmer, D.; Todoric, J.; Aszmann, O.; Speiser, M.; Gyori, G.; Zlabinger, G.J.; Stulnig, T.M. Human adipose tissue macrophages are of an anti-inflammatory phenotype but capable of excessive pro-inflammatory mediator production. Int. J. Obesity 2007, 31, 1420–1428. [Google Scholar] [CrossRef]
- Gordon, S. Alternative activation of macrophages. Nat. Rev. Immunol. 2003, 3, 23–35. [Google Scholar] [CrossRef] [PubMed]
- Kosteli, A.; Sugaru, E.; Haemmerle, G.; Martin, J.F.; Lei, J.; Zechner, R.; Ferrante, A.W., Jr. Weight loss and lipolysis promote a dynamic immune response in murine adipose tissue. J. Clin. Investig. 2010, 120, 3466–3479. [Google Scholar] [CrossRef] [PubMed]
- Jiao, P.; Chen, Q.; Shah, S.; Du, J.; Tao, B.; Tzameli, I.; Yan, W.; Xu, H. Obesity-related upregulation of monocyte chemotactic factors in adipocytes: Involvement of nuclear factor-kappaB and c-Jun NH2-terminal kinase pathways. Diabetes 2009, 58, 104–115. [Google Scholar] [CrossRef] [PubMed]
- Fujisaka, S.; Usui, I.; Ikutani, M.; Aminuddin, A.; Takikawa, A.; Tsuneyama, K.; Mahmood, A.; Goda, N.; Nagai, Y.; Takatsu, K.; et al. Adipose tissue hypoxia induces inflammatory M1 polarity of macrophages in an HIF-1α-dependent and Hif-1α-independent manner in obese mice. Diabetologia 2013, 56, 1403–1412. [Google Scholar] [CrossRef] [PubMed]
- Cancello, R.; Henegar, C.; Viguerie, N.; Taleb, S.; Poitou, C.; Rouault, C.; Coupaye, M.; Pelloux, V.; Hugol, D.; Bouillot, J.L.; et al. Reduction of macrophage infiltration and chemoattractant gene expression changes in white adipose tissue of morbidly obese subjects after surgery-induced weight loss. Diabetes 2005, 54, 2277–2286. [Google Scholar] [CrossRef] [PubMed]
- Taylor, C.T.; Kent, B.D.; Crinion, S.J.; McNicholas, W.T.; Ryan, S. Human adipocytes are highly sensitive to intermittent hypoxia induced NF-kappaB activity and subsequent inflammatory gene expression. Biochem. Biophys. Res. Commun. 2014, 447, 660–665. [Google Scholar] [CrossRef] [PubMed]
- Gao, Z.; Hwang, D.; Bataille, F.; Lefevre, M.; York, D.; Quon, M.J.; Ye, J. Serine phosphorylation of insulin receptor substrate 1 by inhibitor kappa B kinase complex. J. Biol. Chem. 2002, 277, 48115–48121. [Google Scholar] [CrossRef] [PubMed]
- Hirosumi, J.; Tuncman, G.; Chang, L.; Gorgun, C.Z.; Uysal, K.T.; Maeda, K.; Karin, M.; Hotamisligil, G.S. A central role for jnk in obesity and insulin resistance. Nature 2002, 420, 333–336. [Google Scholar] [CrossRef] [PubMed]
- Arkan, M.C.; Hevener, A.L.; Greten, F.R.; Maeda, S.; Li, Z.W.; Long, J.M.; Wynshaw-Boris, A.; Poli, G.; Olefsky, J.; Karin, M. IKK-beta links inflammation to obesity-induced insulin resistance. Nat. Med. 2005, 11, 191–198. [Google Scholar] [CrossRef] [PubMed]
- Davis, J.E.; Gabler, N.K.; Walker-Daniels, J.; Spurlock, M.E. The c-Jun N-terminal kinase mediates the induction of oxidative stress and insulin resistance by palmitate and toll-like receptor 2 and 4 ligands in 3T3-L1 adipocytes. Horm. Metab.Res. 2009, 41, 523–530. [Google Scholar] [CrossRef] [PubMed]
- Lacasa, D.; Taleb, S.; Keophiphath, M.; Miranville, A.; Clement, K. Macrophage-secreted factors impair human adipogenesis: Involvement of proinflammatory state in preadipocytes. Endocrinology 2007, 148, 868–877. [Google Scholar] [CrossRef] [PubMed]
- Brown, M.S.; Goldstein, J.L. The srebp pathway: Regulation of cholesterol metabolism by proteolysis of a membrane-bound transcription factor. Cell 1997, 89, 331–340. [Google Scholar] [CrossRef] [PubMed]
- Mandrup, S.; Lane, M.D. Regulating adipogenesis. J. Biol. Chem. 1997, 272, 5367–5370. [Google Scholar] [CrossRef] [PubMed]
- Morrison, R.F.; Farmer, S.R. Hormonal signaling and transcriptional control of adipocyte differentiation. J. Nutr. 2000, 130, 3116S–3121S. [Google Scholar] [PubMed]
- Rosen, E.D.; Walkey, C.J.; Puigserver, P.; Spiegelman, B.M. Transcriptional regulation of adipogenesis. Genes Dev. 2000, 14, 1293–1307. [Google Scholar] [PubMed]
- Tamori, Y.; Masugi, J.; Nishino, N.; Kasuga, M. Role of peroxisome proliferator-activated receptor-gamma in maintenance of the characteristics of mature 3T3-l1 adipocytes. Diabetes 2002, 51, 2045–2055. [Google Scholar] [CrossRef] [PubMed]
- Fajas, L.; Fruchart, J.C.; Auwerx, J. Transcriptional control of adipogenesis. Curr. Opin. Cell Biol. 1998, 10, 165–173. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.; Rosen, E.D.; Brun, R.; Hauser, S.; Adelmant, G.; Troy, A.E.; McKeon, C.; Darlington, G.J.; Spiegelman, B.M. Cross-regulation of C/EBP alpha and PPAR gamma controls the transcriptional pathway of adipogenesis and insulin sensitivity. Mol. Cell 1999, 3, 151–158. [Google Scholar] [CrossRef] [PubMed]
- Spiegelman, B.M. PPAR-gamma: Adipogenic regulator and thiazolidinedione receptor. Diabetes 1998, 47, 507–514. [Google Scholar] [CrossRef] [PubMed]
- Fox, K.E.; Fankell, D.M.; Erickson, P.F.; Majka, S.M.; Crossno, J.T., Jr.; Klemm, D.J. Depletion of cAMP-response element-binding protein/ATF1 inhibits adipogenic conversion of 3T3-L1 cells ectopically expressing CCAAT/enhancer-binding protein (C/EBP) alpha, C/EBP beta, or PPAR gamma 2. J. Biol. Chem. 2006, 281, 40341–40353. [Google Scholar] [CrossRef] [PubMed]
- Siriwardhana, N.; Kalupahana, N.S.; Cekanova, M.; LeMieux, M.; Greer, B.; Moustaid-Moussa, N. Modulation of adipose tissue inflammation by bioactive food compounds. J. Nutr. Biochem. 2013, 24, 613–623. [Google Scholar] [CrossRef] [PubMed]
- Mosimann, A.L.; Wilhelm-Filho, D.; da Silva, E.L. Aqueous extract of Ilex paraguariensis attenuates the progression of atherosclerosis in cholesterol-fed rabbits. Biofactors 2006, 26, 59–70. [Google Scholar] [CrossRef] [PubMed]
- Bastos, D.H.M.; de Oliveira, D.M.; Matsumoto, R.L.T.; Carvalho, P.O.; Ribeiro, M.L. Yerba mate: Pharmacological properties, research and biotechnology. Med. Aromat. Plant Sci. Biotechnol. 2007, 1, 37–46. [Google Scholar]
- Gugliucci, A. Antioxidant effects of Ilex paraguariensis: Induction of decreased oxidability of human ldl in vivo. Biochem. Biophys. Res. Commun. 1996, 224, 338–344. [Google Scholar] [CrossRef] [PubMed]
- Gugliucci, A.; Menini, T. The botanical extracts of Achyrocline satureoides and Ilex paraguariensis prevent methylglyoxal-induced inhibition of plasminogen and antithrombin III. Life Sci. 2002, 72, 279–292. [Google Scholar] [CrossRef] [PubMed]
- Miranda, D.D.; Arcari, D.P.; Pedrazzoli, J., Jr.; Carvalho Pde, O.; Cerutti, S.M.; Bastos, D.H.; Ribeiro, M.L. Protective effects of mate tea (Ilex paraguariensis) on H2O2-induced DNA damage and DNA repair in mice. Mutagenesis 2008, 23, 261–265. [Google Scholar] [CrossRef] [PubMed]
- Boaventura, B.C.; di Pietro, P.F.; Stefanuto, A.; Klein, G.A.; de Morais, E.C.; de Andrade, F.; Wazlawik, E.; da Silva, E.L. Association of mate tea (Ilex paraguariensis) intake and dietary intervention and effects on oxidative stress biomarkers of dyslipidemic subjects. Nutrition 2012, 28, 657–664. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, E.S.; de Oliveira Machado, M.; Becker, A.M.; de Andrade, F.; Maraschin, M.; da Silva, E.L. Yerba mate (Ilex paraguariensis) enhances the gene modulation and activity of paraoxonase-2: In vitro and in vivo studies. Nutrition 2012, 28, 1157–1164. [Google Scholar] [CrossRef] [PubMed]
- Bixby, M.; Spieler, L.; Menini, T.; Gugliucci, A. Ilex paraguariensis extracts are potent inhibitors of nitrosative stress: A comparative study with green tea and wines using a protein nitration model and mammalian cell cytotoxicity. Life Sci. 2005, 77, 345–358. [Google Scholar] [CrossRef] [PubMed]
- Paganini Stein, F.L.; Schmidt, B.; Furlong, E.B.; Souza-Soares, L.A.; Soares, M.C.; Vaz, M.R.; Muccillo Baisch, A.L. Vascular responses to extractable fractions of Ilex paraguariensis in rats fed standard and high-cholesterol diets. Biol. Res. Nurs. 2005, 7, 146–156. [Google Scholar]
- Lunceford, N.; Gugliucci, A. Ilex paraguariensis extracts inhibit age formation more efficiently than green tea. Fitoterapia 2005, 76, 419–427. [Google Scholar] [CrossRef] [PubMed]
- Gugliucci, A.; Bastos, D.H.; Schulze, J.; Souza, M.F. Caffeic and chlorogenic acids in Ilex paraguariensis extracts are the main inhibitors of age generation by methylglyoxal in model proteins. Fitoterapia 2009, 80, 339–344. [Google Scholar] [CrossRef] [PubMed]
- Pereira, D.F.; Kappel, V.D.; Cazarolli, L.H.; Boligon, A.A.; Athayde, M.L.; Guesser, S.M.; da Silva, E.L.; Silva, F.R. Influence of the traditional Brazilian drink Ilex paraguariensis tea on glucose homeostasis. Phytomedicine 2012, 19, 868–877. [Google Scholar] [CrossRef] [PubMed]
- Hussein, G.M.; Matsuda, H.; Nakamura, S.; Akiyama, T.; Tamura, K.; Yoshikawa, M. Protective and ameliorative effects of mate (Ilex paraguariensis) on metabolic syndrome in tsod mice. Phytomedicine 2011, 19, 88–97. [Google Scholar] [CrossRef] [PubMed]
- Kang, Y.R.; Lee, H.Y.; Kim, J.H.; Moon, D.I.; Seo, M.Y.; Park, S.H.; Choi, K.H.; Kim, C.R.; Kim, S.H.; Oh, J.H.; et al. Anti-obesity and anti-diabetic effects of yerba mate (Ilex paraguariensis) in C57Bl/6J mice fed a high-fat diet. Lab. Anim. Res. 2012, 28, 23–29. [Google Scholar] [CrossRef] [PubMed]
- Arcari, D.P.; Bartchewsky, W., Jr.; dos Santos, T.W.; Oliveira, K.A.; DeOliveira, C.C.; Gotardo, E.M.; Pedrazzoli, J., Jr.; Gambero, A.; Ferraz, L.F.; Carvalho Pde, O.; et al. Anti-inflammatory effects of yerba mate extract (Ilex paraguariensis) ameliorate insulin resistance in mice with high fat diet-induced obesity. Mol. Cell. Endocrinol. 2011, 335, 110–115. [Google Scholar] [CrossRef] [PubMed]
- Pimentel, G.D.; Lira, F.S.; Rosa, J.C.; Caris, A.V.; Pinheiro, F.; Ribeiro, E.B.; Oyama, L.M.; Oller do Nascimento, C.M. Yerba mate extract (Ilex paraguariensis) attenuates both central and peripheral inflammatory effects of diet-induced obesity in rats. J. Nutr. Biochem. 2013, 24, 809–818. [Google Scholar] [CrossRef]
- Carmo, L.S.; Rogero, M.M.; Cortez, M.; Yamada, M.; Jacob, P.S.; Bastos, D.H.; Borelli, P.; Ambrosio Fock, R. The effects of yerba mate (Ilex paraguariensis) consumption on IL-1, IL-6, TNF-alpha and IL-10 production by bone marrow cells in wistar rats fed a high-fat diet. Int. J. Vitam. Nutr. Res. 2013, 83, 26–35. [Google Scholar] [CrossRef] [PubMed]
- Borges, M.C.; Vinolo, M.A.; Nakajima, K.; de Castro, I.A.; Bastos, D.H.; Borelli, P.; Fock, R.A.; Tirapegui, J.; Curi, R.; Rogero, M.M. The effect of mate tea (Ilex paraguariensis) on metabolic and inflammatory parameters in high-fat diet-fed wistar rats. Int. J. Food Sci. Nutr. 2013, 64, 561–569. [Google Scholar] [CrossRef] [PubMed]
- Arcari, D.P.; Bartchewsky, W.; dos Santos, T.W.; Oliveira, K.A.; Funck, A.; Pedrazzoli, J.; de Souza, M.F.F.; Saad, M.J.; Bastos, D.H.M.; Gambero, A.; et al. Antiobesity effects of yerba mate extract (Ilex paraguariensis) in high-fat diet-induced obese mice. Obesity 2009, 17, 2127–2133. [Google Scholar] [CrossRef] [PubMed]
- Filip, R.; Sebastian, T.; Ferraro, G.; Anesini, C. Effect of Ilex extracts and isolated compounds on peroxidase secretion of rat submandibulary glands. Food Chem. Toxicol. 2007, 45, 649–655. [Google Scholar] [CrossRef] [PubMed]
- Ramirez-Mares, M.V.; Chandra, S.; de Mejia, E.G. In vitro chemopreventive activity of Camellia sinensis, Ilex paraguariensis and Ardisia compressa tea extracts and selected polyphenols. Mutat. Res. 2004, 554, 53–65. [Google Scholar] [CrossRef] [PubMed]
- Puangpraphant, S.; Berhow, M.A.; Vermillion, K.; Potts, G.; Gonzalez de Mejia, E. Dicaffeoylquinic acids in yerba mate (Ilex paraguariensis st. Hilaire) inhibit NF-kappaB nucleus translocation in macrophages and induce apoptosis by activating caspases-8 and -3 in human colon cancer cells. Mol. Nutr. Food Res. 2011, 55, 1509–1522. [Google Scholar] [CrossRef] [PubMed]
- Puangpraphant, S.; Dia, V.P.; de Mejia, E.G.; Garcia, G.; Berhow, M.A.; Wallig, M.A. Yerba mate tea and mate saponins prevented azoxymethane-induced inflammation of rat colon through suppression of NF-kappaB p65ser(311) signaling via IkappaB-alpha and GSK-3beta reduced phosphorylation. Biofactors 2013, 39, 430–440. [Google Scholar] [CrossRef] [PubMed]
- Martinet, A.; Hostettmann, K.; Schutz, Y. Thermogenic effects of commercially available plant preparations aimed at treating human obesity. Phytomedicine 1999, 6, 231–238. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, D.M.; Freitas, H.S.; Souza, M.F.; Arcari, D.P.; Ribeiro, M.L.; Carvalho, P.O.; Bastos, D.H. Yerba mate (Ilex paraguariensis) aqueous extract decreases intestinal SGLT1 gene expression but does not affect other biochemical parameters in alloxan-diabetic wistar rats. J. Agric. Food Chem. 2008, 56, 10527–10532. [Google Scholar] [CrossRef] [PubMed]
- Lima Nda, S.; Franco, J.G.; Peixoto-Silva, N.; Maia, L.A.; Kaezer, A.; Felzenszwalb, I.; de Oliveira, E.; de Moura, E.G.; Lisboa, P.C. Ilex paraguariensis (yerba mate) improves endocrine and metabolic disorders in obese rats primed by early weaning. Eur. J. Nutr. 2014, 53, 73–82. [Google Scholar]
- Arcari, D.P.; Santos, J.C.; Gambero, A.; Ferraz, L.F.; Ribeiro, M.L. Modulatory effects of yerba mate (Ilex paraguariensis) on the PI3K-AKT signaling pathway. Mol. Nutr. Food Res. 2013, 57, 1882–1885. [Google Scholar] [PubMed]
- Pang, J.; Choi, Y.; Park, T. Ilex paraguariensis extract ameliorates obesity induced by high-fat diet: Potential role of ampk in the visceral adipose tissue. Arch. Biochem. Biophys. 2008, 476, 178–185. [Google Scholar] [CrossRef] [PubMed]
- Arcari, D.P.; Santos, J.C.; Gambero, A.; Ribeiro, M.L. The in vitro and in vivo effects of yerba mate (Ilex paraguariensis) extract on adipogenesis. Food Chem. 2013, 141, 809–815. [Google Scholar] [CrossRef] [PubMed]
- Gosmann, G.; Barlette, A.G.; Dhamer, T.; Arcari, D.P.; Santos, J.C.; de Camargo, E.R.; Acedo, S.; Gambero, A.; Gnoatto, S.C.; Ribeiro, M.L. Phenolic compounds from mate (Ilex paraguariensis) inhibit adipogenesis in 3T3-L1 preadipocytes. Plant Foods Hum. Nutr. 2012, 67, 156–161. [Google Scholar] [CrossRef] [PubMed]
- Hussein, G.M.; Matsuda, H.; Nakamura, S.; Hamao, M.; Akiyama, T.; Tamura, K.; Yoshikawa, M. Mate tea (Ilex paraguariensis) promotes satiety and body weight lowering in mice: Involvement of glucagon-like peptide-1. Biol. Pharm. Bull. 2011, 34, 1849–1855. [Google Scholar] [CrossRef] [PubMed]
- Resende, P.E.; Verza, S.G.; Kaiser, S.; Gomes, L.F.; Kucharski, L.C.; Ortega, G.G. The activity of mate saponins (Ilex paraguariensis) in intra-abdominal and epididymal fat, and glucose oxidation in male wistar rats. J. Ethnopharmacol. 2012, 144, 735–740. [Google Scholar] [CrossRef] [PubMed]
- Martins, F.; Noso, T.M.; Porto, V.B.; Curiel, A.; Gambero, A.; Bastos, D.H.; Ribeiro, M.L.; Carvalho Pde, O. Mate tea inhibits in vitro pancreatic lipase activity and has hypolipidemic effect on high-fat diet-induced obese mice. Obesity 2010, 18, 42–47. [Google Scholar] [CrossRef] [PubMed]
- Bracesco, N.; Sanchez, A.G.; Contreras, V.; Menini, T.; Gugliucci, A. Recent advances on Ilex paraguariensis research: Minireview. J. Ethnopharmacol. 2011, 136, 378–384. [Google Scholar] [CrossRef] [PubMed]
- Gugliucci, A.; Stahl, A.J. Low density lipoprotein oxidation is inhibited by extracts of Ilex paraguariensis. Biochem. Mol. Biol. Int. 1995, 35, 47–56. [Google Scholar] [PubMed]
- Andersen, T.; Fogh, J. Weight loss and delayed gastric emptying following a south American herbal preparation in overweight patients. J. Hum. Nutr. Diet. 2001, 14, 243–250. [Google Scholar] [CrossRef] [PubMed]
- Harrold, J.A.; Hughes, G.M.; OʼShiel, K.; Quinn, E.; Boyland, E.J.; Williams, N.J.; Halford, J.C. Acute effects of a herb extract formulation and inulin fibre on appetite, energy intake and food choice. Appetite 2013, 62, 84–90. [Google Scholar] [CrossRef] [PubMed]
- De Morais, E.C.; Stefanuto, A.; Klein, G.A.; Boaventura, B.C.; de Andrade, F.; Wazlawik, E.; di Pietro, P.F.; Maraschin, M.; da Silva, E.L. Consumption of yerba mate (Ilex paraguariensis) improves serum lipid parameters in healthy dyslipidemic subjects and provides an additional LDL-cholesterol reduction in individuals on statin therapy. J. Agric. Food Chem. 2009, 57, 8316–8324. [Google Scholar]
- Gao, H.; Liu, Z.; Qu, X.; Zhao, Y. Effects of yerba mate tea (Ilex paraguariensis) on vascular endothelial function and liver lipoprotein receptor gene expression in hyperlipidemic rats. Fitoterapia 2013, 84, 264–272. [Google Scholar] [CrossRef] [PubMed]
- Hemmerle, H.; Burger, H.J.; Below, P.; Schubert, G.; Rippel, R.; Schindler, P.W.; Paulus, E.; Herling, A.W. Chlorogenic acid and synthetic chlorogenic acid derivatives: Novel inhibitors of hepatic glucose-6-phosphate translocase. J. Med. Chem. 1997, 40, 137–145. [Google Scholar] [CrossRef] [PubMed]
- Nardini, M.; DʼAquino, M.; Tomassi, G.; Gentili, V.; di Felice, M.; Scaccini, C. Inhibition of human low-density lipoprotein oxidation by caffeic acid and other hydroxycinnamic acid derivatives. Free Radic Biol. Med. 1995, 19, 541–552. [Google Scholar] [CrossRef] [PubMed]
- Han, L.K.; Zheng, Y.N.; Xu, B.J.; Okuda, H.; Kimura, Y. Saponins from platycodi radix ameliorate high fat diet-induced obesity in mice. J. Nutr. 2002, 132, 2241–2245. [Google Scholar] [PubMed]
- Tseng, Y.H.; Butte, A.J.; Kokkotou, E.; Yechoor, V.K.; Taniguchi, C.M.; Kriauciunas, K.M.; Cypess, A.M.; Niinobe, M.; Yoshikawa, K.; Patti, M.E.; et al. Prediction of preadipocyte differentiation by gene expression reveals role of insulin receptor substrates and necdin. Nat. Cell Biol. 2005, 7, 601–611. [Google Scholar] [CrossRef] [PubMed]
- Tong, Q.; Tsai, J.; Tan, G.; Dalgin, G.; Hotamisligil, G.S. Interaction between gata and the C/EBP family of transcription factors is critical in gata-mediated suppression of adipocyte differentiation. Mol. Cell Biol. 2005, 25, 706–715. [Google Scholar] [CrossRef] [PubMed]
- Banerjee, S.S.; Feinberg, M.W.; Watanabe, M.; Gray, S.; Haspel, R.L.; Denkinger, D.J.; Kawahara, R.; Hauner, H.; Jain, M.K. The kruppel-like factor KLF2 inhibits peroxisome proliferator-activated receptor-gamma expression and adipogenesis. J. Biol. Chem. 2003, 278, 2581–2584. [Google Scholar] [CrossRef] [PubMed]
- Hotamisligil, G.S. Inflammation and metabolic disorders. Nature 2006, 444, 860–867. [Google Scholar] [CrossRef] [PubMed]
- Shimomura, I.; Hammer, R.E.; Ikemoto, S.; Brown, M.S.; Goldstein, J.L. Leptin reverses insulin resistance and diabetes mellitus in mice with congenital lipodystrophy. Nature 1999, 401, 73–76. [Google Scholar] [CrossRef] [PubMed]
- Farese, R.V.; Sajan, M.P.; Standaert, M.L. Insulin-sensitive protein kinases (atypical protein kinase C and protein kinase B/AKT): Actions and defects in obesity and type II diabetes. Exp. Biol. Med. (Maywood) 2005, 230, 593–605. [Google Scholar]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gambero, A.; Ribeiro, M.L. The Positive Effects of Yerba Maté (Ilex paraguariensis) in Obesity. Nutrients 2015, 7, 730-750. https://doi.org/10.3390/nu7020730
Gambero A, Ribeiro ML. The Positive Effects of Yerba Maté (Ilex paraguariensis) in Obesity. Nutrients. 2015; 7(2):730-750. https://doi.org/10.3390/nu7020730
Chicago/Turabian StyleGambero, Alessandra, and Marcelo L. Ribeiro. 2015. "The Positive Effects of Yerba Maté (Ilex paraguariensis) in Obesity" Nutrients 7, no. 2: 730-750. https://doi.org/10.3390/nu7020730
APA StyleGambero, A., & Ribeiro, M. L. (2015). The Positive Effects of Yerba Maté (Ilex paraguariensis) in Obesity. Nutrients, 7(2), 730-750. https://doi.org/10.3390/nu7020730