An Ethyl Acetate Fraction of Moringa oleifera Lam. Inhibits Human Macrophage Cytokine Production Induced by Cigarette Smoke

Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. Extractions and Fractionation of MO Leaves
2.3. Determination of Total Phenolic Content
2.4. Determination of Free Radical Scavenging Activity
2.5. Isolation and Differentiation of Human Peripheral Blood Monocytes
2.6. Preparation of Cigarette Smoke Extract
2.7. Determination of Cellular Cytotoxicity
2.8. Induction of an Acute Inflammatory Response
2.9. Investigation of Anti-Inflammatory Activity
2.10. Enzyme-Linked Immunosorbent Assay (ELISA)
2.11. Total RNA Extraction
2.12. Gene Expression Analysis
2.13. Experimental Design and Statistical Analysis

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Oligonucleotide Sequence (5′→3′) | Tm (°C) | Amplicon Size (bp) | |
|---|---|---|---|---|
| TNF | Homo sapiens tumor necrosis factor, mRNA | |||
| F: | CCTCTCTCTAATCAGCCCTCTG | 60.8 | 220 | |
| R: | GAGGACCTGGGAGTAGATGAG | 60.2 | ||
| IL6 | Homo sapiens interleukin 6 (interferon, β2), mRNA | |||
| F: | ACTCACCTCTTCAGAACGAATTG | 60.2 | 149 | |
| R: | CCATCTTTGGAAGGTTCAGGTTG | 61.3 | ||
| IL8 | Homo sapiens interleukin 8 precursor, mRNA | |||
| F: | TTTTGCCAAGGAGTGCTAAAGA | 60.1 | 194 | |
| R: | AACCCTCTGCACCCAGTTTTC | 62.5 | ||
| RelA * | Homo sapiens v-rel reticuloendotheliosis viral oncogene homolog A (avian), transcript variant 2, mRNA | |||
| F: | ATGTGGAGATCATTGAGCAGC | 60.2 | 151 | |
| R: | CCTGGTCCTGTGTAGCCATT | 61.3 | ||
| ACTB | β actin; β cytoskeletal actin [Homo sapiens] | |||
| F: | CATGTACGTTGCTATCCAGGC | 60.8 | 250 | |
| R: | CTCCTTAATGTCACGCACGAT | 60.2 | ||
3. Results
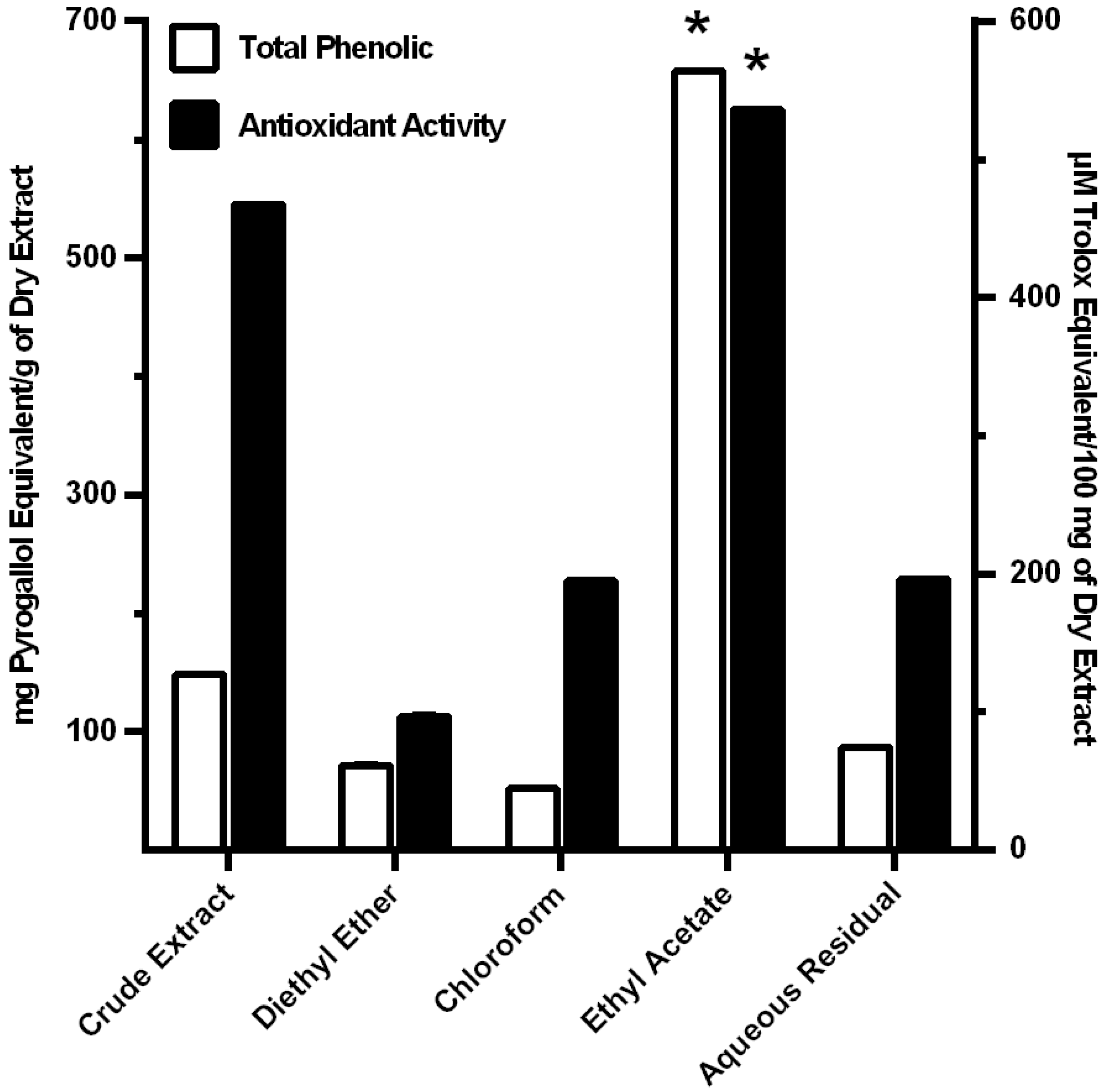
3.1. Extraction and Fractionation of MO Leaves
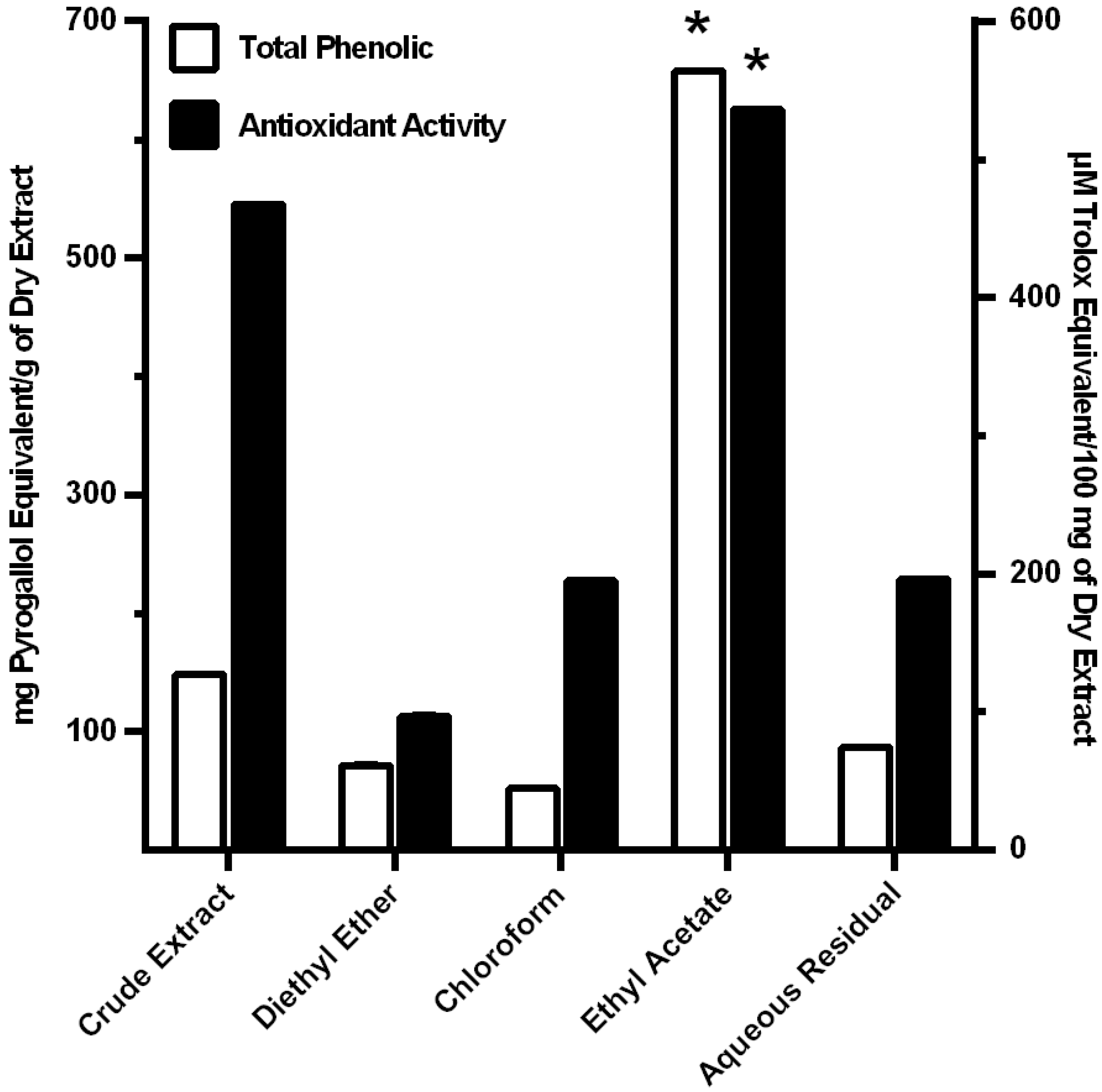
3.2. Antioxidant Activity of MO Extracts
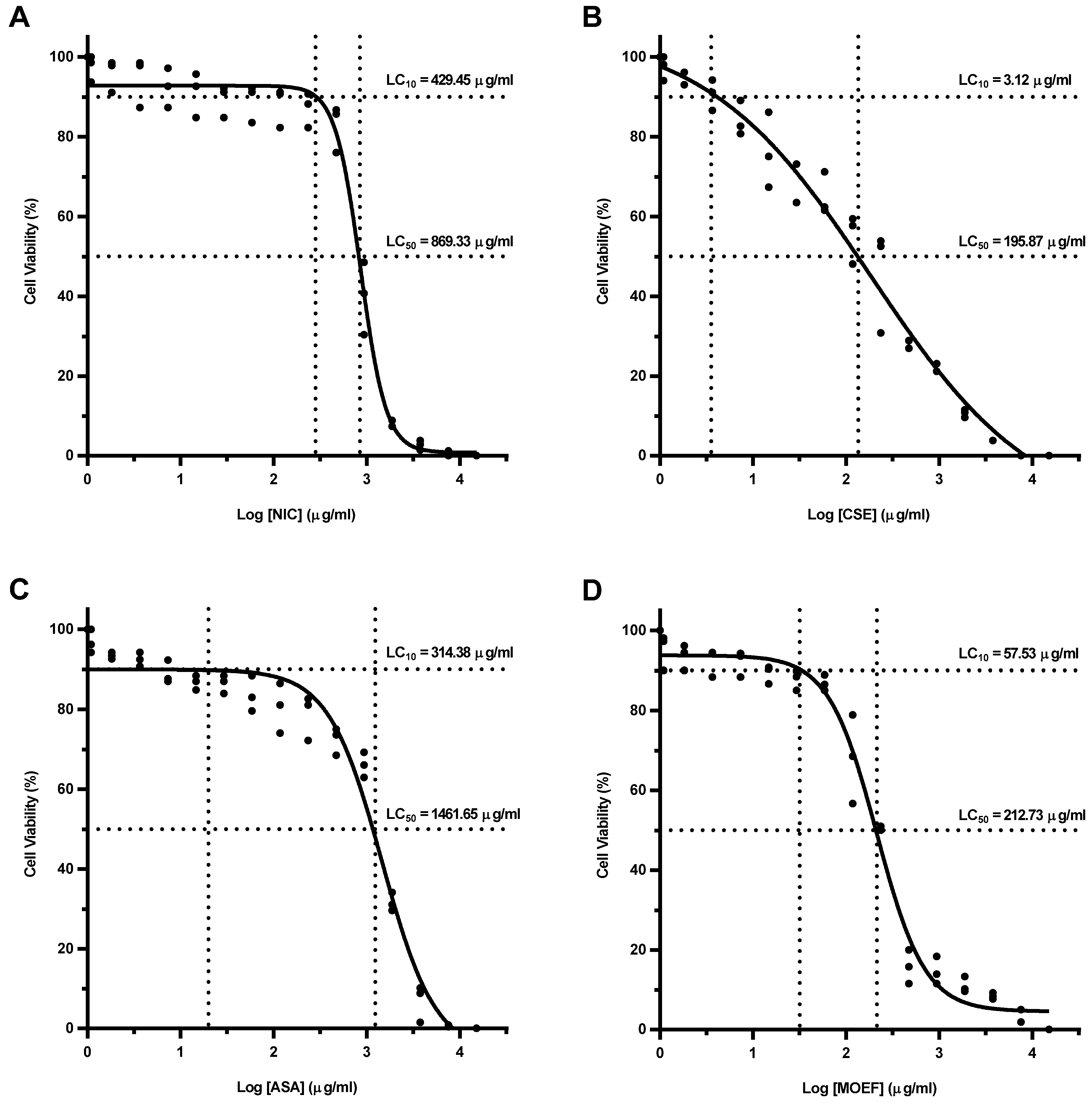
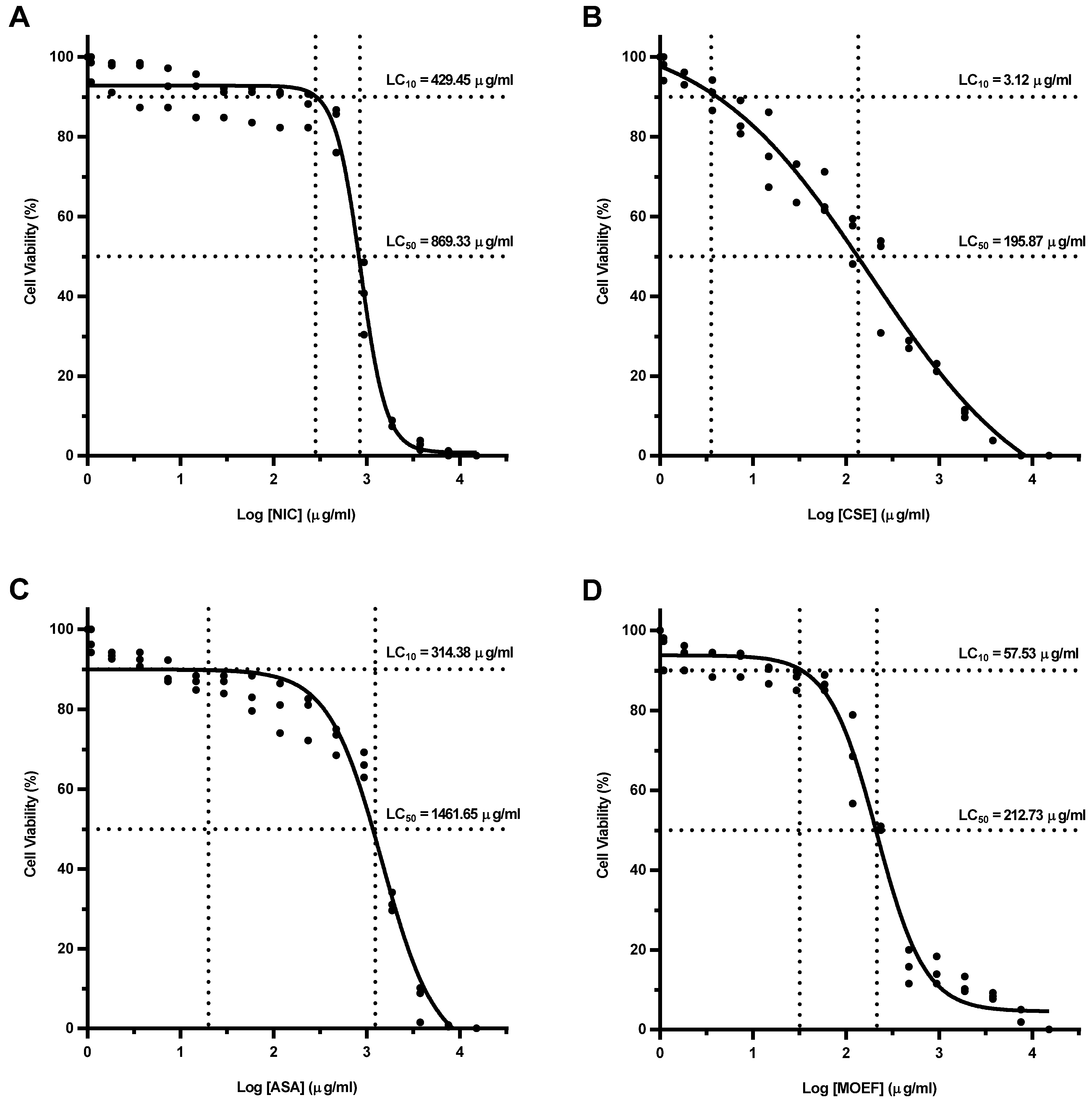
3.3. Cellular Cytotoxicity of Fractions
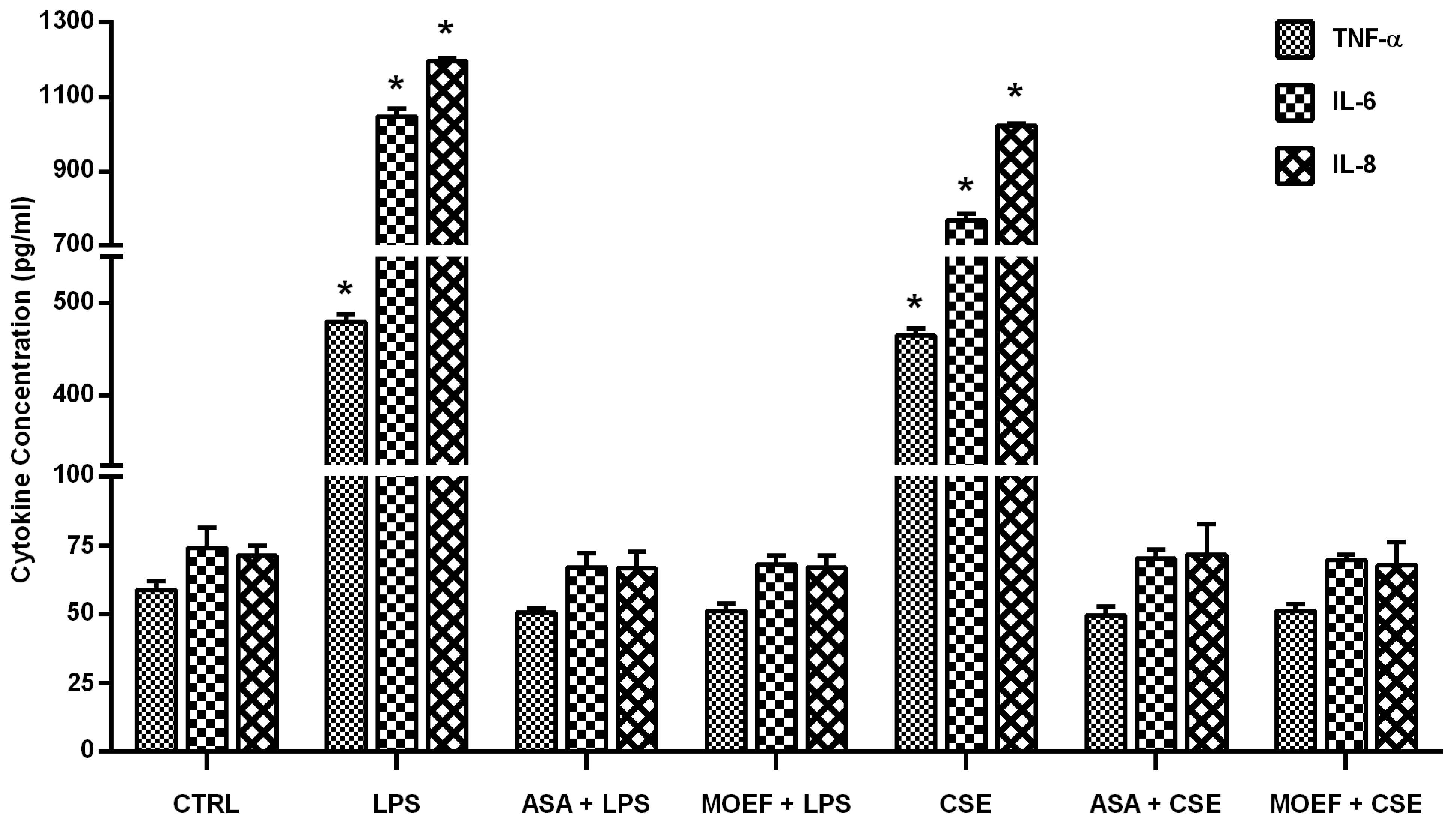
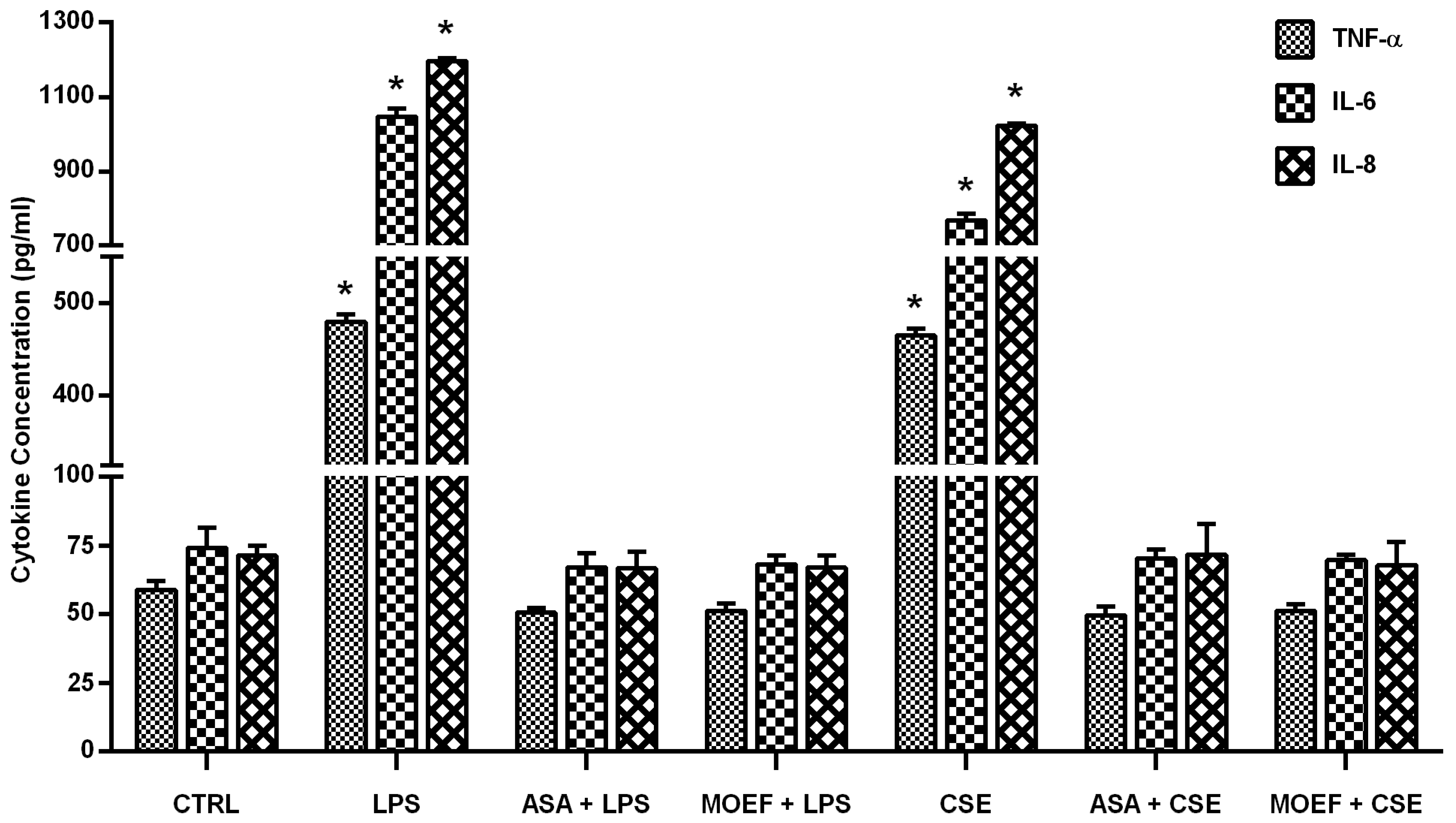
3.4. Effect on Cytokines Production
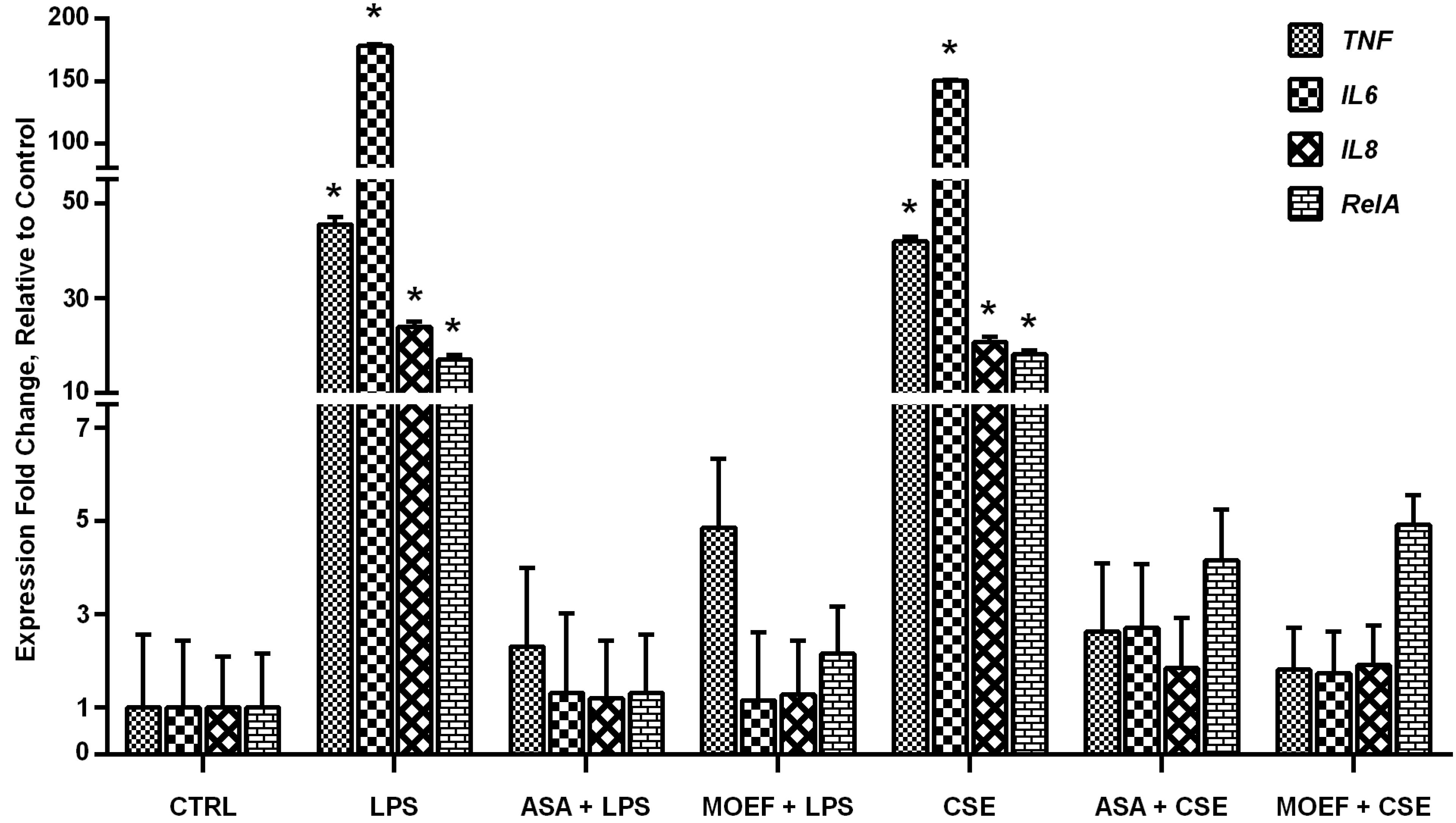
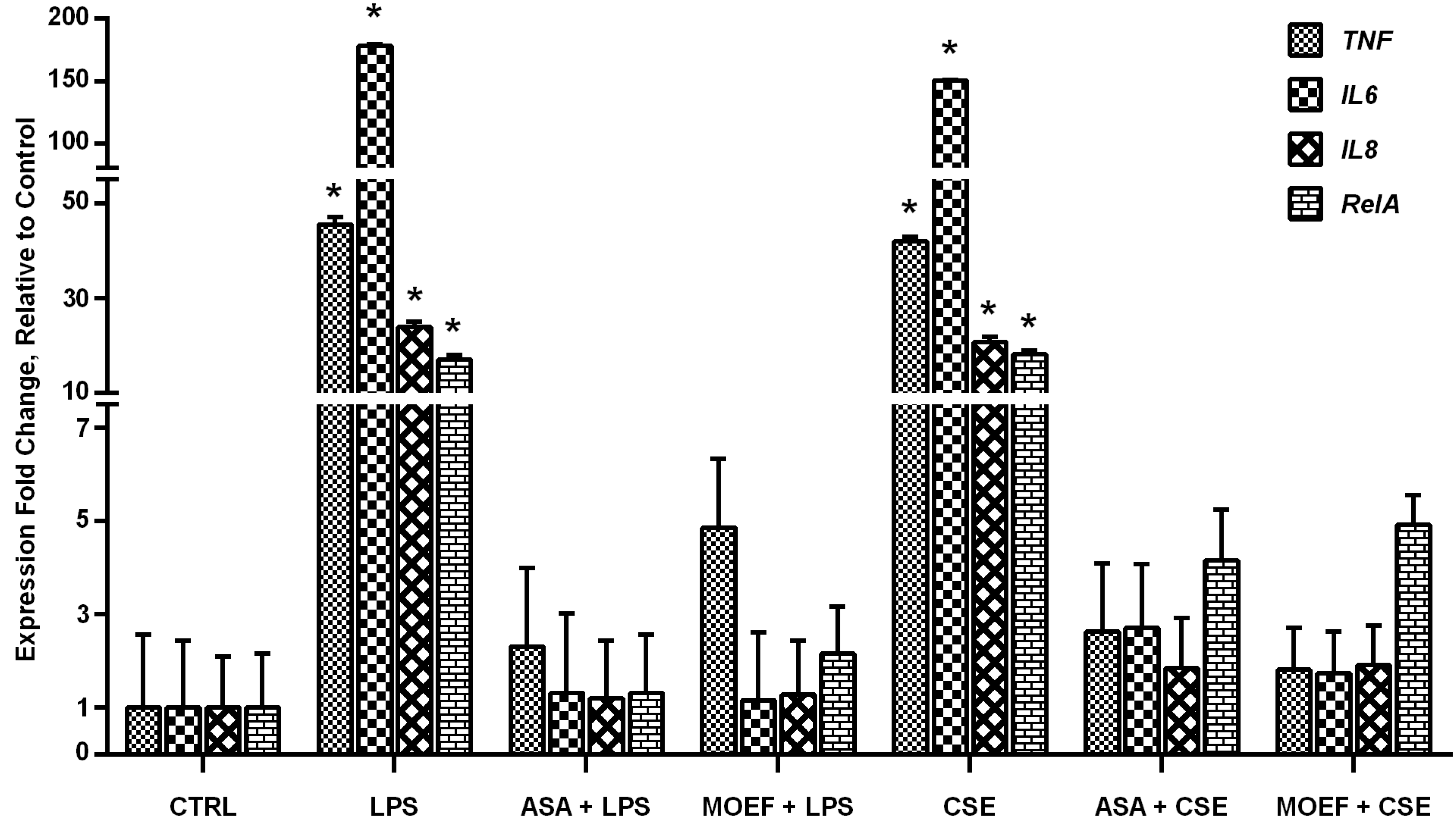
3.5. Modulation of Inflammatory Gene and Transcription Factor Gene Expression
4. Discussion
5. Conclusions
Acknowledgments
Conflicts of Interest
References
- Soto-Vaca, A.; Gutierrez, A.; Losso, J.N.; Xu, Z.; Finley, J.W. Evolution of phenolic compounds from color and flavor problems to health benefits. J. Agric. Food Chem. 2012, 60, 6658–6677. [Google Scholar] [CrossRef]
- Chang, C.L.; Lin, C.S.; Lai, G.H. Phytochemical characteristics, free radical scavenging activities, and neuroprotection of five medicinal plant extracts. Evid. Based Complement. Alternat. Med. 2012, 2012, 984295. [Google Scholar]
- Sreelatha, S.; Padma, P.R. Modulatory effects of Moringa oleifera extracts against hydrogen peroxide-induced cytotoxicity and oxidative damage. Hum. Exp. Toxicol. 2011, 30, 1359–1368. [Google Scholar] [CrossRef]
- Mensah, J.K.; Ikhajiagbe, B.; Edema, N.E.; Emokhor, J. Phytochemical, nutritional and antibacterial properties of dried leaf powder of Moringa oleifera (Lam.) from Edo Central Province, Nigeria. J. Nat. Prod. Plant Resour. 2012, 2, 107–112. [Google Scholar]
- Bamishaiye, E.I.; Olayemi, F.F.; Awagu, E.F.; Bamshaiye, O.M. Proximate and phytochemical composition of Moringa oleifera leaves at three stages of maturation. Adv. J. Food. Sci. Technol. 2011, 3, 233–237. [Google Scholar]
- Farooq, F.; Rai, M.; Tiwari, A.; Khan, A.A.; Farooq, S. Medicinal properties of Moringa oleifera: An overview of promising healer. J. Med. Plants Res. 2012, 6, 4368–4374. [Google Scholar]
- Posmontier, B. The medicinal qualities of Moringa oleifera. Holist. Nurs. Pract. 2011, 25, 80–87. [Google Scholar] [CrossRef]
- Holst, S. Moringa: Nature’s Medicine Cabinet; Santorini Publishing: Secaucus, NJ, USA, 2011. [Google Scholar]
- Marcilla, A.; Martinez, I.; Berenguer, D.; Gomez-Siurana, A.; Beltran, M.I. Comparative study of the main characteristics and composition of the mainstream smoke of ten cigarette brands sold in Spain. Food Chem. Toxicol. 2012, 50, 1317–1333. [Google Scholar] [CrossRef]
- Purkis, S.W.; Mueller, C.; Intorp, M. The fate of ingredients in and impact on cigarette smoke. Food Chem. Toxicol. 2011, 49, 3238–3248. [Google Scholar] [CrossRef]
- Taito, S.; Sekikawa, K.; Domen, S.; Konishi, K.; Kimura, T.; Takahashi, M.; Inamizu, T.; Hamada, H. Pulmonary oxidative stress is induced by maximal exercise in young cigarette smokers. Nicotine Tob. Res. 2012, 14, 243–247. [Google Scholar] [CrossRef]
- Colombo, G.; Dalle-Donne, I.; Orioli, M.; Giustarini, D.; Rossi, R.; Clerici, M.; Regazzoni, L.; Aldini, G.; Milzani, A.; Butterfield, D.A.; et al. Oxidative damage in human gingival fibroblasts exposed to cigarette smoke. Free Radic. Biol. Med. 2012, 52, 1584–1596. [Google Scholar] [CrossRef]
- Leonarduzzi, G.; Sottero, B.; Poli, G. Targeting tissue oxidative damage by means of cell signaling modulators: The antioxidant concept revisited. Pharmacol. Ther. 2010, 128, 336–374. [Google Scholar] [CrossRef]
- Barreiro, E.; Peinado, V.I.; Galdiz, J.B.; Ferrer, E.; Marin-Corral, J.; Sanchez, F.; Gea, J.; Barbera, J.A.; Project, E.I.C. Cigarette smoke-induced oxidative stress: A role in chronic obstructive pulmonary disease skeletal muscle dysfunction. Am. J. Respir. Crit. Care Med. 2010, 182, 477–488. [Google Scholar] [CrossRef]
- Rastrick, J.M.; Stevenson, C.S.; Eltom, S.; Grace, M.; Davies, M.; Kilty, I.; Evans, S.M.; Pasparakis, M.; Catley, M.C.; Lawrence, T.; et al. Cigarette smoke induced airway inflammation is independent of NF-κB signalling. PLoS One 2013, 8, e54128. [Google Scholar] [CrossRef]
- Oeckinghaus, A.; Hayden, M.S.; Ghosh, S. Crosstalk in NF-κB signaling pathways. Nat. Immunol. 2011, 12, 695–708. [Google Scholar] [CrossRef]
- Wright, W.R.; Parzych, K.; Crawford, D.; Mein, C.; Mitchell, J.A.; Paul-Clark, M.J. Inflammatory transcriptome profiling of human monocytes exposed acutely to cigarette smoke. PLoS One 2012, 7, e30120. [Google Scholar]
- Pera, T.; Atmaj, C.; van der Vegt, M.; Halayko, A.J.; Zaagsma, J.; Meurs, H. Role for TAK1 in cigarette smoke-induced proinflammatory signaling and IL-8 release by human airway smooth muscle cells. Am. J. Physiol. Lung Cell. Mol. Physiol. 2012, 303, L272–L278. [Google Scholar] [CrossRef]
- Tamimi, A.; Serdarevic, D.; Hanania, N.A. The effects of cigarette smoke on airway inflammation in asthma and COPD: Therapeutic implications. Respir. Med. 2012, 106, 319–328. [Google Scholar] [CrossRef]
- Kundu, J.K.; Surh, Y.J. Emerging avenues linking inflammation and cancer. Free Radic. Biol. Med. 2012, 52, 2013–2037. [Google Scholar] [CrossRef]
- Huang, R.Y.; Chen, G.G. Cigarette smoking, cyclooxygenase-2 pathway and cancer. Biochim. Biophys. Acta 2011, 1815, 158–169. [Google Scholar]
- Verma, A.R.; Vijayakumar, M.; Mathela, C.S.; Rao, C.V. In vitro and in vivo antioxidant properties of different fractions of Moringa oleifera leaves. Food Chem. Toxicol. 2009, 47, 2196–2201. [Google Scholar] [CrossRef]
- Floegel, A.; Kim, D.-O.; Chung, S.-J.; Koo, S.I.; Chun, O.K. Comparison of ABTS/DPPH assays to measure antioxidant capacity in popular antioxidant-rich US foods. J. Food Compost. Anal. 2011, 24, 1043–1048. [Google Scholar] [CrossRef]
- Fuss, I.J.; Kanof, M.E.; Smith, P.D.; Zola, H. Isolation of Whole Mononuclear Cells from Peripheral Blood and Cord Blood. In Current Protocols in Immunology; Coligan, J.E., Bierer, B.E., Margulies, D.H., Shevach, E.M., Strober, W., Brown, P., Donovan, J.C., Kruisbeek, A.M., Eds.; John Wiley & Sons: Hoboken, NJ, USA, 2001. [Google Scholar]
- Wahl, L.M.; Wahl, S.M.; Smythies, L.E.; Smith, P.D. Isolation of Human Monocyte Populations. In Current Protocols in Immunology; Coligan, J.E., Bierer, B.E., Margulies, D.H., Shevach, E.M., Strober, W., Brown, P., Donovan, J.C., Kruisbeek, A.M., Eds.; John Wiley & Sons: Hoboken, NJ, USA, 2001. [Google Scholar]
- Cederblad, L.; Thunberg, U.; Engstrom, M.; Castro, J.; Rutqvist, L.E.; Laytragoon-Lewin, N. The combined effects of single-nucleotide polymorphisms, tobacco products, and ethanol on normal resting blood mononuclear cells. Nicotine Tob. Res. 2013, 15, 890–895. [Google Scholar] [CrossRef]
- Repetto, G.; del Peso, A.; Zurita, J.L. Neutral red uptake assay for the estimation of cell viability/cytotoxicity. Nat. Protoc. 2008, 3, 1125–1131. [Google Scholar] [CrossRef]
- Russell, M.A.; Jarvis, M.; Iyer, R.; Feyerabend, C. Relation of nicotine yield of cigarettes to blood nicotine concentrations in smokers. Br. Med. J. 1980, 280, 972–976. [Google Scholar] [CrossRef]
- Cohen, A.; George, O. Animal models of nicotine exposure: Relevance to second-hand smoking, electronic cigarette use, and compulsive smoking. Front. Psychiatry 2013, 4. [Google Scholar] [CrossRef]
- Lawrence, T. The nuclear factor NF-κB pathway in inflammation. Cold Spring Harb. Perspect. Biol. 2009, 1, a001651. [Google Scholar] [CrossRef]
- Spandidos, A.; Wang, X.; Wang, H.; Seed, B. PrimerBank: A resource of human and mouse PCR primer pairs for gene expression detection and quantification. Nucleic Acids Res. 2010, 38, D792–D799. [Google Scholar] [CrossRef]
- Van Peer, G.; Mestdagh, P.; Vandesompele, J. Accurate RT-qPCR gene expression analysis on cell culture lysates. Sci. Rep. 2012, 2. [Google Scholar] [CrossRef] [Green Version]
- Sarir, H.; Mortaz, E.; Karimi, K.; Kraneveld, A.D.; Rahman, I.; Caldenhoven, E.; Nijkamp, F.P.; Folkerts, G. Cigarette smoke regulates the expression of TLR4 and IL-8 production by human macrophages. J. Inflamm. 2009, 6. [Google Scholar] [CrossRef]
- Lerner, L.; Weiner, D.; Katz, R.; Reznick, A.Z.; Pollack, S. Increased pro-inflammatory activity and impairment of human monocyte differentiation induced by in vitro exposure to cigarette smoke. J. Physiol. Pharmacol. 2009, 60, 81–86. [Google Scholar]
- Guzman, J.R.; Koo, J.S.; Goldsmith, J.R.; Muhlbauer, M.; Narula, A.; Jobin, C. Oxymatrine prevents NF-κB nuclear translocation and ameliorates acute intestinal inflammation. Sci. Rep. 2013, 3. [Google Scholar] [CrossRef]
- Hwang, J.W.; Oh, J.H.; Yoo, H.S.; Lee, Y.W.; Cho, C.K.; Kwon, K.R.; Yoon, J.H.; Park, J.; Her, S.; Lee, Z.W.; et al. Mountain ginseng extract exhibits anti-lung cancer activity by inhibiting the nuclear translocation of NF-κB. Am. J. Chin. Med. 2012, 40, 187–202. [Google Scholar] [CrossRef]
- Anchoori, R.K.; Harikumar, K.B.; Batchu, V.R.; Aggarwal, B.B.; Khan, S.R. Inhibition of IkB kinase and NF-κB by a novel synthetic compound SK 2009. Bioorg. Med. Chem. 2010, 18, 229–235. [Google Scholar]
- Oeckinghaus, A.; Ghosh, S. The NF-κB family of transcription factors and its regulation. Cold Spring Harb. Perspect. Biol. 2009, 1, a000034. [Google Scholar]
- Muangnoi, C.; Chingsuwanrote, P.; Praengamthanachoti, P.; Svasti, S.; Tuntipopipat, S. Moringa oleifera pod inhibits inflammatory mediator production by lipopolysaccharide-stimulated RAW 264.7 murine macrophage cell lines. Inflammation 2012, 35, 445–455. [Google Scholar] [CrossRef]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Kooltheat, N.; Sranujit, R.P.; Chumark, P.; Potup, P.; Laytragoon-Lewin, N.; Usuwanthim, K. An Ethyl Acetate Fraction of Moringa oleifera Lam. Inhibits Human Macrophage Cytokine Production Induced by Cigarette Smoke. Nutrients 2014, 6, 697-710. https://doi.org/10.3390/nu6020697
Kooltheat N, Sranujit RP, Chumark P, Potup P, Laytragoon-Lewin N, Usuwanthim K. An Ethyl Acetate Fraction of Moringa oleifera Lam. Inhibits Human Macrophage Cytokine Production Induced by Cigarette Smoke. Nutrients. 2014; 6(2):697-710. https://doi.org/10.3390/nu6020697
Chicago/Turabian StyleKooltheat, Nateelak, Rungnapa Pankla Sranujit, Pilaipark Chumark, Pachuen Potup, Nongnit Laytragoon-Lewin, and Kanchana Usuwanthim. 2014. "An Ethyl Acetate Fraction of Moringa oleifera Lam. Inhibits Human Macrophage Cytokine Production Induced by Cigarette Smoke" Nutrients 6, no. 2: 697-710. https://doi.org/10.3390/nu6020697